

Abstract
Background
Colonization of the body is an important step in Staphylococcus aureus infection. S. aureus colonizes skin and mucous membranes in humans and several animal species. One important ecological niche of S. aureus is the anterior nares. More than 60% of the S. aureus in the nose are found in vestibulum nasi. Our aim was to describe the localization of S. aureus in nasal tissue from healthy carriers.
Methods
Punch skin biopsies were taken from vestibulum nasi from healthy volunteers (S. aureus carriers and non−/intermittent carriers, n = 39) attending the population-based Tromsø 6 study. The tissue samples were processed as frozen sections before immunostaining with a specific S. aureus antibody, and finally evaluated by a confocal laser-scanning microscope.
Results
Our results suggest that S. aureus colonize both the upper and lower layers of the epidermis within the nasal epithelium of healthy individuals. The number of S. aureus in epidermis was surprisingly low. Intracellular localization of S. aureus in nasal tissue from healthy individuals was also detected.
Conclusions
Knowledge of the exact localization of S. aureus in nasal tissue is important for the understanding of the host responses against S. aureus. Our results may have consequences for the eradication strategy of S. aureus in carriers, and further work can provide us with tools for targeted prevention of S. aureus colonisation and infection.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-017-0997-3) contains supplementary material, which is available to authorized users.
Keywords: Nasal colonization, Staphylococcus aureus, Carrier, Localization, Epithelial cells, Intracellular
Background
The nose is an important site for Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) colonization in humans, and S. aureus is predominantly located in the anterior nasal vestibule on the septum adjacent to the nasal ostium on the moist squamous epithelium [1, 2]. In this location, S. aureus can exist as a commensal without any signs or symptoms of an infection. The prevalence of persistent nasal carriage with S. aureus is 20–30% in adult human populations [3, 4], and most often it is an endogenous S. aureus strain that causes infection [5–7]. Prevention and elimination of the carrier state may contribute in reducing the S. aureus disease burden [4]. Successful nasal decolonization of S. aureus is difficult to achieve, and S. aureus readily recolonizes the nose, the throat and other sites within one week, commonly with the same S. aureus genotype [8–11]. Today, nasal mupirocin is the most efficacious regimen of S. aureus eradication from the anterior nares [12, 13]. However, the success rate for eradication of MRSA is low 12 months after decolonization [14].
The anterior part of the nasal cavity (vestibulum nasi) is lined with stratified, keratinized, squamous epithelium. More than 60% of S. aureus in the nasal vestibule are found here [1]. It has also been shown that S. aureus can bind to the ciliated nasal epithelial cells in the inner part of the nasal cavity (internal nares) with pseudostratified columnar ciliated epithelium [15–18]. The skin in vestibulum nasi consists of two main layers: epidermis and dermis [19]. Epidermis consists of five strata called the stratum corneum, stratum lucidum, stratum granulosum, stratum spinosum, and stratum basale [20, 21]. The five main strata are characterized by cells at varying stages of development. Stratum corneum, also called the cornified layer, is the outermost layer and contains mature keratinocytes called corneocytes [21, 22]. The stratum lucidum is a thin, translucent layer with no nuclei. Stratum granulosum are flattened keratinocytes which contain cytoplasmic granula and secretory organelles [21, 23]. Stratum spinosum consists of several layers of large polygonal keratinocytes, also called spinous keratinocytes. Stratum basale is the layer that separates dermis from epidermis, and it consists of undifferentiated epidermal cells. The cells in this layer are responsible for constant renewal of cells in epidermis [21]. Dermis is a layer of connective tissue and contains accessory epidermal structures, lymphatic and vascular conduits, nerves and nerve endings, collagen and elastic fibers, in addition to many specialized immune cells [20, 21].
Successful adhesion of microbes to the human host depends on an efficient combination of microbial surface components as well as human host ligands. S. aureus adhesion to human epithelium is mediated by wall teichoic acid (WTA) and microbial surface components recognizing adhesive matrix molecules (MSCRAMMs) [24, 25]. WTA is important for both initial and late phases of S. aureus colonization, while surface proteins are important for long term S. aureus persistence in the nasal cavity [25]. Clumping factor B (ClfB) [26, 27] and the iron-regulated surface determinant A (IsdA) [26, 28] are important for nasal colonization and adhere to cytokeratin and loricrin/involucrin in corneocytes [26, 27, 29, 30]. S. aureus adhesion is also dependent on the host phenotype [25, 31, 32], but very few ligands in the host are known. Recently, it was shown that the interaction between the serine-aspartate repeat protein D (SdrD) and the human desmosome protein, desmoglein 1, is important for adhesion to keratinocytes in vitro [33].
S. aureus not only adhere to cell surfaces during colonization, but also invade both phagocytic and non-phagocytic cells where they can survive antibiotic treatment [16, 34, 35]. Here, they constitute a significant risk factor for recurrent episodes of disease, e.g. rhinosinusitis [15, 16], tonsillitis [36] and osteomyelitis [37].
For developing new strategies to eradicate S. aureus and diminish the risk of invasive disease, it is crucial to understand the host-pathogen interaction in colonization, invasion and infection. Fundamental questions of these aspects include; where in the epithelial layer is S. aureus located and how does S. aureus interact with cells in the nasal vestibule in healthy individuals? Thus, our aim was to determine where S. aureus localize in the nasal epithelium, and to evaluate a possible intracellular residency.
Methods
Study design and population
In 2010 an invitation letter for participation in the study was sent to a random selection of men and women age 40–42 years (n = 193) with known S. aureus nasal carrier status in the sixth Tromsø study (Tromsø 6) from October 2007–August 2008, a large population-based multipurpose health study [38, 39]. The S. aureus carrier status of the participants included in this study was defined in Tromsø 6 [38, 39]. Our working definition for nasal carriage types was according to van Belkum et al. [4]. There are two types of human nasal S. aureus carriers: persistent carriers and others, i.e. noncarriers and intermittent carriers are grouped together. A total of 48 individuals (25% of the invited) were willing to participate in this cross-sectional sub-study in 2010–2011. Among these volunteers, we performed a selection based on S. aureus carriage results from Tromsø 6 with the purpose of recruiting approximately 50% carriers of S. aureus and 50% non−/intermittent carriers, with an equal distribution of women and men. The selection represents a homogenous population according to age, with small age-related tissue-changes. The S. aureus noncarriers were considered as normal controls. Before tissue sampling, the volunteers were interviewed by a physician concerning known allergies, medicinal use, and ongoing cold/upper respiratory infection or fever. Exclusion criteria included known allergies against local anesthetics, and current use of anticoagulants (warfarin, acetylsalicylic acid) or other medication that preclude tissue sampling. If the subject was undergoing antibiotic treatment, symptomatic respiratory infection or fever at the time of sampling, the donor was asked to return for sampling after resolution of symptoms. Thirty-nine (n = 39) volunteers were finally included for nasal biopsy sampling at the University Hospital of North Norway (UNN HF), i.e. 18 females and 21 males, 18 S. aureus carriers and 21 non- /intermittent carriers.
Ethics statement
This work was performed in compliance with the ethical guidelines established by UiT- The Arctic University of Norway. Each participant gave written informed consent prior to the tissue sampling. Subject recruitment, enrollment and sampling of nasal tissue samples have been approved by the Regional Committee for Medical and Health Research Ethics, Rec North, Norway (Document reference 2010/146–7).
Microbiological sampling for detection and spa-typing of S. aureus
After interviewing the participants, a microbiological swab sample was taken from one nostril for confirmation of S. aureus carriage according to procedures described earlier [39]. S. aureus isolates and MRSA were identified using standard bacteriological criteria and laboratory methods, and all S. aureus isolates were spa-typed according to Sangvik et al. [39].
Tissue collection
A tissue sample was harvested from one nostril, while a microbiological sample was harvested from the other nostril. Selection of nostril (left or right side) for 3 mm punch biopsy sampling was done randomly by flipping a coin. Before tissue sampling, local anesthetic was injected (Xylocaine 1% infiltration anesthetic, 0.4 mm diameter hypodermic needle, approx. 1 ml) subcutaneously at the sampling site. Entry of injection cannula was at least 5 mm from sampling site. The thickness of the histological specimen was 2–4 mm. It included the whole of epidermis and parts of dermis. The biopsy was harvested from the transition zone between skin and mucosa in the upper lateral vestibulum nasi. In total, 33 samples were obtained from keratinized squamous epithelium without nasal hair and six samples from the same region containing nasal hair. The biopsy sampling was divided between two physicians.
Embedding of tissue and sectioning of frozen sections
Immediately after tissue sampling the tissue was embedded in Frozen Section Medium (OCT) (Fisher Scientific/Thermo Scientific), frozen in precooled liquid isopentane, and stored at −80 °C. The frozen tissue was sectioned in 10 μm sections with a glass/cryostat knife. The biopsy was sectioned vertically from the dermal site towards the epidermis with a cryostat blade that was cleaned with alcohol after obtaining each section. We started sectioning from one side of the biopsy towards the middle of the biopsy where most of our sections were obtained. Sections were placed on Super Frost Plus glass slides (Thermo Scientific) for immunohistochemistry and ordinary glasses for Hematoxylin-Eosin (HE)-staining. Sections and remaining tissue were stored at −80 °C for further use.
Fluorescence labeling/staining
To detect S. aureus, 10 μm cryostat sections of nasal tissue were fixed (4% Formaldehyde), blocked (2% Goat serum, Sigma-Aldrich; 0.1% cold Fish Skin Gelatin; 1% bovine serum albumin, Sigma-Aldrich; 0.1% Triton X-100; 0.05% Tween-20; diluted in 1X phosphate buffered saline PBS, ThermoFisher Scientific), and incubated with rabbit polyclonal S. aureus antibody (ab20920, Abcam) 1:5000 dilution over night at 4 °C followed by Alexa Fluor 546® goat anti-rabbit IgG (A-11010, Molecular Probes™, Thermo Fisher Scientific) 1:1000 dilution labeling, or Alexa Fluor 488® goat anti-rabbit IgG (A-11008, Molecular Probes™, Thermo Fisher Scientific) 1:1000 dilution labeling for 30–60 min. The slides were incubated with DRAQ5 1:1000 (BioStatus) to stain keratinocyte nuclei, and Alexa Fluor 594 Phalloidin (A12381, Molecular Probes™, Thermo Fisher Scientific) 1:40 dilution for staining actin red. The samples were dried and the glass slides mounted by adding Prolong Gold Antifade reagent with DAPI (P36935, Molecular Probes™, Thermo Fisher Scientific). Samples were stored at +4 °C before immediate microscopy.
Tissue analysis/microscopic analysis
Slides were analyzed by confocal microscopy using a Zeiss confocal microscope LSM510 Meta (Carl Zeiss Microscopy GmbH, Germany), with ×63 or ×40 objectives. Images were captured using the LSM510-imaging software system, version 4.2.0.121 (Carl Zeiss Inc.). For determining intracellular localization we used the Zeiss confocal microscope LSM510 Meta (Carl Zeiss Microscopy GmbH, Germany) using the LSM510 imaging software or Leica TCS SP5 (Leica Microsystems CMS GmbH, Germany) using the LAS AF software, version 2.3.0. For inspection of intracellular residency, multiple consecutive images were taken in the axial z-axis of tissue samples with a slice thickness of 0.2 μm or 0.5 μm. The tissue sections were assessed for evidence of S. aureus, visualized as bright green coccoid structures fluorescing on stimulation with a 488-nm laser. Samples were categorized as intracellular positive if S. aureus could be identified in at least one z-plane and closely localized adjacent to intraepithelial nuclei.
The relative amount of S. aureus present in the nasal tissue was calculated by randomly selecting minimum 10 consecutive tissue sections from 14 donors (eight carriers and six non−/intermittent carriers randomly selected), labeling the sections with S. aureus specific antibody, performing confocal laser scanning microscopy (CLSM) and calculating the mean number of S. aureus per tissue section. We counted all positive green cocci. Microscope images/slides were read and analyzed by two independent, blinded observers (RSO and AMH) for the presence of S. aureus in epidermis and intracellular localization.
Hematoxylin-eosin (HE) staining
We fixed 10 μm tissue sections in 4% formaldehyde and immersed with hematoxylin (Harris, Thermo Scientific), stained in Scott’s solution (NaHCO3, MgSO4, and H2O), and immersed in eosin (Thermo Scientific). The sections were dehydrated in 96% ethanol and 100% ethanol, immersed in xylene (Sigma-Aldrich), and finally mounted onto glass slides with Histokit (Fisher Scientific) before examination in light microscope. The HE stained sections were used to investigate histopathological alterations.
Gram-staining
For identification of Gram-positive cocci, frozen sections were stained following standard Gram-staining procedures (Sigma Aldrich). The Gram-stained sections were examined by light microscopy.
Bacterial controls and antibody specificity
To avoid cross-reaction with Staphylococcus epidermidis and to test the specificity of the S. aureus polyclonal antibody we included bacterial controls in the immuno-fluorescence staining. We used S. aureus ATCC 25923 as positive control for primary antibody binding, and S. epidermidis ATCC 12228 as negative control. Overnight cultures of the controls were made in 5 ml Brain-Heart Infusion broth. 1 ml o.n. culture was centrifuged for 10 min at 3000 rpm, the supernatant was removed and the pellet resuspended in 1 ml 1× PBS. Centrifugation was repeated for 10 min at 3000 rpm, the pellet was resuspended in 1 ml 1× PBS, and 1 μl suspension was spotted on a glass slide, before flame fixation. From this point, the positive and negative controls were prepared in exactly the same way as the frozen tissue sections. Green fluorescence was detected from the positive controls only.
Determination of S. aureus colony forming units (CFU) in the nose of healthy volunteers
To estimate the likelihood of detecting S. aureus in nasal tissue biopsies from healthy individuals, we performed a pilot study where 14 volunteers (colleagues working at the Department for Medical Biology, UiT – The Arctic University of Norway) were sampled by nasal swabbing according to Sangvik et al. 2011 [39], and the S. aureus CFU in the nose was determined. One nostril of each volunteer was swabbed in the anterior region, 1–1.5 cm inside the nostril. The swab was resuspended thoroughly in 1 ml of 0.9% sodium chloride. Twenty microliters from each tenfold dilution (100–108) was applied onto blood agar plates and ChromID S. aureus agar (Biomerieux) and incubated at 37 °C for 24 h. Colonies were counted in spots from two dilutions (3 if possible). The CFU/nostril was calculated, both total CFU and S. aureus CFU.
Results
S. aureus Carrier status versus results from confocal microscopic analyses and nasal swabbing
Thirty-nine healthy volunteers were included in the study, and their carrier status was determined in the previous Tromsø 6 study, i.e. 18 carriers and 21 non−/intermittent carriers. According to one swab sampling in this study, 16 out of 39 individuals were positive for S. aureus (Table 1), while 23 out of 39 individuals were negative for S. aureus. No MRSA were detected. S. aureus was observed in biopsy sections by CLSM in ten out of 18 persistent carriers (Table 1). Among the 21 non−/intermittent carriers, we found S. aureus in three of the biopsies by CLSM (Table 1). Thus, detection of S. aureus by one nasal swabbing and CLSM showed some inconsistency, i.e. three individuals that were negative for S. aureus by swabbing, both in the Tromsø 6 study and in the present study, were positive for S. aureus by CLSM, whereas three individuals with positive swab cultures in both studies, were negative for S. aureus by CLSM (Table 1). Two individuals defined as non−/intermittent carriers in the Tromsø 6 study, were positive for S. aureus by one swabbing, but negative by CLSM in our study (Table 1). Four individuals that were defined as persistent S. aureus carriers in the Tromsø 6 study were negative for S. aureus in our study based on swabbing and CLSM analysis (Table 1). Seven out of 39 biopsies were not examined in our study due to damaged sections, which was most likely caused by tissue sampling errors or sectioning errors (Table 1, not determined).
Table 1.
Results from nasal swab culture and confocal laser scanning microscopy (CLSM) of tissue sections
| S. aureus carriage status in the Tromsø 6 studya (n = 39) | Presence of S. aureus by one nasal swab sampling present studyc (n = 39) | Presence of S. aureus by CLSM present studyd (n = 39) | ||
|---|---|---|---|---|
| Persistent carriers (n = 18) | + | 14 | + | 10 |
| − | 3 | |||
| nd | 1 | |||
| − | 4 | + | 0 | |
| − | 4 | |||
| nd | 0 | |||
| Othersb (n = 21) | + | 2 | + | 0 |
| − | 2 | |||
| nd | 0 | |||
| − | 19 | + | 3 | |
| − | 10 | |||
| nd | 6 | |||
aResults from nasal swabbing (two samplings with median time interval of 28 days) and spa-typing in the Tromsø 6 study in October 2007–August 2008. S. aureus carriage status as defined by van Belkum et al. [4]
bIncludes both noncarriers and intermittent carriers
cResults from one nasal swab sampling and spa-typing in the present study in 2010–2011; +, positive for S. aureus; −, negative for S. aureus
dConfocal laser scanning microscopy (CLSM); +, positive for S. aureus; −, negative for S. aureus; nd, not determined, CLSM not performed
Fourteen different spa-types were found among the S. aureus isolates from the 39 healthy participants in this study (Additional file 1: Table S1). Twelve spa-types were found in one individual each, while t084 and t246 were found in two individuals each. Six individuals had changed spa-types since the Tromsø 6 study (Additional file 1: Table S1).
Estimation of S. aureus in nasal vestibule in healthy individuals
We performed a pilot study to estimate the likelihood of detecting S. aureus in biopsies. From nasal swabbing of 14 healthy colleagues, we found between 0 and 1.5 × 104 S. aureus CFU/nostril (Table 2). By a rough estimate, this should correspond to approximately 0–200 S. aureus per biopsy surface area.
Table 2.
Number of S. aureus found in nares of healthy individuals
| Number of S. aureus CFU/nostril (n = 14)a | |||||
|---|---|---|---|---|---|
| n = 8 | n = 2 | n = 1 | n = 1 | n = 1 | n = 1 |
| 0 | 50 | 1.5 × 102 | 5.0 × 102 | 6.0 × 102 | 1.5 × 104 |
aResults from a pilot study where we determined S. aureus CFU/nostril in 14 healthy colleagues from Department of Medical Biology by one nasal swab culture
Biopsies were then obtained from the 39 participants and HE staining and CLSM/immunohistochemistry (IHC) were performed. The cryo-sectioning of nasal tissue showed all layers of epidermis, confirming the quality of the biopsies (Fig. 1).
Fig. 1.
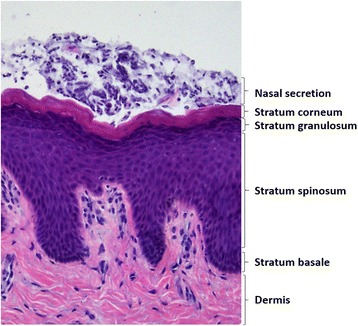
Representative histological view of epidermis in an S. aureus nasal carrier. Nasal secretion and four stratified cell layers in epidermis are marked. Sections were stained with hematoxylin-eosin, and examined in light microscope
Localization of S. aureus in epidermis and in nasal secretions
We detected S. aureus in both the upper and lower layers of epidermis by CLSM/IHC (Figs. 2, 3, 4, 5 and 6). S. aureus was most often found in stratum corneum (Fig. 2). We also found S. aureus in stratum spinosum (Figs. 3, 4 and 5), as far down as in stratum basale (Figs. 5 and 6), and on the border to dermis (Fig. 6b). Nasal secretion was observed in sections from both carriers and non−/intermittent carriers, and S. aureus was often observed in association with nasal secretion in pairs or clusters (Fig. 2b). S. aureus was often observed as single bacterial cells (Fig. 2a and c, Figs. 3, 4, 5 and 6) or in pairs (Fig. 2b and d) in the epidermal layers. Gram-staining confirmed the presence of Gram-positive cocci in the epidermis of both carriers and non−/intermittent carriers (results not shown).
Fig. 2.
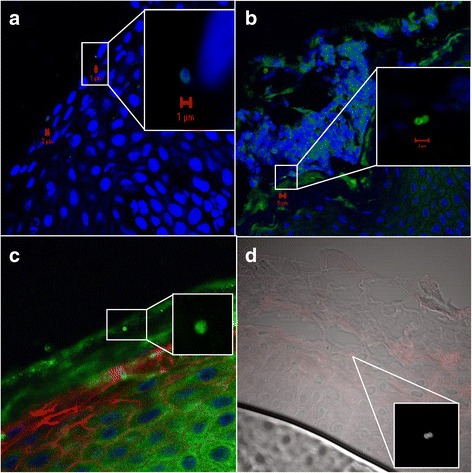
Localization of S. aureus in the upper part of epidermis. Inset represents a zoomed portion of the image. Scale bar is in micrometers. We used a confocal laser scanning microscopy (CLSM) ×63 objective. a-b S. aureus is localized on the border between stratum corneum and stratum granulosum. Green fluorescence labeling of S. aureus, blue fluorescence labeling of keratinocyte cell nuclei and S. aureus DNA. Primary antibody rabbit polyclonal antibody to S. aureus (ab20920, Abcam), secondary antibody Alexa Fluor 488® goat anti-rabbit IgG (Molecular Probe™, Thermo Fisher Scientific), and DRAQ5 (BioStatus). c S. aureus in stratum corneum. The same labeling is used as in a-b, but in addition, we used Alexa Fluor 594 Phalloidin (A12381, Molecular Probes™, Thermo Fisher Scientific) staining actin red. d Combined CLSM and light microscopy of nasal tissue section. S. aureus seen as red cocci in pairs, presumably on the border between stratum corneum and stratum granulosum. Primary S. aureus rabbit polyclonal antibody (ab20920, Abcam) and a secondary antibody Alexa Fluor 546 goat anti-rabbit IgG (Molecular Probes™, Thermo Fisher Scientific) were used
Fig. 3.
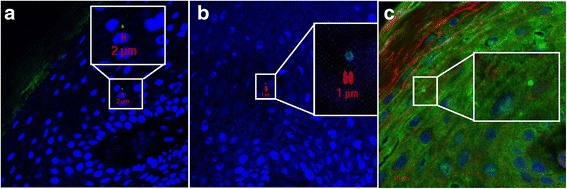
Localization of S. aureus in stratum spinosum by CLSM. Inset represents a zoomed portion of the image. Scale bar is in micrometers. We used a CLSM ×63 objective. The sections are oriented so that the outermost epidermis layer is shown in the upper left corner. a-b Green fluorescence labeling of S. aureus, blue fluorescence labeling of keratinocyte cell nuclei and S. aureus DNA. c Same labeling as in a and b, but in addition we used red fluorescence labeling of actin. Primary antibody rabbit polyclonal antibody to S. aureus (ab20920, Abcam), secondary antibody Alexa Fluor 488® goat anti-rabbit IgG (Molecular Probe™, Thermo Fisher Scientific), Alexa Fluor 594 Phalloidin (Molecular Probes™, Thermo Fisher Scientific) and DRAQ5 (BioStatus)
Fig. 4.
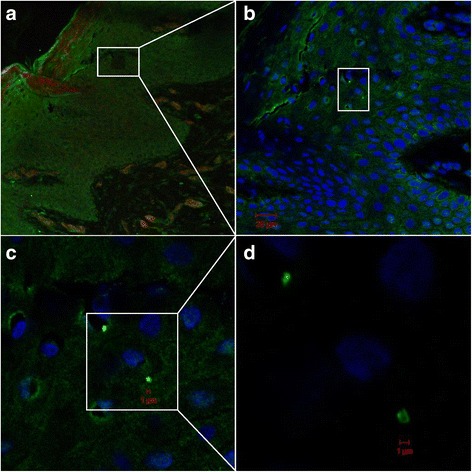
Confocal laser scanning microscopy of frozen section of nasal tissue from an S. aureus carrier. Overview (a-b) and detailed/zoomed portion of image (c-d) showing S. aureus in stratum spinosum. Scale bar is in micrometers. Green fluorescence labeling of S. aureus, blue fluorescence labeling of keratinocyte cell nuclei and S. aureus DNA, red fluorescence labeling of actin. Primary antibody rabbit polyclonal antibody to S. aureus (ab20920, Abcam), secondary antibody Alexa Fluor 488® goat anti-rabbit IgG (Molecular Probe™, Thermo Fisher Scientific), Alexa Fluor 594 Phalloidin (Molecular Probes™, Thermo Fisher Scientific) and DRAQ5 (BioStatus). We used a CLSM ×63 objective
Fig. 5.
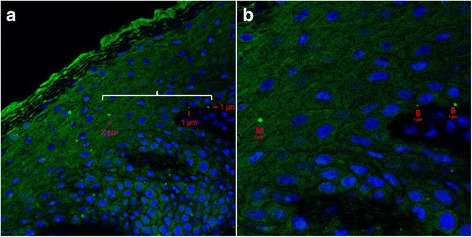
Localization of S. aureus on the border between stratum spinosum and stratum basale. Overview (a) and detailed image (b) of nasal tissue from an S. aureus carrier by confocal laser scanning microscopy. Scale bar is in micrometers. White brace indicates the region which is magnified in (b). Green fluorescence labeling of S. aureus, blue fluorescence labeling of keratinocyte cell nuclei and S. aureus DNA. Primary antibody rabbit polyclonal antibody to S. aureus (ab20920, Abcam), secondary antibody Alexa Fluor 488® goat anti-rabbit IgG (Molecular Probe™, Thermo Fisher Scientific), and DRAQ5 (BioStatus). We used a CLSM ×63 objective
Fig. 6.
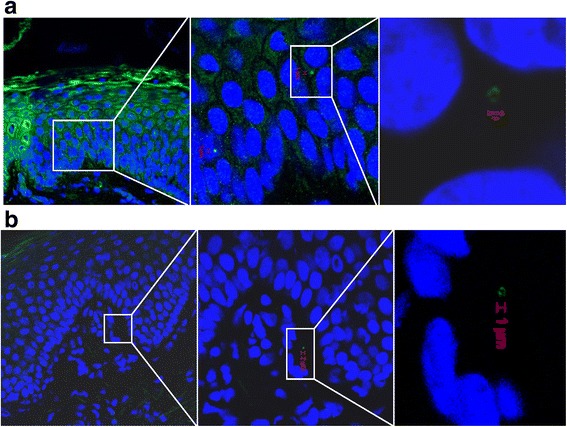
Localization of S. aureus in the lower part of epidermis by CLSM. Overview and detailed image/zoomed portion of the image showing (a) S. aureus on the border between stratum spinosum and stratum basale, and (b) S. aureus on the border between the stratum basale and dermis. Inset represents a zoomed portion of the image. Scale bar is in micrometers. Green fluorescence labeling of S. aureus, blue fluorescence labeling of keratinocyte cell nuclei and S. aureus DNA. Primary antibody rabbit polyclonal antibody to S. aureus (ab20920, Abcam), secondary antibody Alexa Fluor 488® goat anti-rabbit IgG (Molecular Probe™, Thermo Fisher Scientific), and DRAQ5 (BioStatus). We used a CLSM ×63 objective
Intracellular localization of S. aureus was assessed by CLSM/IHC (Fig. 7) using specific antibodies to S. aureus and staining of different structures (nucleus and actin). Three-dimensional reconstruction of highly magnified z-stacks was possible using CLSM. Consecutive z-plane images (Z-scan) showed fluorescent S. aureus (green) in close proximity/closely localized to the cellular nuclei of epithelial cells (blue) (Fig. 7).
Fig. 7.
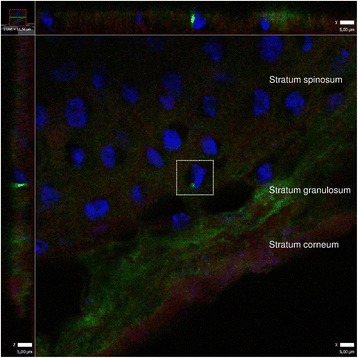
Intracellular localization of S. aureus in nasal epithelial cells. S. aureus is labeled with primary rabbit polyclonal antibody to S. aureus (Abcam), secondary antibody Alexa Fluor 488 goat anti-rabbit IgG (green) (Molecular Probes™, Thermo Fisher Scientific), DRAQ5 (BioStatus) for keratinocyte nuclei (blue) and Alexa Fluor 594 Phalloidin (A12381, Molecular Probes™; Thermo Fisher Scientific) for actin (red). Confocal laser scanning microscopy of frozen sections. Projection is constructed from confocal Z-stacks (0,2 um thick), 63× objective. Image to the left and on top corresponds to a vertical view in the z-plane. Z-plane images reveal a single cellular nucleus (blue) closely related to fluorescing S. aureus (green)
Among the 39 biopsy donors, we selected 14 donors randomly for calculating the number of S. aureus per tissue section by CLSM/IHC. The counting was performed by blinding the sample numbers and the examiner was unaware whether a carrier or non−/intermittent carrier was examined when performing the confocal microscopy. On average, there were 0.61 S. aureus per tissue section among all the 14 selected individuals in our study. Of note, S. aureus was detected in sections from three of the six selected non−/intermittent carriers, while no S. aureus was detected in the blinded sections from three out of eight S. aureus carriers.
Discussion
In the present study, we have assessed the S. aureus localization within vestibulum nasi in healthy individuals. By using a confocal scanning laser microscopy approach, we found that S. aureus is localised in both the upper and lower layers of epidermis within the nasal epithelium of healthy individuals. We also observed intracellular localization of S. aureus in the different layers of epidermis.
As expected, we identified the majority of S. aureus in the stratum corneum. In artificially colonized cotton rats, localization of S. aureus has been shown in the stratum corneum of intranasal squamous epithelium [24] where S. aureus appeared as single cells or diplococci, a finding that is corroborated by our study. This is also in accordance with other studies showing that adherence of S. aureus to nasal cells is directly related with the age of the epithelial cells. The most differentiated cells bind most S. aureus, while the youngest cells (less differentiated keratinocytes in the lower epidermis layer) bind fewer S. aureus [40]. Identical findings have been shown for S. epidermidis [41].
A histological study revealed S. aureus in the stratified squamous epithelium and in hair follicles in vestibulum nasi post mortem [42]. The study suggested that there is a difference between persistent and intermittent S. aureus carriers concerning niche localization, and difficulties in decolonization might be related to S. aureus confined to hair follicles and subsequent re-colonization. We took samples from regions with nasal hair in only six donors, but unfortunately we did not observe hair follicles in any of these samples.
The number of S. aureus captured from swabbing of the nasal vestibule is quite low [1], but the number of S. aureus in the nostrils is in accordance with other studies [1, 26, 43]. As expected, the low density of S. aureus was confirmed by the microscopic analyses. Normally, there are higher bacterial densities in diseased tissue compared with tissue from healthy carriers [16]. A previous study using confocal microscopy on tissue sections from rhinosinusitis patients, found that approximately 1/3 of S. aureus infected cells (regardless of cell type) carried more than 10 bacteria [16]. We confirmed our IHC/fluorescence staining protocol by using a skin tissue section from a patient with confirmed S. aureus skin infection as positive control (results not shown). In this positive control, we observed large clusters of S. aureus in both upper and lower layers of epidermis. Our findings show a clear difference between S. aureus colonization in healthy and inflamed vestibular skin, as the density of S. aureus was low in the healthy volunteers in our study.
The squames (stratum corneum) is constantly being shed from the skin surface, and shedding contributes to the clearing of attached bacteria [44]. This might explain why we saw only a few S. aureus in the nasal tissue sections. In many of our tissue sections, the stratum corneum was detached from the epithelial layer, and was visible as loose appendages in the proximity of the epithelium.
One important deviation in our study when defining colonization status, is that when we screen for S. aureus, we normally swab both nostrils [45], but in this study we only swabbed one of the nostrils and harvested our histologic sample from the other nostril. The reason for this was that we wanted to keep an intact skin structure and not alter the skin surface before biopsy sampling. It has previously been shown that carriers are more likely to carry S. aureus in one nostril than in both nostrils [46], and this could perhaps explain the cases where we detected S. aureus by swab culture, but not by CLSM of biopsied nasal tissue from the same individual, or vice versa.
We observed a switch of S. aureus colonization status in the period between the Tromsø 6 study and our study in some of the participants. Three individuals defined as non−/intermittent carriers in the Tromsø 6 study, were negative for S. aureus by nasal swab sampling and positive for S. aureus by CLSM of nasal epithelium in the present study. We also observed the opposite, i.e. four individuals defined as S. aureus carriers in the Tromsø 6 study were S. aureus negative in our study by both swabbing and CLSM. A change in colonization status of some individuals was expected [39]. Another expected observation was that some persistent carriers exhibited different spa types in the two studies. These observations are in line with earlier reports [39, 47], and could be examples of strain replacement or co-colonization.
Intracellular persistence of S. aureus in connection with infection, e.g. recurrent sinusitis, has been shown in previous studies [15, 16]. We confirmed intracellular S. aureus localization in the stratum spinosum in healthy individuals by CLSM images at multiple z-planes within the nasal tissue by using specific antibodies. We did not perform a systematic investigation of intracellular localization and it was only performed in a few randomly selected donors. S. aureus can live as a commensal within humans without causing overt disease, and there is considerable evidence of intracellular localization of S. aureus [16, 34, 35] in neutrophils [48], osteoblasts [49], nasal epithelial cells [16, 50], and endothelial cells [51]. However, we did not expect to find intracellular S. aureus in nasal tissue from healthy individuals.
The intracellular localization of S. aureus has been suggested to provide a sanctuary for the bacteria, protecting them from the innate host defense system, and perhaps also from antimicrobial agents [16], hence serving as a reservoir for chronic or relapsing infection. Some of the participants in our study were negative by S. aureus nasal swabbing, but positive for S. aureus by CLSM indicating that intracellular localization of S. aureus could be difficult to detect by nasal swabbing. A possible explanation for the discrepancy could be intracellular localization or it could be due to a single nostril S. aureus carrier status.
Our work shows that S. aureus is primarily observed in the debris overlying the nasal epithelium, whereas single staphylococci can also be observed in deeper intraepithelial layers. We observed nasal secretions in most of the tissue samples from both carriers and non−/intermittent carriers. Clusters of S. aureus were very often localized within the nasal secretions in carriers, and especially in two of our donors. One could hypothesize that staphylococci are in a dormant state in their intraepithelial localization. In the healthy nasal vestibule, the stratum corneum is not surveilled by the dendritic cells [52], which could mean that staphylococci might remain undetected by the body’s immune system. In addition, nasal crust formation could be considered a natural growth medium for staphylococci, leading to colony formation under the right host conditions.
Conclusions
Our results suggest that S. aureus colonize both the upper and lower layers of the epidermis within the nasal epithelium of healthy individuals. The number of S. aureus in epidermis was surprisingly low. Intracellular localization of S. aureus in nasal tissue from healthy individuals was also detected. The intracellular state may protect the bacteria from traditional decolonization procedures and cause problems with re-colonization. Thus, decolonization strategies targeting intracellular S. aureus should be considered for future testing. Much is still unknown about S. aureus persistence in nasal tissue. Knowledge of the exact localization of S. aureus in nasal tissue will be important for the understanding of the host responses against S. aureus. Further work on the identification of bacterial and host factors can provide us with tools for targeted prevention of S. aureus colonisation and infection.
Acknowledgements
We acknowledge Department for Clinical Pathology, University Hospital of North Norway, Tromsø, and Marit N. Nilsen, Institute of Medical Biology, UiT- the Arctic University of Norway, for performing the cryo-sectioning. Appreciated is also MD Rune Flaa, University Hospital in North Norway, Tromsø, Norway, for assisting in collecting the nasal tissue biopsies. Acknowledged are also all the Tromsø 6 participants who volunteered and donated nasal tissue samples for this study.
Funding
This work was supported by grants from The Norwegian Research Council (NFR) 191,264 (2009–13); The Northern Norway Regional Health Authority (Helse Nord RHF) projects SFP877–09 and Miljøstøtte MIL963–10 (2010–2012). The funding bodies represented by Johanna Ulrica Ericson Sollid have taken part in design of the study and collection, analysis, and interpretation of data and in writing the manuscript. The publication charges for this article have been funded by a grant from the publication fund of UiT The Arctic University of Norway.
Availability of data and materials
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Authors’ contributions
AMH designed experiments, performed the microscopic analyses, analyzed results and prepared the manuscript. BK designed and performed most of the experiments, and reviewed the manuscript. NCS assisted in design of the experiments, collected the nasal vestibular biopsies, prepared and reviewed the manuscript. ASF, MJ and SF assisted in design of experiments and reviewed the manuscript. RSO performed most of the experiments and reviewed the manuscript. JUES designed the study, analyzed results and prepared the manuscript. All authors gave final approval of the version to be published.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
All authors read the manuscript and have provided their consent to publish. All authors declare no conflict of interests. See ethics approval and consent to participate.
Ethics approval and consent to participate
This work was performed in compliance with the ethical guidelines established by UiT- The Arctic University of Norway. Each participant gave written informed consent prior to the tissue sampling. Subject recruitment, enrollment and sampling of nasal tissue samples have been approved by the Regional Committee for Medical and Health Research Ethics, Rec North, Norway (Document reference 2010/146–7).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- ATCC
American type culture collection
- CFU
Colony forming unit
- ClfB
Clumping factor B
- CLSM
Confocal laser scanning microscopy
- IsdA
Iron-regulated surface determinant
- MRSA
Methicillin-resistant Staphylococcus aureus
- MSCRAMMs
Microbial surface components recognizing adhesive matrix molecules
- PBS
Phosphate buffered saline
- SdrD
Serine-aspartate repeat protein D
- Spa
Staphylococcal protein A
- UNN
University hospital of North Norway
- WTA
Wall teichoic acid
Additional file
Results from spa-typing in the previous Tromsø 6 study and the present study. Additional documentation. (PDF 91 kb)
Contributor Information
Anne-Merethe Hanssen, Phone: +47 776 60749, Email: anne-merethe.hanssen@uit.no.
Bert Kindlund, Email: bert.k@spray.se.
Niels Christian Stenklev, Email: Niels.Christian.Stenklev@unn.no.
Anne-Sofie Furberg, Email: anne-sofie.furberg@uit.no.
Silje Fismen, Email: Silje.Fismen@unn.no.
Renate Slind Olsen, Email: renate.slind.olsen@rjl.se.
Mona Johannessen, Email: mona.johannessen@uit.no.
Johanna Ulrica Ericson Sollid, Email: johanna.e.sollid@uit.no.
References
- 1.Cole AM, Tahk S, Oren A, Yoshioka D, Kim YH, Park A, Ganz T. Determinants of Staphylococcus aureus nasal carriage. Clin Diagn Lab Immunol. 2001;8:1064–1069. doi: 10.1128/CDLI.8.6.1064-1069.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.VandenBergh MF, Verbrugh HA. Carriage of Staphylococcus aureus: epidemiology and clinical relevance. J Lab Clin Med. 1999;133:525–534. doi: 10.1016/S0022-2143(99)90181-6. [DOI] [PubMed] [Google Scholar]
- 3.Kluytmans JA, Wertheim HF. Nasal carriage of Staphylococcus aureus and prevention of nosocomial infections. Infection. 2005;33:3–8. doi: 10.1007/s15010-005-4012-9. [DOI] [PubMed] [Google Scholar]
- 4.van Belkum A, Verkaik NJ, de Vogel CP, Boelens HA, Verveer J, Nouwen JL, Verbrugh HA, Wertheim HF. Reclassification of Staphylococcus aureus nasal carriage types. J Infect Dis. 2009;199:1820–1826. doi: 10.1086/599119. [DOI] [PubMed] [Google Scholar]
- 5.Wertheim HF, Vos MC, Ott A, van Belkum A, Voss A, Kluytmans JA, van Keulen PH, Vandenbroucke-Grauls CM, Meester MH, Verbrugh HA. Risk and outcome of nosocomial Staphylococcus aureus bacteraemia in nasal carriers versus non-carriers. Lancet. 2004;364:703–705. doi: 10.1016/S0140-6736(04)16897-9. [DOI] [PubMed] [Google Scholar]
- 6.Mertz D, Frei R, Periat N, Zimmerli M, Battegay M, Fluckiger U, Widmer AF. Exclusive Staphylococcus aureus throat carriage: at-risk populations. Arch Intern Med. 2009;169:172–178. doi: 10.1001/archinternmed.2008.536. [DOI] [PubMed] [Google Scholar]
- 7.von Eiff C, Becker K, Machka K, Stammer H, Peters G. Nasal carriage as a source of Staphylococcus aureus bacteremia. Study group. N Engl J Med. 2001;344:11–16. doi: 10.1056/NEJM200101043440102. [DOI] [PubMed] [Google Scholar]
- 8.Coates T, Bax R, Coates A. Nasal decolonization of Staphylococcus aureus with mupirocin: strengths, weaknesses and future prospects. J Antimicrob Chemother. 2009;64:9–15. doi: 10.1093/jac/dkp159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lee CJ, Sankaran S, Mukherjee DV, Apa ZL, Hafer CA, Wright L, Larson EL, Lowy FD. Staphylococcus aureus Oropharyngeal carriage in a prison population. Clin Infect Dis. 2011;52:775–778. doi: 10.1093/cid/cir026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kluytmans J, van Belkum A, Verbrugh H. Nasal carriage of Staphylococcus aureus: epidemiology, underlying mechanisms, and associated risks. Clin Microbiol Rev. 1997;10:505–520. doi: 10.1128/cmr.10.3.505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mody L, Kauffman CA, McNeil SA, Galecki AT, Bradley SF. Mupirocin-based decolonization of Staphylococcus aureus carriers in residents of 2 long-term care facilities: a randomized, double-blind, placebo-controlled trial. Clin Infect Dis. 2003;37:1467–1474. doi: 10.1086/379325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mehta MS, Hacek DM, Kufner BA, Price C, Peterson LR. Dose-ranging study to assess the application of intranasal 2% mupirocin calcium ointment to eradicate Staphylococcus aureus nasal colonization. Surg Infect. 2013;14:69–72. doi: 10.1089/sur.2012.086. [DOI] [PubMed] [Google Scholar]
- 13.Ammerlaan HS, Kluytmans JA, Berkhout H, Buiting A, de Brauwer EI, van den Broek PJ, van Gelderen P, Leenders SA, Ott A, Richter C, Spanjaard L, Spijkerman IJ, van Tiel FH, Voorn GP, Wulf MW, van ZJ, Troelstra A, Bonten MJ. Eradication of carriage with methicillin-resistant Staphylococcus aureus: determinants of treatment failure. J Antimicrob Chemother. 2011;66:2418–2424. doi: 10.1093/jac/dkr250. [DOI] [PubMed] [Google Scholar]
- 14.Gilpin DF, Small S, Bakkshi S, Kearney MP, Cardwell C, Tunney MM. Efficacy of a standard methicillin-resistant Staphylococcus aureus decolonisation protocol in routine clinical practice. J Hosp Infect. 2010;75:93–98. doi: 10.1016/j.jhin.2009.11.025. [DOI] [PubMed] [Google Scholar]
- 15.Plouin-Gaudon I, Clement S, Huggler E, Chaponnier C, Francois P, Lew D, Schrenzel J, Vaudaux P, Lacroix JS. Intracellular residency is frequently associated with recurrent Staphylococcus aureus rhinosinusitis. Rhinology. 2006;44:249–254. [PubMed] [Google Scholar]
- 16.Clement S, Vaudaux P, Francois P, Schrenzel J, Huggler E, Kampf S, Chaponnier C, Lew D, Lacroix JS. Evidence of an intracellular reservoir in the nasal mucosa of patients with recurrent Staphylococcus aureus rhinosinusitis. J Infect Dis. 2005;192:1023–1028. doi: 10.1086/432735. [DOI] [PubMed] [Google Scholar]
- 17.Weidenmaier C, Goerke C, Wolz C. Staphylococcus aureus Determinants for nasal colonization. Trends Microbiol. 2012;20:243–250. doi: 10.1016/j.tim.2012.03.004. [DOI] [PubMed] [Google Scholar]
- 18.Ramakrishnan VR, Feazel LM, Gitomer SA, Ir D, Robertson CE, Frank DN. The microbiome of the middle meatus in healthy adults. PLoS One. 2013;8:e85507. doi: 10.1371/journal.pone.0085507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.SanMiguel A, Grice EA. Interactions between host factors and the skin microbiome. Cell Mol Life Sci. 2015;72:1499–1515. doi: 10.1007/s00018-014-1812-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fritsch H, Kuehnel W. Color atlas of human anatomy; Vol. 2.; internal organs.; p.406. 5th edition. Stuttgart: Georg Thieme Verlag; 2008. [Google Scholar]
- 21.Nestle FO, Di Meglio P, Qin JZ, Nickoloff BJ. Skin immune sentinels in health and disease. Nat Rev Immunol. 2009;9:679–691. doi: 10.1038/nri2622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Grice EA, Segre JA. The skin microbiome. Nat Rev Microbiol. 2011;9:244–253. doi: 10.1038/nrmicro2537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Krishna S, Miller LS. Host-pathogen interactions between the skin and Staphylococcus aureus. Curr Opin Microbiol. 2012;15:28–35. doi: 10.1016/j.mib.2011.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Burian M, Rautenberg M, Kohler T, Fritz M, Krismer B, Unger C, Hoffmann WH, Peschel A, Wolz C, Goerke C. Temporal expression of adhesion factors and activity of global regulators during establishment of Staphylococcus aureus nasal colonization. J Infect Dis. 2010;201:1414–1421. doi: 10.1086/651619. [DOI] [PubMed] [Google Scholar]
- 25.Weidenmaier C, Kokai-Kun JF, Kulauzovic E, Kohler T, Thumm G, Stoll H, Gotz F, Peschel A. Differential roles of sortase-anchored surface proteins and wall teichoic acid in Staphylococcus aureus nasal colonization. Int J Med Microbiol. 2008;298:505–513. doi: 10.1016/j.ijmm.2007.11.006. [DOI] [PubMed] [Google Scholar]
- 26.Burian M, Wolz C, Goerke C. Regulatory adaptation of Staphylococcus aureus during nasal colonization of humans. PLoS One. 2010;5:e10040. doi: 10.1371/journal.pone.0010040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.O'Brien LM, Walsh EJ, Massey RC, Peacock SJ, Foster TJ. Staphylococcus aureus Clumping factor B (ClfB) promotes adherence to human type I cytokeratin 10: implications for nasal colonization. Cell Microbiol. 2002;4:759–770. doi: 10.1046/j.1462-5822.2002.00231.x. [DOI] [PubMed] [Google Scholar]
- 28.Clarke SR, Brummell KJ, Horsburgh MJ, McDowell PW, Mohamad SA, Stapleton MR, Acevedo J, Read RC, Day NP, Peacock SJ, Mond JJ, Kokai-Kun JF, Foster SJ. Identification of in vivo-expressed antigens of Staphylococcus aureus and their use in vaccinations for protection against nasal carriage. J Infect Dis. 2006;193:1098–1108. doi: 10.1086/501471. [DOI] [PubMed] [Google Scholar]
- 29.Wertheim HF, Walsh E, Choudhurry R, Melles DC, Boelens HA, Miajlovic H, Verbrugh HA, Foster T, van Belkum A. Key role for clumping factor B in Staphylococcus aureus nasal colonization of humans. PLoS Med. 2008;5:e17. doi: 10.1371/journal.pmed.0050017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Clarke SR, Andre G, Walsh EJ, Dufrene YF, Foster TJ, Foster SJ. Iron-regulated surface determinant protein a mediates adhesion of Staphylococcus aureus to human corneocyte envelope proteins. Infect Immun. 2009;77:2408–2416. doi: 10.1128/IAI.01304-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Aly R, Shinefield HI, Strauss WG, Maibach HI. Bacterial adherence to nasal mucosal cells. Infect Immun. 1977;17:546–549. doi: 10.1128/iai.17.3.546-549.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Foster TJ, Geoghegan JA, Ganesh VK, Hook M. Adhesion, invasion and evasion: the many functions of the surface proteins of Staphylococcus aureus. Nat Rev Microbiol. 2014;12:49–62. doi: 10.1038/nrmicro3161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Askarian F, Ajayi C, Hanssen AM, van Sorge NM, Pettersen I, Diep DB, Sollid JU, Johannessen M. The interaction between Staphylococcus aureus SdrD and desmoglein 1 is important for adhesion to host cells. Sci Rep. 2016;6:22134. doi: 10.1038/srep22134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Garzoni C, Kelley WL. Staphylococcus aureus: new evidence for intracellular persistence. Trends Microbiol. 2009;17:59–65. doi: 10.1016/j.tim.2008.11.005. [DOI] [PubMed] [Google Scholar]
- 35.Garzoni C, Francois P, Huyghe A, Couzinet S, Tapparel C, Charbonnier Y, Renzoni A, Lucchini S, Lew DP, Vaudaux P, Kelley WL, Schrenzel J. A global view of Staphylococcus aureus whole genome expression upon internalization in human epithelial cells. BMC Genomics. 2007;8:171. doi: 10.1186/1471-2164-8-171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zautner AE, Krause M, Stropahl G, Holtfreter S, Frickmann H, Maletzki C, Kreikemeyer B, Pau HW, Podbielski A. Intracellular persisting Staphylococcus aureus is the major pathogen in recurrent tonsillitis. PLoS One. 2010;5:e9452. doi: 10.1371/journal.pone.0009452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ellington JK, Harris M, Webb L, Smith B, Smith T, Tan K, Hudson M. Intracellular Staphylococcus aureus. A mechanism for the indolence of osteomyelitis. J Bone Joint Surg Br. 2003;85:918–921. [PubMed] [Google Scholar]
- 38.Jacobsen BK, Eggen AE, Mathiesen EB, Wilsgaard T, Njolstad I. Cohort profile: the Tromso study. Int J Epidemiol. 2012;41:961–967. doi: 10.1093/ije/dyr049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sangvik M, Olsen RS, Olsen K, Simonsen GS, Furberg AS, Sollid JU. Age- and gender-associated Staphylococcus aureus spa types found among nasal carriers in a general population: the Tromso staph and skin study. J Clin Microbiol. 2011;49:4213–4218. doi: 10.1128/JCM.05290-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bibel DJ, Aly R, Shinefield HR, Maibach HI, Strauss WG. Importance of the keratinized epithelial cell in bacterial adherence. J Invest Dermatol. 1982;79:250–253. doi: 10.1111/1523-1747.ep12500072. [DOI] [PubMed] [Google Scholar]
- 41.Brooker BE, Fuller R. The adhesion of coagulase negative staphylococci to human skin and its relevance to the bacterial flora of milk. J Appl Bacteriol. 1984;57:325–332. doi: 10.1111/j.1365-2672.1984.tb01397.x. [DOI] [PubMed] [Google Scholar]
- 42.Ten Broeke-Smits NJ, Kummer JA, Bleys RL, Fluit AC, Boel CH. Hair follicles as a niche of Staphylococcus aureus in the nose; is a more effective decolonisation strategy needed? J Hosp Infect. 2010;76:211–214. doi: 10.1016/j.jhin.2010.07.011. [DOI] [PubMed] [Google Scholar]
- 43.Nouwen JL, Ott A, Kluytmans-Vandenbergh MF, Boelens HA, Hofman A, van Belkum A, Verbrugh HA. Predicting the Staphylococcus aureus nasal carrier state: derivation and validation of a “culture rule”. Clin Infect Dis. 2004;39:806–811. doi: 10.1086/423376. [DOI] [PubMed] [Google Scholar]
- 44.Margadant C, Charafeddine RA, Sonnenberg A. Unique and redundant functions of integrins in the epidermis. FASEB J. 2010;24:4133–4152. doi: 10.1096/fj.09-151449. [DOI] [PubMed] [Google Scholar]
- 45.Olsen K, Falch BM, Danielsen K, Johannessen M, Ericson Sollid JU, Thune I, Grimnes G, Jorde R, Simonsen GS, Furberg AS. Staphylococcus aureus Nasal carriage is associated with serum 25-hydroxyvitamin D levels, gender and smoking status. The Tromso staph and skin study. Eur J Clin Microbiol Infect Dis. 2012;31:465–473. doi: 10.1007/s10096-011-1331-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kildow BJ, Conradie JP, Robson RL. Nostrils of healthy volunteers are independent with regard to Staphylococcus aureus carriage. J Clin Microbiol. 2012;50:3744–3746. doi: 10.1128/JCM.01488-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sakwinska O, Blanc DS, Lazor-Blanchet C, Moreillon M, Giddey M, Moreillon P. Ecological temporal stability of Staphylococcus aureus nasal carriage. J Clin Microbiol. 2010;48:2724–2728. doi: 10.1128/JCM.02091-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Gresham HD, Lowrance JH, Caver TE, Wilson BS, Cheung AL, Lindberg FP. Survival of Staphylococcus aureus inside neutrophils contributes to infection. J Immunol. 2000;164:3713–3722. doi: 10.4049/jimmunol.164.7.3713. [DOI] [PubMed] [Google Scholar]
- 49.Ellington JK, Reilly SS, Ramp WK, Smeltzer MS, Kellam JF, Hudson MC. Mechanisms of Staphylococcus aureus invasion of cultured osteoblasts. Microb Pathog. 1999;26:317–323. doi: 10.1006/mpat.1999.0272. [DOI] [PubMed] [Google Scholar]
- 50.Dziewanowska K, Patti JM, Deobald CF, Bayles KW, Trumble WR, Bohach GA. Fibronectin binding protein and host cell tyrosine kinase are required for internalization of Staphylococcus aureus by epithelial cells. Infect Immun. 1999;67:4673–4678. doi: 10.1128/iai.67.9.4673-4678.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Menzies BE, Kourteva I. Internalization of Staphylococcus aureus by endothelial cells induces apoptosis. Infect Immun. 1998;66:5994–5998. doi: 10.1128/iai.66.12.5994-5998.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Simpson CL, Patel DM, Green KJ. Deconstructing the skin: cytoarchitectural determinants of epidermal morphogenesis. Nat Rev Mol Cell Biol. 2011;12:565–580. doi: 10.1038/nrm3175. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.