

Interest in the biology of the intervertebral disc has grown significantly over the past 2 decades, driven mainly by studies aimed at developing biological therapies for repairing degenerate discs (Alini et al. 2002, Sakai and Grad 2015). Most interest has focused on cellular therapies, where cells, capable of synthesizing appropriate disc tissue, are implanted into the damaged tissue to replace resident cells that have died or have acquired a degenerative phenotype. This appears to be an attractive strategy, and has led to a significant increase in information about disc cellular biology. It follows the approach used clinically for repairing damaged cartilage (Hunziker et al. 2015); however, cell therapy for the disc faces more obstacles than that for cartilage repair and has not yet entered routine clinical practice.
In this review, we discuss some of the challenges in successful cellular repair of the disc. We first review the function, organization, and composition of a normal disc, outline the changes that occur in degeneration, and consider how these might influence function. We then summarize cell therapy approaches to repairing the disc in relation to the choice of cells and cell support. We outline the challenges facing the implanted cells in the degenerate disc, and ask whether these therapies can be evaluated in animal models. Finally, we outline the important, but often neglected, problem of patient selection.
The disc is complex in structure, composition, and function. What are we aiming to repair/regenerate?
The normal disc
Morphology and composition
The intervertebral discs are large load-bearing cartilaginous tissues that lie interspersed between the bony vertebral bodies. Morphologically, the disc appears to consist of 2 main regions (Figure 1), with an inner, more gelatinous region, the nucleus pulposus (or nucleus), encircled by a stiffer, collagenous annulus fibrosus (or annulus), consisting of concentric lamellae. The nucleus and annulus are separated from the bone by a thin (approx. 1-mm) layer of hyaline cartilage, the cartilage endplate; annulus insertions anchor the disc to the bone (Nosikova et al. 2012). The normal disc is virtually avascular, with blood vessels and nerves being found only in the periphery of the annulus.
Figure 1.
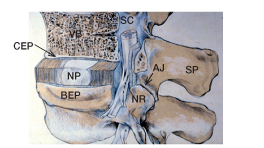
A schematic view of the vertebral joint. Here it is partly cut away to show the annulus fibrosus (AF) surrounding the nucleus pulposus (NP) of the intervertebral disc, the cartilaginous endplate (CEP) and bony endplate (BEP) interspersed between the disc and vertebral body (VB), and the spinal canal (SC) lying behind the vertebral bodies and the disc. The spinal canal—surrounded by the discs, the spinal processes (SP), and apophyseal joints (AJ)—encloses the spinal cord which gives rise to the nerve roots (NR) running adjacent to the posterior portion of the disc. (Adapted from Urban and Roberts 1986).
The composition and organization of the macromolecules that make up its extracellular matrix enable the disc to fulfill its mechanical role. Fibrillar collagens provide the structural framework of the disc (Eyre et al. 1991). The collagen network of the nucleus is formed from fine fibrils of (mainly type-II) collagen. Parallel bundles of fibrils (mainly type-I), running obliquely between the adjacent vertebral bodies, form the concentric lamellae of the annulus (Takeda 1975, Pezowicz et al. 2006). The lamellae are held together by elastic proteins (Yu et al. 2015), which help to give the disc its flexibility. Aggrecan, the other major macromolecular component, is a large polyanionic proteoglycan that imparts a high osmotic pressure to the disc matrix (Sivan et al. 2006); the matrix thus tends to imbibe water, inflating the collagen network until the osmotic swelling pressure balances the applied load. Apart from aggrecan and collagens, the disc matrix also contains a large number of other proteins (Figure 2A), which, although present in low concentrations, are also important in regulating the stability and function of the disc matrix (Feng et al. 2006).
Figure 2.
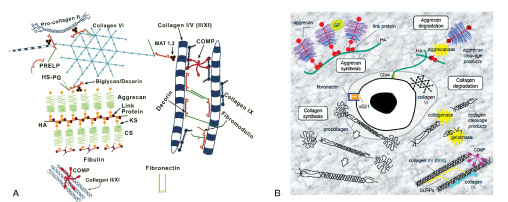
A. Schematic illustration of assemblies of matrix proteins in the intervertebral disc. Aggrecan monomer is synthesized intracellularly and secreted into the ECM where it forms supramolecular aggregates with HA that are stabilized by link proteins. Collagen synthesis involves removal of the N- and C-terminal propeptides from procollagen to generate tropocollagen which self-assembles into polymeric collagen fibrils. Cartilage oligomeric matrix protein (COMP) acts as a catalyst in collagen fibrillogenesis, and small leucine-rich proteoglycans (SLRPs; e.g. decorin, biglycan, fibromodulin, lumican) and collagen IX regulate fibril thickness and interfibrillar spacing. CS: chondroitin sulfate; KS: keratan sulfate; HA: hyaluronan; HS-PG, heparan sulfate proteoglycan; MAT: matrilin; PRELP: proline arginine-rich end leucine-rich repeat protein. (Reproduced from Feng et al. (2006) with permission). B. Schematic illustration depicting the synthesis and degradation of the disc extracellular matrix. In normal, healthy discs, there is a fine balance between matrix synthesis, assembly, and turnover, which becomes perturbed during disc degeneration. Aggrecanases (ADAMTS-4 and -5) within the ECM cause cleavage and fragmentation of the aggrecan core protein. Degradation of collagen fibrils occurs through the activity of collagenases (MMP-1 and -13) and gelatinases (MMP-2 and -9). α5β1: α5β1 integrin (fibronectin receptor); CS: chondroitin sulfate; CD44: hyaluronic acid receptor; G1, G2, and G3: globular domains of aggrecan; GF growth factors: cytokines and other bioactive signaling molecules; HA: hyaluronic acid; KS, keratan sulfate. (Reproduced from Sivan et al. (2014a) with permission).
Disc cells
The human disc contains a small population of resident cells (Pattappa et al. 2012) that make and maintain the disc’s macromolecules. The cells also produce proteases that are capable of degrading all matrix components. In a healthy disc, the rates at which the macromolecules are made and broken down are in balance (Figure 2B), but because of the low cell density, the turnover in human discs is very slow (Sivan et al. 2014a).
The cell type—and hence the composition—of the matrix synthesized varies across the disc and changes with age. The nucleus pulposus of all mammals is initially populated by clusters of large notochordal cells that produce a highly hydrated, aggrecan-rich, collagen-poor matrix. In humans and in some other species, the cell phenotype changes during growth, with the notochord cells being replaced by several phenotypically distinct but poorly characterized subpopulations of chondrocyte-like cells (Molinos et al. 2015). These chondrocyte-like cells produce matrix that becomes more collagenous and less hydrated during development in humans. In the outer annulus, fibroblast-like cells synthesize the highly organized collagen-rich lamellae. The disc also contains a small number of progenitor cells that are potentially able to differentiate into the appropriate disc cell phenotypes (Henriksson et al. 2009, Sakai et al. 2012, Gruber et al. 2016). Little is known about the cells of the cartilage endplate.
The degenerate disc
Disc degeneration is a loose term that encompasses progressive biochemical, cellular, and structural changes to the disc—with consequent changes in its load-bearing properties. Although little is understood about the factors that initiate disc degeneration, the process appears to be driven by changes in the behavior of its resident cells, which begin to increase the production of proteases and reduce production of the matrix macromolecules. Hence, macromolecules are degraded and lost from the disc at a faster rate than they can be replaced.
B. MRIs showing discs at different stage of Pfirrmann degeneration grade. Grading is based on signal intensity, distinction between nucleus and annulus, degree of homogeneity of disc structure, and loss of disc height. Features which are apparent morphologically (Figure 3a), such as fissures, changes in the endplate and even herniations are not taken into account in this grading scheme (adapted from Pfirrmann et al. 2001)
Figure 3.
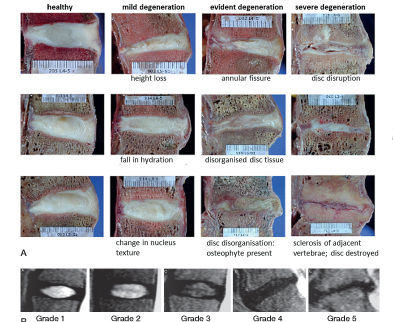
A. Sagittal sections of human lumbar intervertebral discs at various stages of degeneration. Features such as height loss, fall in water content, annular tears, osteophytes, and endplate sclerosis observed at different stages of degeneration are indicated (Adapted from Galbursera et al. 2014).
Information on the changes in disc composition and organization with degeneration has been obtained from examination of discs taken at autopsy or removed at surgery (Lyons et al. 1981, Boos et al. 2002, Roberts et al. 2006). Degenerate discs have high concentrations of proteases that tend to degrade the macromolecules of the disc, particularly aggrecan—the concentration of which falls on disc degeneration (Sivan et al. 2014b) (Figure 2B); degenerate discs thus retain less water and lose it faster under load. As the disc degrades and becomes more dehydrated, the lamellae become disorganized and the disc loses structural integrity, with formation of fissures and defects at the bone-disc interface (Figure 3A). The cartilaginous endplate tends to calcify, decreasing nutrient transport to the cells; many of them become senescent and die (Kletsas 2009). Blood vessels and nerves invade the previously avascular, aneural disc along with inflammatory cells such as macrophages. The changes seen in disc degeneration vary from individual to individual, may start early in life, appear to be strongly genetic (Boos et al 2002, Battié et al. 2009), and are an ongoing process with the severity and number of degenerative changes increasing with age.
Functional changes in disc degeneration
The morphological and biochemical changes resulting from disc degeneration influence the mechanical behavior of the disc, and therefore of the whole spinal column (Adams 2004, Galbusera et al. 2014, Von Forell et al. 2015, Muriuki et al. 2016). Degeneration, with its loss of aggrecan, results in a fall in hydration and a reduction in disc height, an increase in disc bulge, and a change in stiffness. Loss of the integrity of the disc results in instability of the spinal motion segment, possibly leading to spondylolisthesis. Inappropriate loads are thus transmitted to other spinal structures such as the facet joints, which may become osteoarthritic—and also to the posterior ligaments, which may thicken, leading to spinal stenosis. Profound degenerative changes in the spinal column triggered by a series of these degenerative events may end in onset of complex spinal deformities.
Diagnosis of disc degeneration in vivo
In vivo, disc degeneration is detected using magnetic resonance imaging (MRI). It is often classified using MRI scores (Pfirrmann et al. 2001) (Figure 3B), based on changes in disc height and signal intensity without considering other degradative features. MRI grade-3 discs, for instance, may include discs with very different degrees of endplate irregularity, disc bulge, or radial or circumferential tears (Figure 3A). Currently, degenerative changes at the tissue and cellular level cannot be detected non-invasively.
What degenerative changes are the biological therapies aimed at repairing?
Currently, disc cell therapies are mostly aimed at restoring macromolecular components, with aggrecan in the nucleus being the major focus, as the mechanical consequences of its loss are very apparent. However, while desirable mechanical properties for repair have been defined (Cortes et al. 2014), little is known about what other components of the complex matrix—apart from collagens—are necessary for functional repair. Moreover, while restoration of nucleus hydration is the aim of many studies, fewer studies have examined repair of the annulus (Sakai and Grad 2015) or cartilage endplate (Bendtsen et al. 2011, Nosikova et al. 2012), yet the integrity of these structures is also essential for disc health. Thus, would functional and stable cellular disc repair require an approach that integrates all disc regions (Nosikova et al. 2012)?
Clinical trials of cellular therapies for intervertebral disc repair
| Title | Place | ClinicalTrials.gov identifier | Status |
|---|---|---|---|
| Autologous adipose derived stem cell therapy for intervertebral disc degeneration | Bundang CHA Hospital, Korea | NCT02338271 | Recruiting |
| Treatment of degenerative disc disease with allogeneic mesenchymal stem cells | Hospital Clinico Universitario, Valladolid, Spain | NCT01860417 | Ongoing, not recruiting |
| Autologous adipose tissue derived mesenchymal stem cell transplantation in patient with lumbar intervertebral disc degeneration | Biostar, Korea University Anam Hospital | NCT01643681 | Unknown |
| Safety and preliminary efficacy study of mesenchymal precursor cells (MPCs) in subjects with lumbar back pain | Mesoblast Ltd. | NCT01290367 | Completed but no results posted |
| Safety and efficacy study of rexlemestrocel-L (viz. allogenic MSCs) in subjects with chronic discogenic lumbar back pain | Mesoblast Ltd. | NCT02412735 | Recruiting |
| Lumbar degenerative disc disease treatment with bone marrow autologous mesenchymal stem cells (MSV) | Red de Terapia Celular, Spain | NCT02440074 | Withdrawn |
| Human autograft mesenchymal stem cell mediated stabilization of the degnerative lumbar spine | Trinity Stem Cell Institution, Odessa, Florida, USA | NCT02529566 | Enrolling by invitation |
| Adipose cells for degenerative disc disease | Bioheart Inc. | NCT02097862 | Recruiting |
| Safety and efficacy with NOVOCART disc plus (ADCT) for the treatment of degenerative disc disease in lumbar spine (NDisc) | Tetec AG | NCT01640457 | Ongoing, not recruiting |
| A study comparing the safety and effectiveness of cartilage cells injected into the lumbar disc as compared to a placebo | ISTO Technologies Inc., USA | NCT01771471 | Ongoing, not recruiting |
Cellular repair
Which cells are appropriate for cellular repair of the disc?
It is a challenge to find an appropriate source of cells for disc repair (Kregar-Velikonja et al. 2014, Sakai and Andersson 2015). Human disc cells can only be harvested during surgical procedures. As no autologous cells from healthy discs are available, cells from other cartilages have been used for animal studies, while the use of notochord cells to stimulate resident cells is under investigation (Arkesteijn et al. 2015). Most researchers have, however, concentrated on differentiating stem cells or progenitor cells towards a nucleus pulposus-like cell type. Many studies have investigated the use of autologous mesenchymal stem cells (MSCs); allogenic MSCs are being tested in clinical trials (Table). A few studies have investigated differentiation of progenitor cells, or embryonic or induced pluripotent stem cells, towards the notochord- or adult nucleus pulposus cell phenotype. Success in differentiation is judged by expression of phenotypic nucleus pulposus markers (Risbud et al. 2015), which may not be specific (Thorpe et al. 2016), and through expression of matrix macromolecules such as collagen II and aggrecan, which are also expressed by other cartilages.
Currently, strategies tend to implant only 1 cell type into the disc—albeit that there are different cellular subpopulations even in the nucleus—and disc degeneration almost invariably involves more than 1 disc region (Figure 3A). Will stem cells implanted directly into the disc differentiate into the populations required to regenerate a stable nucleus, and repair the annulus and endplate? Strategies such as the use of notochord cells and chondrocyte-like cells generated from human stem cells may restore the dialogue between both cell types, based on the secretion of growth factors including TGF-β, CTGF, and SHH, and lead to the survival of nucleus cells and an increase in proteoglycan synthesis (Dahia et al. 2012). Would such differentiation strategies be sufficient, or would each region have to be directly targeted with appropriate cells?
Can implanted cells survive and function in the challenging environment found in degenerate discs?
As the dense matrix of the cartilaginous endplate and matrix of the normal disc acts as a permeability barrier between the disc cells and circulating macromolecules, the activity of the disc cells is governed to a large extent by their extracellular physical environment, and by signals from contacts with the extracellular matrix.
Nutrient levels limit the number of viable cells that can be implanted into the disc
Extracellular nutrient concentrations are of particular importance in the avascular disc (Figure 4A) (Grunhagen et al. 2011), which obtains its energy by aerobic glycolysis. Nutrient levels fall with distance from the blood supply and must remain above critical levels (0.2 mM glucose, pH 6.7) for cells to remain viable. Although much interest has been expressed in the hypoxic environment of the disc and the role of HIF-1 and HIF-2 (Risbud et al. 2010), nucleus cells can survive without oxygen; even so, they consume it, and matrix synthesis is affected by oxygen concentrations. As in other avascular cartilages (Stockwell 1971), viable cell density varies inversely with disc height, being only 1–5 million cells/mL in healthy human lumbar discs but over 50 million cells/mL in mouse discs (Figure 4B).
Figure 4.
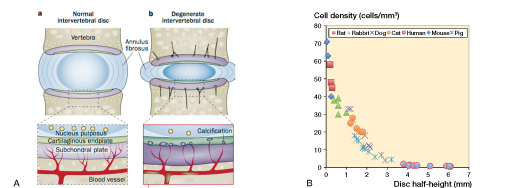
A. Schematic illustration showing nutrient pathways in a normal disc (a) and changes seen in disc degeneration (b). Most of the disc is supplied with nutrients by diffusion from capillaries arising in the vertebral body, which penetrate the subchondral plate and terminate at the junction with the cartilage endplate. Nutrients diffuse from these capillaries, through the cartilage endplate and disc matrix to the cells, which, in the center of a human disc, may be up to 8 mm from the nearest capillary. Nutrient supply is adversely affected in disc degeneration; disc degeneration is associated with atherosclerosis of the lumbar arteries and calcification of the cartilaginous endplate. Loss of nutrient supply leads to a fall in the number of active and viable cells that can be supported in the disc. (Reproduced from Huang et al. (2014) with permission). B. The inverse relationship between disc cell density across the nucleus pulposus and disc height. Cell density was measured in histological sections of discs taken from mice, rats, rabbits, cats, dogs, pigs, and humans. Here it has been plotted against disc half-height (adapted from Holm and Nachemson 1983).
The supply of nutrients thus limits the number of viable cells that can be implanted into even a healthy disc. In degenerate discs, calcification of the endplate further restricts nutrient supply and the number of viable cells (Figure 4A). Cells implanted into a degenerate disc may therefore have limited access to nutrients, compromising their activity and survival.
Signals from the matrix are disturbed in degenerate discs
Disc cells are sensitive to the level of extracellular osmolarity, which is regulated by aggrecan concentrations. Loss of aggrecan and hence osmolarity in degenerate discs both reduces rates of matrix production (Takeno et al. 2007) and initiates inflammatory changes (van Dijk et al. 2015). In addition, cells in degenerate discs produce more active proteases (Roberts et al. 2000, Pockert et al. 2009), which will tend to work against the ability of implanted cells to produce new matrix.
The inflammatory environment of degenerate discs can have an adverse effect on implanted cells
Inflammation is almost invariably encountered in degenerate discs (Risbud and Shapiro 2014). Inflammatory cytokines upregulate matrix degradation, thus slowing the rates of matrix accumulation and hindering attempts at repair; they can also induce pain. Moreover, these cytokines lead to further nutritional stresses, increasing rates of glycolysis, and thus further reducing glucose levels and pH levels—thereby compromising the activity and viability of implanted cells (Wuertz et al. 2009). Inflammation therefore appears to provide an unfavorable environment for implanted cells.
Can scaffolds drive cells towards repair?
The highly hydrated networks of hydrogels make them particularly suitable as a cell support for nucleus regeneration. While synthetic scaffolds with mechanical properties matching those of the nucleus are of interest, natural biopolymers have advantages in mimicking the native extracellular environment regarding mechanical, permeability, and biochemical properties—and in providing a bioresorbable temporary 3-dimensional microenvironment. Some, such as injectable alginate (Zeng et al. 2015) and hyaluronan hydrogels (Peroglio et al. 2013), may optimize stem cell differentiation and synthesis of an appropriate extracellular matrix. However, there are still no hydrogels that are able to fulfill needs regarding both cell biocompatibility and load-bearing capacity, and yet can also act as a reservoir of bioactive molecules.
Results from animal models may be misleading
Numerous in vivo studies have examined the process of cellular repair in animals ranging from mice to larger animals such as pigs and goats (Sakai and Andersson 2015), with apparently favorable outcomes. However, can such promising results be expected in humans? The discs of these animals, even those of cattle, are considerably smaller than human lumbar discs (Figure 5). The animal discs can consequently support a much greater cell density than human discs (Figure 4B). Moreover, the animals used are generally young or even immature, with degeneration produced by an acute intervention that may not produce inflammatory changes similar to those seen in humans, and may leave the nutrient supply unimpaired. Here, implanted cells appear to be able to survive and produce repair tissue relatively rapidly (in weeks or months). By contrast, the half-life of aggrecan in a degenerate human disc is around 4 years, and that of collagen and elastin is more than 50 years (Sivan et al. 2014b). Hence, results from animal models must be viewed with caution (Alini et al. 2008).
Figure 5.
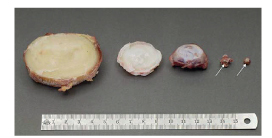
Relative sizes of intervertebral discs from different species. From left to right: human lumbar L4–L5 disc; bovine tail C1–C2 disc; sheep thoracic T11–T12 disc; rat lumbar and tail discs (with arrows showing the intervertebral disc location). (Reproduced from Alini et al. 2008 with permission).
Which patients would benefit from disc repair?
The important question of which patients would be suitable for cellular therapies has seldom been addressed (Kandel et al. 2008, Tibiletti et al. 2014, Benneker et al. 2014, Sakai and Andersson 2015). Patients come to see a clinician because they have back pain, not because they are worried about disc degeneration. Indeed, many people with even severe disc degeneration are asymptomatic and are unaware of having any spinal problems (Brinjikji et al. 2015). Thus, should pain rather than disc degeneration be the clinical target?
Currently, there is no reliable means of diagnosing whether a disc is the source of pain or not; discography has been discredited and may indeed cause harm (Carragee et al. 2009), and there are no validated MRI indications. In most cases, it is not known whether low back pain even arises from the disc; other structures such as the facets may also be involved, so regenerating the disc alone may not be effective. Moreover neuropathic pain, central nervous system changes, and disorders of muscular control are evident in many back pain patients (Freynhagen and Baron 2009, Yu et al. 2014, Schabrun et al. 2015), so even complete regeneration of the disc may not cure the pain.
Summary
Because of the complex nature of degenerative changes, biological repair of the disc invokes challenges in many areas. An integrated approach that involves not only the choice of appropriate cells and scaffolds for the different regions of the disc (including the endplate), but also targets inflammation and nutrient supply, might be necessary for successful and stable repair—and restoration of function. Although small clinical studies using single cell populations have been published showing apparent success (Meisel et al. 2006, Yoshikawa et al. 2010, Orozco et al. 2011, Mochida et al. 2015), information on outcomes is still awaited from randomized clinical trials (Table), which are currently in progress.
Conclusions
Over the past decade, the growing interest in the development of cell therapies has led to real progress with not only some promising results in this field in animal studies, but also in furthering our understanding of the biology of the intervertebral disc in general. However, a number of biological challenges must be overcome before these cellular therapies can be put into routine clinical use in humans.
One challenge is to improve characterization of the phenotype of the various disc cell populations, and then to determine how they interact under normal conditions and also in the nutrient-poor and inflammatory environment of degenerate discs—and importantly, to characterize the matrix macromolecules that they produce at the protein level. Without this information, it would be difficult to develop rational strategies for differentiation of stem or progenitor cells into cell phenotypes that can survive implantation and produce a stable and functional matrix.
Another challenge is to develop strategies for coping with the long repair process (years) in large human discs. This might necessitate designing scaffolds that, as well as supporting cells, would be able to restore load-bearing function to the degenerate disc and that can be maintained safely in the tissue until an appropriate matrix is synthesized by the low number of viable cells that are able to survive in human lumbar discs.
Yet another challenge, as in other regenerative cell-based therapies, is to reduce costs. Currently, the high cost of autologous donor cell preparations, and regulatory barriers, prevent routine clinical application for disorders such as disc degeneration.
Probably the most difficult challenge is to improve diagnosis in order to determine which patients would benefit most from disc regeneration, remembering that patients seek medical help for pain, not for disc degeneration. Even though current strategies using anti-TNF antibodies to treat pain have not always met with success (Cohen et al. 2009, Freeman et al. 2013), patients might still be better served by developing cellular therapies that are aimed at damping down inflammation and pain (Pettine et al. 2015, Willems et al. 2015), rather than through therapies aimed at biological regeneration of the disc.
References
- Adams M A. Biomechanics of back pain. Acupunct Med 2004; 22(4): 178–88. [DOI] [PubMed] [Google Scholar]
- Alini M, Roughley P J, Antoniou J, Stoll T, Aebi M.. A biological approach to treating disc degeneration: not for today, but maybe for tomorrow. Eur Spine J 2002; 11Suppl2: S215–S220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alini M, Eisenstein S M, Ito K, Little C, Kettler A A, Masuda K, Melrose J, Ralphs J, Stokes I, Wilke H J.. Are animal models useful for studying human disc disorders/degeneration? Eur Spine J 2008; 17(1): 2–19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arkesteijn I T, Smolders L A, Spillekom S, Riemers F M, Potier E, Meij B P, Ito K, Tryfonidou M A.. Effect of coculturing canine notochordal, nucleus pulposus and mesenchymal stromal cells for intervertebral disc regeneration. Arthritis Res Ther 2015; 17: 60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Battié M C, Videman T, Kaprio J, Gibbons L E, Gill K, Manninen H, Saarela J, Peltonen L.. The Twin Spine Study: Contributions to a changing view of disc degeneration. Spine J 2009; 9(1): 47–59. [DOI] [PubMed] [Google Scholar]
- Bendtsen M, Bunger C E, Zou X, Foldager C, Jorgensen H. S. Autologous stem cell therapy maintains vertebral blood flow and contrast diffusion through the endplate in experimental intervertebral disc degeneration. Spine (Phila Pa 1976) 2011; 36(6): E373–E379. [DOI] [PubMed] [Google Scholar]
- Benneker L M, Andersson G, Iatridis J C, Sakai D, Hartl R, Ito K, Grad S.. Cell therapy for intervertebral disc repair: advancing cell therapy from bench to clinics. Eur Cell Mater 2014; 27: 5–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boos N, Weissbach S, Rohrbach H, Weiler C, Spratt K F, Nerlich A G.. Classification of age-related changes in lumbar intervertebral discs: 2002 Volvo Award in basic science. Spine 2002; 27(23): 2631–44. [DOI] [PubMed] [Google Scholar]
- Brinjikji W, Luetmer P H, Comstock B, Bresnahan B W, Chen L E, Deyo R A, Halabi S, Turner J A, Avins A L, James K, Wald J T, Kallmes D F, Jarvik J G.. Systematic literature review of imaging features of spinal degeneration in asymptomatic populations. AJNR Am J Neuroradiol 2015; 36(4): 811–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carragee E J, Don A S, Hurwitz E L, Cuellar J M, Carrino J A, Herzog R.. 2009 ISSLS Prize Winner: Does discography cause accelerated progression of degeneration changes in the lumbar disc: a ten-year matched cohort study. Spine (Phila Pa 1976) 2009; 34(21): 2338–45. [DOI] [PubMed] [Google Scholar]
- Cohen S P, Bogduk N, Dragovich A, Buckenmaier C C III, Griffith S, Kurihara C, Raymond J, Richter P J, Williams N, Yaksh T L.. Randomized, double-blind, placebo-controlled, dose-response, and preclinical safety study of transforaminal epidural etanercept for the treatment of sciatica. Anesthesiology 2009; 110(5): 1116–26. [DOI] [PubMed] [Google Scholar]
- Cortes D H, Jacobs N T, DeLucca J F, Elliott D M.. Elastic, permeability and swelling properties of human intervertebral disc tissues: A benchmark for tissue engineering. J Biomech 2014; 47(9): 2088–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahia C L, Mahoney E, Wylie C.. Shh signaling from the nucleus pulposus is required for the postnatal growth and differentiation of the mouse intervertebral disc. PLoS One 2012; 7(4): e35944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eyre D R, Caterson B, Benya P, Heinegard D, Oegema T R, Pearce R H, Pope M H, Urban J P G.. The intervertebral disc. In: New Perspectives on Low Back Pain. (Eds. Gordon S, Frymoyer J). Am Inst Orthop Surg., Philadelphia, 1991; 147–209. [Google Scholar]
- Feng H, Danfelter M, Stromqvist B, Heinegard D.. Extracellular matrix in disc degeneration. J Bone Joint Surg Am 2006; 88Suppl2: 25–9. [DOI] [PubMed] [Google Scholar]
- Freeman B J, Ludbrook G L, Hall S, Cousins M, Mitchell B, Jaros M, Wyand M, Gorman J R.. Randomized, double-blind, placebo-controlled, trial of transforaminal epidural etanercept for the treatment of symptomatic lumbar disc herniation. Spine (Phila Pa 1976) 2013; 38(23): 1986–94. [DOI] [PubMed] [Google Scholar]
- Freynhagen R, Baron R.. The evaluation of neuropathic components in low back pain. Curr Pain Headache Rep 2009; 13(3): 185–190. [DOI] [PubMed] [Google Scholar]
- Galbusera F, van R M, Ito K, Huyghe J M, Brayda-Bruno M, Wilke H J.. Ageing and degenerative changes of the intervertebral disc and their impact on spinal flexibility. Eur Spine J 2014; 23Suppl3: S324–S332. [DOI] [PubMed] [Google Scholar]
- Gruber H E, Riley F E, Hoelscher G L, Ingram J A, Bullock L, Hanley E N Jr.. Human annulus progenitor cells: Analyses of this viable endogenous cell population. J Orthop Res 2016; 34(8): 1351–60. [DOI] [PubMed] [Google Scholar]
- Grunhagen T, Shirazi-adl A, Fairbank J C, Urban J P.. Intervertebral disk nutrition: a review of factors influencing concentrations of nutrients and metabolites. Orthop Clin North Am 2011; 42(4): 465–77, vii. [DOI] [PubMed] [Google Scholar]
- Henriksson H, Thornemo M, Karlsson C, Hagg O, Junevik K, Lindahl A, Brisby H.. Identification of cell proliferation zones, progenitor cells and a potential stem cell niche in the intervertebral disc region: a study in four species. Spine (Phila Pa 1976) 2009; 34(21): 2278–87. [DOI] [PubMed] [Google Scholar]
- Hunziker E B, Lippuner K, Keel M J, Shintani N.. An educational review of cartilage repair: precepts & practice–myths & misconceptions–progress & prospects. Osteoarthritis Cartilage 2015; 23(3): 334–50. [DOI] [PubMed] [Google Scholar]
- Kandel R, Roberts S, Urban J P G.. Tissue engineering and the intervertebral disc: the challenges. Eur Spine J 2008; 17Suppl4: S480–S491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kletsas D. Senescent cells in the intervertebral disc: numbers and mechanisms. Spine J 2009; 9(8): 677–8. [DOI] [PubMed] [Google Scholar]
- Kregar-Velikonja N, Urban J, Frohlich M, Neidlinger-Wilke C, Kletsas D, Potocar U, Turner S, Roberts S.. Cell sources for nucleus pulposus regeneration. Eur Spine J 2014; 23Suppl3: S364–S374. [DOI] [PubMed] [Google Scholar]
- Lyons G, Eisenstein S M, Sweet M B.. Biochemical changes in intervertebral disc degeneration. Biochim Biophys Acta 1981; 673(4): 443–53. [DOI] [PubMed] [Google Scholar]
- Meisel H J, Ganey T, Hutton W C, Libera J, Minkus Y, Alasevic O.. Clinical experience in cell-based therapeutics: intervention and outcome. Eur Spine J 2006; 15Suppl3: S397–S405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mochida J, Sakai D, Nakamura Y, Watanabe T, Yamamoto Y, Kato S.. Intervertebral disc repair with activated nucleus pulposus cell transplantation: a three-year, prospective clinical study of its safety. Eur Cell Mater 2015; 29: 202–12. [DOI] [PubMed] [Google Scholar]
- Molinos M, Almeida C R, Goncalves R M, Barbosa M A.. Improvement of bovine nucleus pulposus cells Isolation leads to identification of three phenotypically distinct cell subpopulations. Tissue Eng Part A 2015; 21(15–16): 2216–27. [DOI] [PubMed] [Google Scholar]
- Muriuki M G, Havey R M, Voronov L I, Carandang G, Zindrick M R, Lorenz M A, Lomasney L, Patwardhan A G.. Effects of motion segment level, Pfirrmann intervertebral disc degeneration grade and gender on lumbar spine kinematics. J Orthop Res 2016: 34(8): 1389–98. [DOI] [PubMed] [Google Scholar]
- Nosikova Y S, Santerre J P, Grynpas M, Gibson G, Kandel R A.. Characterization of the annulus fibrosus-vertebral body interface: identification of new structural features. J Anat 2012; 221(6): 577–89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orozco L, Soler R, Morera C, Alberca M, Sanchez A, Garcia-Sancho J.. Intervertebral disc repair by autologous mesenchymal bone marrow cells: a pilot study. Transplantation 2011; 92(7): 822–828. [DOI] [PubMed] [Google Scholar]
- Pattappa G, Li Z, Peroglio M, Wismer N, Alini M, Grad S.. Diversity of intervertebral disc cells: phenotype and function. J Anat 2012; 221(6): 480–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peroglio M, Eglin D, Benneker L M, Alini M, Grad S.. Thermoreversible hyaluronan-based hydrogel supports in vitro and ex vivo disc-like differentiation of human mesenchymal stem cells. Spine J 2013; 13(11): 1627–39. [DOI] [PubMed] [Google Scholar]
- Pettine K A, Murphy M B, Suzuki R K, Sand T T.. Percutaneous injection of autologous bone marrow concentrate cells significantly reduces lumbar discogenic pain through 12 months. Stem Cells 2015; 33(1): 146–56. [DOI] [PubMed] [Google Scholar]
- Pezowicz C A, Robertson P A, Broom N D.. The structural basis of interlamellar cohesion in the intervertebral disc wall. J Anat 2006; 208(3): 317–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfirrmann C W, Metzdorf A, Zanetti M, Hodler J, Boos N.. Magnetic resonance classification of lumbar intervertebral disc degeneration. Spine 2001; 26(17): 1873–8. [DOI] [PubMed] [Google Scholar]
- Pockert A J, Richardson S M, Le Maitre C L, Lyon M, Deakin J A, Buttle D J, Freemont A J, Hoyland J A.. Modified expression of the ADAMTS enzymes and tissue inhibitor of metalloproteinases 3 during human intervertebral disc degeneration. Arthritis Rheum 2009; 60(2): 482–91. [DOI] [PubMed] [Google Scholar]
- Risbud M V, Shapiro I M.. Role of cytokines in intervertebral disc degeneration: pain and disc content. Nat Rev Rheumatol 2014; 10(1): 44–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Risbud M V, Schipani E, Shapiro I M.. Hypoxic regulation of nucleus pulposus cell survival: from niche to notch. Am J Pathol 2010; 176(4): 1577–1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Risbud M V, Schoepflin Z R, Mwale F, Kandel R A, Grad S, Iatridis J C, Sakai D, Hoyland J A.. Defining the phenotype of young healthy nucleus pulposus cells: Recommendations of the Spine Research Interest Group at the 2014 Annual ORS Meeting. J Orthop Res 2015; 33(3): 283–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts S, Caterson B, Menage J, Evans E H, Jaffray D C, Eisenstein S M.. Matrix metalloproteinases and aggrecanase: their role in disorders of the human intervertebral disc. Spine 2000; 25(23): 3005–13. [DOI] [PubMed] [Google Scholar]
- Roberts S, Evans H, Trivedi J, Menage J.. Histology and pathology of the human intervertebral disc. J Bone Joint Surg Am 2006; 88Suppl2: 10–4. [DOI] [PubMed] [Google Scholar]
- Sakai D, Andersson G B.. Stem cell therapy for intervertebral disc regeneration: obstacles and solutions. Nat Rev Rheumatol 2015; 11(4): 243–256. [DOI] [PubMed] [Google Scholar]
- Sakai D, Grad S.. Advancing the cellular and molecular therapy for intervertebral disc disease. Adv Drug Deliv Rev 2015; 84: 159–171. [DOI] [PubMed] [Google Scholar]
- Sakai D, Nakamura Y, Nakai T, Mishima T, Kato S, Grad S, Alini M, Risbud M V, Chan D, Cheah K S, Yamamura K, Masuda K, Okano H, Ando K, Mochida J.. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat Commun 2012; 3: 1264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schabrun S M, Elgueta-Cancino E L, Hodges P W.. Smudging of the motor cortex is related to the severity of low back pain. Spine (Phila Pa 1976) 2015. [Epub ahead of print]. [DOI] [PubMed] [Google Scholar]
- Sivan S, Merkher Y, Wachtel E, Ehrlich S, Maroudas A.. Correlation of swelling pressure and intrafibrillar water in young and aged human intervertebral discs. J Orthop Res 2006; 24(6): 1292–8. [DOI] [PubMed] [Google Scholar]
- Sivan S S, Hayes A J, Wachtel E, Caterson B, Merkher Y, Maroudas A, Brown S, Roberts S.. Biochemical composition and turnover of the extracellular matrix of the normal and degenerate intervertebral disc. Eur Spine J 2014a; 23Suppl3: S344–S353. [DOI] [PubMed] [Google Scholar]
- Sivan S S, Wachtel E, Roughley P.. Structure, function, aging and turnover of aggrecan in the intervertebral disc. Biochim Biophys Acta 2014b; 1840(10): 3181–9. [DOI] [PubMed] [Google Scholar]
- Stockwell R. The inter-relationship of cell density and cartilage thickness in mammalian articular cartilage. J Anat 1971; 109: 411–22. [PMC free article] [PubMed] [Google Scholar]
- Takeda T. Three-dimensional observations of collagen framework of human lumbar discs. J Japan Orthop Assoc 1975; 49: 45–57. [DOI] [PubMed] [Google Scholar]
- Takeno K, Kobayashi S, Negoro K, Uchida K, Miyazaki T, Yayama T, Shimada S, Baba H.. Physical limitations to tissue engineering of intervertebral disc cells: effect of extracellular osmotic change on glycosaminoglycan production and cell metabolism. Laboratory investigation. J Neurosurg Spine 2007; 7(6): 637–44. [DOI] [PubMed] [Google Scholar]
- Thorpe A A, Binch A L, Creemers L B, Sammon C, Le Maitre C L.. Nucleus pulposus phenotypic markers to determine stem cell differentiation: fact or fiction? Oncotarget 2016; 7(3): 2189–200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tibiletti M, Kregar V N, Urban J P, Fairbank J C.. Disc cell therapies: critical issues. Eur Spine J 2014; 23Suppl3: S375–S384. [DOI] [PubMed] [Google Scholar]
- Urban J P, Roberts S.. Development and degeneration of the intervertebral discs. Mol Med Today 1995; 1(7): 329–35. [DOI] [PubMed] [Google Scholar]
- van Dijk B, Potier E, van D M, Langelaan M, Papen-Botterhuis N, Ito K.. Reduced tonicity stimulates an inflammatory response in nucleus pulposus tissue that can be limited by a COX-2-specific inhibitor. J Orthop Res 2015; 33(11): 1724–31. [DOI] [PubMed] [Google Scholar]
- Von Forell G A, Stephens T K, Samartzis D, Bowden A E.. Low Back Pain: A Biomechanical Rationale Based on "Patterns" of Disc Degeneration. Spine (Phila Pa 1976) 2015; 40(15): 1165–1172. [DOI] [PubMed] [Google Scholar]
- Willems N, Yang H Y, Langelaan M L, Tellegen A R, Grinwis G C, Kranenburg H J, Riemers F M, Plomp S G, Craenmehr E G, Dhert W J, Papen-Botterhuis N E, Meij B P, Creemers L B, Tryfonidou M A.. Biocompatibility and intradiscal application of a thermoreversible celecoxib-loaded poly-N-isopropylacrylamide MgFe-layered double hydroxide hydrogel in a canine model. Arthritis Res Ther 2015; 17: 214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshikawa T, Ueda Y, Miyazaki K, Koizumi M, Takakura Y.. Disc regeneration therapy using marrow mesenchymal cell transplantation: a report of two case studies. Spine (Phila Pa 1976) 2010; 35(11): E475–E480. [DOI] [PubMed] [Google Scholar]
- Yu R, Gollub R L, Spaeth R, Napadow V, Wasan A, Kong J.. Disrupted functional connectivity of the periaqueductal gray in chronic low back pain. Neuroimage Clin 2014; 6: 100–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J, Schollum M L, Wade K R, Broom N D, Urban J P.. ISSLS Prize Winner: A Detailed examination of the elastic network leads to a new understanding of annulus fibrosus organization. Spine (Phila Pa 1976) 2015; 40(15): 1149–57. [DOI] [PubMed] [Google Scholar]
- Zeng Y, Chen C, Liu W, Fu Q, Han Z, Li Y, Feng S, Li X, Qi C, Wu J, Wang D, Corbett C, Chan B P, Ruan D, Du Y.. Injectable microcryogels reinforced alginate encapsulation of mesenchymal stromal cells for leak-proof delivery and alleviation of canine disc degeneration. Biomaterials 2015; 59: 53–65. [DOI] [PubMed] [Google Scholar]