

Abstract
Background and Purpose
Eucalyptol (1,8‐cineol), the major ingredient in the essential oil of eucalyptus leaves and other medicinal plants, has long been known for its anti‐inflammatory properties. Eucalyptol interacts with the TRP cation channels among other targets, but it is unclear which of these mediates its anti‐inflammatory effects.
Experimental Approach
Effects of eucalyptol were compared in wild‐type and TRPM8 channel‐deficient mice in two different models: footpad inflammation elicited by complete Freund's adjuvant (CFA) and pulmonary inflammation following administration of LPS. Oedema formation, behavioural inflammatory pain responses, leukocyte infiltration, enzyme activities and cytokine and chemokine levels were measured.
Key Results
In the CFA model, eucalyptol strongly attenuated oedema and mechanical allodynia and reduced levels of inflammatory cytokines (IL‐1β, TNF‐α and IL‐6), effects comparable with those of ibuprofen. In the LPS model of pulmonary inflammation, eucalyptol treatment diminished leukocyte infiltration, myeloperoxidase activity and production of TNF‐α, IL‐1β, IFN‐γ and IL‐6. Genetic deletion of TRPM8 channels abolished the anti‐inflammatory effects of eucalyptol in both models. Eucalyptol was at least sixfold more potent on human, than on mouse TRPM8 channels. A metabolite of eucalyptol, 2‐hydroxy‐1,8‐cineol, also activated human TRPM8 channels.
Conclusion and Implications
Among the pharmacological targets of eucalyptol, TRPM8 channels were essential for its anti‐inflammatory effects in mice. Human TRPM8 channels are more sensitive to eucalyptol than rodent TRPM8 channels explaining the higher potency of eucalyptol in humans. Metabolites of eucalyptol could contribute to its anti‐inflammatory effects. The development of more potent and selective TRPM8 agonists may yield novel anti‐inflammatory agents.
Abbreviations
- CFA
complete Freund's adjuvant
- TRP
transient receptor potential
Tables of Links
| TARGETS |
|---|
| Voltage‐gated ion channels |
| TRPA1 channels |
| TRPM8 channels |
| LIGANDS | |
|---|---|
| Eucalyptol | IL‐1β |
| LPS | IL‐6 |
| TNF‐α |
These Tables list key protein targets and ligands in this article which are hyperlinked to corresponding entries in http://www.guidetopharmacology.org, the common portal for data from the IUPHAR/BPS Guide to PHARMACOLOGY (Southan et al., 2016), and are permanently archived in the Concise Guide to PHARMACOLOGY 2015/16 (Alexander et al., 2015).
Introduction
Eucalyptol (1,8‐cineol) is the major medicinally active component of Eucalyptus essential oil, and it is present in other plant extracts used for medicinal purposes since antiquity, including myrtle, camphor, rosemary and sage oils (Hippocrates and Adams, 1886; Dymock et al., 1972; Barr, 1988). Eucalyptol‐containing products are widely used as a remedy for symptoms of the common cold and other respiratory infections and for alleviating pain (Juergens, 2014). In animal studies, eucalyptol was shown to counteract carrageenan‐induced oedema formation, acid‐, formalin‐ and heat‐evoked pain behaviour, gastric injury, colitis, allergen‐induced bronchoconstriction and inflammation, respiratory irritation due to inhalation of smoke irritants and endotoxin‐induced pulmonary inflammation and liver failure (Santos and Rao, 2000, 2001; Santos et al., 2001, 2004; Liapi et al., 2007; Bastos et al., 2009; Nascimento et al., 2009; Willis et al., 2011; Liu et al., 2013b; Zhao et al., 2014; Ha et al., 2015; Rocha Caldas et al., 2015). Clinical studies administering eucalyptol observed anti‐inflammatory effects in asthmatics, alleviation of symptoms of rhino‐sinusitis and reduced chronic obstructive pulmonary disease (COPD) exacerbations (Juergens et al., 2003; Kehrl et al., 2004; Worth et al., 2009; Juergens, 2014). Standardized preparations of essential oils containing eucalyptol as a major component had similar clinical effects (Meister et al., 1999).
Eucalyptol interacts with transient receptor potential (TRP) ion channels such as TRPM8, the cold/menthol receptor in peripheral sensory neurons, and TRPA1, an irritant receptor expressed in nociceptors (McKemy et al., 2002; Takaishi et al., 2012; Liu et al., 2013a). Eucalyptol is an agonist of TRPM8 channels and these channels are essential for at least some of eucalyptol's analgesic actions, as eucalyptol was shown to inhibit acid‐induced visceral pain in wild‐type mice, but not in TRPM8‐deficient mice (Liu et al., 2013b). Analgesia mediated by TRPM8 channels is likely to involve activation of central inhibitory circuits activated by input from TRPM8 channel‐expressing peripheral neurons (Proudfoot et al., 2006; Liu et al., 2013b; Vinuela‐Fernandez et al., 2014). TRPA1 channels are inhibited by eucalyptol and eucalyptol diminished pain elicited by application of TRPA1 channel agonists to human skin (Liu et al., 2013a; Takaishi et al., 2012; Takaishi et al., 2014).
Whether interactions with TRP channels underlie eucalyptol's anti‐inflammatory effects remains unclear. TRPM8 channels were essential for the suppression of inflammation by icilin, a potent TRPM8 channel agonist, in mouse models of colitis (Ramachandran et al., 2013). Inhibition of TRPA1 channels can also suppress inflammatory responses, as observed in animal models of asthma, dermatitis, arthritis and other inflammatory conditions (Escalera et al., 2008; Caceres et al., 2009; Bautista et al., 2013; Liu et al., 2013a; Rocha Caldas et al., 2015). Several studies have shown that eucalyptol may also engage targets in the immune system (Juergens, 2014). Eucalyptol interfered with arachidonic acid metabolism in blood monocytes of asthma patients, inhibited LPS‐induced IL‐1β production in human monocytes, suppressed LPS‐induced production of nitric oxide in a macrophage cell line and diminished LPS‐induced expression, translocation and transcriptional activities of early growth response factor 1 and NF‐κB in human cell lines (Juergens et al., 1998a,b; Zhou et al., 2007; Kim et al., 2015). Finally, eucalyptol also has antioxidant and antimicrobial activity that may contribute to its anti‐inflammatory effects in some pathological conditions (Keinan et al., 2005; Hendry et al., 2009; Sadlon and Lamson, 2010; Greiner et al., 2013).
The aim of the present study was to examine the role of TRPM8 channels in the anti‐inflammatory effects of eucalyptol in mouse models of inflammation replicating the most common conditions for which treatment with eucalyptol is frequently advised: pain accompanied by tissue swelling and respiratory inflammation. Eucalyptol treatment of mice strongly inhibited tissue oedema and inflammatory cytokine production elicited by paw injection of complete Freund's adjuvants (CFA) and prevented neutrophil infiltration and cytokine production in lungs of LPS‐exposed animals. While TRPM8 channel‐deficient mice developed normal inflammatory responses in these models, the anti‐inflammatory effects of eucalyptol were significantly diminished. These findings suggest that TRPM8 channels are the key mediators of eucalyptol's anti‐inflammatory effects and reinforce the notion that development of selective and potent agonists of TRPM8 channels may yield novel analgesic and anti‐inflammatory therapeutic agents.
Methods
Mice
All animal care and experimental procedures were approved by the Institutional Animal Care and Use Committees. Mice were housed in facilities accredited by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) under standard environmental conditions (12 h light–dark cycles and 23°C). Food and water were provided ad libitum. A total of 165 mice were used in the studies presented here. Animal studies are reported in compliance with the ARRIVE guidelines (Kilkenny et al., 2010; McGrath and Lilley, 2015).
Trpm8−/− mice were a gift from David Julius (University of California, San Francisco, CA, USA) and were transferred to Charles River Laboratories (Wilmington, MA, USA) for marker‐assisted backcrossing (Max‐Bax Speed Congenics) into C57BL/6 background (C57BL/6NCrl, Strain Code 027), confirming >99.9% strain congenicity after 12 crosses (Bautista et al., 2007; Liu et al., 2013b). Animals were then imported by embryo transfer into the Duke Laboratory Animal Resources breeding facility. C57BL/6 wild‐type founders were purchased from parent strain stock from Charles River Laboratories (C57BL/6NCrl, Strain Code 027) and bred in the same room to provide wild‐type control animals raised under identical conditions. Mice were identically matched for age (8–12 weeks) and sex and randomly distributed into the required number of experimental groups by breeding facility personnel before transfer into the research housing space.
CFA‐induced skin inflammation/paw swelling
CFA was injected (10 μL) into the plantar surface of the right hind paw. Saline was injected in control mice. Paw volumes were measured with a plethysmometer (IITC, Woodland Hills, CA, USA) immediately before (0 h, Vbefore) and 2, 4, 24 and 48 h (Vafter) after CFA injection. Changes in paw volume were shown as % increase in paw volume and calculated as follows: % increase in paw volume = (Vafter − Vbefore)/Vbefore. For therapy, eucalyptol (30, 100 or 300 mg·kg−1), ibuprofen (30 mg·kg−1) or vehicle (corn oil) were injected i.p. (25 μL·10 g−1 body weight injection volume) 30 min before CFA injection, and then again at 6, 24, 30 and 48 h after CFA injection for a total of five times for each group.
von Frey hair analysis of mechanical allodynia in CFA‐injected mice
Mice were habituated for 45 min to the wire mesh screen surface before testing. Paw withdrawal thresholds were determined using a series of von Frey filaments (0.008 to 6.00 g) pressed against the plantar surface of the hind paw in ascending order beginning with the finest fibre, following standard procedures (Chaplan et al., 1994; Liu et al., 2013b). The minimum force (g) that caused the mouse to withdraw its hind paw away from the filament was considered as the withdrawal threshold. For each paw, a von Frey hair was applied five times at 10 s intervals. The threshold was determined when paw withdrawal was observed in more than three out of five applications. A withdrawal response was considered valid only if the hind paw was removed completely from the platform. If the paw withdrawal response was ambiguous, the application was repeated. The test was performed 2 h after the third eucalyptol treatment injection or 26 h after the CFA paw injection. Behavioural tests were performed by an experimenter blinded to experimental conditions.
Skin sample collection and ELISA
Mice were killed 3 h after the last eucalyptol treatment. The skin of the CFA or saline‐treated paw was collected by a 4 mm biopsy punch (Miltex Inc., PA, USA), weighed and immediately frozen in liquid nitrogen. Tissue was homogenized using a Bullet Blender (NextAdvance, Averill Park, NY, USA) in 50 mM Tris‐base (pH 7.4) and 150 mM NaCl with protease inhibitors (Roche, Indianapolis, IN, USA) and 0.2% Triton‐X100 for 25 min at full speed. Then the samples were centrifuged at 10 000 g for 15 min at 4°C. The supernatant was tested by ELISA for IL‐1β, TNF‐α and IL‐6 (R&D Systems, Minneapolis, MN, USA), according to the manufacturer's instructions.
LPS‐induced pulmonary inflammation
Mice received a single intranasal dose of 10 μg LPS in 40 μL sterile saline under sevoflurane anaesthesia. Controls received saline only. For therapy, eucalyptol was injected i.p. at 200 mg·kg−1 in corn oil as vehicle (25 μL·10 g−1 injection volume). Treated mice received four i.p. injections: one 12 h before LPS administration, a second dose 1 h before LPS, a third dose 12 h after LPS and a fourth dose 1 h before killing. Mice were killed for sample collections 24 h after LPS administration, a time point where the acute lung inflammation (ALI) peaks (Hirano, 1997; Santos et al., 2006). Control animals received vehicle (corn oil) at the same intervals above.
Collection and analysis of bronchoalveolar lavage (BAL) fluid
BAL was performed by cannulation of the trachea and gentle instillation/aspiration (three times) of 1.0 mL of PBS with 0.1% BSA and protease inhibitors (Roche, Indianapolis, IN) as published before (Caceres et al., 2009). The lavage fluid was centrifuged at 1500 × g for 5 min, and the supernatant was frozen at −80°C for later assessment of cytokines and myeloperoxidase (MPO) activity. The cell pellet was treated with red‐blood‐cells lysing buffer (BD Biosciences, San Diego, CA, USA), washed and resuspended in 200 μL of PBS. Total cell counts were determined with a haematology analyser (Scil Vet ABC, Gurnee, IL, USA), spun onto cytoslides (Cytospin 3, Shandon Inc, Pittsburg, PA, USA) and stained with Diff‐Quick (Dade‐Behring Inc., Newark, DE, USA). Differential cell counts were obtained by microscopic counting of a minimum of 200 cells per slide and using standard morphological and staining criteria.
Quantitative analysis of cytokines and chemokines in BAL fluid
Cytokines and chemokines in the BAL fluid were measured using the Luminex xMAP technology. Analyte levels were determined using a Milliplex MAP mouse cytokine/chemokine kit (Millipore, Billerica, MA, USA) following the manufacturer's instructions. Briefly, 50 μL of BALF were added in duplicate to each well of a 96‐well plate. Antibody‐conjugated beads solution was added to each well and incubated in the dark prior to incubation with the biotinylated detection antibody specific for each analyte. Fluorescence intensity was determined by incubation with streptavidin‐phycoerythrin solution. All readings were carried out using a Luminex 200 analyzer (Luminex, Austin, TX, USA). MPO activity in the BAL fluid was assayed using a chlorination assay kit (Cayman Chemical Company, MI, USA).
RNA extraction and cDNA synthesis
After obtaining the BAL, lungs were perfused with PBS, surgically removed and snap‐frozen by immersion in liquid nitrogen. Total RNA was isolated from the tissue homogenates of each sample using the RNeasy Mini Kit 50 (Qiagen, MD, USA). The homogenization of all the tissues in Buffer RLT with 2‐mercaptoethanol was carried out in a NextAdvance bullet blender (max speed, 5 min). cDNA synthesis was performed from 500 ng of RNA right after the RNA isolation following the manufacturer's instructions (High Capacity Reverse Transcription Kit, Applied Biosystems).
Real‐time PCR (qPCR)
TaqMan® Gene Expression Assays from Applied Biosystems (Foster City, CA, USA) were used for expression assessment. About 20 μL reactions contained 10 μL of TaqMan Fast Universal PCR Master Mix (2×), 1 μL of the specific TaqMan assay, 1 μL cDNA and water up to 20 μL. Real‐Time PCR was performed on a LightCycler 480 (Roche, Indianapolis, IN) with cycling parameters of 10 min initial denaturation at 95°C, followed by 45 cycles at 95°C for 15 s and 1 min at 60°C. Each reaction was performed in triplicate and normalized to the endogenous 18S and Act‐b gene expression. The CT value of each well was determined using the LightCycler 480 software, and the average of the triplicates was calculated. The relative quantification was determined by the ΔΔCT method (Livak and Schmittgen, 2001). TaqMan Gene Expression Assays used are as follows: mTrpm8: Mm01299593_m1; mTrpm8: Mm00454566_m1; mTRPV4: Mm00499025_m1; mActb: Mm00607939_s1; 18SrRNA: Mm03928990_g1; TNF‐α: Mm99999068_m1; IL‐1b: Mm00434228_m1; IFM‐g: Mm99999071_m1; and IL‐6: Mm00446190_m1.
Cell culture and calcium influx imaging
HEK293T cells were cultured in DMEM and supplemented with 10% FBS, 100 units·mL−1 penicillin and 0.1 mg·mL−1 streptomycin. Cells were plated on poly‐D‐lysine‐coated 100 mm tissue culture plastic dishes and grown overnight to 60–70% confluency. The cells were transiently transfected with human TRPM8 plasmid DNA (Origene Technologies, Rockville, MD, USA, Cat. RC220615, Genbank NM_024080) using Fugene 6 transfection reagent and Opti‐MEM according to manufacturer's protocols for 16–24 h. Cells were re‐suspended and plated onto poly‐D‐lysine‐coated 96‐well plates at 100 000 cells per well (100 μL per well) and allowed to grow for another 16–24 h prior to the experiments. Cells were maintained as monolayers in a 5% CO2 incubator at 37°C.
Cells plated in 96‐well plates (Krystal black walled plates, Genesee Scientific) were loaded with FLIPR Calcium 6 no wash dye for 2 h (Molecular Devices) according to manufacturer's instructions. In preliminary experiments, we determined that the optimal dye concentration was one‐third of that suggested by the manufacturer. After 2 h, the plate was transferred to a FlexStation III benchtop scanning fluorometer chamber. Fluorescence measurements were performed at 37°C. The cells were excited at 485 nm, and Ca2+‐bound FLIPR Calcium 6 emission was recorded at 525 nm at every 1.62 s intervals. After recording baseline fluorescence for 18 s, 50 μL of a 5× concentration of eucalyptol or 2‐OH‐1,8, cineole was added to the cells yielding a final volume of 250 μL per well; the fluorescence was monitored for an additional 100 s. The FLIPR Calcium 6 dye fluorescence was expressed as Fmax − F0, where Fmax is the maximum and F0 is the basal fluorescence measured in each well. The EC50 values and their 95% confidence intervals for eucalyptol and 2‐OH‐1,8 cineole stimulation of calcium influx were determined by non‐linear regression analysis with a three‐parameter logistic equation (Graphpad Prism software).
Data and statistical analysis
The data and statistical analysis in this study comply with the recommendations on experimental design and analysis in pharmacology (Curtis et al., 2015). Data in bar graphs are expressed as means ± SEM. Statistical comparisons were made between groups using Student's t‐test (for comparison between two groups) or one‐way ANOVA (for comparison among ≥3 groups) followed by Tukey's post hoc test, with values of P < 0.05 considered significant.
Materials
Eucalyptol (1,8‐cineol) was purchased from Acros Organics (Pittsburgh, PA, USA), LPS (Escherichia coli, 0111:B4) from Calbiochem (Darmstadt, Germany), 2‐hydroxy‐1,8‐cineol (CAS 60761–00‐4) from Alfa Chemistry (Stony Brook, NY, USA), Ibuprofen from Sigma (St. Louis, MO, USA), CFA from Rockland Immunochemicals (Gilbertsville, PA, USA) and corn oil from Sigma (C‐8267).
Results
Inhibition of tissue oedema by eucalyptol in the mouse CFA model
Eucalyptol‐containing medicinal preparations are often used to treat inflammatory tissue swelling and pain. We therefore examined the effects of eucalyptol in the mouse CFA model in which CFA is injected into the mouse hind paw, eliciting tissue oedema and hypersensitivity to mechanical stimulation. Changes in mouse paw volume were determined 2, 4, 24 and 48 h after CFA injection (Figure 1A). For eucalyptol treatment, the dose (30, 100 or 300 mg·kg−1, i.p.) and regimen were chosen based on published protocols effective in rodent inflammation models (Santos et al., 2001, 2004; Zhao et al., 2014). Animals were treated with eucalyptol 30 min before CFA injection and again 6, 24, 30 and 48 h after CFA injection for a total of five times for each group (Figure 1A). Injection of CFA induced robust swelling lasting until the endpoint (48 h) of our observations (Figure 1B). Comparison of paw volumes measured 2 h after CFA injection showed that 100 and 300 mg·kg−1 eucalyptol effectively reduced paw swelling (Figure 1B). The higher eucalyptol dose (300 mg·kg−1) suppressed tissue swelling throughout the 48 h time course, so we chose this dose for the subsequent experiments (Figure 1B).
Figure 1.
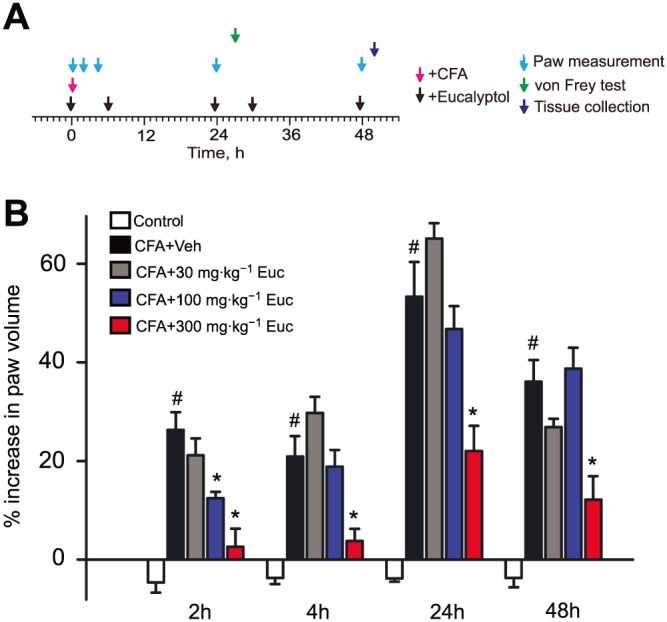
Effects of eucalyptol on CFA‐induced paw swelling in mice. (A) Schedule of treatment and experimental procedures. Paw oedema (% increase in paw volume) was measured by plethysmography directly (0 h), 2, 4, 24 and 48 h after injection of CFA (10 μL per paw) into the hind paw. Vehicle (corn oil), eucalyptol (30, 100 or 300 mg·kg−1) or ibuprofen (30 mg·kg−1) were injected (i.p.) 30 min before testing time points at 0, 24 and 48 h and in between at 6 and 30 h after CFA injection (total of five injections). (B) Percent increase in paw volume of CFA‐injected mice treated (i.p.) with eucalyptol (Euc) doses of 30, 100 or 300 mg·kg−1 or vehicle (Veh). The control group (Control) received saline injection into the hind paw instead of CFA and treatment with vehicle (corn oil) thereafter. n = 6–8 mice per group. # P < 0.05, significantly different from control group; *P < 0.05, significantly different from CFA + Veh group, one‐way ANOVA.
TRPM8 channels are essential for the anti‐inflammatory effects of eucalyptol in the CFA model
After establishing the efficacious dose for eucalyptol in the CFA model, we compared eucalyptol's anti‐inflammatory and analgesic effects in wild‐type and Trpm8−/− mice. Trpm8−/− mice developed a similar degree of hind paw swelling upon CFA injection as wild‐type mice (Figure 2A, B). However, eucalyptol (300 mg·kg−1, i.p.) was completely ineffective for suppressing paw swelling in TRPM8 channel‐deficient mice (Figure 2A, B, P > 0.05). In contrast to eucalyptol, ibuprofen (30 mg·kg−1, i.p.) was equally effective in suppressing paw swelling in wild‐type and Trpm8−/− mice, suggesting that other anti‐inflammatory pathways remain intact in mice lacking TRPM8 channels (Figure 2B, C). We also compared the weights of skin punch biopsies obtained from CFA‐injected hind paws of wild‐type and Trpm8−/− mice, excised 50 h after CFA injection and 2 h after the last treatment injection. Both eucalyptol and ibuprofen suppressed the increase in biopsy weights in wild‐type mice, whereas eucalyptol was ineffective (P > 0.05) in skin biopsies from Trpm8−/− mice (Figure 2D).
Figure 2.
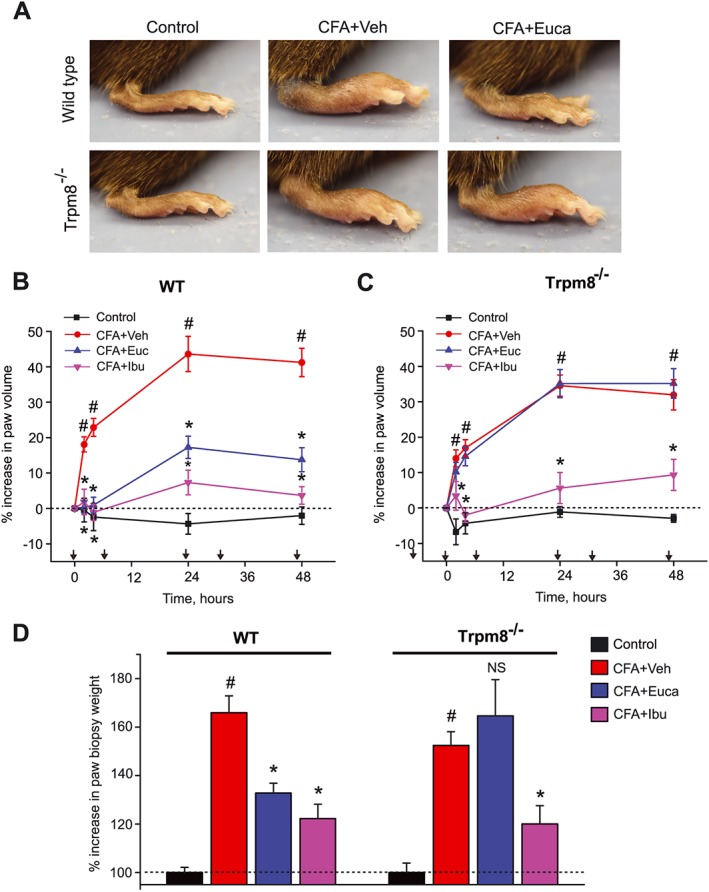
Comparison of the effects of eucalyptol on CFA‐induced paw oedema in wild‐type and Trpm8−/− mice, with animals treated as in Figure 1A. (A) Representative photographs of CFA‐induced paw oedema in wild‐type (top row) or Trpm8−/− mice (bottom row) injected i.p. with vehicle (Veh, corn oil) or 300 mg·kg−1 eucalyptol (Euc), taken 50 h after intraplantar CFA injection. (B) Time course of CFA‐induced paw oedema in wild‐type mice, treated with vehicle, eucalyptol (300 mg·kg−1) or the reference drug ibuprofen (Ibu, 30 mg·kg−1). Black arrows indicate the time points of treatment. (C) Time course of CFA‐induced paw oedema in Trpm8−/− mice. (D) Comparisons of weights of paw biopsies from wild‐type and Trpm8−/− mice of control group (no CFA), CFA groups treated with vehicle, eucalyptol or ibuprofen respectively. Tissues were collected 50 h after CFA injection from mice tested in Figure 2B, C. n = 6–8 mice per group. # P < 0.05, significantly different from control group, *P < 0.05, significantly different from CFA + Veh group, NS, no significance (P > 0.05), one‐way ANOVA.
Mice develop robust mechanical allodynia in the CFA model. Using von Frey hair analysis, we tested whether eucalyptol would diminish mechanical allodynia in this model, and whether this effect is altered in Trpm8−/− mice. Eucalyptol treatment (regimen as above, Figure 1A) strongly reduced paw withdrawal thresholds to a similar degree as ibuprofen, measured 24 h after CFA injection (Figure 3). In Trpm8−/− mice, eucalyptol was ineffective (P > 0.05), while ibuprofen retained its analgesic effect (Figure 3).
Figure 3.
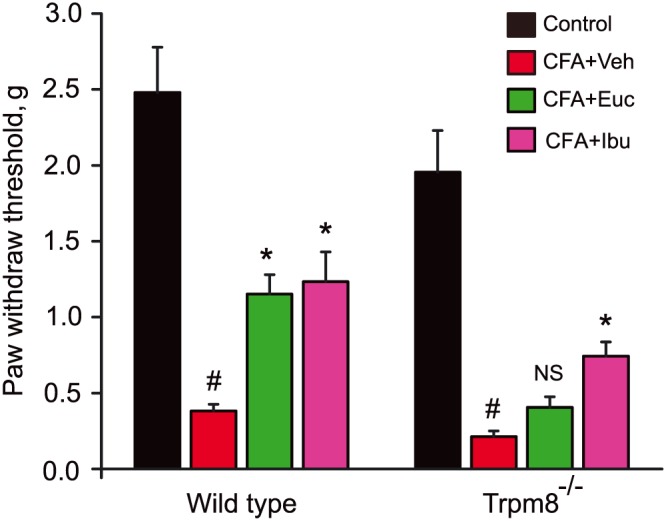
Effects of eucalyptol on CFA‐induced mechanical allodynia in wild‐type and Trpm8−/− mice, measured by von Frey hair analysis (administration as in Figure 1A). Testing was performed 26 h after CFA injection, 2 h after the mice received the third treatment of vehicle (Veh, corn oil), eucalyptol (Euc, 300 mg·kg−1) or ibuprofen (Ibu, 30 mg·kg−1). # P < 0.05, significantly different from control group; *P < 0.05, significantly different from CFA + Veh group, NS, no significance (P > 0.05), one‐way ANOVA. n = 6–12 mice per group.
CFA also triggers the production of key pro‐inflammatory cytokines. We examined the effects of eucalyptol on cytokine levels in hind paw skin biopsies from the CFA‐treated mice. Using ELISA, we detected markedly increased levels of TNF‐α, IL‐1β and IL‐6 in hind paw skin homogenates of CFA‐treated wild‐type mice. Eucalyptol inhibited the production of these cytokines in wild‐type mice (Figure 4). In Trpm8−/− mice, CFA injection caused increases in TNF‐α, IL‐1β and IL‐6 to similar levels as observed in wild‐type mice. However, eucalyptol treatment (300 mg·kg−1, i.p.) did not reduce the levels of any of these inflammatory cytokines in TRPM8 channel‐deficient mice (Figure 4, P > 0.05). Ibuprofen significantly reduced the levels of IL‐1β, IL‐6 and TNF‐α in skin homogenates from both wild‐type and Trpm8−/− mice treated with CFA (Figure 4). Taken together, these results show that TRPM8 channels are crucial for both the anti‐inflammatory and analgesic effects of eucalyptol in the mouse CFA model.
Figure 4.
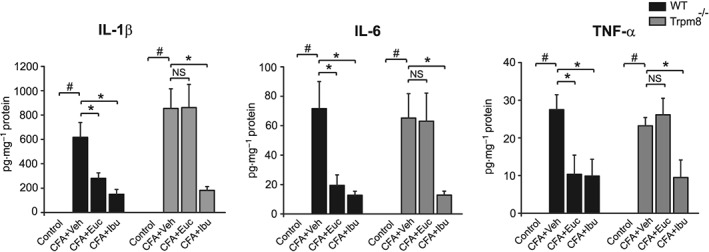
Influence of eucalyptol treatment on CFA‐induced inflammatory cytokines in hind paw skin of wild‐type and Trpm8−/− mice. Mice were treated (i.p.) with vehicle (Veh, corn oil), eucalyptol (Euc, 300 mg·kg−1) or ibuprofen (Ibu, 30 mg·kg−1) as in Figure 1A. Hind paw biopsies were collected 50 h after CFA injection, and levels of IL‐1β, IL‐6 and TNF‐α were determined by ELISA in tissue homogenates. # P < 0.05, significantly different from control group; *P < 0.05, significantly different from CFA + Veh group, NS, no significance (P > 0.05), one‐way ANOVA. n = 6–12 mice per group.
Mitigation of LPS‐induced pulmonary inflammation by eucalyptol is dependent on TRPM8 channels
Eucalyptol is frequently used to counteract respiratory inflammation and discomfort due to the common cold and other respiratory infections, as well as in asthma and COPD (Bastos et al., 2009). To study the actions of eucalyptol in the respiratory system, we used the mouse LPS model of ALI, a model in which LPS (10 μg in 40 μL of sterile saline) is instilled intranasally to mimic respiratory infections or environmental exposures to endotoxins. Respiratory inflammation was monitored by measurements of leukocyte concentrations in BAL, levels of cytokines in bronchoalveolar lavage fluid (BALF) and whole lung and MPO activity as a measure of neutrophil function. As described earlier (Zhao et al., 2014), eucalyptol (200 mg·kg−1) was injected 12 and 1 h prior to LPS instillation and also 8 and 23 h thereafter, with responses analysed 24 h after (Figure 5A). In wild‐type mice, eucalyptol treatment strongly reduced neutrophil counts in BALF of LPS‐exposed animals (Figure 5B). MPO enzyme activity was also diminished by eucalyptol treatment (Supporting Information Figure S1). Eucalyptol decreased protein concentrations and gene transcription of key pro‐inflammatory cytokines in BALF, including TNF‐α, IFN‐γ, KC, IL‐6 and G‐CSF, measured by ELISA in BALF and quantitative real time PCR of cDNA prepared from whole lungs after BAL and perfusion (Figures 5C and 6A). The anti‐inflammatory effects of eucalyptol in this model were absent in Trpm8−/− mice (Figures 5B, C and 6B).
Figure 5.
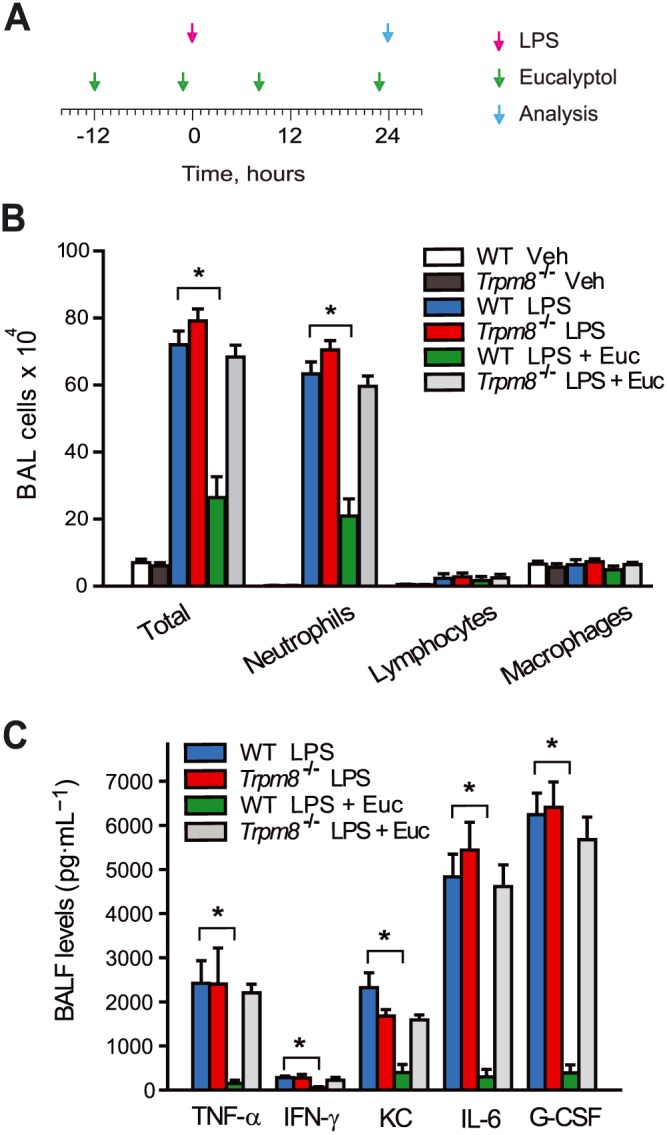
TRPM8‐dependent suppression of LPS‐induced pulmonary inflammation by eucalyptol. (A) LPS administration and eucalyptol treatment timeline. (B) Comparison of cell differential counts (total cells, neutrophils, lymphocytes and macrophages) in BALF of wild‐type or Trpm8−/− mice exposed to vehicle (saline) or LPS and injected with treatment vehicle (corn oil) or eucalyptol (200 mg·kg−1). (C) Cytokine levels in BALF of mice in Figure 5B, tested by bead array multiplex analysis. Protein concentrations are shown for TNF‐α, IFN‐g, KC, IL‐6 and G‐CCSF. (n = 4–8 mice per group). *P < 0.05, WT + LPS significantly different from WT + LPS + Eucalyptol.
Figure 6.
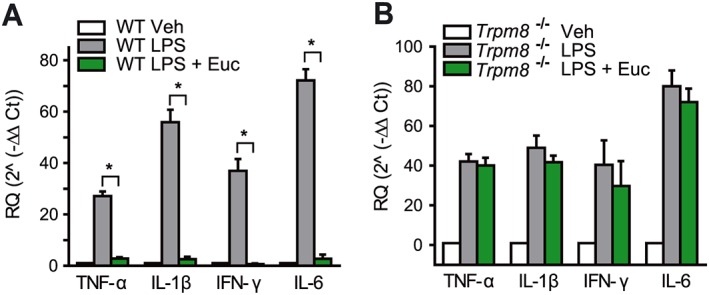
Cytokine transcripts analysed by quantitative real time qPCR in cDNA from whole lungs of LPS‐ and vehicle‐exposed mice treated and untreated with eucalyptol. (A) Impaired induction of cytokines in Trpm8−/− mice treated with eucalyptol before and after intranasal LPS administration. (B) Unaltered expression of cytokines in Trpm8−/− mice treated with eucalyptol before and after LPS exposure. (n = 4–6 mice per group). *P < 0.05, significantly different as indicated.
The treatment protocol for eucalyptol we used specified two pretreatment injections, administered 12 and 1 h prior to LPS instillation (Zhao et al., 2014). We then tested eucalyptol injections given after LPS instillation for their effects, injecting eucalyptol 1 and 6 h after LPS (200 mg·kg−1 each). This treatment protocol failed to suppress inflammatory responses (Supporting Information Figure S2, P > 0.05).
It remains unclear whether TRPM8 is transcribed and expressed locally in the lung where it may contribute to eucalyptol's anti‐inflammatory effects. Using two quantitative PCR hydrolysis probes covering key sequences of the cDNA of TRPM8 wild‐type and reported variants (Mm01299593_m1 and Mm00454566_m1), we found TRPM8 transcripts to be undetectable in cDNA prepared from mouse whole lungs (Figure 7, Supporting Information Figure S3). In contrast, cDNA of TRPV4, a TRP ion channel expressed in lung epithelial and vascular cells, was readily detectable (Figure 7). cDNA obtained from peripheral sensory ganglia (trigeminal and dorsal root ganglia) contained very high levels of TRPM8 transcripts, confirming the utility of the probes used and the highly selective expression of TRPM8 channels in sensory neurons (Figure 7, Supporting Information Figure S3). Levels of TRPA1 and TRPV1 transcripts in sensory ganglia did not show significant differences between wild‐type and Trpm8−/− mice (Supporting Information Figure S4, P > 0.05).
Figure 7.
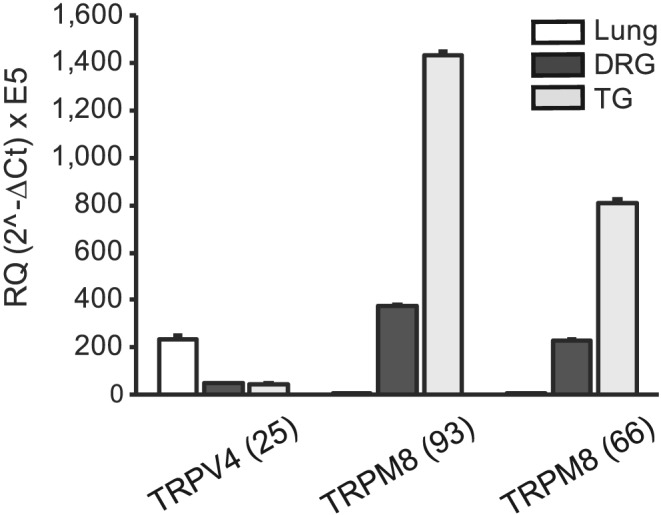
Comparison of TRPV4 and TRPM8 expression in lung and sensory ganglia. Averaged relative quantities (RQ) of TRPV4 and TRPM8 transcript levels were measured by real‐time Taqman PCR of cDNA from whole lung, DRG and TG of wild‐type mice. We used one TRPV4 Taqman probe (mTRPV4: Mm00499025_m1, 25) and two mTRPM8 probes covering different cDNA segments (mTrpm8: Mm01299593_m1, 93 and mTrpm8: Mm00454566_m1, 66). Actin was used as reference gene (n = 4 mice per group).
Dose–response relationship of human TRPM8 channels to eucalyptol and their sensitivity to the major eucalyptol metabolite, 2‐hydroxy‐1,8‐cineol
The effective dosages of eucalyptol used in clinical trials are lower than the doses required to suppress inflammation in mice in our study and in previous rodent studies. This may be due to different dose–response relationships of eucalyptol with rodent and human TRPM8 channels. While the EC50 of eucalyptol for mouse TRPM8 channels was determined as 7.7 ± 2.0 mM (Behrendt et al., 2004), the dose–response relationship of eucalyptol‐induced activation of human TRPM8 channels has not been published. We therefore used calcium fluorimetry to analyse eucalyptol's dose–response relationship for agonism of human TRPM8 channels expressed in HEK293T cells. Eucalyptol activated human TRPM8 channels with a much lower EC50 than that for the mouse channels (Figure 8A, C).
Figure 8.
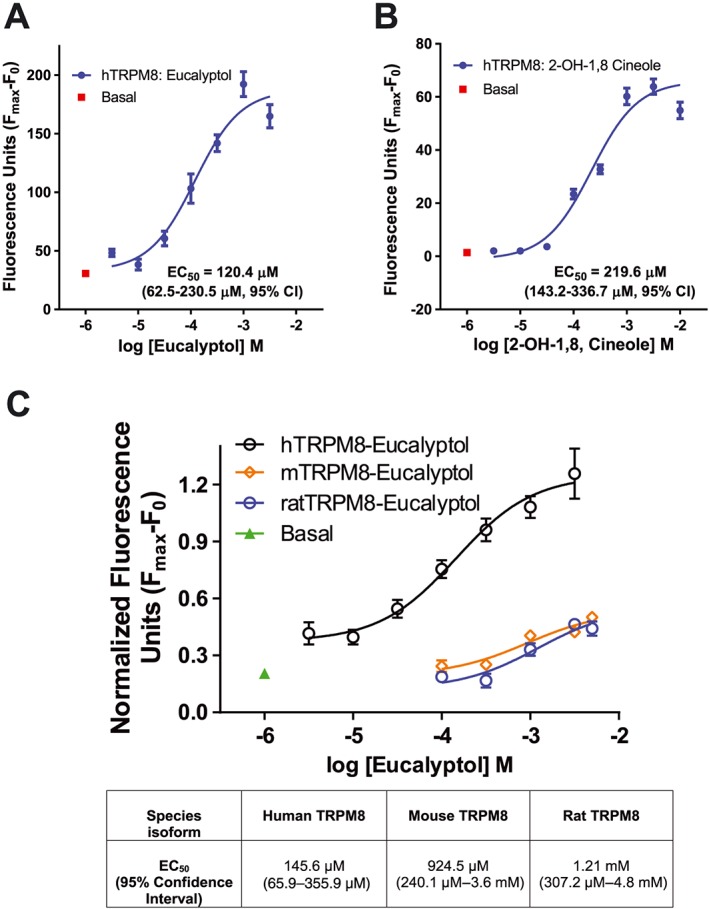
Comparison of eucalyptol sensitivities of TRPM8 channel species orthologues (human, mouse and rat) and determination of sensitivity of human TRPM8 channels to a eucalyptol metabolite, measured by Ca2+ fluorimetry in HEK293T cells. (A) Concentration response analysis of eucalyptol activation of human TRPM8 channels,with EC50 values as shown. (B) Concentration–response relationship of human TRPM8 channel activation by 2‐hydroxy‐1,8‐cineol. (C) Dose–response relationship of eucalyptol activation of human, mouse and rat TRPM8. The estimated EC50,values are shown in the Figure.
Eucalyptol is metabolized to compounds that persist in human tissues for a long time after dosing. Some of these metabolites resemble both eucalyptol and menthol, but it is not known whether they also activate TRPM8 channels. We examined the effects of 2‐hydroxy‐1,8‐cineol, the major metabolite of eucalyptol, on human TRPM8 channels by calcium fluorimetry. 2‐hydroxy‐1,8‐cineol activated hTRPM8 channels with an EC50 of 220 μM (140–340 μM, 95% CI) and lower efficacy than eucalyptol (Figure 8B). In contrast to human TRPM8 channels, the rodent orthologues (rat and mouse) responded to eucalyptol only at higher concentration, with EC50s around 1 mM (Figure 8C).
Discussion and conclusions
In the present study, we examined the anti‐inflammatory effects of eucalyptol in two mouse models of inflammation and explored the involvement of TRPM8 channels as potential mediators of the actions of eucalyptol. Eucalyptol effectively reduced paw oedema and cytokine production in the CFA‐induced mouse model of inflammatory pain. Eucalyptol also attenuated inflammation, leukocyte infiltration, MPO activity and cytokine production in the lungs of LPS‐exposed mice. Anti‐inflammatory effects of eucalyptol were completely absent in TRPM8 channel‐deficient mice, suggesting an important role of TRPM8 channels in this therapeutic activity.
Eucalyptol also exhibited strong analgesic effects. In the CFA model, eucalyptol significantly attenuated mechanical allodynia. This analgesic effect was abolished in TRPM8 channel‐deficient mice. This finding in a model of inflammatory pain correlates with our previous observations in an acute pain model induced by i.p. injection of acetic acid in which eucalyptol or L‐menthol effectively reduced nocifensive behaviour through a TRPM8 channel‐dependent mechanism (Liu et al., 2013b). While eucalyptol analgesia was absent in TRPM8 channel‐deficient mice, the analgesic and anti‐inflammatory effects of ibuprofen were retained, suggesting no general defect in analgesic pathways in this mouse strain.
In addition to TRPM8 channels, eucalyptol is known to interact with other TRP ion channels. For example, eucalyptol was found to inhibit mouse and human TRPA1 channels activated by the noxious agonist, mustard oil, in vitro, and may counteract pain induced by a TRPA1 channel agonist in human subjects, when applied topically (Takaishi et al., 2012; Liu et al., 2013b). However, this outcome can also be explained by eucalyptol acting on TRPM8 channels and exerting analgesia of TRPA1‐mediated noxious sensations. As demonstrated in several recent studies, TRPA1 channels are key mediators of CFA‐induced pain and inflammation, the model also used in the present study (da Costa et al., 2010; Fernandes et al., 2011; Lennertz et al., 2012). If TRPA1 channels were the direct anti‐inflammatory and analgesic targets of eucalyptol, eucalyptol's effects would have been retained in Trpm8−/− mice. However, this was not the case suggesting that, in the mouse, TRPM8 channels are the more likely analgesic and anti‐inflammatory pharmacological target of eucalyptol. This conclusion applies to the systemic administration with eucalyptol in the present study. It remains possible that other pathways, including inhibition of TRPA1 channels, could contribute to the analgesic effects of eucalyptol when large dosages are applied topically (Takaishi et al., 2012).
Assuming equal distribution without any metabolism in a mouse of 25 g body weight, the effective tissue levels of eucalyptol at the dose given in our present study would not exceed 3.2 mM. While this concentration will maintain activity at TRPM8 channels, eucalyptol is metabolized at a significant rate, effectively reducing its tissue levels. According to the dose–response analysis in the present study (mouse TRPM8 channels EC50 = 924.5 μM) and in other studies, this concentration would maintain significant activity at mouse TRPM8 channels (Behrendt et al., 2004). However, mice metabolize eucalyptol quite rapidly with an initial half‐life of 6 min for similar dosages (Kovar et al., 1987). These limitations may explain why eucalyptol is only effective in rodents when administered prophylactically, and not when administered when inflammation is already established.
In human patients, eucalyptol is administered orally or inhaled as a vapour. In clinical trials in bronchitis, COPD and asthma patients 200 mg were dosed three times per day for 12 weeks, reducing cough frequency, exacerbations and dyspnoea and improving lung function (Juergens et al., 2003; Worth et al., 2009; Fischer and Dethlefsen, 2013). Our dose–response analysis revealed that eucalyptol is a significantly more potent agonist of human TRPM8 channels (EC50 = 120.4 μM) than of mouse TRPM8 channels (EC50 = 924.5 μM) or rat TRPM8 channels (EC50 = 1.21 mM), suggesting that the lower dosages of eucalyptol used in human clinical trials may indeed be sufficient to activate human TRPM8 channels and counteract inflammation, even when administered when inflammation is already established. Eucalyptol and its metabolites can still be detected at significant levels in human body fluids, some hours after dosing (Kirsch and Buettner, 2013). Some of eucalyptol's major metabolites such as 2‐hydroxy‐1,8‐cineol used in the present study structurally resemble eucalyptol and menthol (Madyastha and Chadha, 1986; Horst and Rychlik, 2010). 2‐hydroxy‐1,8‐cineol activated human TRPM8 channels suggesting that persistent eucalyptol metabolites may prolong the anti‐inflammatory and analgesic effects of eucalyptol.
The mechanism through which activation of TRPM8 channels suppresses inflammation remains poorly understood. These ion channels are mainly expressed in the peripheral sensory neurons responding to cool thermal stimuli, menthol and other cooling natural or synthetic products (McKemy et al., 2002). Recent studies observed that input from TRPM8 channel‐expressing neurons excited central inhibitory interneurons that act as a gate to control behaviour elicited by noxious and pruritic stimuli (Kardon et al., 2014). Along the same lines, eucalyptol, through activation of TRPM8 channels, may strengthen a central neuronal gate controlling neuronally‐driven inflammatory responses elicited by C‐fibres expressing TRPA1 and TRPV1 channels that trigger the release of pro‐inflammatory neuropeptides and transmitters in the CFA model examined in our study and also promote pulmonary inflammatory responses (Petrus et al., 2007; Caceres et al., 2009; Lennertz et al., 2012). Subsets of TRPM8 channel‐positive fibres were shown to contain CGRP that, in some inflammation models, inhibits the activity of pro‐inflammatory immune cells (de Jong et al., 2015). While eucalyptol may also act locally – on TRPM8 channels expressed in cells in the inflamed tissue – our qPCR analysis was unable to detect significant levels of TRPM8 gene expression in pulmonary tissues or BALF cells, and no local TRPM8 transcripts were detected in the skin, the location of CFA injection (Peier et al., 2002). In contrast, TRPM8 transcript levels in DRG and trigeminal ganglia were very high. The data confirm previous studies detecting little TRPM8 expression outside the nervous system and prostate (McKemy et al., 2002; Fonfria et al., 2006; Jang et al., 2012). These findings favour a TRPM8‐dependent neuronal mechanism, mediating the broad anti‐inflammatory effects of eucalyptol in a number of tissues and disease models.
We have provided strong evidence identifying TRPM8 channels as essential for the anti‐inflammatory effects of eucalyptol. TRPM8 channels may therefore represent a promising pharmacological target to counteract inflammation, in addition to analgesia and acute irritation. The development of more potent and selective TRPM8 channel agonists may yield novel anti‐inflammatory therapeutic agents with lower toxicity and adverse effects than the current available options, and with improved compatibility with other treatments.
Author contributions
A.I.C., B.L., S.V.J. and S.A. completed the experiments. J.B.M. and S.E.J. designed the experiments and evaluated the results. A.I.C., B.L., S.V.J., J.B.M. and S.E.J. wrote the manuscript.
Conflict of interest
S.E.J. serves on the Scientific Advisory Board of Hydra Biosciences LLC (Cambridge, MA, USA), a developer of TRP ion channel inhibitors.
Declaration of transparency and scientific rigour
This Declaration acknowledges that this paper adheres to the principles for transparent reporting and scientific rigour of preclinical research recommended by funding agencies, publishers and other organisations engaged with supporting research.
Supporting information
Figure S1 Comparison of MPO activity levels in BALF from LPS‐exposed mice with or without treatment with eucalyptol. *P < 0.05 (n = 4 mice per group).
Figure S2 Lack of therapeutic effect of eucalyptol when administered after LPS. (A) Cell counts in BALF extracted from mice 24 h after intranasal exposure to LPS. Mice were injected i.p. with eucalyptol (200 mg·kg−1) 1 h after the intranasal LPS administration and a second dose was given 6 h later. (n = 4–6 mice per group) (B) BALF cytokine levels in the same mice as in (A). None of the differences were significant (P > 0.05).
Figure S3 TRPM8 transcription levels in lung. (A) Representative amplification fluorescence curves during RT‐PCR showing minimal amplification of TRPM8 transcript cDNA from mouse lung cDNA compared to TRPV4 and to TRPM8 transcript cDNA amplification from DRG and TG cDNA. (B) Comparison of TRPM8 transcript cDNA levels in lung, DRG and TG cDNA using TRPV4 as positive control (n = 4 mice per group).
Figure S4 Comparison of transcript levels of TRPA1 and TRPV1 in sensory neurons of wild‐type (grey) and Trpm8−/− mice, by qPCR of cDNAs extracted from dorsal root (DRG, left) or trigeminal (TG, right) ganglia. (n = 4 mice per group). None of the data were significantly different (P > 0.05).
Acknowledgements
Supported by grants from the National Heart Lung and Blood Institute (NHLBI, R01HL105635 and R01HL105635‐S1 to S.E.J. and J.B.M.), and the National Institute on Drug Abuse (NIDA, Yale Tobacco Center of Regulatory Science, P50DA036151 Project #1 to S.E.J.) of the National Institutes of Health (NIH) of the United States. Contents are solely the responsibility of the authors and do not necessarily represent the official views of the federal government of the United States.
Caceres, A. I. , Liu, B. , Jabba, S. V. , Achanta, S. , Morris, J. B. , and Jordt, S.‐E. (2017) Transient Receptor Potential Cation Channel Subfamily M Member 8 channels mediate the anti‐inflammatory effects of eucalyptol. British Journal of Pharmacology, 174: 867–879. doi: 10.1111/bph.13760.
References
- Alexander SPH, Catterall WA, Kelly E, Marrion N, Peters JA, Benson HE et al. (2015). The Concise Guide to PHARMACOLOGY 2015/16: Voltage‐gated ion channels. Br J Pharmacol 172: 5904–5941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barr A (1988). Traditional Bush Medicines: An Aboriginal Pharmacopoeia: Aboriginal Communities of the Northern Territory of Australia. Greenhouse Publications: Richmond, Victoria, Australia. [Google Scholar]
- Bastos VP, Brito TS, Lima FJ, Pinho JP, Lahlou S, Abreu Matos FJ et al. (2009). Inhibitory effect of 1,8‐cineole on guinea‐pig airway challenged with ovalbumin involves a preferential action on electromechanical coupling. Clin Exp Pharmacol Physiol 36: 1120–1126. [DOI] [PubMed] [Google Scholar]
- Bautista DM, Siemens J, Glazer JM, Tsuruda PR, Basbaum AI, Stucky CL et al. (2007). The menthol receptor TRPM8 is the principal detector of environmental cold. Nature 448: 204–208. [DOI] [PubMed] [Google Scholar]
- Bautista DM, Pellegrino M, Tsunozaki M (2013). TRPA1: a gatekeeper for inflammation. Annu Rev Physiol 75: 181–200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Behrendt HJ, Germann T, Gillen C, Hatt H, Jostock R (2004). Characterization of the mouse cold‐menthol receptor TRPM8 and vanilloid receptor type‐1 VR1 using a fluorometric imaging plate reader (FLIPR) assay. Br J Pharmacol 141: 737–745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caceres AI, Brackmann M, Elia MD, Bessac BF, del Camino D, D'Amours M et al. (2009). A sensory neuronal ion channel essential for airway inflammation and hyperreactivity in asthma. Proc Natl Acad Sci U S A 106: 9099–9104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL (1994). Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods 53: 55–63. [DOI] [PubMed] [Google Scholar]
- da Costa DS, Meotti FC, Andrade EL, Leal PC, Motta EM, Calixto JB (2010). The involvement of the transient receptor potential A1 (TRPA1) in the maintenance of mechanical and cold hyperalgesia in persistent inflammation. Pain 148: 431–437. [DOI] [PubMed] [Google Scholar]
- Curtis MJ, Bond RA, Spina D, Ahluwalia A, Alexander SP, Giembycz MA et al. (2015). Experimental design and analysis and their reporting: new guidance for publication in BJP. Br J Pharmacol 172: 3461–3471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dymock W, Warden CJH, Hooper DH (1972). Pharmacographia Indica; A History of the Principal Drugs of Vegetable Origin Met with in British India. Hamdard National Foundation for the Institute of Health and Tibbi Research: Karachi. [Google Scholar]
- Escalera J, von Hehn CA, Bessac BF, Sivula M, Jordt SE (2008). TRPA1 mediates the noxious effects of natural sesquiterpene deterrents. J Biol Chem 283: 24136–24144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandes ES, Russell FA, Spina D, McDougall JJ, Graepel R, Gentry C et al. (2011). A distinct role for transient receptor potential ankyrin 1, in addition to transient receptor potential vanilloid 1, in tumor necrosis factor alpha‐induced inflammatory hyperalgesia and Freund's complete adjuvant‐induced monarthritis. Arthritis Rheum 63: 819–829. [DOI] [PubMed] [Google Scholar]
- Fischer J, Dethlefsen U (2013). Efficacy of cineole in patients suffering from acute bronchitis: a placebo‐controlled double‐blind trial. Cough 9: 25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fonfria E, Murdock PR, Cusdin FS, Benham CD, Kelsell RE, McNulty S (2006). Tissue distribution profiles of the human TRPM cation channel family. J Recept Signal Transduct Res 26: 159–178. [DOI] [PubMed] [Google Scholar]
- Greiner JF, Muller J, Zeuner MT, Hauser S, Seidel T, Klenke C et al. (2013). 1,8‐Cineol inhibits nuclear translocation of NF‐kappaB p65 and NF‐kappaB‐dependent transcriptional activity. Biochim Biophys Acta 1833: 2866–2878. [DOI] [PubMed] [Google Scholar]
- Ha MA, Smith GJ, Cichocki JA, Fan L, Liu YS, Caceres AI et al. (2015). Menthol attenuates respiratory irritation and elevates blood cotinine in cigarette smoke exposed mice. PLoS One 10: e0117128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hendry ER, Worthington T, Conway BR, Lambert PA (2009). Antimicrobial efficacy of eucalyptus oil and 1,8‐cineole alone and in combination with chlorhexidine digluconate against microorganisms grown in planktonic and biofilm cultures. J Antimicrob Chemother 64: 1219–1225. [DOI] [PubMed] [Google Scholar]
- Hippocrates, Adams F (1886). The Genuine Works of Hippocrates. W. Wood and company: New York. [Google Scholar]
- Hirano S (1997). Quantitative time‐course profiles of bronchoalveolar lavage cells following intratracheal instillation of lipopolysaccharide in mice. Ind Health 35: 353–358. [DOI] [PubMed] [Google Scholar]
- Horst K, Rychlik M (2010). Quantification of 1,8‐cineole and of its metabolites in humans using stable isotope dilution assays. Mol Nutr Food Res 54: 1515–1529. [DOI] [PubMed] [Google Scholar]
- Jang Y, Lee Y, Kim SM, Yang YD, Jung J, Oh U (2012). Quantitative analysis of TRP channel genes in mouse organs. Arch Pharm Res 35: 1823–1830. [DOI] [PubMed] [Google Scholar]
- de Jong PR, Takahashi N, Peiris M, Bertin S, Lee J, Gareau MG et al. (2015). TRPM8 on mucosal sensory nerves regulates colitogenic responses by innate immune cells via CGRP. Mucosal Immunol 8: 491–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Juergens UR (2014). Anti‐inflammatory properties of the monoterpene 1.8‐cineole: current evidence for co‐medication in inflammatory airway diseases. Drug Res (Stuttg) 64: 638–646. [DOI] [PubMed] [Google Scholar]
- Juergens UR, Stober M, Schmidt‐Schilling L, Kleuver T, Vetter H (1998a). Antiinflammatory effects of euclyptol (1.8‐cineole) in bronchial asthma: inhibition of arachidonic acid metabolism in human blood monocytes ex vivo. Eur J Med Res 3: 407–412. [PubMed] [Google Scholar]
- Juergens UR, Stober M, Vetter H (1998b). Inhibition of cytokine production and arachidonic acid metabolism by eucalyptol (1.8‐cineole) in human blood monocytes in vitro. Eur J Med Res 3: 508–510. [PubMed] [Google Scholar]
- Juergens UR, Dethlefsen U, Steinkamp G, Gillissen A, Repges R, Vetter H (2003). Anti‐inflammatory activity of 1.8‐cineol (eucalyptol) in bronchial asthma: a double‐blind placebo‐controlled trial. Respir Med 97: 250–256. [DOI] [PubMed] [Google Scholar]
- Kardon AP, Polgar E, Hachisuka J, Snyder LM, Cameron D, Savage S et al. (2014). Dynorphin acts as a neuromodulator to inhibit itch in the dorsal horn of the spinal cord. Neuron 82: 573–586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehrl W, Sonnemann U, Dethlefsen U (2004). Therapy for acute nonpurulent rhinosinusitis with cineole: results of a double‐blind, randomized, placebo‐controlled trial. Laryngoscope 114: 738–742. [DOI] [PubMed] [Google Scholar]
- Keinan E, Alt A, Amir G, Bentur L, Bibi H, Shoseyov D (2005). Natural ozone scavenger prevents asthma in sensitized rats. Bioorg Med Chem 13: 557–562. [DOI] [PubMed] [Google Scholar]
- Kilkenny C, Browne W, Cuthill IC, Emerson M, Altman DG (2010). Animal research: reporting in vivo experiments: the ARRIVE guidelines. Br J Pharmacol 160: 1577–1579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim KY, Lee HS, Seol GH (2015). Eucalyptol suppresses matrix metalloproteinase‐9 expression through an extracellular signal‐regulated kinase‐dependent nuclear factor‐kappa B pathway to exert anti‐inflammatory effects in an acute lung inflammation model. J Pharm Pharmacol 67: 1066–1074. [DOI] [PubMed] [Google Scholar]
- Kirsch F, Buettner A (2013). Characterisation of the metabolites of 1,8‐cineole transferred into human milk: concentrations and ratio of enantiomers. Metabolites 3: 47–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kovar KA, Gropper B, Friess D, Ammon HP (1987). Blood levels of 1,8‐cineole and locomotor activity of mice after inhalation and oral administration of rosemary oil. Planta Med 53: 315–318. [DOI] [PubMed] [Google Scholar]
- Lennertz RC, Kossyreva EA, Smith AK, Stucky CL (2012). TRPA1 mediates mechanical sensitization in nociceptors during inflammation. PLoS One 7: e43597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liapi C, Anifandis G, Chinou I, Kourounakis AP, Theodosopoulos S, Galanopoulou P (2007). Antinociceptive properties of 1,8‐Cineole and beta‐pinene, from the essential oil of Eucalyptus camaldulensis leaves, in rodents. Planta Med 73: 1247–1254. [DOI] [PubMed] [Google Scholar]
- Liu B, Escalera J, Balakrishna S, Fan L, Caceres AI, Robinson E et al. (2013a). TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB J 27: 3549–3563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu B, Fan L, Balakrishna S, Sui A, Morris JB, Jordt SE (2013b). TRPM8 is the principal mediator of menthol‐induced analgesia of acute and inflammatory pain. Pain 154: 2169–2177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Livak KJ, Schmittgen TD (2001). Analysis of relative gene expression data using real‐time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 25: 402–408. [DOI] [PubMed] [Google Scholar]
- Madyastha KM, Chadha A (1986). Metabolism of 1,8‐cineole in rat: its effects on liver and lung microsomal cytochrome P‐450 systems. Bull Environ Contam Toxicol 37: 759–766. [DOI] [PubMed] [Google Scholar]
- McGrath JC, Lilley E (2015). Implementing guidelines on reporting research using animals (ARRIVE etc.): new requirements for publication in BJP. Br J Pharmacol 172: 3189–3193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKemy DD, Neuhausser WM, Julius D (2002). Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 416: 52–58. [DOI] [PubMed] [Google Scholar]
- Meister R, Wittig T, Beuscher N, de Mey C (1999). Efficacy and tolerability of myrtol standardized in long‐term treatment of chronic bronchitis. A double‐blind, placebo‐controlled study. Study Group Investigators. Arzneimittelforschung 49: 351–358. [DOI] [PubMed] [Google Scholar]
- Nascimento NR, Refosco RM, Vasconcelos EC, Kerntopf MR, Santos CF, Batista FJ et al. (2009). 1,8‐Cineole induces relaxation in rat and guinea‐pig airway smooth muscle. J Pharm Pharmacol 61: 361–366. [DOI] [PubMed] [Google Scholar]
- Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Story GM et al. (2002). A TRP channel that senses cold stimuli and menthol. Cell 108: 705–715. [DOI] [PubMed] [Google Scholar]
- Petrus M, Peier AM, Bandell M, Hwang SW, Huynh T, Olney N et al. (2007). A role of TRPA1 in mechanical hyperalgesia is revealed by pharmacological inhibition. Mol Pain 3: 40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Proudfoot CJ, Garry EM, Cottrell DF, Rosie R, Anderson H, Robertson DC et al. (2006). Analgesia mediated by the TRPM8 cold receptor in chronic neuropathic pain. Curr Biol 16: 1591–1605. [DOI] [PubMed] [Google Scholar]
- Ramachandran R, Hyun E, Zhao L, Lapointe TK, Chapman K, Hirota CL et al. (2013). TRPM8 activation attenuates inflammatory responses in mouse models of colitis. Proc Natl Acad Sci U S A 110: 7476–7481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rocha Caldas GF, Oliveira AR, Araujo AV, Lafayette SS, Albuquerque GS, Silva‐Neto Jda C et al. (2015). Gastroprotective mechanisms of the monoterpene 1,8‐cineole (eucalyptol). PLoS One 10: e0134558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sadlon AE, Lamson DW (2010). Immune‐modifying and antimicrobial effects of eucalyptus oil and simple inhalation devices. Altern Med Rev 15: 33–47. [PubMed] [Google Scholar]
- Santos FA, Rao VS (2000). Antiinflammatory and antinociceptive effects of 1,8‐cineole a terpenoid oxide present in many plant essential oils. Phytother Res 14: 240–244. [DOI] [PubMed] [Google Scholar]
- Santos FA, Rao VS (2001). 1,8‐cineol, a food flavoring agent, prevents ethanol‐induced gastric injury in rats. Dig Dis Sci 46: 331–337. [DOI] [PubMed] [Google Scholar]
- Santos FA, Silva RM, Tome AR, Rao VS, Pompeu MM, Teixeira MJ et al. (2001). 1,8‐cineole protects against liver failure in an in‐vivo murine model of endotoxemic shock. J Pharm Pharmacol 53: 505–511. [DOI] [PubMed] [Google Scholar]
- Santos FA, Silva RM, Campos AR, De Araujo RP, Lima Junior RC, Rao VS (2004). 1,8‐cineole (eucalyptol), a monoterpene oxide attenuates the colonic damage in rats on acute TNBS‐colitis. Food Chem Toxicol 42: 579–584. [DOI] [PubMed] [Google Scholar]
- Santos FB, Nagato LK, Boechem NM, Negri EM, Guimaraes A, Capelozzi VL et al. (2006). Time course of lung parenchyma remodeling in pulmonary and extrapulmonary acute lung injury. J Appl Physiol (1985) 100: 98–106. [DOI] [PubMed] [Google Scholar]
- Southan C, Sharman JL, Benson HE, Faccenda E, Pawson AJ, Alexander SP et al. (2016). The IUPHAR/BPS guide to PHARMACOLOGY in 2016: towards curated quantitative interactions between 1300 protein targets and 6000 ligands. Nucleic Acids Res 44: D1054–D1068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takaishi M, Fujita F, Uchida K, Yamamoto S, Sawada Shimizu M, Hatai Uotsu C et al. (2012). 1,8‐cineole, a TRPM8 agonist, is a novel natural antagonist of human TRPA1. Mol Pain 8: 86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takaishi M, Uchida K, Fujita F, Tominaga M (2014). Inhibitory effects of monoterpenes on human TRPA1 and the structural basis of their activity. J Physiol Sci 64: 47–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinuela‐Fernandez I, Sun L, Jerina H, Curtis J, Allchorne A, Gooding H et al. (2014). The TRPM8 channel forms a complex with the 5‐HT(1B) receptor and phospholipase D that amplifies its reversal of pain hypersensitivity. Neuropharmacology 79: 136–151. [DOI] [PubMed] [Google Scholar]
- Willis DN, Liu B, Ha MA, Jordt SE, Morris JB (2011). Menthol attenuates respiratory irritation responses to multiple cigarette smoke irritants. FASEB J 25: 4434–4444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worth H, Schacher C, Dethlefsen U (2009). Concomitant therapy with cineole (eucalyptole) reduces exacerbations in COPD: a placebo‐controlled double‐blind trial. Respir Res 10: 69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao C, Sun J, Fang C, Tang F (2014). 1,8‐cineol attenuates LPS‐induced acute pulmonary inflammation in mice. Inflammation 37: 566–572. [DOI] [PubMed] [Google Scholar]
- Zhou JY, Wang XF, Tang FD, Zhou JY, Lu GH, Wang Y et al. (2007). Inhibitory effect of 1,8‐cineol (eucalyptol) on Egr‐1 expression in lipopolysaccharide‐stimulated THP‐1 cells. Acta Pharmacol Sin 28: 908–912. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1 Comparison of MPO activity levels in BALF from LPS‐exposed mice with or without treatment with eucalyptol. *P < 0.05 (n = 4 mice per group).
Figure S2 Lack of therapeutic effect of eucalyptol when administered after LPS. (A) Cell counts in BALF extracted from mice 24 h after intranasal exposure to LPS. Mice were injected i.p. with eucalyptol (200 mg·kg−1) 1 h after the intranasal LPS administration and a second dose was given 6 h later. (n = 4–6 mice per group) (B) BALF cytokine levels in the same mice as in (A). None of the differences were significant (P > 0.05).
Figure S3 TRPM8 transcription levels in lung. (A) Representative amplification fluorescence curves during RT‐PCR showing minimal amplification of TRPM8 transcript cDNA from mouse lung cDNA compared to TRPV4 and to TRPM8 transcript cDNA amplification from DRG and TG cDNA. (B) Comparison of TRPM8 transcript cDNA levels in lung, DRG and TG cDNA using TRPV4 as positive control (n = 4 mice per group).
Figure S4 Comparison of transcript levels of TRPA1 and TRPV1 in sensory neurons of wild‐type (grey) and Trpm8−/− mice, by qPCR of cDNAs extracted from dorsal root (DRG, left) or trigeminal (TG, right) ganglia. (n = 4 mice per group). None of the data were significantly different (P > 0.05).