

Abstract
Fatal Familial Insomnia (FFI) is a genetic prion disease caused by a point mutation in the prion protein gene (PRNP) characterized by prominent thalamic atrophy, diffuse astrogliosis and moderate deposition of PrPSc in the brain. Here, for the first time, we demonstrate that the olfactory mucosa (OM) of patients with FFI contains trace amount of PrPSc detectable by PMCA and RT-QuIC. Quantitative PMCA analysis estimated a PrPSc concentration of about 1 × 10−14 g/ml. In contrast, PrPSc was not detected in OM samples from healthy controls and patients affected by other neurodegenerative disorders, including Alzheimer’s disease, Parkinson’s disease and frontotemporal dementia. These results indicate that the detection limit of these assays is in the order of a single PrPSc oligomer/molecule with a specificity of 100%.
Transmissible Spongiform Encephalopathies (TSEs) are a group of fatal and incurable neurodegenerative diseases that might present as sporadic, acquired or genetic conditions. The most common form of sporadic TSE is represented by the Creutzfeldt-Jakob disease (CJD)1,2. Acquired forms include variant CJD (vCJD) which is linked to the consumption of cattle infected with bovine spongiform encephalopathy (BSE)3,4. Genetic forms are associated to mutations and insertions in the PRNP gene encoding the cellular prion protein (PrPC)5. Fatal Familial Insomnia (FFI) is caused by a point mutation at codon 178 of PRNP resulting in aspartic acid to asparagine substitution in coupling phase with methionine at polymorphic position 129 (D178N/129M)6. FFI is associated with sleep alterations, autonomic dysfunction, loss of weight and motor signs7. After symptoms onset, the disease usually leads to death within 2 years and no treatments are available at present8.
Although characterized by a great heterogeneity of clinical and neuropathological phenotypes, these diseases share a common infectious agent which is a conformationally altered form of the PrPC, known as PrPSc, which is able to induce the conformational conversion of PrPC into new PrPSc. Compared to PrPC, PrPSc has higher content in β-sheet structures, is insoluble in mild detergents and partially resistant to protease degradation. Thus, after digestion with proteinase K (PK), PrPC is completely digested while PrPSc shows a PK resistant core (27–30 kDa)9. Mutations in the PRNP gene might favor the process of PrPC misfolding and aggregation thus favoring the formation of PrPSc.
Both PrPC and PrPSc can be variably glycosylated and give rise to diglycosylated, monoglycosylated and unglycosylated species10. After PK digestion, only the unglycosylated band can migrate at around 21 kDa (type 1) or 19 kDa (type 2)1. The relative abundance of each PrP glycoform (glycoform ratio) and the electrophoretic mobility of the unglycosylated form enable a biochemical classification of different prion strains. For instance, the sporadic forms of CJD are generally characterized by a predominance of the monoglycosylated PrP band, with unglycosylated PrP migrating at 21 kDa (sCJD-type1) or 19 kDa (sCJD-type2). BSE, vCJD and FFI are characterized by an abundance of the diglycosylated band with the unglycosylated one migrating at 19 kDa (type 2)3,8,11.
Recently, two innovative assays named Protein Misfolding Cyclic Amplification (PMCA) and Real Time Quaking-Induced Conversion (RT-QuIC) were generated to model the process of prion misfolding in vitro in an accelerated manner. PMCA12 consists of cycles of incubation and sonication of samples containing small amount of PrPSc in the presence of an excess of PrPC. This enables the exponential amplification of PrPSc, and can begin the reaction with the equivalent of a single molecule of PrPSc, which after amplification can give rise to billions of PrPSc molecules13. Therefore, PMCA analysis allowed to detect extremely low levels of PrPSc in urine14 and blood15,16 of patients with vCJD as well as in blood of sheep and primates experimentally infected with vCJD in preclinical stages17. RT-QuIC18 alternates phases of incubation to phases of vigorous shaking where soluble recombinant prion proteins (recPrP) are used as substrate for detecting femtomolar amounts of PrPSc. The reaction is monitored by Thioflavin-T (ThT) fluorescence and enabled detection of PrP seeds in the cerebrospinal fluid (CSF) of patients with sporadic CJD (sCJD) or genetic forms of prion disease (e.g. GSS and FFI) either in the symptomatic19 or pre-symptomatic stage of the disease20,21. RT-QuIC has been further adapted to detect PrPSc in other tissues, such as the olfactory mucosa (OM) of patients suffering from sporadic CJD22. Taken together, these findings suggest that RT-QuIC and PMCA have a huge potential to detect trace-amount of PrPSc (≥1 femtogram) in peripheral tissues.
By taking advantage of recent observations23,24,25, we have optimized PMCA and RT-QuIC and extended their use for the analysis of OM samples collected from two patients with FFI, 16 patients with other neurodegenerative disorders including Alzheimer’s disease (AD), Parkinson’s disease (PD), Frontotemporal Dementia (FTD) and 10 healthy control (HC). Our results showed that both techniques were able to detect PrPSc in FFI-OM samples in the order of a single molecule with 100% specificity.
Results
PMCA and RT-QuIC efficiently detect PrPSc in serial dilutions of FFI brain homogenates
To assess whether PrPSc associated to FFI (FFI-PrPSc) was able to amplify by means of PMCA, we have performed spiking experiment diluting FFI brain homogenate (from 10−5 to 10−12 dilution of the brain) into 10% healthy bank vole brain homogenates (BvBH) carrying methionine at position 109 of the prion protein (M109). After one round of amplification, we could efficiently detect up to a 10−9 dilution while, after two rounds, we were able to amplify all dilutions (Fig. 1A). Similarly, dilutions of FFI brain homogenate were prepared in PBS and analyzed by means of RT-QuIC. The experiments were carried out using recombinant truncated form of the bank vole PrP (BvPrP(90-231)) as substrate and were seeded in triplicate with 2 μL of each dilution (Fig. 1B). All reactions were performed at least three times by different operators. We could detect up to 10−9 dilution of PrPSc, while we did not efficiently detect any signal from higher dilutions. In particular, lower dilutions of brain homogenate induced a faster aggregation of BvPrP(90-231) (at around 8 hours), while intermediate dilutions induced aggregation after 14 hours. A sample was considered positive when the mean of the highest two fluorescence values (AU) of the replicates was higher than 10.000 AU and at least two out of three replicates crossed this value before 30 hours (see materials and methods for details). Samples that did not cross the threshold before 30 hours were considered negative. Brain homogenates of patient with AD and FTD were used as control and induced aggregation of BvPrP(90-231) after 30 hours, thus were considered negative (Fig. 1B).
Figure 1. Serial dilutions of FFI brain homogenate (D178N/129M) were analyzed by means of PMCA and RT-QuIC for estimating their ability to detect PrPSc.

(A) Ten % bank vole brain homogenates (BvBH) were spiked with serial dilution of FFI brain homogenate (from 10−5 to 10−12). The signal of PrPSc was assessed by means of Western blotting, after PK digestion, with the 6D11 antibody. Mw refers to the molecular weight. PrPC refers to normal bank vole brain homogenate not digested with PK. (B) Serial dilutions of FFI brain homogenate were analyzed by means of RT-QuIC reactions using BvPrP (90-231) as substrate. Average ThT fluorescence were plotted against time. AD brain homogenate (dilution 10−5) was used as negative control. BvPrP refers to unseeded recombinant protein.
PMCA and RT-QuIC can detect PrPSc from samples of olfactory mucosa
OM samples (n = 28) collected from patients with FFI (n = 2), AD (n = 6), PD (n = 6), FTD (n = 4) and HC (n = 10) were blindly analyzed by means of PMCA and RT-QuIC. After three rounds of PMCA, we could detect PrPSc only in FFI-OM samples (Fig. 2A). All the other samples remained negative even after 7 rounds of amplification (Supplementary Fig. 1). Similarly, through RT-QuIC analysis we could detect prion seeding activity in both FFI-OM samples, while PrPSc was never detected in any of the OM belonging to the other patients analyzed (Fig. 2B). The final RT-QuIC reaction products were collected (at 30 hours) digested with PK and analyzed by means of Western blotting. As previously observed24, seeds collected from OM of patients with FFI, sporadic or iatrogenic forms of CJD were able to produce final RT-QuIC aggregates partially resistant to PK digestion and characterized by a molecular weight ranging from 14 to 18 kDa (Fig. 3A). On the contrary, OM samples collected from patients with PD, AD or healthy subjects did not produce any PK resistant aggregate. Brain homogenate of patients with FFI and iCJD were used as positive controls.
Figure 2. PrPSc detection in OM of FFI subjects using PMCA and RT-QuIC assays.

(A) Representative image of PMCA analysis of olfactory mucosa samples (n = 28) that were blindly performed. After three rounds of amplification PrPSc could be detected in two samples (FFI-OM#1 and FFI-OM#2) that belong to subjects that were symptomatic at the time of OM collection. 10−5 and 10−8 refer to dilutions of FFI brain homogenate in PMCA substrate that were used as positive control for reaction. The signal of PrPSc was assessed by means of Western blotting, after PK digestion, with the 6D11 antibody. Numbers in the right indicate the position of Mw. PrPC refers to normal bank vole brain homogenate that was not digested with PK. Dashed line indicates cropped images from separate gels. (B) RT-QuIC reactions were seeded with olfactory mucosa specimens of all patients. FFI brain homogenate (FFI-BH 10−8) was used as positive control for the reaction. Two OM samples were found positive before the threshold set at 30 hours. Average ThT fluorescence were plotted against time.
Figure 3. Amplified OM-PrPSc comparison with that of FFI or sCJD-type 1 and sCJD-type 2 brain homogenates.
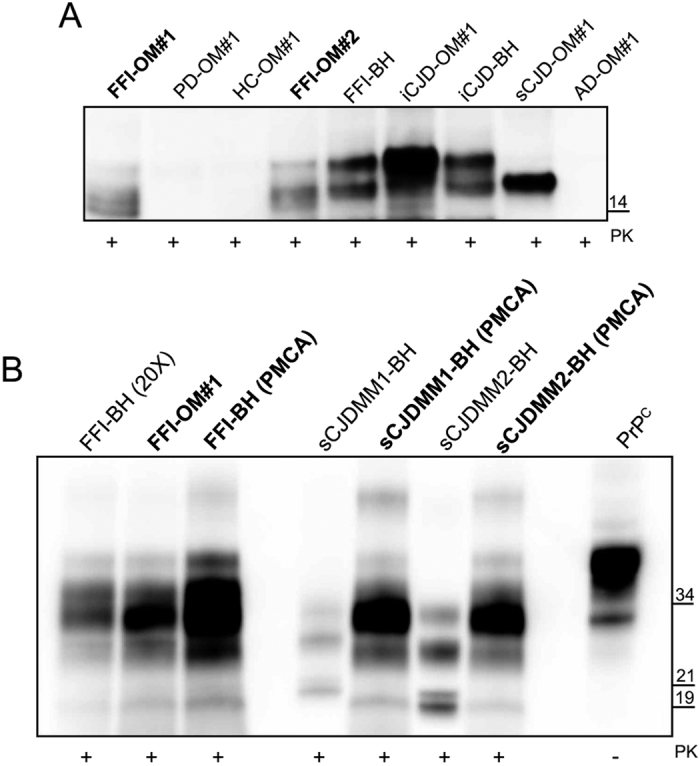
(A) Western blot of the final products of RT-QuIC reaction seeded with brain homogenates or olfactory mucosa from sCJD, iCJD, FFI, AD, PD patients and healthy controls (HC). Samples were digested with PK [50 μg/mL] and immunoblot was probed with the C-terminal antibody SAF-84. Number in the right indicates the position of Mw. (B) Biochemical profile of amplified products (both brain homogenates and olfactory mucosa) was compared to that of FFI, sCJD-type 1 and sCJD-type 2 brain homogenates assessed before and after PMCA. FFI brain homogenate was concentrated 20-fold for PrPSc detection. The signal of PrPSc was assessed by means of Western blotting, after PK digestion, with the 6D11 antibody. Numbers in the right indicate the position of Mw: 21 kDa refers to type 1 PrPSc, while 19 kDa refers to type 2 PrPSc.
Estimation of PrPSc concentration in the olfactory mucosa by means of quantitative PMCA (qPMCA)
To estimate the concentration of PrPSc present in the olfactory mucosa of an FFI subject, we have performed a quantitative PMCA, as previously described14. First, we have carefully estimated the concentration of PrPSc present in a FFI brain homogenate used to calibrate the PMCA reaction. The estimation was carried out by comparing the PrPSc signal by Western blot with a known concentration of human recombinant PrP (Supplementary Fig. 2A). By this procedure we calculated that PrPSc concentration in the 10% brain homogenate of this patient was approximately 141 ng/ml. Thereafter, we have performed serial dilutions of this brain homogenate and used them for PrPSc detection after various rounds of PMCA. The results indicate that 1 round of PMCA enables to detect at around 10−10 to 10−11 g/ml of PrPSc. Two or more rounds of PMCA allowed detection of 10−13–10−14 g/ml of PrPSc (Supplementary Fig. 2B). Olfactory mucosa of FFI patient resulted positive after 2 rounds of PMCA. According to our estimation the olfactory mucosa of this subject has a PrPSc concentration of 1.41 × 10−14 g/ml.
Biochemical characterization of PMCA reaction products
The biochemical profile of the amplified PrPSc obtained from both FFI-OM samples was characterized by a predominance of the diglycosylated band of PrP with the unglycosylated one migrating at around 19 kDa1. As expected, PrPSc obtained from FFI brain homogenate showed a similar biochemical profile that did not change after amplification by PMCA (Fig. 3B)26. This pattern was then compared to that of PrPSc sCJD-type 1 or PrPSc sCJD-type 2 before and after PMCA. Although, sCJD brain homogenates showed their characteristic PrPSc biochemical profiles following PMCA the biochemical profile of both PrPSc changed and acquired a unique feature characterized by the prevalence of the diglycosylated band with the unglycosylated fragment migrating at around 19 kDa.
Discussion
RT-QuIC and PMCA efficiently demonstrated the presence of PrPSc in urine14, CSF19, blood15 and olfactory mucosa22 of patients with prion diseases, at concentrations that are six orders of magnitude below the detection limits of the current diagnostic techniques, such as Western blot. RT-QuIC assay on OM samples obtained by nasal swabbing has been largely tested in patients with sporadic or genetic forms of CJD, showing a specificity of 100% and sensitivity of 97 and 75%, respectively, but never in patients with FFI or at risk of developing FFI.
In this work, we analyzed OM samples of 28 subjects 2 of whom carry the D178N mutation in the PRNP gene and already died with a clinical diagnosis of FFI. The other samples were collected from patients with different neurodegenerative disorders (including AD, PD and FTD) and healthy controls. RT-QuIC and PMCA analysis revealed the presence of PrPSc in both FFI-OM samples that were collected 4 and 10 months after the disease onset. In both cases, three rounds of amplification were enough to enable detection of PrPSc. Moreover, through quantitative PMCA we have estimated that the OM sample analyzed contained about 1 × 10−14 g/ml of PrPSc. This estimation should be carefully considered before being extended to other samples since it suffers from some limitations that might influence PMCA sensitivity. For instance, the efficiency of the nasal brushing itself and the dilution of samples before analysis might introduce technical variables to the assay. Even more important, the olfactory neurons are unique since they are replaced through life and the amount of PrPSc collected during each nasal brushing might also depend on this phenomenon27. PrPSc was not detected in patients affected by other neurodegenerative diseases. At this point, we wondered whether PMCA was able to faithfully amplify the PrPSc originally detected in the OM of FFI patients. To this aim, we have performed a biochemical characterization of the amplified products and, after PK digestion, we observed a PrP banding profile characterized by a predominance of the diglycosylated species with the unglycosylated one migrating at around 19 kDa. This profile was compared to that of a PrPSc collected from the brain homogenate of a patient who died for FFI8. Both profiles were similar and suggested that PMCA of OM samples might be able to maintain the biochemical feature of the PrPSc typically associated to FFI. To confirm our observation, we decided to perform PMCA analysis in FFI brain homogenates and in two cases of sporadic sCJD-type 1 or sCJD-type 2 with PrPSc biochemical profiles different to that of FFI. Following PMCA amplification, FFI-PrPSc did not change its biochemical profile, while PrPSc associated to each sCJD type converted to a PrPSc glycotype characterized by a predominance of the diglycosylated band with the unglycosylated one migrating at 19 kDa (type 2). Therefore, we have realized that, even though our PMCA conditions enable PrPSc amplification, the original biochemical features of the prion strain were not retained. Recent PMCA experiments23,28 revealed that the glycoform ratio or the electrophoretic mobility of different prion strains are not faithfully maintained after amplification using bank vole homogenates as substrate for the reaction. These results seem to be corroborated by in vivo experiments which demonstrated that intracerebral inoculation of different prion strain (including sCJD-type 1 and type 2) in bank voles29 or transgenic mice expressing the bank vole prion protein25 with methionine at codon 109 (Tg109M) gave rise to PrPSc with electrophoretic mobilities similar to that of the original inoculum but with a glycoform ratio characterized by a predominance of the diglycosylated PrP band30. All these results suggest that, although bank voles can amplify different strains of prion, they might amplify PrPSc with biochemical properties slightly different to that of the original prion strain. Certainly, additional factors other than PrP itself could play an important role in driving the fidelity of prion replication and further studies are necessary to clarify this intricate mechanism of amplification. PK digestion of RT-QuIC amplification products enabled detection of protease resistant recPrP useful for identifying reactions seeded with prion infected olfactory mucosa samples. However, differences in the Western blot banding profiles (between sCJD, iCJD or FFI) do not seem to have any correlation with distinct disease phenotypes.
Since the involvement of the olfactory system, including the olfactory bulb, in FFI has never been deeply investigated, we have hypothesized that PrPSc could have spread from the central nervous system (CNS) to the OM through axonal transport along nerve fibers, as it happens for the sporadic forms of CJD31. Otherwise, some mutations of the PrPC gene, especially the D178N which is located in the hydrophobic core of the protein, are known to reduce the stability of the protein which is subsequently more prone to misfold32,33. This process might be favored in the nasal cavity, which is constantly exposed to chemical and physical stress34 which predisposes PrPC to acquire less stable conformations, eventually leading to the formation of PrPSc. If this were the case, the presence of PrPSc in the OM is not directly linked to disease progression but instead represents a completely independent event.
Our results demonstrated that the OM collected from symptomatic FFI patients contain minimal amount of PrPSc. Therefore, considering the extremely high levels of sensitivity and specificity of PMCA and RT-QuIC, we are planning to investigate whether PrPSc might be detectable in subjects carrying the D178N mutation in the preclinical stage. There is only one study showing pre-symptomatic alterations in the thalamus of a D178N carrier who underwent to serial 18FDG-PET that was characterized by a selective hypometabolism in the thalamus 13 months before the clinical onset. No correlations between thalamic dysfunction and presence of PrPSc were analyzed35. Prospective longitudinal studies will enable to evaluate whether detection of PrPSc in OM collected from subjects of a well characterized Italian FFI family, which is currently involved in a preventive clinical trial with doxycycline, might be used as quantifiable biomarker to monitor the effect of the drug on PrP replication and disease progression33.
Materials and Methods
Ethical approval
The study was approved by the institutional review board of Carlo Besta Neurological Institute and performed according to the guidelines approved by the ethics committee. Written informed consent for participation in research was done in accordance with the Declaration of Helsinki (1964–2008) and the Additional Protocol on the Convention of Human Rights and Biomedicine concerning Biomedical Research (2005).
Olfactory mucosa brushing procedure
A step-by-step tutorial video of the nasal brushing procedure is available at: https://www.youtube.com/watch?v=wYb9W3u6uMY. This procedure was performed using flocked swabs (FLOQSwabsTM Copan Italia, Brescia, Italy), as previously described22,36.
Brain homogenates preparation
Frontal cortex of patients with Fatal Familial Insomnia (FFI-D178N), Alzheimer’s disease (AD), Frontotemporal Dementia (FTD) sporadic Creutzfeldt-Jakob disease (sCJD) and iatrogenic Creutzfeldt-Jakob disease (iCJD) were homogenized in phosphate buffer (pH 7.4, Sigma) at 10% (weight/volume). Samples were centrifuged (Eppendorf Centrifuge) at 4 °C, 800 × g, 1 minute in order to remove cellular debris. Supernatant was collected and stored at −80 °C for further use.
Olfactory mucosa cells collections
Olfactory mucosa (OM) cells were collected from the mid-part of the inferior turbinate by brushing with a cotton swab, after treating the nasal mucosa of the patients with a topical anesthetic. Cotton swab was subsequently immersed in saline solution and olfactory cells collected from the brush by means of vortexing.
Olfactory mucosa cells preparation for PMCA analysis
Olfactory mucosa cells were pelleted for 20 minutes at 800 × g at 4 °C. The supernatant was removed and approximately 2 μl of the pellet was collected and transferred into a tube containing 25 μl of PBS. The latter was sonicated (approximately 280 W) with a Microsonicator QSonica (Q700) until the pellet was dispersed and 10 μL of this solution was added to 90 μL of specific PMCA substrate and subjected to several rounds of amplification.
Substrate preparation for PMCA analysis
Bank voles carrying the M109 PrP genotype were sacrificed with carbon dioxide and immediately perfused using phosphate-buffered saline (PBS) plus 5 mM ethylendiaminetetraacetic acid (EDTA, Sigma). Brains were collected and homogenized in conversion buffer (PBS, 150 mM NaCl, 1% Triton X-100) at 10% (weight/volume). Heparin (100 μg/mL, Sigma), digitonin (0.05%, Sigma) and 3 teflon beads were added to the homogenates to increase PMCA efficiency. To avoid contamination, perfusion and brain homogenates were carried out in a prion-free laboratory.
PMCA procedure
PMCA was performed as previously reported36. Briefly, 10 μL of sonicated olfactory mucosa samples was added to 90 μL of fresh substrate and subjected to 96 cycles of PMCA with the use of a microsonicator (model Q700, Qsonica). Each cycle consisted of 29 minutes 30 seconds of incubation at 37–40 °C, followed by a 30-second pulse of sonication set at a potency of 260–280 W. After one round of PMCA, an aliquot of the amplified material was diluted by a factor of 10 into fresh brain homogenate and an additional round was performed. Each sample was subjected to 7 rounds of PMCA. Considering our ultrasensitive level of amplification, all PMCA reactions were prepared under rigorous prion-free conditions. Overall, in order to avoid any contamination, appropriate number of negative controls were added in each PMCA assay. No contamination or de novo generation of prions were observed.
SDS-PAGE and Western blotting
Ten microliters of final PMCA products was treated with 50 μg/mL of PK (Invitrogen) for 1 hour at 37 °C under shaking (550 rpm). Digestion was stopped by the addition of LDS-PAGE loading buffer (Life Technologies), samples were heated at 100 °C for 10 minutes and loaded into 12% Bolt Bis-Tris Plus gels (Life Technologies). Proteins were separated by means of SDS-PAGE, transferred onto Polyvinylidene difluoride (PVDF, Millipore) membrane and incubated with 5% (wt/vol) dry nonfat milk in 0.05% (vol/vol) Tween-20 (prepared in Tris-HCl) for 1 h at room temperature with shaking. PVDF membranes were finally incubated with anti-PrP antibody 6D11 (epitopes 93–109, Covance) and developed with chemiluminescent system (ECL Prime, GE Healthcare Amersham).
Ten microliters of RT-QuIC reaction products was digested with 50 μg/mL of PK at 37 °C for 30 minutes under shaking (550 rpm). Digestion was stopped by the addition of LDS-PAGE loading buffer, samples were heated at 100 °C for 10 minutes and loaded into 17% Bolt Bis-Tris Plus gels. Proteins were separated by means of SDS-PAGE, transferred onto Polyvinylidene difluoride (PVDF) membrane and incubated with 5% (wt/vol) dry nonfat milk in 0.05% (vol/vol) Tween-20 (prepared in Tris-HCl) for 1 h at room temperature with shaking. PVDF membranes were finally incubated with anti-PrP antibody SAF-84 (epitopes 160–170, Spi Bio) and developed with chemiluminescent system (ECL Prime).
Recombinant PrP production for RT-QuIC analysis
Truncated Bank voles PrP (BvPrP(90-231)) construct was purchased from GenScript. The construct was expressed in Escherichia coli BL21 (DE3) cells (Stratagene). Freshly transformed overnight culture was inoculated into Luria Bertani (LB) medium and 100 μg/mL ampicillin. At 0.8 OD600 expression was induced with isopropyl b-D galactopyranoside (IPTG) to a final concentration of 0.75 mM. Cells were grown in a BioStat-B plus fermentor (Sartorius). The cells were lysed by a homogenizer (PandaPLUS 2000) and the inclusion bodies were suspended in buffer containing 25 mM Tris-HCl, 5 mM EDTA, 0.8% TritonX100, pH 8, and then in bi-distilled water several times. Inclusion bodies containing BvPrP(90-231) were dissolved in 5 volumes of 8 M guanidine hydrochloride (GndHCl), loaded onto pre-equilibrated HiLoad 26/60 Superdex 200-pg column, and eluted in 25 mM Tris–HCl (pH 8), 5 mM ethylenediaminetetraacetic acid (EDTA), and 6 M GndHCl at a flow/rate of 2 mL/min. Proteins refolding was performed by dialysis against refolding buffer (20 mM sodium acetate and 0.005% NaN3 (pH 5.5)) using a Spectrapor membrane. Purified protein was analyzed by SDS-polyacrylamide gel electrophoresis under reducing conditions and Western blot.
Olfactory mucosa cells preparation for RT-QuIC assay
Olfactory cells were collected from the brush by means of vortexing. The brush was then removed and the cells were pelleted for 20 minutes at 800 × g at 4 °C. The supernatant was removed, and approximately 2 μl of the pellet was collected and transferred into a tube containing 25 μl of PBS. The latter was sonicated (approximately 280 W) until the pellet was dispersed and 2 μL of this solution was further diluted in 18 μL of freshly prepared PBS. Two μL of the final solution was added to each reaction well.
Real-Time Quaking Induced Conversion procedure
After purification, aliquots of the recombinant BvPrP(90-231) were stored at −80 °C in 10 mM phosphate buffer (pH 5.8). Before each test, the protein solution was allowed to thaw at room temperature and filtered through a 100 kDa Nanosep centrifugal device (Pall Corporation). The concentration of recPrP was determined by measuring the adsorbance at 280 nm. Samples were blindly analyzed at least three times (by different operators) in triplicate in a black 96-well optical flat bottom plate (Thermoscientific) and results showed a high degree of consistency.
The final reaction volume was 100 μL and the reagents (Sigma) were concentrated as follow: 150 mM NaCl, 0.002% SDS, 1X PBS, 1 mM EDTA, 10 μM ThT and 0.1 mg recPrP ml−1. To avoid contamination, reaction mix was prepared in a prion-free laboratory. After the addition of 2 μL of olfactory mucosa samples, the plate was sealed with a sealing film (Thermoscientific) and inserted into a FLUOstar OPTIMA microplate reader (BMG Labtech). The plate was shaken for 1 minute at 600 rpm (double orbital) and incubated for 1 minute at 55 °C. Fluorescence reading (480 nm) were taken every 15 minutes (30 flashes per well at 450 nm). Given the rapid response, a specific threshold was set to decrease the likelihood of false positives. A sample was considered positive if the mean of the highest two fluorescence values (AU) of the replicates was higher than 10.000 AU and at least two out of three replicates crossed the threshold that was set at 30 hours. This reaction cutoff was established because in all the experiments that were seeded with the brain homogenates of patients with Alzheimer’s disease, no positive RT-QuIC reactions were observed until after 30 hours. In rare occasions, we could observe the aggregation of 1 replicate out of 3 before the threshold. In these cases, we have performed additional analysis of the samples in quadruplicate before considering it negative. The sample was confirmed negative if none or only one replicate (out of four) crossed the threshold before 30 hours. This allowed us to maximize the accuracy of the analysis thus reducing the risk of false positive or false negative results.
Additional Information
How to cite this article: Redaelli, V. et al. Detection of prion seeding activity in the olfactory mucosa of patients with Fatal Familial Insomnia. Sci. Rep. 7, 46269; doi: 10.1038/srep46269 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Acknowledgments
This work was supported in part by the Italian Ministry of Health (GR-2013-02355724), MJFF, ALZ, Alzheimer’s Research UK and the Weston Brain Institute (BAND2015), and La Foundation pour la Recherche Medicale (FABS201402) to FM; the Italian Ministry of Health and Associazione Italiana Encefalopatie da Prioni (AIEnP) to FT; Telethon (GSP14001) to GF. The funders had no role in the study design, data collection and analysis or the preparation of the manuscript.
Footnotes
The authors declare no competing financial interests.
Author Contributions V.R. and E.B. performed all PMCA and RT-QuIC experiments, analyzed the data and prepared the figures of the manuscript; G.Z. helped to coordinate the study, performed part of the experiments, data analysis and critically reviewed the manuscript; G.S. produced recombinant bank vole PrP used for RT-QuIC analysis; L.S. and S.M.P. performed the nasal brushing procedures; M.R. and C.M.G.D.L. performed part of the RT-QuIC analysis and Western blot experiments; M.D.B. and U.A. collected bank voles brain homogenates and critically reviewed the manuscript; G.L., I.R., G.F. and F.T. helped to coordinate the study and critically reviewed the manuscript. V.R., E.B. and Z.G. helped in writing the manuscript. F.M. supervised the work and wrote the final version of the manuscript. All the authors read and approved the final version of the manuscript.
References
- Parchi P. et al. Classification of sporadic Creutzfeldt-Jakob disease based on molecular and phenotypic analysis of 300 subjects. Ann Neurol 46, 224–233 (1999). [PubMed] [Google Scholar]
- Bishop M. T., Will R. G. & Manson J. C. Defining sporadic Creutzfeldt-Jakob disease strains and their transmission properties. Proc Natl Acad Sci USA 107, 12005–12010, doi: 10.1073/pnas.1004688107 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill A. F. et al. The same prion strain causes vCJD and BSE. Nature 389, 448–450, 526, doi: 10.1038/38925 (1997). [DOI] [PubMed] [Google Scholar]
- Collinge J. Prion diseases of humans and animals: their causes and molecular basis. Annu Rev Neurosci 24, 519–550, doi: 10.1146/annurev.neuro.24.1.519 (2001). [DOI] [PubMed] [Google Scholar]
- Mastrianni J. A. In GeneReviews(R) (eds Pagon R. A. et al.) (1993). [Google Scholar]
- Medori R. et al. Fatal familial insomnia: a second kindred with mutation of prion protein gene at codon 178. Neurology 42, 669–670 (1992). [DOI] [PubMed] [Google Scholar]
- Krasnianski A. et al. A proposal of new diagnostic pathway for fatal familial insomnia. J Neurol Neurosurg Psychiatry 85, 654–659, doi: 10.1136/jnnp-2013-305978 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossi G. et al. Fatal familial insomnia: genetic, neuropathologic, and biochemical study of a patient from a new Italian kindred. Neurology 50, 688–692 (1998). [DOI] [PubMed] [Google Scholar]
- McKinley M. P., Bolton D. C. & Prusiner S. B. A protease-resistant protein is a structural component of the scrapie prion. Cell 35, 57–62 (1983). [DOI] [PubMed] [Google Scholar]
- Levavasseur E. et al. Regulating factors of PrP glycosylation in Creutzfeldt-Jakob disease–implications for the dissemination and the diagnosis of human prion strains. PLoS One 3, e2786, doi: 10.1371/journal.pone.0002786 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wadsworth J. D., Hill A. F., Beck J. A. & Collinge J. Molecular and clinical classification of human prion disease. Br Med Bull 66, 241–254 (2003). [DOI] [PubMed] [Google Scholar]
- Saborio G. P., Permanne B. & Soto C. Sensitive detection of pathological prion protein by cyclic amplification of protein misfolding. Nature 411, 810–813, doi: 10.1038/35081095 (2001). [DOI] [PubMed] [Google Scholar]
- Saa P., Castilla J. & Soto C. Ultra-efficient replication of infectious prions by automated protein misfolding cyclic amplification. J Biol Chem 281, 35245–35252, doi: 10.1074/jbc.M603964200 (2006). [DOI] [PubMed] [Google Scholar]
- Moda F. et al. Prions in the urine of patients with variant Creutzfeldt-Jakob disease. N Engl J Med 371, 530–539, doi: 10.1056/NEJMoa1404401 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Concha-Marambio L. et al. Detection of prions in blood from patients with variant Creutzfeldt-Jakob disease. Sci Transl Med 8, 370ra183, doi: 10.1126/scitranslmed.aaf6188 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bougard D. et al. Detection of prions in the plasma of presymptomatic and symptomatic patients with variant Creutzfeldt-Jakob disease. Sci Transl Med 8, 370ra182, doi: 10.1126/scitranslmed.aag1257 (2016). [DOI] [PubMed] [Google Scholar]
- Lacroux C. et al. Preclinical detection of variant CJD and BSE prions in blood. PLoS pathogens 10, e1004202, doi: 10.1371/journal.ppat.1004202 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atarashi R. et al. Simplified ultrasensitive prion detection by recombinant PrP conversion with shaking. Nat Methods 5, 211–212, doi: 10.1038/nmeth0308-211 (2008). [DOI] [PubMed] [Google Scholar]
- McGuire L. I. et al. Real time quaking-induced conversion analysis of cerebrospinal fluid in sporadic Creutzfeldt-Jakob disease. Ann Neurol 72, 278–285, doi: 10.1002/ana.23589 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verde F. et al. MRI abnormalities found 1 year prior to symptom onset in a case of Creutzfeldt-Jakob disease. Journal of neurology 263, 597–599, doi: 10.1007/s00415-016-8022-6 (2016). [DOI] [PubMed] [Google Scholar]
- Sano K. et al. Early detection of abnormal prion protein in genetic human prion diseases now possible using real-time QUIC assay. PLoS One 8, e54915, doi: 10.1371/journal.pone.0054915 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orru C. D. et al. A test for Creutzfeldt-Jakob disease using nasal brushings. N Engl J Med 371, 519–529, doi: 10.1056/NEJMoa1315200 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cosseddu G. M. et al. Ultra-efficient PrP(Sc) amplification highlights potentialities and pitfalls of PMCA technology. PLoS Pathog 7, e1002370, doi: 10.1371/journal.ppat.1002370 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orru C. D. et al. Bank Vole Prion Protein As an Apparently Universal Substrate for RT-QuIC-Based Detection and Discrimination of Prion Strains. PLoS Pathog 11, e1004983, doi: 10.1371/journal.ppat.1004983 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watts J. C. et al. Evidence that bank vole PrP is a universal acceptor for prions. PLoS Pathog 10, e1003990, doi: 10.1371/journal.ppat.1003990 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parchi P. et al. Molecular pathology of fatal familial insomnia. Brain Pathol 8, 539–548 (1998). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghanbari H. A. et al. Oxidative damage in cultured human olfactory neurons from Alzheimer’s disease patients. Aging Cell 3, 41–44 (2004). [DOI] [PubMed] [Google Scholar]
- Vanni I. et al. In vitro replication highlights the mutability of prions. Prion 8, 154–160, doi: 10.4161/pri.28468 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nonno R. et al. Efficient transmission and characterization of Creutzfeldt-Jakob disease strains in bank voles. PLoS Pathog 2, e12, doi: 10.1371/journal.ppat.0020012 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espinosa J. C. et al. PrPC Governs Susceptibility to Prion Strains in Bank Vole, While Other Host Factors Modulate Strain Features. J Virol 90, 10660–10669, doi: 10.1128/JVI.01592-16 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zanusso G. et al. Detection of pathologic prion protein in the olfactory epithelium in sporadic Creutzfeldt-Jakob disease. N Engl J Med 348, 711–719, doi: 10.1056/NEJMoa022043 (2003). [DOI] [PubMed] [Google Scholar]
- van der Kamp M. W. & Daggett V. Pathogenic mutations in the hydrophobic core of the human prion protein can promote structural instability and misfolding. J Mol Biol 404, 732–748, doi: 10.1016/j.jmb.2010.09.060 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen R. B., Parchi P., Richardson S. L., Urig C. B. & Gambetti P. Effect of the D178N mutation and the codon 129 polymorphism on the metabolism of the prion protein. J Biol Chem 271, 12661–12668 (1996). [DOI] [PubMed] [Google Scholar]
- Elad D., Naftali S., Rosenfeld M. & Wolf M. Physical stresses at the air-wall interface of the human nasal cavity during breathing. J Appl Physiol (1985) 100, 1003–1010, doi: 10.1152/japplphysiol.01049.2005 (2006). [DOI] [PubMed] [Google Scholar]
- Cortelli P. et al. Pre-symptomatic diagnosis in fatal familial insomnia: serial neurophysiological and 18FDG-PET studies. Brain 129, 668–675 , doi: 10.1093/brain/awl003 (2006). [DOI] [PubMed] [Google Scholar]
- Morales R., Duran-Aniotz C., Diaz-Espinoza R., Camacho M. V. & Soto C. Protein misfolding cyclic amplification of infectious prions. Nat Protoc 7, 1397–1409, doi: 10.1038/nprot.2012.067 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.