

Abstract
Accumulating evidences suggest that oxidative stress caused and deteriorated the aging related osteoporosis and pyrroloquinoline quinone (PQQ) is a powerful antioxidant. However, it is unclear whether PQQ can prevent testosterone deficiency-induced osteoporosis. In this study, the orchidectomized (ORX) mice were supplemented in diet with/without PQQ for 48 weeks, and compared with each other and with sham mice. Results showed that bone mineral density, trabecular bone volume, collagen deposition and osteoblast number were decreased significantly in ORX mice compared with shame mice, whereas PQQ supplementation largely prevented these alterations. In contrast, osteoclast surface and ratio of RANKL and OPG mRNA relative expression levels were increased significantly in ORX mice compared with shame mice, but were decreased significantly by PQQ supplementation. Furthermore, we found that CFU-f and ALP positive CFU-f forming efficiency and the proliferation of mesenchymal stem cells were reduced significantly in ORX mice compared with shame mice, but were increased significantly by PQQ supplementation. Reactive oxygen species (ROS) levels in thymus were increased, antioxidant enzymes SOD-1, SOD-2, Prdx I and Prdx IV protein expression levels in bony tissue were down-regulated, whereas the protein expression levels of DNA damage response related molecules including γ-H2AX, p53, Chk2 and NFκB-p65 in bony tissue were up-regulated significantly in ORX mice compared with shame mice, whereas PQQ supplementation largely rescued these alterations observed in ORX mice. Our results indicate that PQQ supplementation can prevent testosterone deficiency-induced osteoporosis by inhibiting oxidative stress and DNA damage, stimulating osteoblastic bone formation and inhibiting osteoclastic bone resorption.
Keywords: Male osteoporosis, orchiectomy, pyrroloquinoline quinone, oxidative stress
Introduction
Osteoporosis is a disease that thins and weakens the bones to the point that they become fragile and break easily. It is well-known that osteoporosis-related fractures (low-trauma or fragility fractures) cause substantial disability, health-related costs and mortality among postmenopausal women and older men [1].
Osteoporosis is now recognized as a major threat to health in elderly [2], despite not undergoing a menopausal transition as women do, men sustain bone loss of approximately 0.5-1% per year beginning at the sixth decade [3]. Although the role of declining sex steroid levels in bone loss is well recognized in women, remarkable changes in sex steroid levels also occur over the lifespan in men. With aging, the change of sex steroid hormone levels seems to be a leading cause of osteoporosis in men. The testosterone deficiency-induced osteoporosis in men occurs later than postmenopausal osteoporosis in women, but the morbidity and mortality after osteoporotic fractures are greater [4]. With aging, bone formation decreases due to reduction in osteoblast number, activity, and life span, whereas bone resorption increases as a result of sex hormone deprivation. These two mechanisms contribute to the decreased bone mass and increased risk of fractures seen in the aging population. Current effective antiresorbing drugs reduce bone remodeling in osteoporotic subjects. An ideal way to prevent age-related bone loss would be not only to reduce bone resorption, but also to promote bone formation. There is therefore an important need to develop therapeutic strategies capable of promoting bone formation in osteoporotic subjects.
Reactive oxygen species (ROS) are a number of reactive molecules and free radicals generated in the process of endogenous mitochondrial oxidative phosphorylation or from the reaction with exogenous compounds in environment. Previous studies showed that oxidative stress or ROS caused aging, atherosclerosis, tumorigenesis, arthritis and osteoporosis [5-11]. In bone, low level of ROS was reported to promote the differentiation of hypertrophic chondrocytes into osteoblasts to form bones [12], while high levels of ROS influenced the bone formation and remodeling by inhibiting the proliferation and differentiation of mesenchymal stem cells (MSCs) into osteoblasts and the activity of osteoblasts as well [13-17]. Interestingly, aged human MSCs had elevated levels of ROS and inhibition of ROS level rescued the ability of these MSCs to proliferate normally. Therefore, it is no doubt that there is a critical role of ROS in modulating the cell proliferation and survival.
Recently, accumulating evidences based on the animal studies and epidemiological researches inferred that the high oxidative stress caused and deteriorated the aging related osteoporosis [18]. Although literatures showed that testosterone deficiency resulted in increased oxidative stress which accelerated skeleton aging, however, the mechanism of the testosterone deficiency-induced osteoporosis is not clear [19].
Pyrroloquinoline quinone (PQQ) is a powerful antioxidant [20]. Many studies [21-24] suggest that antioxidants act as inhibitors of osteoclastogenesis and promote osteoblastogenesis. Whether PQQ could improve the testosterone deficiency-induced osteoporosis has not been investigated and related mechanism is unknown. To determine the effect of PQQ on the testosterone deficiency-induced osteoporosis, we established a testosterone deficiency mouse model by orchiectomy (ORX) and ORX mice were supplemented with/without PQQ in the diet. The control mice received shame surgery on a normal diet. We examined the alterations of bone formation and resorption in these mice by morphological, histopathological, and molecular biological methods to investigate whether PQQ can prevent testosterone deficiency-induced osteoporosis by inhibiting oxidative stress.
Materials and methods
Animals
2-month-old C57BL/6 background wild type mice used in this study were bred and maintained in specific pathogen free (SPF) laboratory animal center in Nanjing Medical University. All animals were housed in the room with the appropriate temperature (22°C~26°C) and humidity (45%~75%) range. Animals were randomly divided into three groups: 1) sham surgery with normal diet; 2) ORX group with normal diet; 3) ORX group supplemented with PQQ in the diet. ORX mice were received orchiectomy surgery at both sides [25] and given PQQ supplement immediately after the surgery or a control normal diet. PQQ diet was obtained by supplementing 4 mg PQQ per Kg diet. All mice were sacrificed at 48 weeks after the surgery. All animal procedures were approved by the Institutional Animal Care and Use Committee.
X-ray, micro-CT scanning and 3D reconstruction
Vertebrae from three groups were fixed in 4% paraformaldehyde and applied for X-ray radiograph and micro-CT scanning. Radiographs were taken as described previously [26]. Same samples were then scanned on a micro-CT scanner (Sky Scan 1072 Scanner) using 100 kV energy, and 98 µA intensity. 3D images were generated using the 3D Creator software supplied with the instrument as described previously [26].
Histology
All animals were sacrificed by cervical dislocation after inhaled anesthesia with ether. Vertebrae were removed and dissected free of soft tissue, fixed with 4% paraformaldehyde, decalcifed with EDTA glycerol solution and embedded in paraffin. Sections were stained with H&E, or histochemically for total collagen or tartrate-resistant acid phosphatase (TRAP). For total collagen staining, sections were stained with saturated 1% Sirius Red in saturated picric acid solution for 1 hour at room temperature after dewaxing and hydration, counterstained with hematoxylin and mounted with permount. For TRAP staining, sections were stained with TRAP staining buffer (25 mg Naphthol AS-MX phosphate and 5 mg Fast Garnet GBC dissolved in 50 mM sodium acetate, 40 mM potassium sodium tartrate buffer (pH 5.0)) after dewaxing and hydration, counterstained with methyl green and mounted with hydromounting medium. Stained sections were observed and imaged under fluorescent microscopy and DP70 camera (Olympus, Japan). Osteoblast number relative to bone tissue area (N.Ob./T.Ar, #/mm2) was measured in sections stained with H&E. Total collagen positive area was measured in sections stained histochemically for total collagen. Osteoclast surface along the bone surface (Oc.S/B.S, %) was measured in sections stained histochemically for TRAP. All the histomorphometric analysis of each group were carried out using a Northern Eclipse analysis software system described previously [27,28] and statistically analyzed with SPSS 13.0 software.
Immunohistochemistry
Following dewaxing of the paraffin sections in xylene and hydration in a descending ethanol series, antigen retrieval was performed by hyaluronidase digestion for 30 mins. After washing with phosphate buffered saline solution (PBS) 0.1 M at pH 7.4, sections were immersed in 3% hydrogen peroxide for one hour to block endogenous peroxidase and treated with 10% goat serum for 1 hour to block nonspecific binding sites. Then sections were subjected to incubation with the following primary antibodies: Primary antibodies against type Ι collagen (Southern Biotech, Birmingham, AL, USA) were used followed by secondary antibodies (Biotinylated goat anti rabbit or rabbit antigoat IgG; Sigma-Aldrich) for 1 hour at room temperature.
Real-time RT-PCR and Western blotting
For examination of gene expression at RNA level, total RNA was isolated from mouse long bones using Trizol reagent (Invitrogen, Inc., Carlsbad, CA, USA) according to the manufacturer’s protocol. Reverse-transcription reactions were performed using the SuperScript First-Strand Synthesis System (Invitrogen), as described previously [29]. Real-time PCR was performed using a LightCycler system (Roche, Indianapolis, IN, USA) as described previously [30]. For examination of protein expression level, whole cell lysates (15 μg) from long bone tissues were loaded in 10% SDS-PAGE gels, transferred to PVDF membranes. Membranes were blotted with primary antibodies against SOD1 (Abcam), SOD2 (Novus Biological), Prdx1 (Santa Cruz Biotechnol-ogy, Inc.), Prdx IV (Santa Cruz Biotechnology, Inc), p53 (Cell Signaling Technology), p21 (Santa Cruz Biotechnology, Inc.), γ-H2AX (Ser139) (Cell Signaling Technology), CHK2 (Novus Biological), NFκB-p65 (Cell Signaling Technology, Beverly, MA, USA) or β-actin (Bioworld Technology, St. Louis Park, MN, USA) were used as loading control. Immunoblotting was carried out as described previously [27]. Bands were quantitated by Scion Image Beta 4.02 (Scion Corporation, Frederick, MD, USA). All the experiments were repeated independently for three times.
Cell cultures
Bone marrow cells were flushed out from femurs and tibias. For fibroblast colony-forming unit (CFU-F) and ALP-expressing colony-forming unit (CFU-fap) assays, total bone marrow cells were cultured in 10 cm dishes at 1×106 cells/dish in 10 mL of a modified essential medium (a-MEM) containing 10% fetal calf serum (FBS) (Hyclone Laboratories, Logan, UT, USA), 50 mg/mL ascorbic acid and 10 mM β-glycerophosphate for 28 days. At the end of the culture period, cells were stained for CFU-f or CFU-fap. Cell cycle fractions (G0, G1, S, and G2M phases) were determined by flow cytometry. After cultured for 12 days in the presence of medium containing 10% FBS, BM-MSC were removed and a Cell-Light EdU Apollo 488 in vitro imaging kit was used to investigate the EdU expression levels in each group. Briefly, 100 ml EdU medium (50 µM) was added to each well. After incubation for 2 h, the EdU medium was removed and BM-MSC were washed twice with PBS and fixed with 75% ethanol. After lysing with Triton X-100 (0.5%) for 15 min, staining solution was added to each well and incubated for 30 min. DAPI was used to stain the cell nuclei. EdU expression levels were examined by immunofluorescence staining and the area of stained EdU-positive nodules relative to the total culture surface was measured by Image-Pro Plus (IPP) analysis. Ten images were captured for each well, and the mean percentage was calculated.
Tissue reactive oxygen species (ROS) examination
Thymus tissues were homogenized in 1 ml cold PBS. Cells were filtered and incubated with 5 mM diacetyl dichlorofluorescein (DCFDA) (Invitrogen) and 10% FBS for 30 min at 37°C. Then cells were analyzed with a FACS calibur flow cytometer (Becton Dickinson, Heidelberg, Germany) for the mean fluorescent intensity (MFI) and relevant fluorescent intensity to MFI of shame-operated mice [29].
Statistical analysis
Data were presented as mean ± SEM. Differences between groups were compared using one-way ANOVA for comparisons. Statistic differences were considered to be significant when P value was <0.05.
Results
Effect of PQQ on ORX-induced osteoporosis
To examine the effect of PQQ on ORX-induced osteoporosis, we first confirm that the ORX mouse model was established successfully by evaluating the bone phenotype of ORX mice and examining the bone density and bone volume of vertebrae from ORX compared with those from shame-operated mice using X-ray and micro-CT, respectively. Data showed that ORX mice have decreased bone density (Figure 1A, 1C) and bone volume (Figure 1B, 1D) of T9-T12 vertebrae at 48 weeks after surgery. PQQ supplement in the diet after ORX surgery partially rescued the decreased bone mineral density (Figure 1A, 1C) and bone volume (Figure 1B, 1D) compared with the shame group. Data suggest that PQQ could prevent the bone loss in ORX mice.
Figure 1.
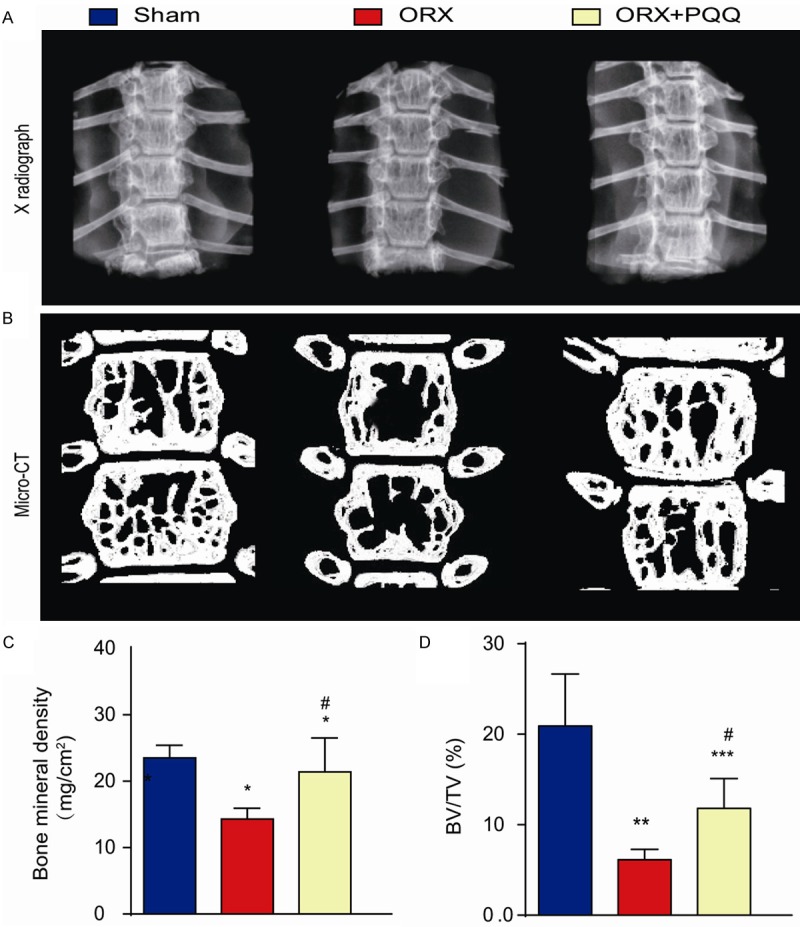
Effect of PQQ on ORX-induced osteoporosis. 2-month-old C57BL/6J wild-type mice were received ORX surgery at both sides or shame surgery. PQQ supplementary diet was given to ORX mice after surgery. T9-T12 vertebrae were harvested 48 weeks after PQQ treatment. A. Representative X-ray images. B. Representative micro-CT scans. C. Bone density by densitometry analysis on X-ray images. D. Bone volume (BV/TV) from micro-CT analysis. *, P<0.05; **, P<0.01; ***, P<0.001, vs shame group; #, P<0.05, vs ORX mice.
Effect of PQQ on osteoblastic bone formation in ORX mice
To investigate whether PQQ prevents bone loss induced by ORX through regulating osteoblastic bone formation, we assessed the number of osteoblasts, the deposition of total and type I collagen in mice with or without PQQ supplement by staining with H&E, histochemically for total collagen and immunohistochemically for type I collagen at 48 weeks after PQQ supplement (Figure 2A-C). Our results showed that the positive areas of total and type I collagen were decreased significantly in ORX mice compared to shame mice, however, they were increased significantly in PQQ-treated ORX mice compared to untreated ORX mice (Figure 2D). To further investigate the change of bone volume is due to osteoblast number change, we measured and compared the osteoblast number among 3 groups. Our results showed that the alterations of osteoblast number were consistent with the alterations of the positive areas of total and type I collagen (Figure 2E). These results suggest that PQQ prevented ORX-induced osteoporosis through promoting osteoblastic bone formation in ORX mice.
Figure 2.
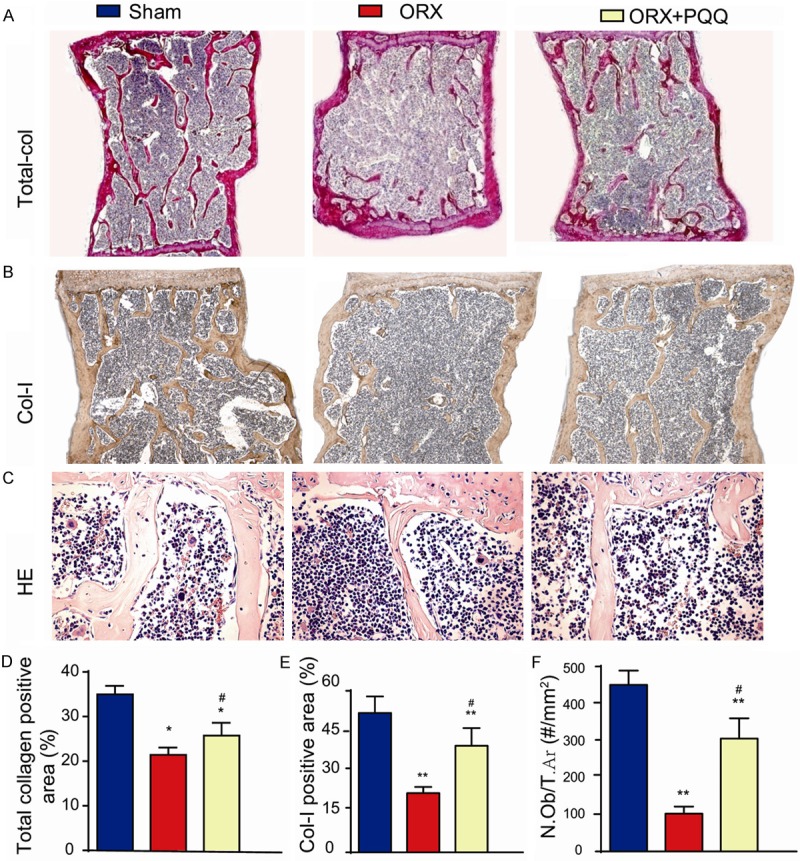
Effect of PQQ on osteoblastic bone formation of ORX mice. (A) T12 vertebrae sections from shame-operated mice, ORX mice and PQQ treated ORX mice were stained histochemically for total collagen (×100); (B) Immunohistochemically for Col-I (×100) and (C) with H&E (×200). (D) The relative total collagen positive area to tissue area was counted from (A). (E) The relative Col-I positive area to tissue area was counted from (B). (F) The osteoblast number were counted in H&E stained vertebrae sections. *, P<0.05; **, P<0.01, vs shame-operated mice; #, P<0.05, vs ORX mice.
Effect of PQQ on osteoclastic bone resorption in ORX mice
To investigate whether PQQ prevents bone loss induced by ORX through inhibiting osteoclastic bone resorption of ORX mice, we assessed the osteoclast surface along the bone surface by histochemical staining for TRAP (Figure 3A). We found that the osteoclast surface was increased significantly in ORX mice compared to shame-operated mice, which was inhibited by PQQ supplement in ORX mice (Figure 3B). Meanwhile, the gene expression the receptor activator of nuclear factor-kappa B ligand (RANKL) levels and the ratio of RANKL/osteoprotegerin (OPG) in T12 vertebrate were increased in ORX mice compared to shame-operated mice, which were reduced by PQQ supplement in ORX mice (Figure 3C, 3D). These results suggest that PQQ not only promoted osteoblastic bone formation, but also inhibited the osteoclastic bone resorption in ORX mice.
Figure 3.
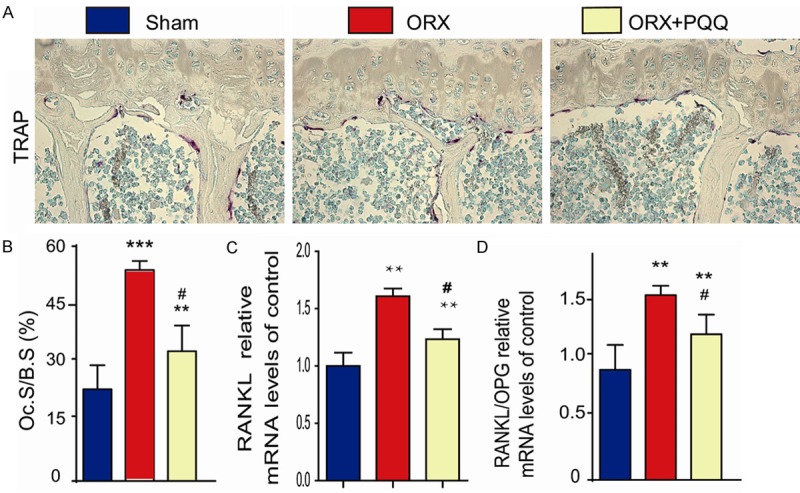
Effect of PQQ on osteoclastic bone resorption of ORX mice. (A) T12 vertebrae sections from shame-operated, ORX and PQQ treated ORX mice were histochemically stained for TRAP. (B) The osteoclast surface along the trabecular bone surface was measured. (C) The mRNA expression of RANKL and (D) The ratio of RANKL/OPG in T10 vertebral bone tissue of shame-operated mice, ORX and PQQ treated ORX mice was examined by qPCR. **, P<0.01; ***, P<0.001, vs shame-operated mice; #, P<0.05, vs ORX mice.
Effect of PQQ on BM-MSC proliferation and differentiation into osteoblasts
To investigate whether increased osteoblastic bone formation in ORX mice by PQQ supplement is associated with the alterations of BM-MSC proliferation and differentiation, we performed the CFU-f, EDU incorporation and CFU-fap assays by ex vivo total bone marrow cell cultures (Figure 4A-C). Our data revealed that the percentage of CFU-f relative area and EDU positive cells are significantly reduced in ORX mice compared to shame-operated mice, but partially rescued by PQQ supplement in ORX mice (Figure 4A, 4D, 4C, 4F). These results suggest that the proliferation of BM-MSCs in ORX mice was impaired, but was partially rescued by PQQ treatment. Furthermore, we also found that the percentage of CFU-fap relative area was reduced in ORX mice compared to shame-operated mice, but partially rescued by PQQ supplement in ORX mice (Figure 4B, 4E). Consistently, we also measured the expressions of osteoblast marker genes and found the reduced expressions of osteopontin (OPN), Runx2, type I collagen and osteocalcin in long bones from ORX mice compared to those from shame-operated mice, which were partially rescued by PQQ supplement in ORX mice (Figure 4G-J). Thus, PQQ can increase osteoblastic bone formation in ORX mice by stimulating the proliferation and differentiation of BM-MSCs.
Figure 4.
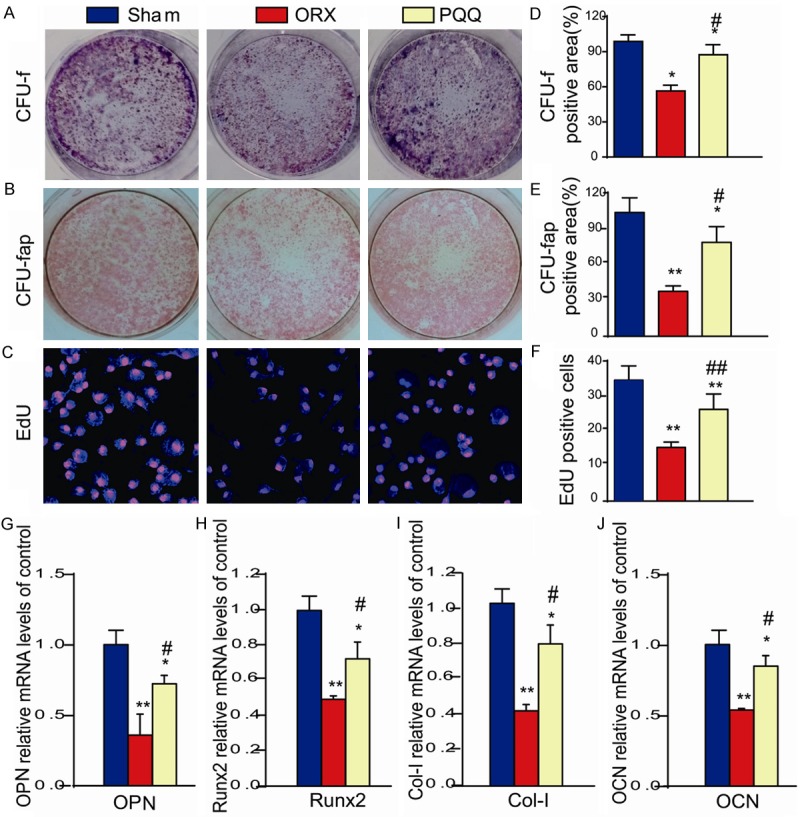
Effect of PQQ on BM-MSC proliferation and differentiation into osteoblasts. (A) CFU-f and (B) CFU-fap. (C)CFU-f cells were incubated with EDU for 2 hours and cells were stained for EDU with anti-EDU antibody. (D) The CFU-f positive area/dish area was counted. (E) The CFU-fap positive area/dish area was counted. (F) The EDU+ CFU-f cells was counted. (G-J) The mRNA expressions of osteoblast marker genes, OPN, Runx2, Col-I and OCN were examined by real-time RT-PCR in long bones. *, P<0.05; **, P<0.01, vs shame-operated mice; #, P<0.05; ##, P<0.01, vs ORX mice.
Effect of PQQ on redox balance
A number of studies showed that oxidative stress induced ROS expression results in the decreased osteoblast differentiation and MSC proliferation [31-36], but increased osteoblast apoptosis [37-40] and osteoclast formation [41,42]. To investigate whether the effect of PQQ on osteoblastic bone formation and osteoclastic bone resorption is associated with the regulation of oxidative stress, we measured the ROS levels in thymus (Figure 5A) and the protein levels of antioxidant enzymes in tibiae (Figure 5C). ROS levels were increased significantly in thymus tissues from ORX mice, while they were reduced significantly by PQQ supplement in ORX mice, but did not return to the level of shame-operated mice (Figure 5A, 5B). Protein levels of antioxidant enzymes, including SOD-1, SOD-2, Prdx I and Prdx IV were down-regulated in ORX mice, which were partially rescued by PQQ supplement in ORX mice (Figure 5C-G). To further examine the effect of PQQ on osteoblastic and osteoclastic cell function, we examined the alterations of the levels of DNA damage response related proteins, including γ-H2AX, p53 and Chk2 expression and osteoclast differentiation regulator, NF-κB-p65. Data showed that the expression levels of γ-H2AX, p53, Chk2, and NF-κB-p65 were up-regulated significantly in ORX mice, but they were down-regulated by PQQ supplement (Figure 5C, 5H-K). Thus, our results indicate that PQQ can stimulate osteoblastic bone formation and inhibit osteoclastic bone resorption in ORX mice by inhibiting oxidative stress and DNA damage.
Figure 5.
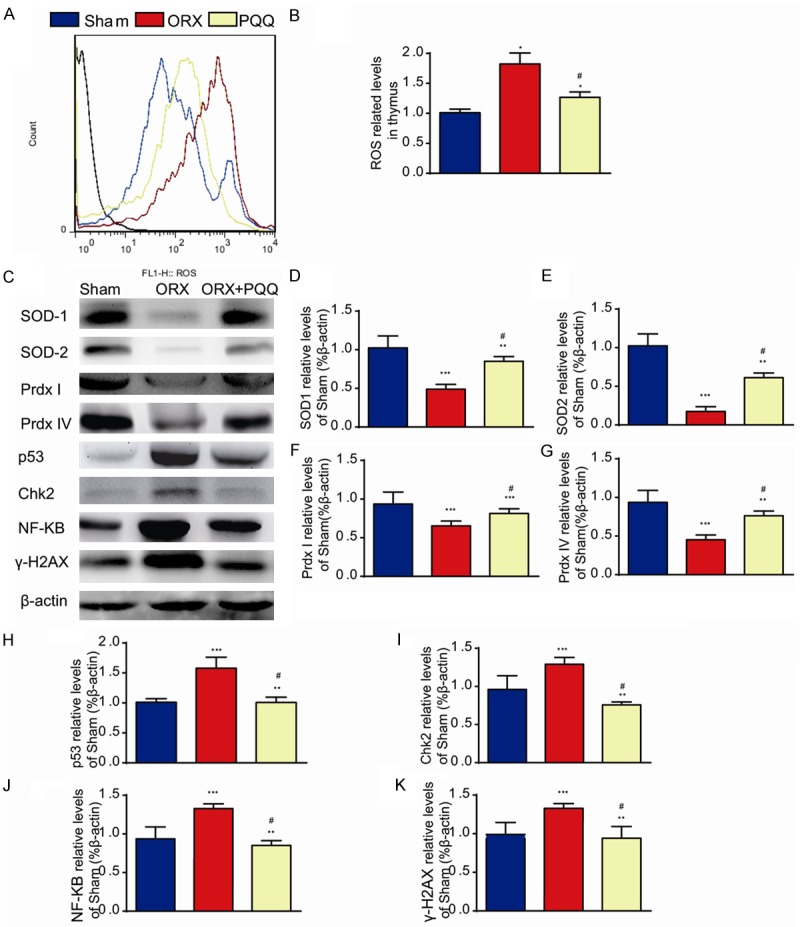
Effect of PQQ on redox balance. (A) Thymus tissues from shame-operated, ORX and PQQ treated ORX mice were examined for the ROS level by FACS analysis and (B). ROS related levels. (C) The expressions of antioxidant enzymes SOD-1, SOD-2, Prdx I and Prdx IV and target proteins of ROS, p53, Chk2, γ-H2AX and NF-κB in long bone by Western blots. (D-K) The densitometry analysis of western blots from 3 independent experiments. *, P<0.05; **, P<0.01; ***, P<0.001, vs shame-operated mice; #, P<0.05, vs ORX mice.
Discussion
In the past ten years, osteoporosis has been becoming to a major health concern for aged men [43]. Osteoporosis reduces the bone strength with increased susceptibility to fracture. Osteoporotic fractures lead to increased mortality and morbidity [44]. Testosterone deficiency has been considered one of important causes for osteoporosis in aged men, but the pathophysiology is not well investigated. Unlike aged women, the medications of osteoporosis in aged men are limited. Most treatments used in men are based on clinic evidences of fracture reduction in women [45]. Testosterone replacement treatment is limited by its side effect [46]. Therefore, the development of new therapeutic approaches for testosterone deficiency-induced osteoporosis is a critical clinical need.
There are three potential targets of testosterone which influence the bone development. First, testosterone directly binds to its receptor on osteoblasts to stimulate the osteoblast proliferation and differentiation. Second, testosterone is transformed to be dihydrotestosterone by 5α-reductase, which was proved to have highest affinity with testosterone receptors. Third, testosterone is transformed to be estrogen by aromatase, which binds to estrogen receptors [43]. In the present study, we used 2-month-old male WT mice, who have the peak bone volume during development, to generate the ORX model and observed the smaller size and decreased bone volume in ORX mice, compared with shame-operated mice, which indicate the osteoporosis and aging phenomena in ORX mice and showed the successful establishment of animal model for testosterone deficiency-induced osteoporosis. In this study, we measured the alterations of bone volumes in shame-operated mice, ORX mice and PQQ treated ORX mice at 13 months of age. In such age mice, almost no any trabecular bone was existed at the metaphyseal region of long bones including famous and tibiae in above 3 group mice, so that in this study we measured the trabecular bone volume in vertebral bodies. We examined the osteoblastic bone formation and osteoclastic bone resorption in ORX mice. Both decreased osteoblast formation and increased osteoclast resorption were found in ORX mice, which may respond to the decreased bone volumes. However, which played the major role in ORX-induced osteoporosis remains to be determined. To understand the cellular mechanism in which the osteoblastic bone formation was affected by testosterone deficiency, we examined the proliferation and differentiation of MSCs into osteoblasts. Our data clearly demonstrated that testosterone deficiency suppressed the proliferation of MSCs and their differentiation into osteoblasts as shown decreased CFU-f and CFU-fap formation efficiency and down-regulated expression levels of osteoblast differentiation marker genes. Therefore, results from this study suggest that the reduction of osteoblastic bone formation with decreased MSC proliferation and differentiation contributed to testosterone deficiency-induced osteoporosis.
Recently, epidemiological and mechanistic studies indicate that aging and its subsequent change in ROS are the major cause of sex hormone deficiency induced osteoporosis [47]. In aged men, there is no doubt that the level of testosterone was decreased by age. On the other hand, the decreased level of testosterone accelerates the aging progress and exogenous testosterone administrations attenuate the ROS level in aged mice [2]. Our results confirmed that testosterone deficiency caused high level of ROS and osteoporosis.
To date, ROS were considered to be exclusively harmful to proteins, lipids and DNA leading to cell death. A large effort of ROS on aged osteoporosis revealed several genes and signaling pathways which contribute to regulation of MSC proliferation, differentiation and apoptosis. Wnt/β-catenin signaling pathway is essential for MSC proliferation and differentiation [48]. In addition to promoting osteoblastogenesis, Wnt/β-catenin signaling also inhibits adipogenesis by blocking the expression of peroxisomal proliferator-activated receptor-γ (PPARγ) and cEBPα [49] and osteoclastogenesis and bone resorption by increasing the expression of osteoprotegerin (OPG) [50]. Wnt/β-catenin signaling has been reported to associate with oxidative stress via forkhead box O (FOXO) transcription factors [51]. In the aged mice, it was found that FOXO target genes were up-regulated, but the wnt/β-catenin target genes were down-regulated. ROS activated the transcription of FOXO and its target genes which requires β-catenin binding to FOXO. As a result, ROS induced FOXO target gene transcription at the expanse of wnt, which also required for canonical wnt/β-catenin/TCF mediated transcription [47]. On the another hand, ROS caused oxidized lipids may act as the ligand of PPARγ, promote the binding of PPARγ to β-catenin and decrease level of β-catenin in cells, which then attenuated the wnt directed signaling activation [31]. Additionally, ROS was also associated with TNF induced osteoblast apoptosis through PKCβ/JNK/p53 signaling cascade. PKCβ inhibitor inhibits the activation of JNK signaling and JNK inhibitor abrogates the TNF induced apoptosis in osteoblasts [51]. In osteoclast, ROS not only decreased OPG expression by inhibiting wnt/β-catenin signaling, but also directly involved in RANKL induced NFκB activation and osteoclast formation [41,42]. Therefore, ROS have diverse effect on osteoblasts and osteoclasts, and consequently inhibited the osteoblastic bone formation and promoted osteoclastic bone resorption.
To prevent the damage on bone by ROS, strategies involving in enzymatic reactions and altered ROS related genes expression were utilized [17]. PQQ as a nature herb derivates is an effective antioxidant to eliminate the free radicals and decrease the ROS level in vivo [52]. To investigate whether PQQ can rescue the osteoporotic bone phenotype in ORX mice, we treated the ORX mice with PQQ in supplementary diet for 48 weeks and found that PQQ treatment did partially rescued the ORX mice in body size, cortical bone thickness (data not shown), trabecular bone number and thickness in vertebrae. The increased bone volume can attribute to increased osteoblastic bone formation and/or decreased osteoclastic bone resporption, which were further examined by histological or immunohistological staining. Data suggest that PQQ treatment not only increased the osteoblastic bone formation, but also reduced the osteoclastic bone resorption. To understand the cellular mechanism in which PQQ increased the osteoblastic bone formation, we examined the MSC proliferation and osteoblast differentiation in PQQ treated ORX mice. Ex vivo cell cultures for CFU-f and CFU-fap revealed that PQQ increased the MSC proliferation and osteoblast differentiation in ORX mice, which responded to the increased the osteoblastic bone formation after PQQ treatment in ORX mice. EDU incorporation analysis in CFU-f cells and osteoblast differentiation marker gene expression confirmed these findings.
ROS as discussed above influenced the MSC proliferation, osteoblast differentiation and apoptosis, and promoted osteoclast formation as well. It was reported previously that antioxidants treatment prevented the estrogen deficiency induced bone loss, in which ROS was also involved [53]. In the present study, we investigated whether PQQ treatment can affect the oxidative stress by examining the ROS level, the protein expression levels of antioxidants and the downstream protein expressions. Our data suggest that PQQ treatment could efficiently suppress the ROS level and increase expression levels of antioxidant enzymes SOD1, SOD2, Prdx I and Prdx IV in ORX mice. Next, we examined the downstream proteins of oxidative stress and found that PQQ suppressed the higher expressions of apoptotic regulator, p53 [51], cyclin dependent kinase inhibitor, Chk2 [54] and DNA damage response related protein γ-H2AX in ORX mice, which infer that PQQ treatment did regulated cell proliferation and apoptosis in bone through inhibiting oxidative stress. ROS was reported to be produced by osteoclasts with RANKL stimulation [55] and then ROS leads to the activation of NFκB signaling and osteoclast formation as secondary messengers [56]. In terms of inhibition of ROS by PQQ treatment, we also found that the expression of NF-κB-p65 was also suppressed by PQQ treatment in ORX mice. Therefore, these data suggest that PQQ treatment inhibited osteoclast bone resorption in ORX mice through inhibiting ROS activated NFκB signaling.
In conclusion, this study demonstrated that the PQQ plays a prevention role in testosterone deficiency-induced osteoporosis by inhibiting oxidative stress and DNA damage, cell apoptosis and promoting the MSC proliferation and differentiation into osteoblasts and by inhibiting the NF-κB signaling in bone to reduce the osteoclastic bone resorption. Our results from this study provided experimental evidence for the clinical application of PQQ to treat osteoporosis in aged men.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (No. 81471501 & 81230009), from the National Basic Research Program of China (2014CB942900) to D.M.
Disclosure of conflict of interest
None.
References
- 1.Martin-Fernandez M, Martinez E, Diaz-Curiel M, Guede D, Caeiro JR, De la Piedra C. Effects of PTH (1-84) on bone quality in a validated model of osteoporosis due to androgenic deprivation. Aging Male. 2014;17:42–50. doi: 10.3109/13685538.2013.821697. [DOI] [PubMed] [Google Scholar]
- 2.Drake MT, Khosla S. Male osteoporosis. Endocrinol Metab Clin North Am. 2012;41:629–641. doi: 10.1016/j.ecl.2012.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Melton LJ 3rd, Khosla S, Achenbach SJ, O’Connor MK, O’Fallon WM, Riggs BL. Effects of body size and skeletal site on the estimated prevalence of osteoporosis in women and men. Osteoporos Int. 2000;11:977–983. doi: 10.1007/s001980070037. [DOI] [PubMed] [Google Scholar]
- 4.Kamel HK. Male osteoporosis: new trends in diagnosis and therapy. Drugs Aging. 2005;22:741–748. doi: 10.2165/00002512-200522090-00003. [DOI] [PubMed] [Google Scholar]
- 5.Ames BN, Gold LS, Willett WC. The causes and prevention of cancer. Proc Natl Acad Sci U S A. 1995;92:5258–5265. doi: 10.1073/pnas.92.12.5258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Basu S, Michaelsson K, Olofsson H, Johansson S, Melhus H. Association between oxidative stress and bone mineral density. Biochem Biophys Res Commun. 2001;288:275–279. doi: 10.1006/bbrc.2001.5747. [DOI] [PubMed] [Google Scholar]
- 7.Hitchon CA, El-Gabalawy HS. Oxidation in rheumatoid arthritis. Arthritis Res Ther. 2004;6:265–278. doi: 10.1186/ar1447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Huang H, Manton KG. The role of oxidative damage in mitochondria during aging: a review. Front Biosci. 2004;9:1100–1117. doi: 10.2741/1298. [DOI] [PubMed] [Google Scholar]
- 9.Nohl H. Involvement of free radicals in ageing: a consequence or cause of senescence. Br Med Bull. 1993;49:653–667. doi: 10.1093/oxfordjournals.bmb.a072638. [DOI] [PubMed] [Google Scholar]
- 10.Pelicano H, Carney D, Huang P. ROS stress in cancer cells and therapeutic implications. Drug Resist Updat. 2004;7:97–110. doi: 10.1016/j.drup.2004.01.004. [DOI] [PubMed] [Google Scholar]
- 11.Takano H, Zou Y, Hasegawa H, Akazawa H, Nagai T, Komuro I. Oxidative stress-induced signal transduction pathways in cardiac myocytes: involvement of ROS in heart diseases. Antioxid Redox Signal. 2003;5:789–794. doi: 10.1089/152308603770380098. [DOI] [PubMed] [Google Scholar]
- 12.Fragonas E, Pollesello P, Mlinarik V, Toffanin R, Grando C, Godeas C, Vittur F. Sensitivity of chondrocytes of growing cartilage to reactive oxygen species. Biochim Biophys Acta. 1998;1425:103–111. doi: 10.1016/s0304-4165(98)00055-5. [DOI] [PubMed] [Google Scholar]
- 13.Feng YF, Wang L, Zhang Y, Li X, Ma ZS, Zou JW, Lei W, Zhang ZY. Effect of reactive oxygen species overproduction on osteogenesis of porous titanium implant in the present of diabetes mellitus. Biomaterials. 2013;34:2234–2243. doi: 10.1016/j.biomaterials.2012.12.023. [DOI] [PubMed] [Google Scholar]
- 14.Kondo H, Limoli C, Searby ND, Almeida EA, Loftus DJ, Vercoutere W, Morey-Holton E, Giedzinski E, Mojarrab R, Hilton D, Globus RK. Shared oxidative pathways in response to gravity-dependent loading and gamma-irradiation of bone marrow-derived skeletal cell progenitors. Radiats Biol Radioecol. 2007;47:281–285. [PubMed] [Google Scholar]
- 15.Lee JS, Lee MO, Moon BH, Shim SH, Fornace AJ Jr, Cha HJ. Senescent growth arrest in mesenchymal stem cells is bypassed by Wip1-mediated downregulation of intrinsic stress signaling pathways. Stem Cells. 2009;27:1963–1975. doi: 10.1002/stem.121. [DOI] [PubMed] [Google Scholar]
- 16.Wildburger R, Borovic S, Zarkovic N, Tatzber F. Post-traumatic dynamic changes in the antibody titer against oxidized low density lipoproteins. Wien Klin Wochenschr. 2000;112:798–803. [PubMed] [Google Scholar]
- 17.Zhou H, Newnum AB, Martin JR, Li P, Nelson MT, Moh A, Fu XY, Yokota H, Li J. Osteoblast/osteocyte-specific inactivation of Stat3 decreases load-driven bone formation and accumulates reactive oxygen species. Bone. 2011;49:404–411. doi: 10.1016/j.bone.2011.04.020. [DOI] [PubMed] [Google Scholar]
- 18.Stolzing A, Jones E, McGonagle D, Scutt A. Age-related changes in human bone marrow-derived mesenchymal stem cells: consequences for cell therapies. Mech Ageing Dev. 2008;129:163–173. doi: 10.1016/j.mad.2007.12.002. [DOI] [PubMed] [Google Scholar]
- 19.Filipovic B, Sosic-Jurjevic B, Ajdzanovic V, Pantelic J, Nestorovic N, Milosevic V, Sekulic M. The effects of sex steroids on thyroid C cells and trabecular bone structure in the rat model of male osteoporosis. J Anat. 2013;222:313–320. doi: 10.1111/joa.12013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ohwada K, Takeda H, Yamazaki M, Isogai H, Nakano M, Shimomura M, Fukui K, Urano S. Pyrroloquinoline quinone (PQQ) prevents cognitive deficit caused by oxidative stress in rats. J Clin Biochem Nutr. 2008;42:29–34. doi: 10.3164/jcbn.2008005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jun JH, Lee SH, Kwak HB, Lee ZH, Seo SB, Woo KM, Ryoo HM, Kim GS, Baek JH. N-acetylcysteine stimulates osteoblastic differentiation of mouse calvarial cells. J Cell Biochem. 2008;103:1246–1255. doi: 10.1002/jcb.21508. [DOI] [PubMed] [Google Scholar]
- 22.Kim HJ, Chang EJ, Kim HM, Lee SB, Kim HD, Su Kim G, Kim HH. Antioxidant alpha-lipoic acid inhibits osteoclast differentiation by reducing nuclear factor-kappaB DNA binding and prevents in vivo bone resorption induced by receptor activator of nuclear factor-kappaB ligand and tumor necrosis factor-alpha. Free Radic Biol Med. 2006;40:1483–1493. doi: 10.1016/j.freeradbiomed.2005.10.066. [DOI] [PubMed] [Google Scholar]
- 23.Lee YS, Choi EM. Apocynin stimulates osteoblast differentiation and inhibits bone-resorbing mediators in MC3T3-E1 cells. Cell Immunol. 2011;270:224–229. doi: 10.1016/j.cellimm.2011.05.011. [DOI] [PubMed] [Google Scholar]
- 24.Moon HJ, Ko WK, Jung MS, Kim JH, Lee WJ, Park KS, Heo JK, Bang JB, Kwon IK. Coenzyme q10 regulates osteoclast and osteoblast differentiation. J Food Sci. 2013;78:H785–891. doi: 10.1111/1750-3841.12116. [DOI] [PubMed] [Google Scholar]
- 25.Inada M, Matsumoto C, Miyaura C. [Animal models for bone and joint disease. Ovariectomized and orchidectomized animals] . Clin Calcium. 2011;21:164–170. [PubMed] [Google Scholar]
- 26.Zhang Z, Yin S, Xue X, Ji J, Tong J, Goltzman D, Miao D. Transplantation of bone marrow-derived mesenchymal stem cells rescues partially rachitic phenotypes induced by 1,25-Dihydroxyvitamin D deficiency in mice. Am J Transl Res. 2016;8:4382–4393. [PMC free article] [PubMed] [Google Scholar]
- 27.Miao D, Bai X, Panda D, McKee M, Karaplis A, Goltzman D. Osteomalacia in hyp mice is associated with abnormal phex expression and with altered bone matrix protein expression and deposition. Endocrinology. 2001;142:926–939. doi: 10.1210/endo.142.2.7976. [DOI] [PubMed] [Google Scholar]
- 28.Xue Y, Karaplis AC, Hendy GN, Goltzman D, Miao D. Genetic models show that parathyroid hormone and 1,25-dihydroxyvitamin D3 play distinct and synergistic roles in postnatal mineral ion homeostasis and skeletal development. Hum Mol Genet. 2005;14:1515–1528. doi: 10.1093/hmg/ddi160. [DOI] [PubMed] [Google Scholar]
- 29.Jin J, Lv X, Chen L, Zhang W, Li J, Wang Q, Wang R, Lu X, Miao D. Bmi-1 plays a critical role in protection from renal tubulointerstitial injury by maintaining redox balance. Aging Cell. 2014;13:797–809. doi: 10.1111/acel.12236. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 30.Sun W, Wu J, Huang L, Liu H, Wang R, Karaplis A, Goltzman D, Miao D. PTHrP nuclear localization and carboxyl terminus sequences modulate dental and mandibular development in part via the action of p27. Endocrinology. 2016;157:1372–1384. doi: 10.1210/en.2015-1555. [DOI] [PubMed] [Google Scholar]
- 31.Almeida M, Ambrogini E, Han L, Manolagas SC, Jilka RL. Increased lipid oxidation causes oxidative stress, increased peroxisome proliferator-activated receptor-gamma expression, and diminished pro-osteogenic Wnt signaling in the skeleton. J Biol Chem. 2009;284:27438–27448. doi: 10.1074/jbc.M109.023572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Coipeau P, Rosset P, Langonne A, Gaillard J, Delorme B, Rico A, Domenech J, Charbord P, Sensebe L. Impaired differentiation potential of human trabecular bone mesenchymal stromal cells from elderly patients. Cytotherapy. 2009;11:584–594. doi: 10.1080/14653240903079385. [DOI] [PubMed] [Google Scholar]
- 33.Monroe DG, McGee-Lawrence ME, Oursler MJ, Westendorf JJ. Update on Wnt signaling in bone cell biology and bone disease. Gene. 2012;492:1–18. doi: 10.1016/j.gene.2011.10.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Scaffidi P, Misteli T. Lamin A-dependent misregulation of adult stem cells associated with accelerated ageing. Nat Cell Biol. 2008;10:452–459. doi: 10.1038/ncb1708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Song L, Liu M, Ono N, Bringhurst FR, Kronenberg HM, Guo J. Loss of wnt/beta-catenin signaling causes cell fate shift of preosteoblasts from osteoblasts to adipocytes. J Bone Miner Res. 2012;27:2344–2358. doi: 10.1002/jbmr.1694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stolzing A, Scutt A. Age-related impairment of mesenchymal progenitor cell function. Aging Cell. 2006;5:213–224. doi: 10.1111/j.1474-9726.2006.00213.x. [DOI] [PubMed] [Google Scholar]
- 37.Almeida M, Han L, Ambrogini E, Weinstein RS, Manolagas SC. Glucocorticoids and tumor necrosis factor alpha increase oxidative stress and suppress Wnt protein signaling in osteoblasts. J Biol Chem. 2011;286:44326–44335. doi: 10.1074/jbc.M111.283481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bonewald LF. The amazing osteocyte. J Bone Miner Res. 2011;26:229–238. doi: 10.1002/jbmr.320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Mariappan N, Elks CM, Sriramula S, Guggilam A, Liu Z, Borkhsenious O, Francis J. NF-kappaB-induced oxidative stress contributes to mitochondrial and cardiac dysfunction in type II diabetes. Cardiovasc Res. 2010;85:473–483. doi: 10.1093/cvr/cvp305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Trinei M, Berniakovich I, Beltrami E, Migliaccio E, Fassina A, Pelicci P, Giorgio M. P66Shc signals to age. Aging (Albany NY) 2009;1:503–510. doi: 10.18632/aging.100057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ha H, Kwak HB, Lee SW, Jin HM, Kim HM, Kim HH, Lee ZH. Reactive oxygen species mediate RANK signaling in osteoclasts. Exp Cell Res. 2004;301:119–127. doi: 10.1016/j.yexcr.2004.07.035. [DOI] [PubMed] [Google Scholar]
- 42.Teitelbaum SL, Ross FP. Genetic regulation of osteoclast development and function. Nat Rev Genet. 2003;4:638–649. doi: 10.1038/nrg1122. [DOI] [PubMed] [Google Scholar]
- 43.Vanderschueren D, Van Herck E, De Coster R, Bouillon R. Aromatization of androgens is important for skeletal maintenance of aged male rats. Calcif Tissue Int. 1996;59:179–183. doi: 10.1007/s002239900106. [DOI] [PubMed] [Google Scholar]
- 44.Nih Consensus Development Panel on Osteoporosis Prevention Diagnosis and Therapy. Osteoporosis prevention, diagnosis, and therapy. JAMA. 2001;285:785–795. [Google Scholar]
- 45.Walsh JS, Eastell R. Osteoporosis in men. Nat Rev Endocrinol. 2013;9:637–645. doi: 10.1038/nrendo.2013.171. [DOI] [PubMed] [Google Scholar]
- 46.Basaria S, Coviello AD, Travison TG, Storer TW, Farwell WR, Jette AM, Eder R, Tennstedt S, Ulloor J, Zhang A, Choong K, Lakshman KM, Mazer NA, Miciek R, Krasnoff J, Elmi A, Knapp PE, Brooks B, Appleman E, Aggarwal S, Bhasin G, Hede-Brierley L, Bhatia A, Collins L, LeBrasseur N, Fiore LD, Bhasin S. Adverse events associated with testosterone administration. N Engl J Med. 2010;363:109–122. doi: 10.1056/NEJMoa1000485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Almeida M, Han L, Martin-Millan M, O’Brien CA, Manolagas SC. Oxidative stress antagonizes Wnt signaling in osteoblast precursors by diverting beta-catenin from T cell factor-to forkhead box O-mediated transcription. J Biol Chem. 2007;282:27298–27305. doi: 10.1074/jbc.M702811200. [DOI] [PubMed] [Google Scholar]
- 48.Kolpakova E, Olsen BR. Wnt/beta-catenin--a canonical tale of cell-fate choice in the vertebrate skeleton. Dev Cell. 2005;8:626–627. doi: 10.1016/j.devcel.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 49.Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol. 2006;7:885–896. doi: 10.1038/nrm2066. [DOI] [PubMed] [Google Scholar]
- 50.Glass DA 2nd, Bialek P, Ahn JD, Starbuck M, Patel MS, Clevers H, Taketo MM, Long F, McMahon AP, Lang RA, Karsenty G. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev Cell. 2005;8:751–764. doi: 10.1016/j.devcel.2005.02.017. [DOI] [PubMed] [Google Scholar]
- 51.Nozaki H, Hiramatsu K, Arimori S. Platelet aggregation and intraplatelet adenine nucleotides in diabetic retinopathy. Scand J Clin Lab Invest. 1987;47:441–445. [PubMed] [Google Scholar]
- 52.Zhang P, Xu Y, Li L, Jiang Q, Wang M, Jin L. In vitro protective effects of pyrroloquinoline quinone on methylmercury-induced neurotoxicity. Environ Toxicol Pharmacol. 2009;27:103–110. doi: 10.1016/j.etap.2008.08.010. [DOI] [PubMed] [Google Scholar]
- 53.Lean JM, Davies JT, Fuller K, Jagger CJ, Kirstein B, Partington GA, Urry ZL, Chambers TJ. A crucial role for thiol antioxidants in estrogen-deficiency bone loss. J Clin Invest. 2003;112:915–923. doi: 10.1172/JCI18859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Neumann J, Yang Y, Kohler R, Giaisi M, Witzens-Harig M, Liu D, Krammer PH, Lin W, Li-Weber M. Mangrove dolabrane-type of diterpenes tagalsins suppresses tumor growth via ROS-mediated apoptosis and ATM/ATR-Chk1/Chk2-regulated cell cycle arrest. Int J Cancer. 2015;137:2739–2748. doi: 10.1002/ijc.29629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Steinbeck MJ, Appel WH Jr, Verhoeven AJ, Karnovsky MJ. NADPH-oxidase expression and in situ production of superoxide by osteoclasts actively resorbing bone. J Cell Biol. 1994;126:765–772. doi: 10.1083/jcb.126.3.765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lee SH, Ding Y, Yan XT, Kim YH, Jang HD. Scopoletin and scopolin isolated from Artemisia iwayomogi suppress differentiation of osteoclastic macrophage RAW 264.7 cells by scavenging reactive oxygen species. J Nat Prod. 2013;76:615–620. doi: 10.1021/np300824h. [DOI] [PubMed] [Google Scholar]