

Abstract
It has been suggested that isoflurane may cause perioperative liver injury. However, the mechanism of its action remains unknown. The purpose of the present study was to determine this possible mechanism. Sprague-Dawley rats were randomly assigned into one of three groups (all n=12): Control group (exposed to mock anesthesia), isoflurane group (exposed to 2% isoflurane for 90 min), and isoflurane + insulin-like growth factor 1 (IGF-1) group (exposed to 2% isoflurane for 90 min and then treated with IGF-1). Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) and western blotting were conducted to determine the levels of expression of IGF-1 and its receptor IGF-R. Liver necrosis was assessed by histological examination. TUNEL assay was performed to determine the apoptosis of hepatic cells. In addition, the levels of the proteins caspase-3 and B-cell lymphoma-extra large (Bcl-xL) were measured. Compared with the control group, levels of IGF-1 and IGF-1R mRNA and protein were significantly decreased following exposure to isoflurane (all P<0.05). The necrosis rate and liver apoptosis were significantly increased in the group treated with isoflurane alone compared with the control group (P<0.05), but were significantly decreased compared with the isoflurane group following application of IGF-1 (P<0.05). Additionally, isoflurane exposure significantly increased levels of caspase-3 compared with the control group (P<0.05), but decreased levels of Bcl-xL (P<0.05). By contrast, application of IGF-1 reversed these changes. The present study therefore suggests that isoflurane induces liver injury in part by regulating the expression of IGF-1 and that application of IGF-1 may protect against liver injury induced by isoflurane exposure.
Keywords: isoflurane, liver injury, insulin-like growth factor 1, necrosis, apoptosis
Introduction
It has been a long time since the introduction of anesthetic drugs to the clinical application. However, exposure to anesthetic agents may result in adverse outcomes including cellular damage. The anesthetic drugs currently available induce various side effects (1). For example, it has been reported that ~20% patients who receive halothane may have experience nausea, lethargy, and fever side effects (2). The liver is heavily involved in drug metabolism and the majority of anesthetics are metabolized totally or partially, in the liver, explaining why anesthetic drugs induce liver injury (3). Isoflurane is a volatile anesthetic agent, which is widely administered to patients undergoing surgical procedures (4). Isoflurane undergoes minimal biotransformation; however, a wide spectrum of hepatotoxicity ranging from transaminitis to fulminant hepatic failure induced by isoflurane has been observed (5–8). It has been demonstrated that the oxidative stress response, and hepatic necrosis and apoptosis, are involved in the liver injury mechanism (9,10); however, the exact mechanisms underlying this action have not been elucidated.
A number of studies have investigated the role of insulin-like growth factor 1 (IGF-1) in liver disease (11–15). IGF-1 is synthesized in hepatocytes and multiple non-parenchymal cells, and serves an important role in the anabolic and metabolic actions involved in the growth and function of multiple tissues (13,16). Serum IGF-I levels have been regarded as a useful index of hepatocellular dysfunction. Serum IGF-I levels are decreased in chronic liver disease (17,18) and it has been observed that administering IGF-1 may improve liver function (19) and decrease oxidative damage and fibrosis (13). Microarray analysis has suggested that the level of IGF-1 is downregulated following exposure to isoflurane anesthesia (20). Therefore, it has been speculated that IGF-1 may be involved in the liver injury mechanism induced by exposure to isoflurane anesthesia.
To confirm this hypothesis, in the present study, rats were exposed to isoflurane for 90 min then levels of IGF-1 and its receptor (IGF-1R) were measured. Exogenous IGF-1 was subsequently administered to rats to investigate the effect of IGF-1 on liver function, as well as its possible mechanism of action. The results of the present study may provide evidence supporting the use of IGF-1 in the treatment of liver injury induced by anesthesia.
Materials and methods
Animals and grouping
A total of 36 male six-week-old Sprague-Dawley rats, weighing 200–250 g, were purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China) and allowed to acclimatize for one week prior to experiments. The rats were randomly assigned into one of three groups: A control group (n=12, exposed to mock anesthesia), an isoflurane group (n=12, exposed to 2% isoflurane for 90 min) and an isoflurane + IGF-1 group (n=12, exposed to 2% isoflurane for 90 min and then administered IGF-1 intervention). The rats were maintained under a standard 12/12-h light/dark cycle in temperature-controlled cages (22±5°C) with a relative humidity of 60±10% and received standard food pellets and distilled water ad libitum. All animals were treated in accordance with the standards of the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (Bethesda, MD, USA) and all protocols were approved by the Ethics Committee on Animals at Sun Yat-Sen University (Zhuhai, China).
Isoflurane exposure and tissue collection
The rats in the isoflurane groups were placed in an induction chamber and received 2% isoflurane (Forene; Abbott, Queenborough, Kent, UK)/98% medical grade air (vol/vol) from an anesthesia machine (Fabius CE, Dräger Medical, Lübeck, Germany) for 90 min. Rats in the control group received air/oxygen at identical flow rates. The concentration of isoflurane was monitored continuously with a Datex Capnomac gas-analyzer (Datex-Ohmeda, Helsinki, Finland) throughout anesthesia. The rectal temperature was maintained at 37.0±0.5°C by a heating lamp and pulse oximeter oxygen saturation (SpO2) was routinely monitored. The rats in the isoflurane + IGF-1 group received anesthetic exposure, then intravenous injections of IGF-1 (20 µg/kg; Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) at different amounts of time (0 and 72 h and 7 days). Thereafter, all animals were sacrificed by decapitation after administration of IGF-1 at 0 and 72 h and 7 days and liver tissues were collected for further analyses.
Histological assessment of liver necrosis
The extent of liver necrosis was determined in 10% neutral buffered formaldehyde-fixed (4°C for 6 h) paraffin-embedded liver sections stained with hematoxylin and eosin, according to a previously described method (21). Briefly, sections were harvested at different times following the application of IGF-1 (0 h, 72 h and 7 days). Sections were deparaffinzed, hydrated, stained in alum hematoxylin, differentiated with acid alcohol, washed with tap water, stained with eosin and dehydrated. Sections were then observed under a light microscope (BX51; Olympus Corp., Tokyo, Japan) by a pathologist who was blinded to the groups. Ten high-power fields were randomly collected. The area of necrosis was quantified using ImageJ software version 1.45 (National Institutes of Health). The necrosis rate was calculated as the area of necrosis divided by the total area.
TUNEL assay
A TUNEL assay was performed to determine the apoptosis of hepatic cells in rat livers using a commercially available TUNEL kit (APO-DIRECT™ kit; BD Biosciences, Franklin Lakes, NJ, USA) according to the manufacturer's instructions. Briefly, sections were fixed with 4% paraformaldehyde for 10 min at room temperature, washed with phosphate-buffered saline (PBS) twice, maintained with 0.1% Triton X-100 in PBS and then incubated with H2O2 for 10 min in methanol at room temperature. Sections were then washed with PBS, followed by incubation in 50 µl biotin labeling solution at 37°C for 1 h in a humidified chamber. Following washing in PBS, sections were incubated with labeling termination solution for 5 min at room temperature and washed again. Streptavidin-horseradish peroxidase working solution (50 µl; Novolink Polymer; Leica Microsystems, Ltd., Milton Keynes, UK) was added to the sections at room temperature for 30 min. Subsequently, sections were incubated in diaminobenzidine working solution (Novolink Polymer; Leica Microsystems, Inc.) at room temperature, washed twice with PBS and photographed using a fluorescence microscope (BX50/BX-FLA/DP70; Olympus Corp.).
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Levels of IGF-1 and IGF-1R mRNA were measured using standard RT-qPCR techniques. RT-qPCR was performed at least three times. Total RNA was derived from liver tissues using TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA according to the manufacturer's instructions. First-strand cDNA was synthesized with the EasyScript® First-Strand cDNA Synthesis SuperMix (TransGen Biotech, Inc., Beijing, China). Amplification reactions were determined using the GeneAmp PCR System 2400 thermal cycler (Applied Biosystems; Thermo Fisher Scientific, Inc.). qPCR was performed using a QuantiTect SYBR Green RT-PCR kit (Qiagen SA, Courtaboeuf, France). Primer sequences were as follows: IGF-1, forward 5′-GTTCCGATGTTTTGCAGGTT-3′ and reverse 5′-TCTGAGGAGGCTGGAGATGT-3′; IGF-1R, forward 5′-ACTATGCCGGTGTCTGTGTG-3′ and reverse 5′-TGCAAGTTCTGGTTGTCGAG-3′; and GAPDH, forward 5′-TCCTGCACCACCAACTGCTTAG-3′ and reverse 5′-AGTGGCAGTGATGGCATGGACT-3′. PCR conditions were as follows: Initial denaturation was performed at 95°C for 15 sec, followed by 30 cycles of denaturation for 30 sec at 95°C, annealing at 61°C for 5 sec, extension at 72°C for 15 sec, and subsequent final extension was performed at 72°C for 10 min. PCR products were run on an 1.2% agarose gel. The GAPDH gene was used as a reference gene. Fold changes relative to GAPDH were calculated using the 2−ΔΔCq method (22).
Western blot analysis
Tissue was harvested, washed with PBS and homogenized in 10 mM Tris buffer (pH 7.5). Total tissues homogenates were sonicated then centrifuged at 1,000 × g for 20 min at 4°C to extract protein. Protein concentration was assessed using a bicinchoninic acid protein assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Protein samples (20 µg per lane) were resolved with 10% SDS-PAGE. The separated proteins were transferred onto a polyvinylidene fluoride ultrafiltration membrane and the membrane was washed twice with Tris-buffered saline with Tween (TBST). The membranes were immersed in 5% fresh nonfat dry milk in TBST for 2 h at room temperature, washed with TBST and incubated with following polyclonal primary antibodies overnight at 4°C: Anti-IGF-1 antibody (sc-14221; 1:1,000), anti-IGF-1R antibody (sc-80985; 1:1,000), anti-caspase-3 antibody (sc-271759; 1:1,000), anti-B-cell lymphoma-extralarge (Bcl-xL) antibody (sc-136132; 1:1,000) or anti-GAPDH (sc-293335; 1:1,000). All antibodies were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). Following washing with TBST, the membrane was incubated with horseradish peroxidase-labeled goat anti-mouse secondary antibody (ZDR-5307; 1;5,000) (Beijing Zhongshan Golden Bridge Biotechnology, Co., Ltd., Beijing, China) for 2 h at room temperature. Finally, bands were visualized using an enhanced chemiluminescence western blotting substrate (Pierce; Thermo Fisher Scientific, Inc.), exposed to an X-ray film (Midwest Scientific, St. Louis, MO, USA), followed by densitometric analysis.
Statistical analysis
Data are represented as the mean ± standard deviation. The differences were compared using Student's t-test (for two groups) or one-way analysis of variance (for three or more groups). All statistical analyses were performed using SPSS software version 16.0 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to indicate a statistically significant difference.
Results
Expression of IGF-1 and IGF-1R following exposure to isoflurane
To evaluate the role of IGF-1 in the liver injury induced by isoflurane, levels of IGF-1 and IGF-1R mRNA and protein were measured following isoflurane exposure for 90 min. The results indicated that compared with the control group, levels of IGF-1 and IGF-1R mRNA (Fig. 1) and protein (Fig. 2) were significantly decreased following isoflurane exposure (all P<0.05), indicating that isoflurane significantly inhibited the expression of IGF-1 and IGF-1R.
Figure 1.

Levels of IGF-1 and IGF-1R mRNA following isoflurane exposure. mRNA levels of (A) IGF-1 and (B) IGF-1R following in control and following isoflurane exposure. *P<0.05 vs. control group. IGF-1, insulin-like growth factor 1; IGF-1R, insulin-like growth factor 1 receptor.
Figure 2.

Levels of IGF-1 and IGF-1R protein following isoflurane exposure. Protein levels of (A) IGF-1 and (B) IGF-1R in control and following isoflurane exposure. (C) Representative western blot images. *P<0.05 vs. control group. IGF-1, insulin-like growth factor 1; IGF-1R, insulin-like growth factor 1 receptor.
Liver necrosis rates following application of IGF-1
To assess the protective function of IGF-1 on the liver, IGF-1 was applied following exposure to isoflurane. At 0, 72 h and 7 days, the liver tissues were harvested and the necrosis rate was determined. At 7 days, the necrosis rate was significantly increased in the isoflurane group compared with the control group (P<0.05); however, compared with the isoflurane group, the necrosis rate was significantly decreased following the application of IGF-1 (P<0.05; Fig. 3). The results suggest that IGF-1 application may reduce tissue necrosis to some extent.
Figure 3.
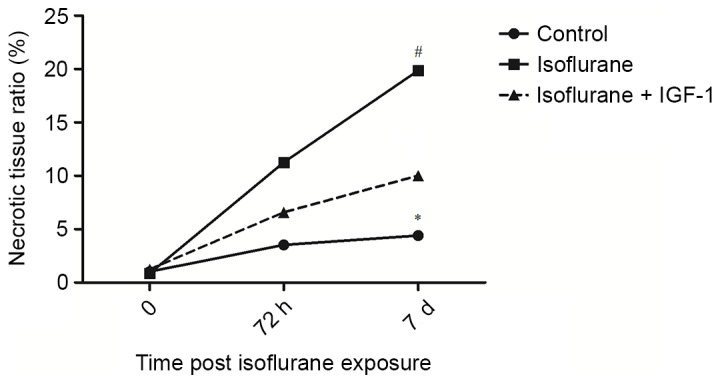
Liver necrosis rates following exposure to isoflurane. *P<0.05 vs. control group; #P<0.05 vs. isoflurane group. IGF-1, insulin-like growth factor 1.
Liver apoptosis rates following application of IGF-1
Subsequently, the effect of IGF-1 application on liver apoptosis was evaluated. As presented in Fig. 4, the number of apoptotic cells detected at day 7 were significantly higher following isoflurane exposure than those in the control group (P<0.05). However, compared with the isoflurane group, the number of apoptotic cells was reduced in the group receiving IGF-1 (P<0.05), demonstrating that IGF-1 application may effectively decrease the rate of apoptosis in the liver.
Figure 4.
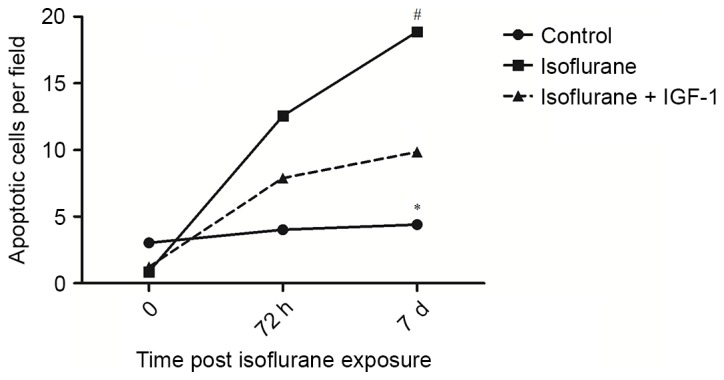
Liver apoptosis rates following exposure to isoflurane. *P<0.05 compared with the control group; #P<0.05 vs. isoflurane group. IGF-1, insulin-like growth factor 1.
Expression of caspase-3 and Bcl-xL following application of IGF-1
The mechanism of apoptosis was further investigated. The expression levels of the apoptosis-related proteins caspase-3 and Bcl-xL were determined. It was demonstrated that isoflurane significantly increased levels of caspase-3 compared with the control group (P<0.05; Fig. 5A), but decreased levels of Bcl-xL (P<0.05; Fig. 5B). However, application of IGF-1 significantly reduced levels of caspase-3 compared with the isoflurane group (P<0.05), but elevated levels of Bcl-xL (P<0.05; Fig. 5A and B). These results indicate that isoflurane induces apoptosis by elevating the expression of pro-apoptotic proteins (caspase-3), and reducing the expression of anti-apoptotic proteins (Bcl-xL). By contrast, IGF-1 treatment appeared to inhibit apoptosis by decreasing the expression of the pro-apoptotic protein caspase-3, and increasing the expression of the anti-apoptotic protein Bcl-xL (Fig. 5).
Figure 5.

Expression of caspase-3 and Bcl-xL following application of IGF-1. Relative protein expression of (A) caspase-3 and (B) Bcl-xL. (C) Representative western blot image. *P<0.05 vs. control group; #P<0.05 vs. isoflurane group. IGF-1, insulin-like growth factor 1; Bcl-xL, B-cell lymphoma-extra large.
Discussion
In the present study, it was demonstrated that exposure to isoflurane may induce liver injury by stimulating liver necrosis and apoptosis, and significantly decreasing levels of IGF-1 and IGF-1R mRNA and protein. Application of IGF-1 following exposure to isoflurane alleviates liver injury by decreasing the rates of liver necrosis and apoptosis. The results of the present study indicate that isoflurane induces liver injury by regulating the expression of IGF-1 and application of IGF-1 may protect against the liver injury induced by isoflurane exposure.
Isoflurane is a halogenated volatile anesthetic, which has generally been considered to be a non-hepatotoxic alternative to halothane (23); nonetheless, an increasing number of cases of hepatic dysfunction following the administration of isoflurane have been reported (24,25). Two distinctive types of hepatotoxicity, mild injury and fulminant hepatotoxicity, have been observed following exposure to halogenated anesthetic agents. Mild injury is associated with mild increases in the levels of aminotransferase enzymes, whereas fulminant hepatotoxicity is characterized by marked increases in levels of liver enzymes and bilirubin and results in mass hepatic necrosis (26). In addition, apoptosis (programmed cell death) has been regarded as a primary mechanism of liver injury that occurs following anesthetic administration (3,27) and may result in mass apoptosis (28). Edmands et al (20) found that exposure to isoflurane anesthesia downregulated IGF-1 levels, indicating that there may be a possible association between isoflurane anesthesia and IGF-1. It has been suggested that IGF-1 and its receptor IGF-1R serve important roles in the liver (29). Decreased serum levels of IGF-1 are associated with chronic liver disease and the levels of circulating IGF-1 may be correlated with the degree of hepatocellular dysfunction (30). The effects of the IGF-1 are mediated through the IGF-IR; however, IGF-1 has little direct effect on hepatocyte function as low levels of IGF-1R are expressed by hepatocytes in normal conditions (31). However, overexpression of IGF-IR has been detected in chronic hepatitis C (32), chronic hepatitis B and liver cirrhosis (33). A number of potential underlying mechanisms may be responsible for the effects of IGF-1 on the liver. Insulin resistance (34), oxidative stress and mitochondrial dysfunction may be responsible for the development of liver diseases (35). It has been demonstrated that IGF-1 improves insulin sensitivity (36), exerts strong anabolic action (29), improves the nutritional condition (37), improves mitochondrial dysfunction, oxidative stress and apoptosis (38), and reverses mitochondrial morphology in the hepatocytes (39). Furthermore, it has been demonstrated in a model of cirrhosis that administration of IGF-1 improves liver dysfunction and fibrosis (40). Therefore, the present study speculated that the liver injury induced by isoflurane anesthesia may occur by regulating IGF-1 expression.
To confirm the hypothesis, levels of IGF-1 following exposure to isoflurane were determined. In accordance with the results from Edmands et al (20), the results of the present study demonstrated that levels of IGF-1 mRNA and protein were significantly decreased following exposure to isoflurane. Moreover, levels of IGF-1R were down-regulated, indicating that the protective effects of IGF-1 on the liver were inhibited by isoflurane. IGF-1 was subsequently administered to rats following exposure to isoflurane. As expected, liver necrosis and apoptosis rates were significantly reduced after administration of IGF-1. The related apoptosis mechanism was then evaluated by assessing levels of caspase-3 and Bcl-xL protein. caspase-3, one of the most important pro-apoptotic proteins, is activated and eventually stimulates apoptosis by inducing DNA fragmentation and plasma membrane blebbing (41). Bcl-xL protein, a member of the Bcl-2 family, promotes cell survival and prevents homo-oligomerization of pro-apoptotic family members (42,43). Intrinsic apoptosis may be stimulated by a number of factors including DNA damage and cytoplasm detachment, and result in the accumulation of pro-apoptotic members of the Bcl-2 family in the mitochondrial membrane (44,45). This may lead to an increase in mitochondrial permeability, and the displacement of cytochrome c and certain pro-apoptotic proteins from the mitochondria to cytoplasm, resulting in the subsequent stimulation of caspase-9 (46) and caspase-3 (47). In the present study, it was demonstrated that levels of caspase-3 were significantly increased by isoflurane, whereas levels of Bcl-xL were significantly decreased, indicating that apoptosis was induced by isoflurane. However, application of IGF-1 significantly downregulated levels of caspase-3 but elevated Bcl-xL levels, suggesting that apoptosis was prevented by IGF-1.
In conclusion, the results of the present study suggest that isoflurane induces liver injury by regulating the expression of IGF-1. Application of IGF-1 may therefore protect against liver injury induced by isoflurane exposure.
References
- 1.Njoku D, Laster MJ, Gong DH, Eger EI, II, Reed GF, Martin JL. Biotransformation of halothane, enflurane, isoflurane, and desflurane to trifluoroacetylated liver proteins: Association between protein acylation and hepatic injury. Anesth Analg. 1997;84:173–178. doi: 10.1097/00000539-199701000-00031. [DOI] [PubMed] [Google Scholar]
- 2.Wright R, Eade OE, Chisholm M, Hawksley M, Lloyd B, Moles TM, Edwards JC, GArdner MJ. Controlled prospective study of the effect on liver function of multiple exposures to halothane. Lancet. 1975;1:817–820. doi: 10.1016/S0140-6736(75)93000-7. [DOI] [PubMed] [Google Scholar]
- 3.Suliburk JW, Gonzalez EA, Kennison SD, Helmer KS, Mercer DW. Differential effects of anesthetics on endotoxin-induced liver injury. J Trauma. 2005;58:711–717. doi: 10.1097/01.TA.0000159245.60495.00. [DOI] [PubMed] [Google Scholar]
- 4.Mohseni M, Safari S, Alavian SM. Volatile anesthetics in ischemic liver injury: Enemy or friend? Hepat Mon. 2014;14:e19880. doi: 10.5812/hepatmon.19880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gunza JT, Pashayan AG. Postoperative elevation of serum transaminases following isoflurane anesthesia. J Clin Anesth. 1992;4:336–341. doi: 10.1016/0952-8180(92)90143-O. [DOI] [PubMed] [Google Scholar]
- 6.Brunt EM, White H, Marsh JW, Holtmann B, Peters MG. Fulminant hepatic failure after repeated exposure to isoflurane anesthesia: A case report. Hepatology. 1991;13:1017–1021. doi: 10.1002/hep.1840130602. [DOI] [PubMed] [Google Scholar]
- 7.Turner GB, O'Rourke D, Scott GO, Beringer TR. Fatal hepatotoxicity after re-exposure to isoflurane: A case report and review of the literature. Eur J Gastroenterol Hepatol. 2000;12:955–959. doi: 10.1097/00042737-200012080-00017. [DOI] [PubMed] [Google Scholar]
- 8.Carrigan TW, Straughen WJ. A report of hepatic necrosis and death following isoflurane anesthesia. Anesthesiology. 1987;67:581–583. doi: 10.1097/00000542-198710000-00026. [DOI] [PubMed] [Google Scholar]
- 9.Dabbagh A, Rajaei S. The role of anesthetic drugs in liver apoptosis. Hepat Mon. 2013;13:e13162. doi: 10.5812/hepatmon.13162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Gelven PL, Cina SJ, Lee JD, Nichols CA. Massive hepatic necrosis and death following repeated isoflurane exposure: Case report and review of the literature. Am J Forensic Med Pathol. 1996;17:61–64. doi: 10.1097/00000433-199603000-00011. [DOI] [PubMed] [Google Scholar]
- 11.Kaseb AO, Morris JS, Hassan MM, Siddiqui AM, Lin E, Xiao L, Abdalla EK, Vauthey JN, Aloia TA, Krishnan S, Abbruzzese JL. Clinical and prognostic implications of plasma insulin-like growth factor-1 and vascular endothelial growth factor in patients with hepatocellular carcinoma. J Clin Oncol. 2011;29:3892–3899. doi: 10.1200/JCO.2011.36.0636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sanz S, Pucilowska JB, Liu S, Rodríguez-Ortigosa CM, Lund PK, Brenner DA, Fuller CR, Simmons JG, Pardo A, Martínez-Chantar ML, et al. Expression of insulin-like growth factor I by activated hepatic stellate cells reduces fibrogenesis and enhances regeneration after liver injury. Gut. 2005;54:134–141. doi: 10.1136/gut.2003.024505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Svegliati-Baroni G, Ridolfi F, Di Sario A, Casini A, Marucci L, Gaggiotti G, Orlandoni P, Macarri G, Perego L, Benedetti A, Folli F. Insulin and insulin-like growth factor-1 stimulate proliferation and type I collagen accumulation by human hepatic stellate cells: Differential effects on signal transduction pathways. Hepatology. 1999;29:1743–1751. doi: 10.1002/hep.510290632. [DOI] [PubMed] [Google Scholar]
- 14.Aksu I, Baykara B, Kiray M, Gurpinar T, Sisman AR, Ekerbicer N, Tas A, Gokdemir-Yazar O, Uysal N. Serum IGF-1 levels correlate negatively to liver damage in diabetic rats. Biotech Histochem. 2013;88:194–201. doi: 10.3109/10520295.2012.758311. [DOI] [PubMed] [Google Scholar]
- 15.Rehem RN, El-Shikh WM. Serum IGF-1, IGF-2 and IGFBP-3 as parameters in the assessment of liver dysfunction in patients with hepatic cirrhosis and in the diagnosis of hepatocellular carcinoma. Hepatogastroenterology. 2011;58:949–954. [PubMed] [Google Scholar]
- 16.Clemmons DR. Metabolic actions of insulin-like growth factor-I in normal physiology and diabetes. Endocrinol Metab Clin North Am. 2012;41:425-443, vii-viii. doi: 10.1016/j.ecl.2012.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Friedman SL. Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J Biol Chem. 2000;275:2247–2250. doi: 10.1074/jbc.275.4.2247. [DOI] [PubMed] [Google Scholar]
- 18.Pinzani M, Abboud HE, Aron DC. Secretion of insulin-like growth factor-I and binding proteins by rat liver fat-storing cells: Regulatory role of platelet-derived growth factor. Endocrinology. 1990;127:2343–2349. doi: 10.1210/endo-127-5-2343. [DOI] [PubMed] [Google Scholar]
- 19.Picardi A, de Oliveira AC, Muguerza B, Tosar A, Quiroga J, Castilla-Cortázar I, Santidrián S, Prieto J. Low doses of insulin-like growth factor-I improve nitrogen retention and food efficiency in rats with early cirrhosis. J Hepatol. 1997;26:191–202. doi: 10.1016/S0168-8278(97)80026-8. [DOI] [PubMed] [Google Scholar]
- 20.Edmands SD, Ladow E, Hall AC. Microarray analyses of genes regulated by isoflurane anesthesia in vivo: A novel approach to identifying potential preconditioning mechanisms. Anesth Analg. 2013;116:589–595. doi: 10.1213/ANE.0b013e31827b27b0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vaquero J, Bélanger M, James L, Herrero R, Desjardins P, Côté J, Blei AT, Butterworth RF. Mild hypothermia attenuates liver injury and improves survival in mice with acetaminophen toxicity. Gastroenterology. 2007;132:372–383. doi: 10.1053/j.gastro.2006.11.025. [DOI] [PubMed] [Google Scholar]
- 22.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 23.Ozier Y, Klinck JR. Anesthetic management of hepatic transplantation. Curr Opin Anaesthesiol. 2008;21:391–400. doi: 10.1097/ACO.0b013e3282ff85f4. [DOI] [PubMed] [Google Scholar]
- 24.Stoelting RK. Isoflurane and postoperative hepatic dysfunction. Can J Anaesth. 1987;34:223–226. doi: 10.1007/BF03015155. (In English, French) [DOI] [PubMed] [Google Scholar]
- 25.Sinha A, Clatch RJ, Stuck G, Blumenthal SA, Patel SA. Isoflurane hepatotoxicity: A case report and review of the literature. Am J Gastroenterol. 1996;91:2406–2409. [PubMed] [Google Scholar]
- 26.Weitz J, Kienle P, Böhrer H, Hofmann W, Theilmann L, Otto G. Fatal hepatic necrosis after isoflurane anaesthesia. Anaesthesia. 1997;52:892–895. doi: 10.1111/j.1365-2044.1997.187-az0319.x. [DOI] [PubMed] [Google Scholar]
- 27.Cheng L, You Q, Yin H, Holt M, Franklin C, Ju C. Effect of polyI: C cotreatment on halothane-induced liver injury in mice. Hepatology. 2009;49:215–226. doi: 10.1002/hep.22585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jaeschke H, Gujral JS, Bajt ML. Apoptosis and necrosis in liver disease. Liver Int. 2004;24:85–89. doi: 10.1111/j.1478-3231.2004.0906.x. [DOI] [PubMed] [Google Scholar]
- 29.Takahashi Y. Essential roles of growth hormone (GH) and insulin-like growth factor-I (IGF-I) in the liver. Endocr J. 2012;59:955–962. doi: 10.1507/endocrj.EJ12-0322. [DOI] [PubMed] [Google Scholar]
- 30.Bonefeld K, Møller S. Insulin-like growth factor-I and the liver. Liver Int. 2011;31:911–919. doi: 10.1111/j.1478-3231.2010.02428.x. [DOI] [PubMed] [Google Scholar]
- 31.Caro JF, Poulos J, Ittoop O, Pories WJ, Flickinger EG, Sinha MK. Insulin-like growth factor I binding in hepatocytes from human liver, human hepatoma, and normal, regenerating, and fetal rat liver. J Clin Invest. 1988;81:976–981. doi: 10.1172/JCI113451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Stefano JT, Correa-Giannella ML, Ribeiro CM, Alves VA, Massarollo PC, Machado MC, Giannella-Neto D. Increased hepatic expression of insulin-like growth factor-I receptor in chronic hepatitis C. World J Gastroenterol. 2006;12:3821–3828. doi: 10.3748/wjg.v12.i24.3821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kim SO, Park JG, Lee YI. Increased expression of the insulin-like growth factor I (IGF-I) receptor gene in hepatocellular carcinoma cell lines: Implications of IGF-I receptor gene activation by hepatitis B virus X gene product. Cancer Res. 1996;56:3831–3836. [PubMed] [Google Scholar]
- 34.Bugianesi E, McCullough AJ, Marchesini G. Insulin resistance: A metabolic pathway to chronic liver disease. Hepatology. 2005;42:987–1000. doi: 10.1002/hep.20920. [DOI] [PubMed] [Google Scholar]
- 35.Mantena SK, King AL, Andringa KK, Eccleston HB, Bailey SM. Mitochondrial dysfunction and oxidative stress in the pathogenesis of alcohol- and obesity-induced fatty liver diseases. Free Radic Biol Med. 2008;44:1259–1272. doi: 10.1016/j.freeradbiomed.2007.12.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yakar S, Liu JL, Fernandez AM, Wu Y, Schally AV, Frystyk J, Chernausek SD, Mejia W, Le Roith D. Liver-specific igf-1 gene deletion leads to muscle insulin insensitivity. Diabetes. 2001;50:1110–1118. doi: 10.2337/diabetes.50.5.1110. [DOI] [PubMed] [Google Scholar]
- 37.Conchillo M, de Knegt RJ, Payeras M, Quiroga J, Sangro B, Herrero JI, Castilla-Cortazar I, Frystyk J, Flyvbjerg A, Yoshizawa C, et al. Insulin-like growth factor I (IGF-I) replacement therapy increases albumin concentration in liver cirrhosis: Results of a pilot randomized controlled clinical trial. J Hepatol. 2005;43:630–636. doi: 10.1016/j.jhep.2005.03.025. [DOI] [PubMed] [Google Scholar]
- 38.Hao CN, Geng YJ, Li F, Yang T, Su DF, Duan JL, Li Y. Insulin-like growth factor-1 receptor activation prevents hydrogen peroxide-induced oxidative stress, mitochondrial dysfunction and apoptosis. Apoptosis. 2011;16:1118–1127. doi: 10.1007/s10495-011-0634-9. [DOI] [PubMed] [Google Scholar]
- 39.Nishizawa H, Takahashi M, Fukuoka H, Iguchi G, Kitazawa R, Takahashi Y. GH-independent IGF-I action is essential to prevent the development of nonalcoholic steatohepatitis in a GH-deficient rat model. Biochem Biophys Res Commun. 2012;423:295–300. doi: 10.1016/j.bbrc.2012.05.115. [DOI] [PubMed] [Google Scholar]
- 40.Pérez R, Garcia-Fernández M, Díaz-Sánchez M, Puche JE, Delgado G, Conchillo M, Muntané J and Castilla-Cortázar I. Mitochondrial protection by low doses of insulin-like growth factor-I in experimental cirrhosis. World J Gastroenterol. 2008;14:2731–2739. doi: 10.3748/wjg.14.2731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Canbay A, Feldstein AE, Higuchi H, Werneburg N, Grambihler A, Bronk SF, Gores GJ. Kupffer cell engulfment of apoptotic bodies stimulates death ligand and cytokine expression. Hepatology. 2003;38:1188–1198. doi: 10.1053/jhep.2003.50472. [DOI] [PubMed] [Google Scholar]
- 42.Sattler M, Liang H, Nettesheim D, Meadows RP, Harlan JE, Eberstadt M, Yoon HS, Shuker SB, Chang BS, Minn AJ, et al. Structure of Bcl-xL-Bak peptide complex: Recognition between regulators of apoptosis. Science. 1997;275:983–986. doi: 10.1126/science.275.5302.983. [DOI] [PubMed] [Google Scholar]
- 43.Ku B, Liang C, Jung JU, Oh BH. Evidence that inhibition of BAX activation by BCL-2 involves its tight and preferential interaction with the BH3 domain of BAX. Cell Res. 2011;21:627–641. doi: 10.1038/cr.2010.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gottschalk S, Zwingmann C, Raymond VA, Hohnholt MC, Chan TS, Bilodeau M. Hepatocellular apoptosis in mice is associated with early upregulation of mitochondrial glucose metabolism. Apoptosis. 2012;17:143–153. doi: 10.1007/s10495-011-0669-y. [DOI] [PubMed] [Google Scholar]
- 45.Dejean LM, Ryu SY, Martinez-Caballero S, Teijido O, Peixoto PM, Kinnally KW. MAC and Bcl-2 family proteins conspire in a deadly plot. Biochim Biophys Acta. 2010;1797:1231–1238. doi: 10.1016/j.bbabio.2010.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Stevens MF, Werdehausen R, Gaza N, Hermanns H, Kremer D, Bauer I, Küry P, Hollmann MW, Braun S. Midazolam activates the intrinsic pathway of apoptosis independent of benzodiazepine and death receptor signaling. Reg Anesth Pain Med. 2011;36:343–349. doi: 10.1097/AAP.0b013e318217a6c7. [DOI] [PubMed] [Google Scholar]
- 47.Porter AG, Jänicke RU. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999;6:99–104. doi: 10.1038/sj.cdd.4400476. [DOI] [PubMed] [Google Scholar]