

Abstract
Over the course of the day, the heart is challenged with dramatic fluctuations in energetic demand and nutrient availability. It is therefore not surprising that rhythms in cardiac metabolism have been reported at multiple levels, including the utilization of glucose, fatty acids, and amino acids. Evidence has emerged suggesting that the cardiomyocyte circadian clock is in large part responsible for governing cardiac metabolic rhythms. In doing so, the cardiomyocyte clock temporally partitions ATP generation for increased contractile function during the active period, promotes nutrient storage at the end of the active period, and facilitates protein turnover (synthesis and degradation) during the beginning of the sleep phase. This review highlights the roles of cardiac metabolism rhythms as well as the potential pathological consequences of their impairment.
Keywords: amino acid, circadian clock, fatty acid, glucose, heart, metabolism, transcription
Introduction
Time of day impacts virtually all aspects of life, including death Processes as fundamental as sleep, locomotion, cognitive function, feeding, digestion/absorption, body temperature, neural activity, endocrine factor release, and cardiovascular function all exhibit time-of-day-dependent oscillations.1,2 In terms of the cardiovascular system, both physiological (e.g., blood pressure, heart rate, cardiac output) and pathophysiological (e.g., adverse ischemic events, arrhythmias, sudden cardiac death) parameters vary over the course of the day.3 It is therefore not surprising that at biologic/molecular levels, cardiovascular components are markedly different during the day versus the night.
In the case of the heart, unbiased transcriptome and proteome analyses reveal an array of gene/protein expression differences as a function of time of day, thereby influencing a plethora of cellular processes.4,5 In addition, candidate signaling approaches suggest daily oscillations in the heart's sensitivity to extracellular stimuli such as fatty acids, β-adrenergic agonist, and thyroid hormone.6–9 The focus of this review is on circadian control of cardiac metabolism, including the mechanisms involved in synchronizing metabolic rhythms, their role in maintaining cardiac function, and the pathological consequences of their disruption/impairment.
Synchronization of Metabolic Rhythms
Time-of-day-dependent oscillations in metabolism have been described at various levels in organisms ranging from yeast (e.g., NAD+/NADH ratio) to humans (e.g., body temperature and energy expenditure).10 Evolutionarily speaking, such oscillations are predicted to confer a selective advantage, allowing the organism to adapt to daily fluctuations in the environment—for example, lighting, temperature, humidity, food availability, and likelihood of predator interaction. At a biological level, it is therefore important to synchronize metabolic processes with time-of-day-dependent perturbations in energetic demand and nutrient availability that are associated with normal light/dark, sleep/wake, and fasting/feeding cycles. In mammals, such a synchronization must occur at intra- and inter-organ levels.
Classically, a stimulus-response model has been proposed wherein neurohumoral changes orchestrate oscillations in metabolic processes between cells/organs. With insulin, for example, increased secretion postprandially promotes peripheral tissue glucose utilization concomitant with attenuation of hepatic glucose production, thus facilitating glucose homeostasis by maintaining blood glucose within a physiologic range.11 What has become increasingly apparent is that cells/organs/organisms possess an ability to predict daily metabolic perturbations before they occur. In doing so, anticipation allows a temporally appropriate and rapid response to a stimuli once it occurs. Again, considering post-prandial insulin-mediated glucose utilization, the amplitude of both insulin secretion and sensitivity differs as a function of time of day due to anticipatory mechanisms.10
The process of anticipation is undoubtedly conferred by circadian clocks. Circadian clocks are cell-autonomous, transcriptionally based molecular mechanisms composed of a series of positive and negative feedback loops with a periodicity of approximately 24 hours. Central to the mechanism are BMAL1 and CLOCK, two transcription factors that, upon heterodimerization, bind to E-boxes within the promoter of target genes. The latter include multiple period and cryptochrome isoforms as well as REV-ERBα, which translocate back into the nucleus and inhibit the BMAL1/CLOCK heterodimer (i.e., feedback inhibition).1 In addition, these transcription factors affect expression of a large number of genes that are not core clock components, also known as clock controlled genes (CCGs). By modulating the expression levels of CCGs in a time-of-day-dependent manner, the clock has the potential of temporally regulating a host of cellular processes such as transcription, translation, signal transduction, ion homeostasis, and metabolism.
A key element for maintaining this selective advantage is entrainment; cellular clocks must be synchronized with the environment. Within the hypothalamus, a region of approximately 20,000 specialized neurons known as the suprachiasmatic nucleus (SCN) receives light signals via the retinohypothalamic tract, resulting in entrainment. The SCN, often termed the “master clock,” subsequently sends neurohumoral signals to other tissues, thus resetting “slave clocks.” It is noteworthy that non-SCN clocks can also be entrained through common behaviors such as eating and physical activity. These entrainment mechanisms ensure that cell autonomous clocks can maintain synchrony between environmental/behavioral-associated factors (e.g., energetic demand and nutrient availability) and biological processes such as metabolism.
Cardiac Metabolic Rhythms
With respect to metabolism, the heart has two 24-hr cycles: sleep/wake and fasting/feeding. Importantly, the heart must meet increased energetic demands even if the animal in the wild is unsuccessful in its forage for food, as the animal continues to forage and avoid predation. It is therefore not surprising that dramatic fluctuations in cardiac metabolism have been reported at various levels, including daily rhythms in cardiac glucose, fatty acid, and amino acid utilization.6,12–16 During periods of increased physical activity, the heart relies primarily on augmented glucose utilization to meet energetic needs for force generation and ion homeostasis.17,18 Consistent with this concept, myocardial glycolysis and glucose oxidation both increase during the awake phase (relative to the sleep phase; Figure 1 A).12,15 Importantly, these rhythms in glucose metabolism are observed in ex vivo perfused rat and mouse hearts, wherein both the milieu and workload are maintained at a constant level. This suggests that they are mediated, at least in part, by an intrinsic mechanism. A likely candidate is the cardiomyocyte circadian clock, as genetic ablation of this mechanism markedly attenuates/abolishes myocardial glucose metabolism rhythms.15,19 Collectively, these observations have led to the concept that the cardiomyocyte circadian clock allows anticipation of increased energetic demand during the awake period through modulation of glucose utilization.
Figure 1.
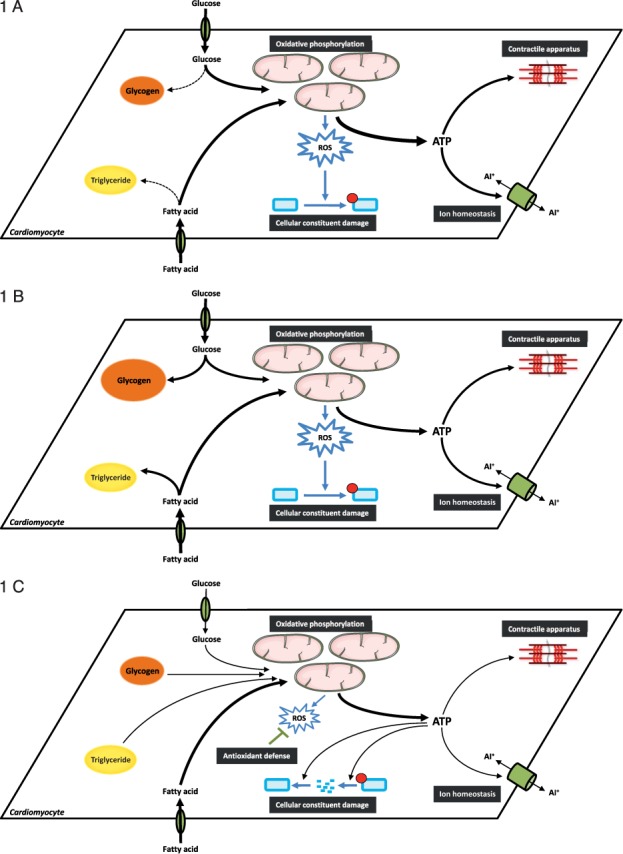
Time-of-day-dependent oscillations in cardiac metabolism. (A) At the sleep-to-wake transition, energetic demand increases in the heart due to increased E-C coupling as the animal forages for food and avoids predation. This elevated energetic demand is met by increased glucose utilization (glucose uptake, glycolysis, and oxidation). During this period, cellular constituents (e.g., protein, phospholipids) are susceptible to oxidative damage. (B) In the latter half of the active period, excess nutrients are utilized for the synthesis of both glycogen and triglyceride. (C) During the sleep period, reliance on stored nutrients increases to meet the energetic demands of the heart; due to lower contractility, ATP can be utilized for the turnover of cellular constituents, thus replacing damaged proteins and phospholipids. At this time, the heart is less susceptible to oxidative damage, potentially through increased antioxidant capacity. In many cases, temporal partitioning of these metabolic processes is mediated by the cardiomyocyte circadian clock.
Diurnal variations in cardiac metabolism extend beyond glucose catabolism. Turnover (i.e., synthesis/degradation cycles) of glycogen, triglycerides, and protein exhibit time-of-day-dependence in rodent hearts. In the cases of glycogen and triglyceride synthesis, peak rates are observed in the middle/end of the active/awake phase (Figure 1 B); similar to glycolysis and glucose oxidation, oscillations in glycogen and triglyceride synthesis persist in ex vivo perfused hearts and are severely attenuated or abolished following genetic ablation of the cardiomyocyte circadian clock.14,15 These findings have led to speculation that the heart increases nutrient storage towards the end of the active period following successful forage for food in anticipation of the upcoming sleep-phase fast. Interestingly, both protein synthesis and degradation processes (ubiquitin-proteasome system and autophagy) are elevated at the beginning of the sleep phase (Figure 1 C), and initial studies suggest that these rhythms are in part dependent on the cardiomyocyte circadian clock.16,20
Protein turnover utilizes large quantities of ATP, leading to the hypothesis that this temporal partitioning is advantageous because it (1) spares ATP for contractile function during the active/awake phase, (2) attenuates protein synthesis during the active period when various prohypertrophic stimuli (e.g., blood pressure, epinephrine, amino acids) are elevated, and (3) allows replacement of myocardial proteins that were damaged during the active/awake period (when oxidative metabolism, respiration rate, and oxidant/toxin consumption from diet is increased) in anticipation of the upcoming active/awake period. Likewise, phospholipid synthesis is increased during the sleep phase, possibly in an attempt to replace recently damaged/oxidized lipids.14 Interestingly, previous studies indicate that the heart is resistant to oxidative stress at the beginning of the sleep phase.21 One possible explanation is that increased antioxidant potential at this time (as observed in extra-cardiac tissues) attenuates misfolding or oxidative damage of proteins and other cellular constituents (Figure 1 C). Thus, temporal control of cardiac metabolism appears to play multiple important roles, ranging from the partitioning of ATP between energetically demanding processes to repair of damaged cellular constituents.
Disrupted Metabolic Rhythms as a Potential Mediator of Cardiac Dysfunction
In some ways, the heart is analogous to a car engine, burning fuel in order to provide the work/force required for other components of the vehicle to function. Indeed, the failing heart has previously been described as “an engine out of fuel.”22 Timing is critical for any engine, as improper timing of fuel ignition adversely affects performance. In addition, the lifespan of the engine is greatly increased through regular preventive maintenance. Metabolic rhythms in the heart serve both timing and preventive maintenance functions. Increased glucose utilization during the active/awake period likely ensures that sufficient ATP is available to meet energetic demands associated with elevated contractile function, while increased protein turnover during the sleep period prevents accumulation of damaged proteins and organelles (e.g., mitochondria), ensuring maintenance of cardiac function. Such concepts suggest that an impairment in metabolic rhythms, either mistiming (i.e., phase shift) or amplitude attenuation, would hinder cardiac performance and ultimately precipitate cardiac dysfunction. Evidence exists to support this theory, including findings that (1) genetic and/or environmental disruption of circadian clocks leads to cardiac dysfunction, and (2) circadian clocks are altered during various cardiometabolic diseases. Herein follows an overview of this evidence.
Whole-body metabolic rhythms diminish with age and are abolished completely as an animal approaches death (by natural causes).23 Interestingly, germline genetic ablation of circadian clock function through knockout of BMAL1 results in an accelerated aging phenotype associated with loss of biological rhythms at multiple levels, particularly metabolism.24 Serial echocardiography revealed development of dilated cardiomyopathy in BMAL1 knockout mice, which correlates with decreased lifespan.25 Underscoring the importance of the clock in the heart, cardiomyocyte-specific BMAL1 ablation recapitulates both cardiac dysfunction and decreased lifespan observed in germline knockout mice.19 Similarly, genetic disruption of the timing of the circadian clock or circadian clock output (as seen in tau mutant hamsters and DBP/HLF/TEF triple knockout mice, respectively) results in cardiac dysfunction.26,27 Circadian clocks are also disrupted through changes in environmental/behavioral factors such as lighting, food consumption, and physical activity at the “wrong” time of day. Indeed, shift workers exhibit impaired circadian rhythms at multiple levels and have an approximate 2-fold increased risk of developing cardiometabolic disease.28,29
The circadian clock in the heart is attenuated during pressure overload and hypertension-induced hypertrophy as well as ischemia/reperfusion.30–32 In contrast, this molecular mechanism is phase shifted during uncontrolled type 1 diabetes mellitus.33 Despite these observations, to date no studies have directly assessed (through flux measurements) whether cardiac metabolic rhythms are impaired or altered during disease states. However, indirect evidence supports this idea. For example, time-of-day-dependent rhythms in expression of genes encoding for metabolic proteins (e.g., glucose transport 4) were markedly attenuated in the heart following pressure-overload induced hypertrophy.12 Should these perturbations in gene expression oscillations translate to metabolic flux, the concept of metabolic inflexibility as a contributing factor in cardiac dysfunction (which usually considers one or more chronic disease states) could lead to a whole new paradigm: that is, the possibility that disease states abolish time-of-day-dependent oscillations in metabolism.
Summary
Temporal partitioning of cardiac metabolism has emerged as a fundamental cornerstone in cardiac biology, governed in large part by the autonomous cardiomyocyte circadian clock. What remains to be fully explored are the molecular links between the circadian clock and metabolic processes as well as the extent to which metabolic rhythms are perturbed during disease states and their precise contribution towards the pathogenesis of cardiac dysfunction. Similarly, the extent to which the circadian clock synergizes with neurohumoral influences (e.g., [nor]epinephrine, insulin) to modulate daily metabolic processes remains to be fully elucidated.
Key Points
The heart exhibits profound time-of-day-dependent oscillations in glucose, fatty acid, and protein metabolism.
The cardiomyocyte circadian clock governs cardiac metabolic rhythms.
Disruption of circadian clocks leads to impaired metabolic rhythms and cardiac dysfunction.
Conflict of Interest Disclosure
This work was supported by funding from the National Heart, Lung, and Blood Institute (HL106199, HL074259, HL123574, HL122975).
References
- 1. Takahashi JS, Hong HK, Ko CH, McDearmon EL.. The genetics of mammalian circadian order and disorder: implications for physiology and disease. Nat Rev Genet. 2008. October; 9( 10): 764– 75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Edery I. Circadian rhythms in a nutshell. Physiol Genomics. 2000. August 9; 3( 2): 59– 74. [DOI] [PubMed] [Google Scholar]
- 3. Young ME. The circadian clock within the heart: potential influence on myocardial gene expression, metabolism, and function. Am J Physiol Heart Circ Physiol. 2006. January; 290( 1): H1– 16. [DOI] [PubMed] [Google Scholar]
- 4. Martino T, Arab S, Straume M, . et al. Day/night rhythms in gene expression of the normal murine heart. J Mol Med (Berl). 2004. April; 82( 4): 256– 64. [DOI] [PubMed] [Google Scholar]
- 5. Podobed P, Pyle WG, Ackloo S, . et al. The day/night proteome in the murine heart. Am J Physiol Regul Integr Comp Physiol. 2014. July 15; 307( 2): R121– 37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Bray M, Shaw CA, Moore MW, . et al. Disruption of the circadian clock within the cardiomyocyte influences myocardial contractile function, metabolism, and gene expression. Am J Physiol Heart Circ Physiol. 2008. February; 294( 2): H1036– 47. [DOI] [PubMed] [Google Scholar]
- 7. Durgan DJ, Trexler NA, Egbejimi O, . et al. The circadian clock within the cardiomyocyte is essential for responsiveness of the heart to fatty acids. J Biol Chem. 2006. August 25; 281( 34): 24254– 69. [DOI] [PubMed] [Google Scholar]
- 8. Collins HE, Rodrigo GC. Inotropic response of cardiac ventricular myocytes to beta-adrenergic stimulation with isoproterenol exhibits diurnal variation: involvement of nitric oxide. Circ Res. 2010. April 16; 106( 7): 1244– 52. [DOI] [PubMed] [Google Scholar]
- 9. Peliciari-Garcia RA, Previde RM, Nunes MT, Young ME.. Interrelationship between 3,5,3′- triiodothyronine and the circadian clock in the rodent heart. Chronobiol Int. 2016. September 23; 1– 11. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Bass J, Takahashi JS. Circadian integration of metabolism and energetics. Science. 2010. December 3; 330( 6009): 1349– 54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Boucher J, Kleinridders A, Kahn CR.. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb Perspect Biol. 2014. January 1; 6( 1). pii: a009191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Young ME, Razeghi P, Cedars AM, Guthrie PH, Taegtmeyer H.. Intrinsic diurnal variations in cardiac metabolism and contractile function. Circ Res. 2001. December 7; 89( 12): 1199– 1208. [DOI] [PubMed] [Google Scholar]
- 13. Durgan DJ, Moore M, Ha NP, . et al. Circadian rhythms in myocardial metabolism and contractile function: influence of workload and oleate. Am J Physiol Heart Circ Physiol. 2007. October 1; 293( 4): H2385– 93. [DOI] [PubMed] [Google Scholar]
- 14. Tsai JY, Kienesberger PC, Pulinilkunnil T, . et al. Direct regulation of myocardial triglyceride metabolism by the cardiomyocyte circadian clock. J Biol Chem. 2010. January 29; 285( 5): 2918– 29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Durgan DJ, Pat BM, Laczy B, . et al. O-GlcNAcylation, novel post-translational modification linking myocardial metabolism and cardiomyocyte circadian clock. J Biol Chem. 2011. December 30; 286( 52): 44606– 19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Rau E, Meyer DK. A diurnal rhythm of incorporation of L-[3H] leucine in myocardium of the rat. Recent Adv Stud Cardiac Struct Metab. 1975; 7: 105– 10. [PubMed] [Google Scholar]
- 17. Goodwin GW, Taylor CS, Taegtmeyer H.. Regulation of energy metabolism of the heart during acute increase in heart work. J Biol Chem. 1998. November 6; 273( 45): 29530– 9. [DOI] [PubMed] [Google Scholar]
- 18. Allard MF, Schonekess BO, Henning SL, English DR, Lopaschuk GD.. Contribution of oxidative metabolism and glycolysis to ATP production in hypertrophied hearts. Am J Physiol. 1994. August; 267( 2 Pt 2): H742– 50. [DOI] [PubMed] [Google Scholar]
- 19. Young ME, Brewer RA, Peliciari-Garcia RA, . et al. Cardiomyocyte-specific BMAL1 plays critical roles in metabolism, signaling, and maintenance of contractile function of the heart. J Biol Rhythms. 2014. August; 29( 4): 257– 76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. He L, Brewer RA, Shanmugam G, . et al. Influence of the cardiomyocyte circadian clock on autophagy in the heart. FASEB J. 2016. April; 30( 4): 1279. [Google Scholar]
- 21. Lapenna D, De Gioia S, Mezzetti A, Porreca E, Ciofani G, Marzio L.. Circadian variations in antioxidant defences and lipid peroxidation in the rat heart. Free Radic Res Commun. 1992; 17( 3): 187– 94. [DOI] [PubMed] [Google Scholar]
- 22. Neubauer S. The failing heart--an engine out of fuel. N Engl J Med. 2007. March 15; 356( 11): 1140– 51. [DOI] [PubMed] [Google Scholar]
- 23. Riera CE, Dillin A. Tipping the metabolic scales towards increased longevity in mammals. Nat Cell Biol. 2015. March; 17( 3): 196– 203. [DOI] [PubMed] [Google Scholar]
- 24. Kondratov RV, Kondratova AA, Gorbacheva VY, Vykhovanets OV, Antoch MP.. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006. July 15; 20( 14): 1868– 73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Lefta M, Campbell KS, Feng HZ, Jin JP, Esser KA.. Development of dilated cardiomyopathy in Bmal1-deficient mice. Am J Physiol Heart Circ Physiol. 2012. August 15; 303( 4): H475– 85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Martino TA, Oudit GY, Herzenberg AM, . et al. Circadian rhythm disorganization produces profound cardiovascular and renal disease in hamsters. Am J Physiol Regul Integr Comp Physiol. 2008. May; 294( 5): R1675– 83. [DOI] [PubMed] [Google Scholar]
- 27. Wang Q, Maillard M, Schibler U, Burnier M, Gachon F.. Cardiac hypertrophy, low blood pressure, and low aldosterone levels in mice devoid of the three circadian PAR bZip transcription factors DBP, HLF, and TEF. Am J Physiol Regul Integr Comp Physiol. 2010. October; 299( 4): R1013– 9. [DOI] [PubMed] [Google Scholar]
- 28. Knutsson A, Kempe A. Shift work and diabetes--a systematic review. Chronobiol Int. 2014. December; 31( 10): 1146– 51. [DOI] [PubMed] [Google Scholar]
- 29. Knutsson A, Akerstedt T, Jonsson BG, Orth-Gomer K.. Increased risk of ischaemic heart disease in shift workers. Lancet. 1986. July 12; 2( 8498): 89– 92. [DOI] [PubMed] [Google Scholar]
- 30. Young ME, Razeghi P, Taegtmeyer H.. Clock genes in the heart: Characterization and attenuation with hypertrophy. Circ Res. 2001. June 8; 88( 11): 1142– 50. [DOI] [PubMed] [Google Scholar]
- 31. Mohri T, Emoto N, Nonaka H, . et al. Alterations of circadian expressions of clock genes in Dahl salt-sensitive rats fed a high-salt diet. Hypertension. 2003. August; 42( 2): 189– 94. [DOI] [PubMed] [Google Scholar]
- 32. Kung TA, Egbejimi O, Cui J, . et al. Rapid attenuation of circadian clock gene oscillations in the rat heart following ischemia-reperfusion. J Mol Cell Cardiol. 2007. December; 43( 6): 744– 53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Young ME, Wilson CR, Razeghi P, Guthrie PH, Taegtmeyer H.. Alterations of the circadian clock in the heart by streptozotocin-induced diabetes. J Mol Cell Cardiol. 2002. February; 34( 2): 223– 31. [DOI] [PubMed] [Google Scholar]