

Abstract
The aim of the present study was to determine the repairing effects of intercellular adhesion molecule (ICAM)-1-expressing mesenchymal stem cells (MSCs) in mice with autoimmune thyroiditis. Following induction of an experimental autoimmune thyroiditis (EAT) model, mice were randomly divided into the following groups (n=10 each): i) Normal control; and experimental groups that were subject to EAT induction, including ii) EAT model; and iii) primary MSC; iv) C3H10T1/2/MSC; v) C3H10T1/2-MIGR1/MSC; and vi) C3H10T1/2-MIGR1-ICAM-1/MSC, which were all administered the relevant cells. MSCs were administered via the caudal vein. A blood sample was harvested from the angular vein of each animal 28 days post-treatment and ELISA was used to determine the serum total triiodothyronine, total thyroxine (T4), thyroid-stimulating hormone (TSH), anti-thyroid peroxidase (TPOAb), anti-thyroid microsomal (TMAb) and anti-thyroglobulin (TGAb) antibodies. Hematoxylin and eosin staining was performed to evaluate injury of the thyroid gland by determining the size of the follicle, inflammatory infiltration, colloidal substance retention and epithelial injury. Reverse transcription-quantitative polymerase chain reaction was performed to determine the mRNA expression of interleukin (IL)-4, IL-10, IL-17 and interferon (INF)-γ. Western blot analysis was performed to determine the expression of p38 mitogen-activated protein kinase (p38) and extracellular signal-regulated kinase (ERK). To observe cellular migration in vivo, mice were divided into the following groups, (n=10 each), which were subject to EAT induction: i) CM-DiI-labeled primary MSC; ii) CM-DiI-labeled C3H10T1/2/MSC; iii) CM-DiI-labeled C3H10T1/2-MIGR1/MSC; and iv) CM-DiI-labeled C3H10T1/2-ICAM-1/MSC, which were all administered the relevant cells via the caudal vein. C3H10T1/2-ICAM-1/MSCs were able to ameliorate the expression of T4, TSH, TPOAb, TMAb and TGAb in vivo, attenuate thyroid follicle injury and decrease the splenic index in mice. They were also able to ameliorate the mRNA expression of IL-4, IL-10, IL-17 and INF-γ, and the modulation of the P38 and ERK-signaling pathways in the mouse spleen. Furthermore, ICAM-1 overexpression was able to modulate the nesting of MSCs in the thyroid gland and lung. These findings suggest that C3H10T1/2-ICAM-1/MSC may affect the differentiation, proliferation and migration of immunocytes through modulating the p38 and ERK signaling pathways, and that ICAM-1 may modulate the immunoregulatory effects of MSCs by affecting the migration of MSCs in vivo.
Keywords: mesenchymal stem cell, experimental autoimmune thyroiditis, intercellular adhesion molecule-1, nesting
Introduction
Autoimmune thyroiditis (AIT), which is also known as chronic lymphocytic thyroiditis or Hashimoto's thyroiditis, is an organ-specific T-cell-mediated disease with a prevalence of 5–10% in the Chinese population (1). Patients with early-stage AIT may present transient hyperthyrea; however in the advanced stage, hypothyroidism may occur (2,3). The pathogenesis of AIT is considered to be associated with various factors, including genetic and environmental factors, age, and sex hormone levels (4); however, its exact mechanism remains to be elucidated. In clinical practice, the treatment of AIT is predominantly dependent on thyroid hormone replacement therapy. However, patients who undergo such therapies typically require additional treatment, potentially for the rest of their lives, to repair potential injuries to the thyroid gland (5).
Mesenchymal stem cells (MSCs) are prototypical adult stem cells with the capacity to self-renew and differentiate in vivo. Recently, Cipriani et al (6) demonstrated that MSCs exhibit immunomodulatory and immunosuppressive properties, as they are able to regulate the immune reactions of the host via various signaling pathways. It was also demonstrated that MSCs were able to contribute to the regeneration of cells in injured organs via cellular migration. Therefore, these cells have been extensively used in tissue engineering due to their extensive distribution in vivo and their capacity for easy isolation and proliferation (7,8). Immunomodulatory properties have previously been reported to be a typical feature of MSCs due to a lack of expression of immuno-co-stimulatory factors such as the major histocompatibility complex class II antigen, cluster of differentiation (CD)80, CD40 and CD86, which supports that MSCs have a low immunogenicity (9). Extensive studies have been performed to investigate the transplantation of cells, tissues or organs in tissue engineering (10), which have also reported that intercellular cell adhesion molecule-1 (ICAM-1) is able to increase the immunosuppressive capacity of MSCs and contribute to the migration of MSCs in vitro (11). Furthermore, ICAM-1 may affect the differentiation of MSCs through modulating the mitogen-activated protein kinase (MAPK)-signaling pathway (12). The aim of the present study was to investigate the effects of ICAM-1-expressing MSCs on the repair of experimental AIT (EAT) mice.
Materials and methods
Animals and cell lines
A total of 60 female C57BL/6 mice (age, 6 weeks) with a body weight of 20±2 g were purchased from Vital River Laboratories Co., Ltd., (Beijing, China), provided with free access to food and water and raised in sterilized conditions for 1 week at 24±2°C in a humidity of 40–50% and a 12 h dark/light cycle. A total of 10 C57BL/6 mice (male, n=5; female, n=5; age, 1 week; weight, 4–5 g) used for the cultivation of primary MSCs were purchased from the Animal Center of the Academy of Military Medical Sciences (Beijing, China), and housed in the same conditions described above. C3H10T1/2 cells were purchased from the American Type Culture Collection (Manassas, VA, USA). The C3H10T1/2-MIGR1/MSC and C3H10T1/2-MIGR1-ICAM-1/MSC cell lines were constructed in the authors' laboratory at Tianjin Medical University General Hospital (Tianjin, China). Study protocols were approved by the Ethics Committee of Tianjin Medical University General Hospital (Tianjin, China).
Reagents
Porcine thyroglobulin, complete Freund's adjuvant and incomplete Freund's adjuvant were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Commercial ELISA kits for total triiodothyronine (TT3; cat. no. 80985; Crystal Chem, Inc., Downers Grove, IL, USA), total thyroxine (TT4; cat. no. 80983; Crystal Chem, Inc.), thyroid stimulating hormone (TSH; cat. no. RTC700R; Xinbosheng Biotech Co., Ltd., Shenzhen, China) and anti-thyroid peroxidase (TPOAb; cat. no. 05–50085; Zhuzhou Zeye Biotech Co., Ltd., Zhuzhou, China), anti-thyroid microsomal (TMAb; cat. no. KB12639; Shanghai Jianglai Biotech Co., Ltd., Shanghai, China) and anti-thyroglobulin antibodies (TGAb; cat. no. MA512408; Beinuo Life Science, Shanghai, China) were used. TRIzol reagent was purchased from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA). TaqDNA polymerase, dNTP mixture, PrimeScript reverse transcription kit (cat. no. 639505), RNase inhibitor and oligo d(T)15 primers were purchased from Takara Bio, Inc., (Otsu, Japan). Rabbit anti-mouse antibodies against total-mitogen-activated protein kinase p38 (p38; cat. no. 9212), phosphorylated (p)-P38 (cat. no. 9216), total extracellular signal-regulated kinase (ERK) (cat. no. 9101) and p-ERK (cat. no. 4696), and goat anti-rabbit horseradish peroxidase (HRP)-conjugated antibodies were purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA). Anti-mouse β-actin antibodies (cat. no. 3700) were also purchased from Cell Signaling Technology, Inc., and CM-DiI was purchased from Sigma-Aldrich (Merck KGaA). Type II collagenase, α-MEM medium and fetal bovine serum were purchased from Gibco (Thermo Fisher Scientific, Inc.).
Experimental design
The EAT model was established in 6-week-old C57BL/6 mice according to a previously described method (13). Subsequently, mice were randomly divided into the following groups (n=10 each): Normal control; EAT model; primary MSC group, which was subjected to EAT induction, followed by administration of MSCs (3×105) via the caudal vein; C3H10T1/2/MSC group with similar function of MSCs, which was subjected to EAT induction, followed by administration of C3H10T1/2/MSCs (3×105) via the caudal vein; C3H10T1/2-MIGR1/MSC group, which was subjected to EAT induction, followed by administration of C3H10T1/2-MIGR1/MSCs (3×105) via the caudal vein; and C3H10T1/2-MIGR1-ICAM-1/MSC group, which was subjected to EAT induction, followed by administration of C3H10T1/2-MIGR1-ICAM-1/MSCs (3×105) via the caudal vein. C3H10T1/2-MIGR1/MSCs were cells transfected with C3H10T1/2-MIGR1 plasmid, and C3H10T1/2-MIGR1-ICAM-1/MSC were cells transected with C3H10T1/2-MIGR1-ICAM-1 plasmid.
ELISA
Blood samples were harvested from the angular vein of each mouse at day 28 post-treatment to determine the serum TT3, TT4, TSH, TPOAb, TMAb and TGAb according to the manufacturer's protocol. All tests were performed a minimum of three times.
Hematoxylin and eosin (H&E) staining
Following anesthesia with 3% pentobarbital (Sigma-Aldrich; Merck KGaA), mice were sacrificed via cervical dislocation. An incision was subsequently made on the skin along the cervical median, and the muscles were separated to expose the thyroid gland. The gland was removed using an ophthalmic scissor. Thyroid gland tissues were fixed using 4% formalin and embedded in wax, followed by preparation of 4 µm-thick sections. H&E staining was subsequently performed to evaluate thyroid gland injury by evaluating the size of the follicles, inflammatory infiltration, colloidal substance retention and epithelial injury.
Calculation of splenic index
Following sacrifice, an incision was made under the inferior margin of the left rib to expose the spleen. The spleen was obtained using an ophthalmic scissor. The width, thickness and craniocaudal length of the spleen were determined and were used to calculate the splenic index according to the previous description (14).
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was extracted from splenic cells using TRIzol reagent according to the manufacturer's protocol. cDNA synthesis was performed using ~2 µg RNA with PrimeScript reverse transcription kit qPCR was performed using SYBR-Green on an Applied Biosystems 7500 Software v2.0.6 system (Thermo Fisher Scientific, Inc., Waltham, MA, USA) with the primers listed in Table I. PCR reactions were performed in a total volume of 10 µl containing 5 µl 2X SYBR Premix, 0.2 µl each specific primer to a final concentration of 200 nM, and 1 µl cDNA template. The PCR conditions consisted of denaturation at 94°C for 3 min, followed by 40 cycles of 94°C for 30 sec, 60°C for 30 sec, and 72°C for 60 sec. The mRNA level was normalized to that of β-actin, and all reactions were performed at least in triplicate. The amplification results for real-time PCR were calculated using the 2−ΔΔCq method (15).
Table I.
Primer sequences for the polymerase chain reaction amplification of IL-4, IL-10, IL-17 and INF-γ mRNA.
| Primer | Sequence (5′-3′) |
|---|---|
| IL-4 | Forward: CGAGTTGACCGTAACAGACAT |
| Reverse: CGTCTTTAGCCTTTCCAAGAAG | |
| IL-10 | Forward: CATTCATGGCCTTGTAGACACCTT |
| Reverse: TCTCCCCTGTGAAAATAAGAGCAAG | |
| Il-17 | Forward: GTCGTTCAGATTGAGAGACAAGG |
| Reverse: CTCATCCTTCAAAGACAGCCTCA | |
| INF-γ | Forward: CTGGTGGACCACTCGGATGA |
| Reverse: TTACTACCTTCTTCAGCAACAGCAA | |
| β-actin | Forward: GGAGATTACTGCCCTGGCTCCTA |
| Reverse: GACTCATCGTACTCCTGCTTGCTG |
IL, interleukin; INF, interferon.
Western blot analysis
Splenic tissues were homogenized at room temperature for 2 min in radioimmunoprecipitation assay buffer containing protease and phosphatase inhibitors (BB-3201-1; Bestbio, Shanghai, China). Protein concentration was determined using a bicinchoninic acid assay, and 40 µg protein per lane was separated by 10% SDS-PAGE and transferred to a Hybond-P membrane. The membrane was subsequently blocked with 5% non-fat milk and incubated with rabbit anti-mouse p-p38 (1:200), total-p38 (1:500), p-ERK (1:1,000) and total-ERK (1:200) antibodies overnight at 4°C. Subsequently, the mixture was incubated with the HRP-conjugated goat anti-rabbit antibody (1:1,000) for 1 h at room temperature. Following washing with Tris-buffered saline Tween-20, the bound primary antibody was visualized using enhanced chemiluminescence and exposed to X-ray film. The relative density of each protein was analyzed using TL120 imaging software (version v2009; TotalLab, Ltd., Newcastle upon Tyne, UK). The same membrane probed with anti-β-actin antibodies (1:2,000) served as the loading control. All experiments were performed a minimum of three times.
Determination of MSC migration in vivo
To determine the migration of MSCs in vivo, 1-week-old C57BL/6 mice were randomly divided into the following groups, subsequent to EAT induction (n=10 each): i) CM-DiI-labeled primary MSCs; ii) CM-DiI-labeled C3H10T1/2/MSC; iii) CM-DiI-labeled C3H10T1/2-MIGR1/MSC; and iv) CM-DiI-labeled C3H10T1/2-MIGR1-ICAM-1/MSC, which were administered the relevant cells via the caudal vein. For the CM-DiI labeling process, MSCs (2×106) were resuspended with 1 ml PBS and labeled with 2 µl CM-DiI at 37°C for 5 min. Subsequently, the mixture was incubated at 4°C for 5 min, and centrifuged at 179 × g for 10 min.
Following anesthesia with 3% pentobarbital (Sigma-Aldrich; Merck KGaA) and sacrifice via cervical dislocation, mice were fixed on an operating table to expose the chest, the chest was shaved and the fascia removed, and the sternum and ribs were removed to expose the heart. PBS was infused from the left ventricle to expel circulated blood from the right atrium. Subsequently, the lung and thyroid gland were washed using PBS, and fixed with 10% paraformaldehyde. Following embedding, 5-µm sections were cut and observed via fluorescence microscopy to determine the positive staining.
Statistical analysis
SPSS 18.0 software (SPSS, Inc., Chicago, IL, USA) was used to perform statistical analysis. Data are presented as the mean ± standard deviation, and Student's t-test was performed for inter-group comparisons. P<0.05 was considered to indicate a statistically significant difference.
Results
MSCs, especially C3H10T1/2-MIGR1-ICAM-1/MSCs, reverse the expression of T4, TSH, TPOAb, TMAb and TGAb in vivo
No statistical difference was observed in the TT3 levels between groups (Fig. 1A). For the expression of TT4, a significant decrease was observed in the EAT model group compared with that of normal controls (P<0.05). However, following administration of MSCs, the level of TT4 was markedly increased compared to the model group. Compared with the C3H10T1/2-MIGR1/MSC group, the level of TT4 was significantly increased in the C3H10T1/2-MIGR1-ICAM-1/MSC group (P<0.05; Fig. 1B). The expression of TSH significantly decreased in the C3H10T1/2-MIGR1-ICAM-1/MSC group compared with the C3H10T1/2-MIGR1/MSC group (P<0.05; Fig. 1C). Levels of TPOAb, TGAb, TMAb and TSH were significantly higher in the EAT model group compared with those of normal controls (all P<0.05; Fig. 1C-F). Similarly, following administration of MSCs, their expression was decreased, particularly in the C3H10T1/2-MIGR1-ICAM-1/MSC and MSC groups. Compared with the C3H10T1/2-MIGR1/MSC group, the levels of TPOAb, TGAb, TMAb and TSH were significantly decreased in the C3H10T1/2-MIGR1-ICAM-1/MSC group (all P<0.05).
Figure 1.

Determination of (A) TT3, (B) TT4, (C) TSH, (D) TPOAb, (E) TMAb and (F) TGAb levels by ELISA. All tests were performed a minimum of three times, and data are presented as the mean ± standard deviation. *P<0.05 vs. C3H10T1/2-MIGR1/MSC-treated group; #P<0.05 vs. model group. TT3, total triiodothyronine; TT4, total thyroxine; TSH, thyroid-stimulating hormone; Ab, antibody; TPO, thyroid peroxidase; TM, thyroid microsomal; TG, thyroglobulin; model, experimental autoimmune thyroiditis model; MSC, mesenchymal stem cell; ICAM, intercellular adhesion molecule.
C3H10T1/2-MIGR1-ICAM-1/MSCs attenuate thyroid follicle injury
Compared with the control group (Fig. 2A), injury of thyroid follicles was determined visually by their uneven size and the presence of epithelial hyperplasia and infiltration of inflammatory cells in the EAT model group (Fig. 2B) as revealed by H&E staining. In the C3H10T1/2-MIGR1-ICAM-1/MSC and MSC groups thyroid gland injury, epithelial hyperplasia and infiltration of inflammatory cells were attenuated;. however, in the other groups, no marked difference was observed following the administration of MSCs (Fig. 2C-F).
Figure 2.

Hematoxylin and eosin staining of thyroid tissue (magnification, ×100). (A) Normal control group; (B) experimental autoimmune thyroiditis model group; (C) MSC group; (D) C3H10T1/2/MSC group; (E) C3H10T1/2-MIGR1/MSC group; and (F) C3H10T1/2-MIGR1-intercellular adhesion molecule-1/MSC group. MSC, mesenchymal stem cell. The scale bar represents 100 µm.
C3H10T1/2-MIGR1-ICAM-1/MSCs decrease splenic index
The splenic index in the EAT model group was significantly higher than that of the normal control group (P<0.01). In the C3H10T1/2-MIGR1-ICAM-1/MSC group, the splenic index was signifantly decreased compared with the EAT model group (P<0.05). However, no significant improvement was observed in the splenic index in the other treatment groups (Fig. 3).
Figure 3.
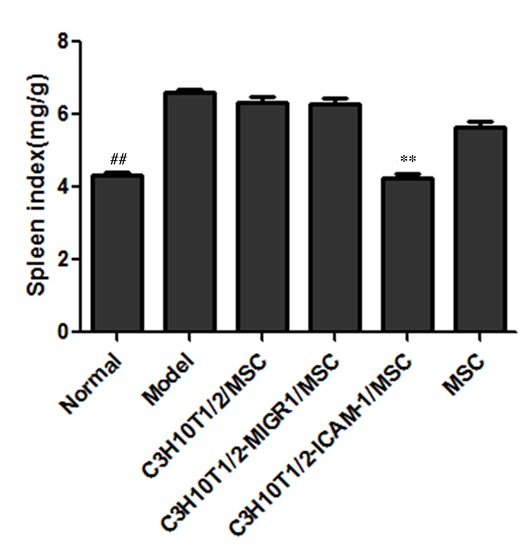
Splenic index in each group. C3H10T1/2-MIGR1-ICAM-1/MSC administration resulted in decreased splenic index compared with the C3H10T1/2-MIGR1/MSC group. Data are presented as the mean ± standard deviation. **P<0.01 vs. C3H10T1/2-MIGR1/MSC group; ##P<0.01 vs. model group. Model, experimental autoimmune thyroiditis model; MSC, mesenchymal stem cell; ICAM, intercellular adhesion molecule.
C3H10T1/2-MIGR1-ICAM-1/MSCs reverse the mRNA expression of IL-4, IL-10, IL-17 and INF-γ
RT-qPCR indicated that IL-4 and IL-10 mRNA expression was downregulated in the EAT model group compared with that of normal controls (P<0.05; Fig. 4A and B). However, this downregulation was reversed in the C3H10T1/2/MSC (IL-4, P<0.01), C3H10T1/2-MIGR1/MSC (IL-4, P<0.05; IL-10, P<0.01), C3H10T1/2-MIGR1-ICAM-1/MSC (IL-4 and IL-10, P<0.01) and MSC groups (IL-4, P<0.05; IL-10, P<0.01) compared with the model group. The mRNA expression of IL-17 and INF-γ was significantly increased in the EAT model group compared with normal controls (both P<0.01; Fig. 4C and D). This upregulation was significantly attenuated following treatment with MSCs (P<0.05). The upregulation of IL-4 and IL-10 mRNA and downregulation of IL-17 and INF-γ mRNA were most apparent in the C3H10T1/2-MIGR1-ICAM-1/MSC group compared with the other MSC groups.
Figure 4.

Expression of (A) IL-4, (B) IL-10, (C) IL-17 and (D) INF-γ mRNA. Data were presented as the mean ± standard deviation. *P<0.05 vs. model group; **P<0.01 vs. model group. IL, interleukin; INF, interferon; model, experimental autoimmune thyroiditis model; MSC, mesenchymal stem cell; ICAM, intercellular adhesion molecule.
C3H10T1/2-MIGR1-ICAM-1/MSCs are associated with the p38 and ERK signaling pathways in the mouse spleen
Western blot analysis revealed the expression of p-ERK and p-p38 was upregulated in the model group compared with the normal control group. Compared with the model group, the expression of p-ERK and p-p38 was decreased in the C3H10T1/2-MIGR1-ICAM-1/MSC group. No statistical difference was observed in the expression of p-ERK and p-p38 in the C3H10T1/2-MIGR1/MSC, C3H10T1/2/MSC and primary MSC groups compared with the model group (Figs. 5 and 6).
Figure 5.
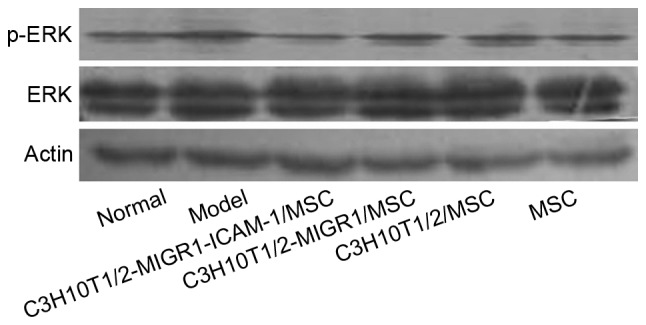
Phosphorylation of ERK in each group determined via western blot analysis. p, phosphorylated; ERK, extracellular signal-regulated kinase; model, experimental autoimmune thyroiditis model; MSC, mesenchymal stem cell; ICAM, intercellular adhesion molecule.
Figure 6.
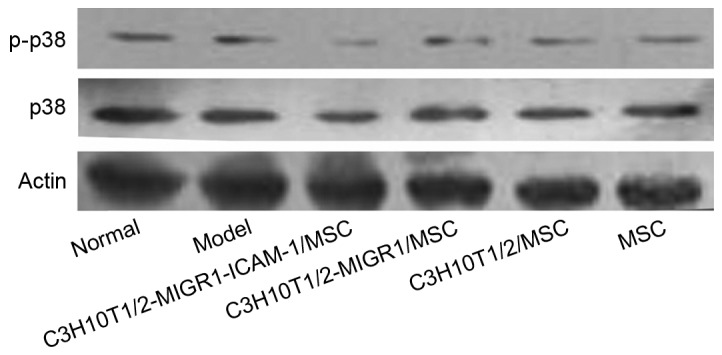
Phosphorylation of p38 in each group determined via western blot analysis. p, phosphorylated, p38, p38 mitogen-activated protein kinase; model, experimental autoimmune thyroiditis model; MSC, mesenchymal stem cell; ICAM, intercellular adhesion molecule.
ICAM-1-overexpression modulates the localization of MSCs in the thyroid gland and lung
Red fluorescence was observed in the thyroid gland (Fig. 7) and lung (Fig. 8), indicating the presence of CM-DiI-labeled MSCs. Localization in these tissues was markedly increased in the C3H10T1/2-MIGR1-ICAM-1/MSCs group in the thyroid gland compared with the other groups. The localization of primary MSCs in the lung was superior to that in the thyroid gland in all groups, which may be associated with the arrest of MSCs by the extensive vascular network in the lung.
Figure 7.

Nesting of CM-DiI-labeled MSCs in thyroid gland tissue observed via fluorescence microscopy (magnification, ×400). White arrows indicate the presence of red fluorescence and therefore MSCs. (A) C3H10T1/2/MSC group; (B) C3H10T1/2-MIGR1/MSC group; (C) C3H10T1/2-MIGR1-intercellular adhesion molecule-1/MSC group; (D) MSC group. The scale bar reperesents 250 µm. MSC, mesenchymal stem cells.
Figure 8.

Nesting of CM-DiI-labeled MSCs in lung tissue observed via fluorescence microscopy (magnification, ×400). White arrows indicate the presence of red fluorescence and therefore MSCs. (A) C3H10T1/2/MSC group; (B) C3H10T1/2-MIGR1/MSC group; (C) C3H10T1/2-MIGR1-intercellular adhesion molecule-1/MSC group; (D) MSC group. The scale bar reperesents 250 µm. MSC, mesenchymal stem cells.
Discussion
The onset of AIT is associated with viral infection and excessive uptake of iodides, which may result in autoimmune disorders (16), potentially leading to injury of the thyroid gland and infiltration of lymph nodes (17). At present, there is no specific treatment option that has been recommended for AIT. Recently, MSCs with the capacity to self-renew and differentiate in vivo, have been demonstrated to have immunomodulatory properties and contribute to the regeneration of cells in injured organs via cellular migration (18,19).
Previous studies have indicated that an imbalance of T cell subtypes serves an important role in the pathogenesis of AIT (20,21), as the imbalance may induce immune dysfunction, expose thyroid antigens and stimulate the generation of antigen-presenting cells. Furthermore, it may induce the formation of precursor cells in the presence of various cytokines, such as IL-10 and IL-4 (22). The present study demonstrated that C3H10T1/2-ICAM-1/MSCs may contribute to the expression of IL-4 and IL-10 in the immune cells, and that they may downregulate the expression of IL-17 and INF-γ. Among these factors, INF-γ and IL-17 are generated by Th1 cells, whereas IL-4 and IL-10 are generated by Th2 cells. Therefore, these findings suggest that C3H10T1/2-ICAM-1/MSCs were associated with the regulation of the immune system via recruiting cytokines and modulating the expression of T cell subtypes.
MAPKs are a highly conserved family of serine/threonine protein kinases involved in a variety of fundamental cellular processes (23). The major components of MAPKs are ERKs, c-Jun N-terminal kinases and p38, which have been reported to have crucial roles in the regulation of gene expression and cytoplasm function (24). MAPKs may also be associated with the proliferation, apoptosis, differentiation, necrosis and expression of cellular adhesion molecules (25). Previous studies have demonstrated that p38 signaling was activated in MSCs under the stimulation of inflammatory factors, which would subsequently trigger the overexpression of adhesion molecules, as well as MSC immune regulation (26,27).
In the present study, the expression of p-ERK and p-p38 was upregulated in the EAT model group, whereas in the C3H10T1/2-ICAM-1/MSC group, a marked increase was observed in the dephosphorylation of ERK and p38 compared with other groups, indicating that C3H10T1/2-ICAM-1/MSCs may modulate the ERK and p38 signaling pathways. These findings suggest that, under the stimulation of exogenous thyroglobulin, immunocytes were stimulated and the ERK and p38 signaling pathways were activated. Additionally, phosphorylation was increased, which further contributed to immunocyte activation. However, following the administration of C3H10T1/2-ICAM-1/MSCs, the activation of these pathways was reduced, and the expression of IL-4, IL-10, IL-17 and IFN-γ was modulated, which suggests that inflammatory factor-induced activation of the p38 signaling pathway may contribute to the expression of ICAM-1 in MSCs and immunocytes, and therefore inhibit the immune response. Furthermore, the level of TMAb was decreased and injury of the thyroid gland and inflammatory infiltration was attenuated.
Nesting of MSCs in the target organ is also of importance for the immunosuppressive effects in vivo (28). In the presence of inflammatory factors, MSCs are able to migrate to injury sites and are distributed in peripheral tissues. At the same time, MSCs may secrete factors involved in the formation of neovessels, and contribute to the regeneration of injury tissues (29). Yen et al (30) previously demonstrated that the expression of adhesion molecules in MSCs and the surface of vascular endothelial cells was upregulated in the presence of inflammatory factors, which increased cellular adhesion between MSCs and vascular endothelial cells and therefore contributed to the migration of MSCs to injury tissues. In the present study, to confirm the migration of MSCs in vivo, they were labeled with CM-DiI. The results indicated that a more notable MSC migration to the injured thyroid gland occurred in the ICAM-1-expressing cells.
In conclusion, the present findings suggest that C3H10T1/2-ICAM-1/MSCs were able to affect the differentiation, proliferation and migration of immunocytes by modulating the p38 and ERK signaling pathways, which suggests that ICAM-1 may affect the immunoregulatory effects of MSCs by modulating the migration of MSCs in vivo. These findings may provide rationale for the development of novel therapies for AIT using MSCs which express ICAM-1.
Acknowledgements
The present study was supported by the Tianjin Natural Science Fund (grant no. 12JCYBJC17100).
References
- 1.Chistiakov DA. Immunogenetics of Hashimoto's thyroiditis. J Autoimmune Dis. 2005;2:1. doi: 10.1186/1740-2557-2-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.de Vries L, Bulvik S, Phillip M. Chronic autoimmune thyroiditis in children and adolescents: At presentation and during long-term follow-up. Arch Dis Child. 2009;94:33–37. doi: 10.1136/adc.2007.134841. [DOI] [PubMed] [Google Scholar]
- 3.DeBoer MD, LaFranchi S. Differential presentation for children with autoimmune thyroiditis discovered because of symptomdevelopment or screening. J Pediatr Endocrinol Metab. 2008;21:753–761. doi: 10.1515/JPEM.2008.21.8.753. [DOI] [PubMed] [Google Scholar]
- 4.Lichiardopol C, Mota M. The thyroid and autoimmunity. Rom J Intern Med. 2009;47:207–215. [PubMed] [Google Scholar]
- 5.Heymann WR. Chronic urticaria and angioedema associated with thyroid autoimmunity: Review and therapeutic implications. J Am Acad Dermatol. 1999;40:229–232. doi: 10.1016/S0190-9622(99)70193-3. [DOI] [PubMed] [Google Scholar]
- 6.Cipriani P, Carubbi F, Liakouli V, Marrelli A, Perricone C, Perricone R, Alesse E, Giacomelli R. Stem cells in autoimmune diseases: Implications for pathogenesis and future trends in therapy. Autoimmun Rev. 2013;12:709–716. doi: 10.1016/j.autrev.2012.10.004. [DOI] [PubMed] [Google Scholar]
- 7.Oreffo RO, Cooper C, Mason C, Clements M. Mesenchymal stem cells: Lineage, plasticity, and skeletal therapeutic potential. Stem Cell Rev. 2005;1:169–178. doi: 10.1385/SCR:1:2:169. [DOI] [PubMed] [Google Scholar]
- 8.Ding DC, Shyu WC, Lin SZ. Mesenchymal stem cells. Cell Transplant. 2011;20:5–14. doi: 10.3727/096368910X. [DOI] [PubMed] [Google Scholar]
- 9.Liu S, Yuan M, Hou K, Zhang L, Zheng X, Zhao B, Sui X, Xu W, Lu S, Guo Q. Immune characterization of mesenchymal stem cells in human umbilical cord Wharton's jelly and derived cartilage cells. Cell Immunol. 2012;278:35–44. doi: 10.1016/j.cellimm.2012.06.010. [DOI] [PubMed] [Google Scholar]
- 10.Wada N, Gronthos S, Bartold PM. Immunomodulatory effects of stem cells. Periodontol 2000. 2013;63:198–216. doi: 10.1111/prd.12024. [DOI] [PubMed] [Google Scholar]
- 11.Buschmann K, Koch L, Braach N, Mueller H, Frommhold D, Poeschl J, Ruef P. CXCL1-triggered interaction of LFA1 and ICAM1 control glucose-induced leukocyte recruitment during inflammation in vivo. Mediators inflamm. 2012;2012:739176. doi: 10.1155/2012/739176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chen JD, Xu FF, Zhu H, Li XM, Tang B, Liu YL, Zhang Y. ICAM-1 regulates differentiation of MSC to adipocytes via activating MAPK pathway. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2014;22:160–165. doi: 10.7534/j.issn.1009-2137.2014.01.031. (In Chinese) [DOI] [PubMed] [Google Scholar]
- 13.Kong YC. Experimental autoimmune thyroiditis in the mouse. Curr Protoc Immunol. 2007;15:Unit 15.7. doi: 10.1002/0471142735.im1507s78. [DOI] [PubMed] [Google Scholar]
- 14.Prassopoulos P, Daskalogiannaki M, Raissaki M, Hatjidakis A, Gourtsoyiannis N. Determination of normal splenic volume on computed tomography in relation to age, gender and body habitus. Eru Radiol. 1997;7:246–248. doi: 10.1007/s003300050145. [DOI] [PubMed] [Google Scholar]
- 15.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-tie quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 16.Nagayama Y, Horie I, Saitoh O, Nakahara M, Abiru N. CD4+ CD25+ naturally occurring regulatory T cells and not lymphopenia play a role in the pathogenesis of iodide-induced autoimmune thyroiditis in NOD-H2 h4 mice. J Autoimmun. 2007;29:195–202. doi: 10.1016/j.jaut.2007.07.008. [DOI] [PubMed] [Google Scholar]
- 17.Armengol MP, Juan M, Lucas-Martín A, Fernández-Figueras MT, Jaraquemada D, Gallart T, Pujol-Borrell R. Thyroid autoimmune disease: Demonstration of thyroid antigen-specific B cells and recombination-activating gene expression in chemokine-containing active intrathyroidal germinal centers. Am J Pathol. 2001;159:861–873. doi: 10.1016/S0002-9440(10)61762-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Choi EW, Shin IS, Lee HW, Park SY, Park JH, Nam MH, Kim JS, Woo SK, Yoon EJ, Kang SK, et al. Transplantation of CTLA4Ig gene-transduced adipose tissue-derived mesenchymal stem cells reduces inflammatory immune response and improves Th1/Th2 balance in experimental autoimmune thyroiditis. J Gene Med. 2011;13:3–16. doi: 10.1002/jgm.1531. [DOI] [PubMed] [Google Scholar]
- 19.Kang SK, Shin IS, Ko MS, Jo JY, Ra JC. Journey of mesenchymal stem cells for homing: Strategies to enhance efficacy and safety of stem cell therapy. Stem Cells Int. 2012;2012:342968. doi: 10.1155/2012/342968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, Fehervari Z, Shimizu J, Takahashi T, Nomura T. Foxp3+ CD25+ CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune disease. Immunol rev. 2006;212:8–27. doi: 10.1111/j.0105-2896.2006.00427.x. [DOI] [PubMed] [Google Scholar]
- 21.Wing K, Sakaguchi S. Regulatory T cells exert checks and balances on self tolerance and autoimmunity. Nat Immunol. 2010;11:7–13. doi: 10.1038/ni.1818. [DOI] [PubMed] [Google Scholar]
- 22.Yudoh K, Matsuno H, Nakazawa F, Yonezawa T, Kimura T. Reduced expression of the regulatory CD4+ T cell subset is related to Th1/Th2 balance and disease severity in rheumatoid arthritis. Arthritis Rheum. 2000;43:617–627. doi: 10.1002/1529-0131(200003)43:3<617::AID-ANR19>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- 23.Arbabi S, Maier RV. Mitogen-activated protein kinases. Crit Care Med. 2002;30(Suppl 1):S74–S79. doi: 10.1097/00003246-200201001-00010. [DOI] [PubMed] [Google Scholar]
- 24.Schett G, Tohidast-Akrad M, Smolen JS, Schmid BJ, Steiner CW, Bitzan P, Zenz P, Redlich K, Xu Q, Steiner G. Activation, differential localization, and regulation of the stress-activated protein kinases, extracellular signal-regulated kinase, c-Jun N-terminal kinase and p38 mitogen-activated protein kinase, in synovial tissue and cells in rheumatoid arthritis. Arthritis Rheum. 2000;43:2501–2512. doi: 10.1002/1529-0131(200011)43:11<2501::AID-ANR18>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 25.Zhao L, Liu X, Liang J, Han S, Wang Y, Yin Y, Luo Y, Li J. Phosphorylation of p38 MAPK mediates hypoxic preconditioning-induced neuroprotection against cerebral ischemic injury via mitochondria translocation of Bcl-xL in mice. Brain Res. 2013;1503:78–88. doi: 10.1016/j.brainres.2013.01.051. [DOI] [PubMed] [Google Scholar]
- 26.Johnson GL, Lapadat R. Mitogen-activated protein kinase pathways mediated by ERK, JNK and p38 protein kinases. Science. 2002;298:1911–1912. doi: 10.1126/science.1072682. [DOI] [PubMed] [Google Scholar]
- 27.Ren G, Zhao X, Zhang L, Zhang J, L'Huillier A, Ling W, Roberts AI, Le AD, Shi S, Shao C, Shi Y. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J immunol. 2010;184:2321–2328. doi: 10.4049/jimmunol.0902023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Luz-Crawford P, Noël D, Fernandez X, Khoury M, Figueroa F, Carrión F, Jorgensen C, Djouad F. Mesenchymal stem cells repress Th17 molecular program through the PD-1 pathway. PLoS One. 2012;7:e45272. doi: 10.1371/journal.pone.0045272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Deng J, Zou ZM, Zhou TL, Su YP, Ai GP, Wang JP, Xu H, Dong SW. Bone marrow mesenchymal stem cells can be mobilized into peripheral blood by G-CSF in vivo and integrate into traumatically injured cerebral tissue. Neurolo Sci. 2011;32:641–651. doi: 10.1007/s10072-011-0608-2. [DOI] [PubMed] [Google Scholar]
- 30.Yen BL, Huang HI, Chien CC, Jui HY, Ko BS, Yao M, Shun CT, Yen ML, Lee MC, Chen YC. Isolation of multipotent cells from human term placenta. Stem Cells. 2005;23:3–9. doi: 10.1634/stemcells.2004-0098. [DOI] [PubMed] [Google Scholar]