

Abstract
Hydrogen sulfide (H2S) exerts synergistic effects with another gaseous signaling molecule nitric oxide (NO) on ion channels and vasculature. However, the mechanism of the synergy is not well understood. Here, we show that the interaction between H2S and NO generates polysulfides (H2Sn), which activate transient receptor potential ankyrin 1 (TRPA1) channels. High performance liquid chromatography with tandem mass spectrometry analysis, along with the imaging of intracellular Ca2+ and H2Sn, showed that H2Sn and their effects were abolished by cyanolysis and by reducing substances such as dithiothreitol (DTT), cysteine, and glutathione (GSH). However, the effects of nitroxyl or nitrosopersulfide, other potential products of H2S and NO interaction, are not affected by cyanolysis or reducing substances. This study demonstrates that H2Sn are products of synergy between H2S and NO and provides a new insight into the signaling mechanisms.
Hydrogen sulfide (H2S) has various physiological roles: neuromodulation, vascular tone regulation, cytoprotection against oxidative stress or ischemia reperfusion injury, and anti-inflammation1,2,3,4,5,6. Cross talk between H2S and another signaling molecule, nitric oxide (NO), was initially reported as a synergistic effect of relaxation on vascular smooth muscle2. A similar synergistic effect of both molecules was observed in the twitch responses of the ileum7.
Two mechanisms of H2S and NO interaction were proposed for angiogenesis: the effect of H2S is mediated by NO through the activation of endothelial NO synthetase (eNOS) in one mechanism8, while the cooperative action between H2S and NO is essential in another9. Increase in H2S production and expression of an H2S-producing enzyme, cystathionine γ–lyase (CSE), by NO were also reported3. H2S enhances the activity of eNOS by facilitating phosphorylation of an active site and de-phosphorylation of the inhibitory site to increase the production of NO, leading to the attenuation of sudden cardiac arrest-induced mitochondrial injury, as well as protection of the heart from ischemia-reperfusion injury10,11.
It has been proposed that molecules generated by the chemical interaction of H2S and NO show a greater activity than the parental molecules or play a role as their carrier12,13,14,15. Nitroxyl (HNO) generated by the interaction of H2S and NO was proposed to activate transient receptor potential ankyrin 1 (TRPA1) channels14. However, nitrosopersulfide (SSNO) was reported to be mainly generated from H2S and NO interaction to act as a NO carrier, releasing NO to relax vascular smooth muscles15. Although H2Sn were detected as additional common products in both studies, they have not been studied in detail14,15,16. The application of H2S- or NO-donor alone can produce H2Sn probably due to the interaction with endogenous H2S or NO in mast cells17.
Recently, we found that H2S3 and H2S2 are produced by 3-mercaptopyruvate sulfurtransferase (3MST), an H2S-producing enzyme, from 3-mercaptopyruvate (3MP), as well as by the oxidation of H2S18,19,20. In addition, H2Sn activate TRPA1 channels by sulfurating two cysteine residues at the amino-termini of the channels21,22,23,24. Various other effects of H2Sn were subsequently reported. H2Sn facilitates the translocation of nuclear factor-like 2 (Nrf2) to the nucleus by modifying its binding partner, kelch-like ECH-associated protein 1 (Keap1), to up-regulate the transcription of antioxidant genes25. It also regulates the activity of a tumor suppressor, phosphatase and tensin homolog (PTEN)26, activates protein kinase G1α to relax vascular smooth muscle27, and suppresses the activity of glyceraldehyde 3-phosphate dehydrogenase (GAPDH)28.
The present study showed that H2S2 and H2S3 were generated by the chemical interaction of H2S and NO. H2Sn and their effects on TRPA1 channels were abolished by cyanolysis and by reducing substances, such as dithiothreitol (DTT), cysteine, and glutathione (GSH). However, HNO is resistant to cyanolysis, and SSNO− is resistant to reducing substances. These observations suggest that H2Sn, rather than HNO or SSNO−, are involved in the activation of TRPA1 channels.
Results
Generation of H2S2 and H2S3 by the chemical interaction of H2S and NO
The oxidation of H2S generates H2Sn20,21,22,23,29, and the interaction of H2S with S-nitroso cysteine generates cysteine persulfide30,31. Therefore, it is possible that the interaction of H2S with NO produces H2Sn. This possibility was examined using high performance liquid chromatography with tandem mass spectrometry analysis (LC-MS/MS). A mixture of Na2S, a sodium salt of sulfide, and diethylamine NONOate (DEA/NO), a donor of NO, was derivatized with monobromobimane, a fluorescence dye specific to thiols, and analyzed using LC-MS/MS. H2S2 and H2S3 were generated by consuming H2S after mixing Na2S and DEA/NO in a concentration dependent manner (Fig. 1).
Figure 1. LC/MS/MS analysis of products generated by the interaction of H2S with NO.

(a,b,c) Chromatograms and relative levels of dibimane-derivatives of H2S (S-dibimane) (a), H2S2 (SS-dibimane) (b), and H2S3 (SSS-dibimane) (c) generated in the mixture of Na2S and DEA/NO. (n = 3).
Activation of TRPA1 channels by H2S2 and H2S3 generated by the interaction of H2S with NO
H2Sn activates TRPA1 channels in astrocytes and dorsal root ganglion (DRG) neurons23,24. These observations and above-mentioned results suggest the possibility that the mixture of H2S and NO activates TRPA1 channels through the generation of H2S2 and H2S3. The activation of TRPA1 channels in DRG neurons by mixture of H2S and NO was examined by measuring the Ca2+ influx with Fluo-4, a Ca2+ sensitive fluorescence dye. The mixture of Na2S and DEA/NO induced Ca2+ influx in DRG neurons, which were sensitive to allyl isothiocyanate (AITC), an agonist of TRPA1 channels, and suppressed by HC-030031, an antagonist of the channels, in a concentration-dependent manner. However, Na2S or DEA/NO alone induced negligible response (Fig. 2a,b and c; Supplemental Fig. 1a and b). These observations suggest that the interaction of H2S with NO generates H2S2 and H2S3 that activate TRPA1 channels.
Figure 2. Generation of H2S2 and H2S3 by the interaction of H2S with NO, and their effect on TRPA1 channels.

(a) Ca2+ influx induced by the product of H2S and NO interaction. The images and traces of Ca2+ influx induced by 20 μM Na2S, 20 μM DEA/NO, a mixture of 20 μM each of Na2S-DEA/NO, 100 μM AITC, and 50 mM KCl detected in fluo-4-loaded AITC-responsive and -unresponsive DRG neurons are shown. The bar in the image indicates 30 μm. (b) Concentration-Ca2+ response relations for H2Sn generated from H2S and NO. Data were obtained from AITC-responsive cells (n = 29–37). Ca2+-responses were normalized to those obtained with 50 mM KCl applied for 5 min after the application of AITC. (c) Ca2+ influx induced on the activation of TRPA1 channels by the products generated by the interaction of H2S with NO. The products from Na2S and DEA/NO induced Ca2+ influx that was suppressed by TRPA1 channel specific inhibitor, 30 μM HC-030031, in DRG neurons pre-loaded with fluo-4. (n = 30–33). (d) The intracellular levels of H2Sn in cells applied with H2S and NO. Responses to 20 μM Na2S, 20 μM DEA/NO, a mixture of 20 μM each of Na2S-DEA/NO, and 30 μM Na2S2 are shown in DRG neurons pre-loaded with SSip-1. (e) Concentration-SSip-1 response relations for H2Sn generated from H2S and NO (n = 28–30). SSip-1 responses were normalized by those to 30 μM Na2S2, applied 20 min after exposure to Na2S-DEA/NO mixture. The bar in the image indicates 30 μm. Data are represented as means ± SEM. **p < 0.01 (unpaired Student’s t-test).
H2S2 and H2S3 generated by the reaction of H2S and NO in DRG neurons were also examined using SSip-1, a fluorescence probe that specifically and reversibly binds to sulfane sulfur32 (Supplemental Fig. 2). The levels of H2Sn generated by a mixture of Na2S and DEA/NO increased in a concentration-dependent manner in DRG neurons, while Na2S alone resulted in a slight increase in H2Sn levels, and no changes were induced by DEA/NO alone (Fig. 2d and e). HNO and H2O2, which activate TRPA1 channels, were not detected by SSip-114,33 (Supplemental Fig. 3a and b). These observations suggest that the interaction of H2S with NO produces H2S2 and H2S3 that activate TRPA1 channels.
Instantaneous generation of H2Sn on exposure of H2S to NO
The rate of H2Sn generation from H2S and NO was examined. Na2S releases H2S immediately after dissolving in the medium, while DEA/NO slowly releases NO after its dissolution34. Therefore, it is possible that the generation of H2Sn depends on the release of NO into the medium. The levels of H2Sn generated from the mixture of Na2S and DEA/NO were measured by SSip1 with or without pre-incubation with DEA/NO. The Ca2+ influx induced by the products from Na2S and DEA/NO was also examined. Thirty seconds after mixing Na2S with DEA/NO solution, which had been dissolved for 5 min, NO was fully released from DEA/NO and generated H2Sn (Fig. 3a and b) that effectively induced Ca2+ influx (Fig. 3c and d). However, the mixture of Na2S and DEA/NO without any pre-release of NO generated much less amount of H2Sn after 30 seconds (Fig. 3a and b) and induced a weak Ca2+ influx (Fig. 3c and d). The release of NO from DEA/NO is a rate-limiting step for the generation of H2Sn. Once NO is released into the reaction mixture, H2Sn is immediately produced (Fig. 3b). Incubation of Na2S with DEA/NO fully for 5 min was thus used for the other experiments in this study, as it fully generates H2Sn.
Figure 3. H2Sn is produced immediately after the exposure of H2S to NO.

(a) A release of NO from DEA/NO is rate-limiting for the generation of H2Sn. SSip-1 responses to Na2S-DEA/NO mixtures with (w) or without (w/o) 5 min incubation of DEA/NO in HBS, before mixing with Na2S, are shown. (b) The time-course of the production of H2Sn after mixing Na2S and DEA/NO with or without 5 min pre-incubation (n = 29–30). SSip-1 responses were normalized by those obtained on exposure to 30 μM Na2S2. (c) Ca2+ influx induced by H2Sn produced by the mixture of H2S with NO. Fifteen traces are shown. Twenty micro molar Na2S was mixed for 30 sec with 20 μM DEA/NO with or without 5 min-pre-incubation in HBS. (d) The amplitude of Ca2+ influx induced by the product of H2S and NO depends on the pre-incubation of DEA/NO to release NO. Release of NO from DEA/NO rate-limits the generation of H2Sn and the induction of Ca2+ influx in AITC-responsive cells (n = 25–28). Data are represented as means ± SEM. **P < 0.01 (unpaired Student’s t-test).
H2Sn generated from H2S and NO was degraded by cyanolysis but not HNO
It has been proposed that H2S interacts with NO to produce HNO, which activates TRPA1 channels14. HNO is resistant, while H2Sn is sensitive, to cyanolysis14,35. We examined sensitivity of the product of H2S and NO interaction to cyanolysis using SSip-1 in DRG neurons. The mixture of Na2S and DEA/NO, which was incubated for 5 min to fully produce H2Sn, was exposed to NaCN, and the mixture was applied to DRG neurons. The exposure to NaCN dramatically decreased the amount of H2Sn generated from H2S and NO (Fig. 4a). A similar vulnerability to cyanolysis was observed for H2S2 and H2S3 (Fig. 4b and c; Supplemental Fig. 3c and d). These observations suggest that molecules generated by the interaction of H2S with NO are H2S2 and H2S3, rather than HNO.
Figure 4. The reaction products of H2S and NO are degraded by cyanolysis.

(a,b and c) The products of H2S and NO (a), H2S2 (b) and H2S3 (c) were degraded by cyanolysis. Mixtures of 20 μM each of Na2S and DEA/NO after incubation for 5 min, and 10 μM each of Na2S2 and Na2S3 were treated with 20 mM NaCN. After incubation with NaCN or NaCl as a control, each solution was applied to SSip-1-loaded DRG neurons (n = 29–30). SSip-1 responses were normalized with those obtained on exposure to 30 μM Na2S2.
We confirmed these results by measuring Ca2+ influx induced by the mixture of H2S with NO in DRG neurons. The Ca2+ influx induced by the mixture of Na2S and DEA/NO, as well as that by Na2S2 and Na2S3, was suppressed by cyanolysis, while that induced by Angeli’s salt, a HNO donor, and H2O2 was not affected by cyanolysis (Fig. 5). These observations confirmed that H2S2 and H2S3, but not HNO, were generated from H2S and NO to activate TRPA1 channels.
Figure 5. Ca2+ influx induced by the mixture of H2S and NO is abolished by cyanolysis, but not that induced by HNO.

(a,b) Ca2+ responses were induced by the mixture of 20 μM each of Na2S and DEA/NO (a) or 0.3 mM Angeli’s salt (b) pre-incubated for 10 min in the presence or absence of 20 mM NaCN. (c) Sensitivity of Ca2+ influx induced by the interaction of H2S with NO, as well as HNO, to cyanolysis. Ca2+ responses induced by 10 μM Na2S2, 10 μM Na2S3, a mixture of 20 μM each Na2S and DEA/NO, 0.3 mM Angeli’s salt, and 1 mM H2O2 in the presence or absence of 20 mM NaCN are shown. Data were obtained from AITC-responsive cells (n = 26–33). Angeli’s salt and H2O2 were treated with NaCN immediately after dilution in HBS. Ca2+-responses were normalized by those obtained in response to 50 mM KCl. Ca2+-responses non-specifically induced by NaCN were subtracted from those induced by each stimulant in the presence of NaCN. Data are represented as means ± SEM. **P < 0.01 (unpaired Student’s t-test).
H2Sn generated by the interaction of H2S and NO was degraded by reducing substances but not SSNO−
SSNO− has also been proposed to be produced by the interaction between H2S and NO15. SSNO− is resistant to reducing substances, such as cysteine, GSH, and DTT, while H2Sn is degraded by them15,31. We examined the generation of H2Sn and SSNO− from H2S and NO, and compared their sensitivity to cysteine, GSH, and DTT by measuring the light absorbance at a specific wavelength. The production of both H2Sn (absorbance at 290–300 nm) and SSNO− (412 nm) increased after mixing a high concentration 5 mM Na2S and 2 mM DEA/NO in a time-dependent manner15 (Fig. 6a). In the presence of a high concentration 3 mM each of cysteine, GSH, and DTT, the levels of H2Sn decreased, while those of SSNO− remained unchanged (Fig. 6b).
Figure 6. Products from H2S and NO are degraded by reducing substances.

(a and b) H2Sn generated from H2S and NO are degraded by cysteine (Cys), GSH, and DTT, while SSNO- is resistant to them. UV-Vis spectra of 5 mM Na2S and 2 mM DEA/NO in the presence (b) or absence (a) of 3 mM each cysteine, GSH, and DTT in 0.5 M HEPES buffer (pH 7.4). Na2S and DEA/NO were mixed at the ‘start’, and cysteine, GSH, and DTT were applied 10 min after the ‘start’. H2Sn is observed at 290–300 nm, and SSNO− at 412 nm. (c) SSip-1 responses to the mixture of 20 μM each Na2S and DEA/NO pretreated with or without 30 μM cysteine for 5 min. (d) Concentration-response relations for degradation of the product from H2S and NO by reducing substances. SSip-1 responses to 10 μM Na2S2, 10 μM Na2S3, and the mixture of 20 μM each of Na2S and DEA/NO with or without pre-treatment with cysteine, GSH, and DTT (n = 28–30). (e) Ca2+ influx induced by the mixture of H2S and NO is suppressed by a pre-treatment with cysteine. Fifteen traces of Ca2+-responses to the mixture of 20 μM each of Na2S and DEA/NO pretreated with or without 30 μM cysteine were shown. (f) The effects of pre-treatment of reducing substances on Ca2+ influx induced by the mixture of H2S and NO. Pre-treatment of 30 μM each cysteine, GSH and DTT suppressed Ca2+-responses to 10 μM Na2S2, 10 μM Na2S3, and the mixture of 20 μM each Na2S and DEA/NO in AITC-responsive cells (n = 21–35). Na2S2, Na2S3, and 5 min pre-incubated mixture of Na2S with DEA/NO were treated with cysteine, GSH, and DTT for 5 min. Data are represented as means ± SEM. **P < 0.01 (Dunnett’s test).
The levels of H2Sn produced from the interaction of physiological concentrations of Na2S with DEA/NO in the presence or absence of reducing substances were also examined using SSip-1 in DRG neurons. The levels of H2Sn increased in DRG neurons on exposure to the mixture of Na2S and DEA/NO (20 μM each), while those exposed to cysteine, GSH, and DTT were suppressed in a concentration dependent manner (Fig. 6c and d). A similar result was observed with Na2S2 and Na2S3 (Fig. 6c and d).
The effect of H2Sn generated from H2S and NO on the Ca2+ influx through the activation of TRPA1 channels was also examined. The mixture of 20 μM each of Na2S and DEA/NO induced Ca2+ influx, while only a weak Ca2+ influx was induced by the mixture exposed to 30 μM each of cysteine, GSH, and DTT (Fig. 6e and f). The effect of 10 μM each of Na2S2 and Na2S3 on Ca2+ influx was suppressed by these reducing substances in a similar manner (Fig. 6f). The higher concentrations of cysteine were required for suppressing the effect of HNO (Supplemental Fig. 1c). These observations suggest that H2S2 and H2S3, rather than SSNO−, are produced from the mixture of H2S and NO to activate TRPA1 channels.
Discussion
The present study shows a mechanism of chemical interaction between H2S and NO to generate other signaling molecules, H2S2 and H2S3, which activate TRPA1 channels (Figs 1 and 2).
The one electron oxidation of sulfide produces thiyl radicals, which readily react with NO radical to generate thionitrous acid (HSNO).
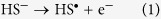 |
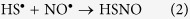 |
At the physiological pH S⦁− must be the major reactive form and produces polysulfides.
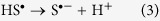 |
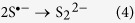 |
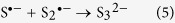 |
HSNO produces nitroxyl in the presence of H2S and persulfide, which in turn reacts with HSNO to produce nitrosopersulfide.
 |
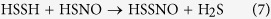 |
Eberhardt et al. reported that HNO and H2Sn are produced by the interaction of H2S with NO14. As shown in equations 1, 2 and 6, the interaction of H2S and NO generates HSNO, which further reacts with H2S to generate HNO14,36. They measured produced HNO in the presence of 2 μM to 75 μM each of H2S and NO by its selective electrode but not characterized the other products H2Sn (equations 3, 4, 5 and 6) nor the stability of the products against cyanide. HNO is stable to cyanide, while H2Sn are degraded in cyanolysis. The present study showed that the products from H2S and NO are degraded in cyanolysis as readily as H2Sn (Figs 4 and 5). Although both HNO and H2Sn can be produced as shown in equation 6, H2Sn must be the chemical entities that activate TRPA1.
Cortese-Krott et al. showed that SSNO− and H2Sn were produced from H2S and NO as shown in equations 2, 6 and 715,37. The study was performed in the presence of 1 to 2 mM NO donors with various ratio of 1 to 10 mM high concentrations of H2S. The present study agreed with their results that SSNO− was detected as absorbance at 412 nm in the presence of 5 mM high concentrations of H2S with 2 mM NO (Fig. 6). However, it was not detected under physiological concentrations of less than 20 μM of Na2S and DEA/NO15 (Figs 1a and 6). SSNO−, which may have a role as a carrier or donor of NO, relaxed vascular smooth muscle, but they suggested that NO-independent effects must attributed to H2Sn15. SSNO− is stable in the presence of high concentration of thiols15, while H2Sn as well as the effective molecules produced from H2S and NO are degraded by reducing substances as shown in the present study (Fig. 6). Under physiological conditions the interaction of H2S and NO can produce H2Sn as major products to induce physiological effects.
H2Sn generation from H2S and NO is a fast reaction. The release of NO from DEA/NO is rate limiting for the generation of H2Sn (Fig. 3), as H2S immediately reacts with the released NO to produce H2Sn. This instantaneous production of H2Sn from H2S and NO is important for its physiological roles. H2S producing enzymes, 3MST, cystathionine β–synthase, and NOS, are localized to neurons and astrocytes in the central nervous system, and H2S and NO interact with each other to produce H2Sn that activates TRPA1 channels to modify synaptic activity18,23,38,39.
In the cardiovascular system, high concentrations of H2S are detected in aortic tissues, and the cysteine aminotransferase/3MST, endothelial NOS and cystathionine γ–lyase pathways are localized to vascular endothelium and smooth muscle, respectively2,3,40. H2Sn produced from H2S and NO can activate PKG1α to induce vascular relaxation27,41.
H2Sn has various other physiological effects. It regulates the activity of a tumor suppressor PTEN26, facilitates the translocation of Nrf225, and suppresses the activity of GAPDH28. H2S2 and H2S3 are also produced by 3MST from 3MP, and from H2S by oxidation or interaction with NO as shown in the present study20,23,30. It is possible that some effects that were previously attributed to H2S or NO alone due to H2Sn generated by the interaction of H2S with NO.
Thus interaction of H2S and NO yielding H2Sn plays an important physiological role and provides therapeutic targets for diseases involving these molecules.
Methods
Chemicals
All methods were performed in accordance with the guidelines and regulations of chemical substance management and approved by the committees of chemical substance management in the National Institute of Neuroscience, National Center of Neurology and Psychiatry. Diethylamine NONOate (DEA/NO) (Cayman Chemical, Ann Arbor, MI) and Angeli’s salt (Cayman Chemical) were dissolved at 0.1 M in 10 mM NaOH. Na2S2 (Dojindo, Kumamoto, Japan), Na2S3 (Dojindo), Na2S (Wako pure chemicals, Osaka, Japan), L-cysteine (Cys) (Wako), glutathione (Wako), dithiothreitol (Wako) were dissolved at 0.1 M in ultrapure water. These stock solutions were stored at −80 °C, and were used within a week. NaCN (Wako) was dissolved at 0.5 M in a 0.1 M HEPES buffer (pH 7.4) and stored at −20 °C.
Isolation of sensory neurons
All the animal procedures were performed in accordance with the guidelines and regulations of animal care and use and approved by the committees of animal care and use in the National Institute of Neuroscience, National Center of Neurology and Psychiatry. DRG were dissected from 1–13 days old Sprague-Dawley rats (CLEA JAPAN, Tokyo, Japan). After the treatment of DRG with collagenase and trypsin, cells were dispersed by pipetting, and suspended in Neurobasal medium supplemented with 2% NS supplement (Wako) and 2% fetal bovine serum, and then seeded onto poly-D-lysine-coated coverslips. The cells, which were incubated in a humidified atmosphere with 5% CO2 at 37 °C, were used for imaging experiments within 32 h after seeding.
Imaging of intracellular Ca2+ and H2Sn
Fluo-4 AM (Thermo Fisher Scientific, Waltham, MA, USA) and SSip-1 DA (synthesized as described in ref. 32) were diluted in a HEPES-buffered saline (HBS; in mM: 137 NaCl, 5.4 KCl, 0.8 MgCl2, 1.8 CaCl2, 10 glucose, 10 HEPES (pH 7.4)). DRG neurons were loaded with 5 μM fluo-4 AM in 0.02% cremophor EL for 45 min at room temperature or with 20 μM SSip-1 DA in 0.02% cremophor EL for 45 min at 37 °C. A coverslip was mounted on an upright microscope (DM LFS, Leica, Heidelberg, Germany) and was perfused with HBS at a rate of 1 ml/min. The recording was started after 15-min perfusion of HBS to let the intracellular probes completely esterized. Fluorescence was recorded every 5 sec with a bandpass filter (excitation at 480/40 nm, emission at 527/30 nm) and a CCD camera (Hamamatsu Photonics, Shizuoka, Japan). Images were acquired using Aquacosmos 2.6 software (Hamamatsu Photonics). Experiments were performed at room temperature.
At the end of the experiments, 50 mM KCl or 30 μM Na2S2 was applied to neurons to induce maximal Ca2+- or SSip-1-responses, respectively. Unless otherwise described, the amplitudes of Ca2+- and SSip-1-responses evoked by tested stimulus were normalized by those to KCl and Na2S2. In Ca2+ imaging, neurons responded to AITC with the amplitudes over 20% of those to KCl were considered as TRPA1-expressing neurons.
LC-MS/MS analysis
Two point five to 5 μl 0.1 M each Na2S2, Na2S3, Na2S, and DEA/NO stock solutions were diluted in a 95 μl 10 mM HEPES buffer (pH 7.4). Immediately after dilution of Na2S2 and Na2S3, or five minutes incubation for the mixture of Na2S and DEA/NO), 40 μl of 0.5 M CHES (pH 8.4) and 2 μl 0.05 M monobromobimane were added. Twenty minutes after incubation at room temperature in the dark, samples were neutralized with 15 μl of 30% acetic acid.
Dibimane derivatives were analyzed by the triple-quadrupole mass spectrometer coupled to HPLC (Shimadzu LCMS-8040). Samples were subjected to a reverse phase Symmetry C18 HPLC column (4.6 × 250 mm, Waters) at the flow rate of 0.8 ml/min. The mobile phase consisted of (A) 5 mM ammonium formate in water and (B) 5 mM ammonium formate in a 1:1 solution of water and methanol. Samples were separated by eluting with a gradient: 40% B at 0 min, and 75% B at 8 min and remained it for 10 min. The column oven was maintained at 40 °C. The effluent was subjected to the mass spectrometer using an electrospray ionization (ESI) interface operating in the positive-ion mode. The source temperature was set at 400 °C, and the ion spray voltage at 4.5 kV. Nitrogen was used as a nebulizer and drying gas. The tandem mass spectrometer was tuned in the multiple reaction monitoring mode to monitor mass transitions m/z Q1/Q3 432.45/192.00 [S-dibimane + NH4]+, 464.55/192.00 [SS-dibimane + NH4]+, 496.60/192.00 [SSS-dibimane + NH4]+.
UV-Vis spectrometry
UV-Vis spectra were recorded with a UV/Vis spectrophotometer (Beckman Coulter, Brea, CA, USA). A silica cuvette with a 1.0-cm light path and 100-μl volume was used for the recording. One μl stock solutions of test compounds were diluted in 100 μl solution buffered with 0.5 M HEPES-NaOH (pH 7.4). UV/Vis spectra were recorded immediately as well as every 1 min after the dilution.
Statistical analysis
All the statistical analysis was performed using OriginLab software (LightStone, Tokyo, Japan). Statistical comparisons between two groups were performed by unpaired Student’s t-test. For comparisons between multiple groups, one-way ANOVA was performed followed by Dunnett’s test. P < 0.05 was considered statistically significant.
Additional Information
How to cite this article: Miyamoto, R. et al. Polysulfides (H2Sn) produced from the interaction of hydrogen sulfide (H2S) and nitric oxide (NO) activate TRPA1 channels. Sci. Rep. 7, 45995; doi: 10.1038/srep45995 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Acknowledgments
This work was supported by the National Institute of Neuroscience, KAKENHI (26460115), a Grant-in-Aid for Scientific Research, the Strategic Research program for Brain Sciences from Japan Agency for Medical Research and development, a AMED, the Uehara Memorial Foundation, Yamazaki Spice Promotion Foundation to H.K., and KAKENHI (26460352), a Grant-in-Aid for Scientific Research to Y.K. and KAKENHI (16K15123), a Grant-in-Aid for Scientific Research to N.S.
Footnotes
The authors declare no competing financial interests.
Author Contributions R.M. and H.K. designed the study, analyzed the data, and wrote the paper. R.M. performed most of the experiments and N.S. and Y.K. supported them. Y.T., K.H., and Y.U. provided SSip-1. S.K. performed the LC-MS/MS experiments and Y.O. supervised them.
References
- Abe K. & Kimura H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 16, 1066–1071 (1996). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hosoki R., Matsuki N. & Kimura H. The possible role of hydrogen sulfide as an endogenous smooth muscle relaxant in synergy with nitric oxide. Biochem. Biophys. Res. Commun. 237, 527–531 (1997). [DOI] [PubMed] [Google Scholar]
- Zhao W., Zhang J., Lu Y. & Wang R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 20, 6008–6016 (2001). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura Y. & Kimura H. Hydrogen sulfide protects neurons from oxidative stress. FASEB J. 18, 1165–1167 (2004). [DOI] [PubMed] [Google Scholar]
- Zanardo R. C. O. et al. Hydrogen sulphide is an endogenous modulator of leukocyte-mediated inflammation. FASEB J. 20, 2118–2120 (2006). [DOI] [PubMed] [Google Scholar]
- Elrod J. W. et al. Hydrogen sulfide attenuates myocardial ischemia-reperfusion injury by preservation of mitochondrial function. Proc. Natl. Acad. Sci. USA 104, 15560–15565 (2007). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teague B., Asiedu S. & Moore P. K. The smooth muscle relaxant effect of hydrogen sulphide in vitro: Evidence for a physiological role to control intestinal contractility. Br. J. Pharmacol. 137, 139–145 (2002). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bir S. C. et al. Hydrogen sulfide stimulates ischemic vascular remodeling through nitric oxide synthase and nitrite reduction activity regulating hypoxia-inducible factor-1α and vascular endothelial growth factor-dependent angiogenesis. J. Am. Heart Assoc. 1, e004093 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coletta C. et al. Hydrogen sulfide and nitric oxide are mutually dependent in the regulation of angiogenesis and endothelium-dependent vasorelaxation. Proc. Natl. Acad. Sci. USA 109, 9161–9166 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minamishima S. et al. Hydrogen sulfide improves survival after cardiac arrest and cardiopulmonary resuscitation via a nitric oxide synthase 3-dependent mechanism in mice. Circulation 120, 888–896 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- King A. L. et al. Hydrogen sulfide cytoprotective signaling is endothelial nitric oxide synthase-nitric oxide dependent. Proc. Natl. Acad. Sci. USA 111, 3182–3187 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whiteman M. et al. Evidence for the formation of a novel nitrosothiol from the gaseous mediators nitric oxide and hydrogen sulphide. Biochem. Biophys. Res. Commun. 343, 303–310 (2006). [DOI] [PubMed] [Google Scholar]
- Miljkovic J. L., Kenkel I., Ivanovic-Burmazovic I. & Filipovic M. R. Generation of HNO and HSNO from nitrite by heme-iron-catalyzed metabolism with H2S. Angew. Chem. Int. Ed. 52, 12061–12064 (2013). [DOI] [PubMed] [Google Scholar]
- Eberhardt M. et al. H2S and NO cooperatively regulate vascular tone by activating a neuroendocrine HNO-TRPA1-CGRP signaling pathway. Nat. Commun. 5, 4381 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortese-Krott M. M. et al. Key bioactive reaction products of the NO/H2S interaction are S/N-hybrid species, polysulfides, and nitroxyl. Proc. Natl. Acad. Sci. USA 112, E4651–4660 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura H. Hydrogen polysulfide (H2Sn) signaling along with hydrogen sulfide (H2S) and nitric oxide (NO). J. Neural Transm. 123, 1235–1245 (2016). [DOI] [PubMed] [Google Scholar]
- Moustafa A. & Habara Y. Cross talk between polysulfide and nitric oxide in rat peritoneal mast cells. Am. J. Physiol. Cell Physiol. 310, C894–C902 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shibuya N. et al. 3-Mercaptopyruvate sulfurtransferease produces hydrogen sulfide and bound sulfane sulfur in the brain. Antioxid. Redox Signal. 11, 703–714 (2009). [DOI] [PubMed] [Google Scholar]
- Shibuya N. et al. A novel pathway for the production of hydrogen sulfide from D-cysteine in mammalian cells. Nat. Commun. 4, 1366 (2013). [DOI] [PubMed] [Google Scholar]
- Kimura Y. et al. Identification of H2S3 and H2S produced by 3-mercaptopyruvate sulfurtransferase in the brain. Sci Rep 5, 14774 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagai Y., Tsugane M., Oka J.-I. & Kimura H. Polysulfides induce calcium waves in rat hippocampal astrocytes. J. Pharmacol. Sci. 100, 200 (2006). [Google Scholar]
- Oosumi K. et al. Polysulfide activates TRP channels and increases intracellular Ca2+ in astrocytes. Neurosci. Res. 685, e109–e222 (2010). [Google Scholar]
- Kimura Y. et al. Polysulfides are possible H2S-derived signaling molecules in rat brain. FASEB J. 27, 2451–2457 (2013). [DOI] [PubMed] [Google Scholar]
- Hatakeyama Y., Takahashi K., Tominaga M., Kimura H. & Ohta T. Polysulfide evokes acute pain through the activation of nociceptive TRPA1 in mouse sensory neurons. Mol. Pain 11:24 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koike S., Ogasawara Y., Shibuya N., Kimura H. & Ishii K. Polysulfide exerts a protective effect against cytotoxicity cuased by t-buthylhydroperoxide through Nrf2 signaling in neuroblastoma cells. FEBS lett 587, 3548–3555 (2013). [DOI] [PubMed] [Google Scholar]
- Greiner R. et al. Polysulfides link H2S to protein thiol oxidation. Antioxid. Redox Signal. 19, 1749–1765 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stubbert D. et al. Protein kinase G Iα oxidation paradoxically underlies blood pressure lowering by the reductant hydrogen sulfide. Hypertension 64, 1344–1351 (2014). [DOI] [PubMed] [Google Scholar]
- Jarosz A. P. et al. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) is inactivated by S-sulfuration in vitro. Free Rad. Biol. Med. 89, 512–521 (2015). [DOI] [PubMed] [Google Scholar]
- Toohey J. I. Sulfur signaling: Is the agent sulfide or sulfane? Anal Biochem 413, 1–7 (2011). [DOI] [PubMed] [Google Scholar]
- Kabil O., Motl N. & Banerjee R. H2S and its role in redox signaling. Biochim. Biophys. Acta 1844, 1355–1366 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishanina T. V., Libiad M. & Banerjee R. Biogenesis of reactive sulfur species for signaling by hydrogen sulfide oxidation pathways. Nat. Chem. Biol. 11, 457–464 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takano Y. et al. Development of a reversible fluorescent probe for reactive sulfur species, sulfane sulfur, and its biological application. Chem. Commun. 53, 1064–1067 (2017). [DOI] [PubMed]
- Andersson D. A., Gentry C., Moss S. & Bevan S. Transient receptor potential A1 is a sensory receptor for multiple products of oxidative stress. J. Neurosci. 28, 2485–2494 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keefer L. K., Nims R. W., Davies K. M. & Wink D. A. “NONOates” (1-substituted diazen-1-ium-1,2-diolates) as nitric oxide donors: convenient nitric oxide dosage forms. Methods Enzym. 268 (1996). [DOI] [PubMed] [Google Scholar]
- Jarabak R. & Westley J. The sulfur-cyanolysis sites of serum albumin: metabolite competition studies. J. Biochem. Toxicol. 4, 255–261 (1989). [DOI] [PubMed] [Google Scholar]
- Filipoovic M. R. et al. Chemical characterization of the smallest S-nitrosothiol, HSNO; cellular cross-talk of H2S and S-nitrosothiols. J. Am. Chem. Soc. 134, 12016–12027 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cortese-Krott M. M. et al. Nitrosopersulfide (SSNO−) accounts for sustained NO bioactivity of S-nitrosothiols following reaction with sulfide. Redox Biol. 2, 234–244 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagahara N., Ito T., Kitamura H. & Nishino T. Tissue and subcellular distribution of mercaptopyruvate sulfurtransferase in the rat: confocal laser fluorescence and immunoelectron microscopic studies combined with biochemical analysis. Histochem. Cell. Biol. 110, 243–250 (1998). [DOI] [PubMed] [Google Scholar]
- Enokido Y. et al. Cystathionine beta-synthase, a key enzyme for homocysteine metabolism, is preferentially expressed in the radial glia/astrocyte lineage of developing mouse CNS. FASEB J. 19, 1854–1856 (2005). [DOI] [PubMed] [Google Scholar]
- Shibuya N., Mikam, , Kimura Y., Nagahara N. & Kimura H. Vascular endothelium exresses 3-mercaptopyruvate sulfurtransferase and produces hydrogen sulfide. J. Biochem. 146, 623–626 (2009). [DOI] [PubMed] [Google Scholar]
- Levitt M. D., Abdel-Rehim M. S. & Furne J. Free and acid-labile hydrogen sulfide concentrations in mouse tissues:Anomalously high free hydrogen sulfide in aortic tissue. Antioxid. Redox Signal. 15, 373–378 (2011). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.