

Correspondence
The production of the iron-regulatory hormone hepcidin is physiologically suppressed during increased erythropoietic activity. Hepcidin suppression increases iron availability to meet the additional iron demand of expanded hemoglobin synthesis. Under the influence of erythropoietin (EPO), the hormone erythroferrone (ERFE) is secreted by erythroid precursors in the bone marrow and spleen, and suppresses hepcidin synthesis to facilitate the recovery from anemia1,2. However, the mechanism by which ERFE suppresses hepcidin is unknown.
In contrast with forms of anemia in which hepcidin is suppressed, patients with iron-refractory iron deficiency anemia (IRIDA)3, a disease caused by mutations in transmembrane serine protease 6 (TMPRSS6) which result in pathological activation of the BMP/Smad signaling, exhibit increased hepcidin production despite a severe anemia and elevated EPO levels. Previous studies have shown that hepcidin production was not affected by EPO in Tmprss6 mutant mice4. More recently, Nai and colleagues suggested that matriptase-2 may act downstream of EPO to dampen the signaling through the BMP-Smad regulatory pathway and allow ERFE to repress hepcidin production5. We therefore examined the crosstalk between ERFE and matriptase-2 in mice.
In agreement with Nai et al, we observed that Erfe mRNA expression was highly increased in the bone marrow and spleen of Tmprss6−/− mice (Supplementary Figure 1). While wild-type mice (WT) mice have undetectable plasma ERFE, plasma ERFE concentration was elevated in Tmprss6−/− to levels comparable to those of WT animals 24 hours after phlebotomy (~3 ng/ml) but was lower than ERFE levels in thalassemic mice (~10 ng/ml) (Figure 1A). To assess the contribution of ERFE to the phenotype of Tmprss6−/− mice, we generated Tmprss6−/− mice with disrupted Erfe (Erfe−/− Tmprss6−/−; Erfe+/− Tmprss6−/− and Erfe+/+ Tmprss6−/−). Ablation of Erfe in Tmprss6−/− mice did not result in any change in hematological parameters compared to Tmprss6−/− animals at 6 weeks of age (Table 1). Mice deficient for Tmprss6 had higher RBC and lower hemoglobin, hematocrit, and MCV compared to WT, regardless of the Erfe genotype. Liver hepcidin mRNA (Figure 1B) and serum hepcidin concentration (Figure 1C) were higher in Tmprss6−/− mice compared to WT mice and no difference was observed after manipulation of Erfe. Consistent with chronically increased hepcidin production, liver iron content (Figure 1D) was decreased in Tmprss6−/− mice compared to WT mice, but this was not influenced by the ablation of Erfe. No statistically significant difference in spleen iron content was observed between the genotypes (Figure 1E). Erfe-deficient Tmprss6−/− mice and Tmprss6−/− mice showed similarly increased spleen weight compared to WT mice (Supplemental Figure 2A). However, we observed a decrease in body weight in double mutant animals at 6 weeks compared to Tmprss6−/− mice (Supplemental Figure 2B) suggesting that ERFE may be important for normal development of Tmprss6−/− mice. As a consequence of smaller body size, spleen index was higher in Erfe−/−, Tmprss6−/− mice compared to Tmprss6−/− mice (Supplemental Figure 2C). Together, these results indicate that the disruption of Erfe has at most a very minor effect on the phenotype of Tmprss6 deficiency in mice.
Figure 1. ERFE and Matriptase 2 regulate hepcidin independently.
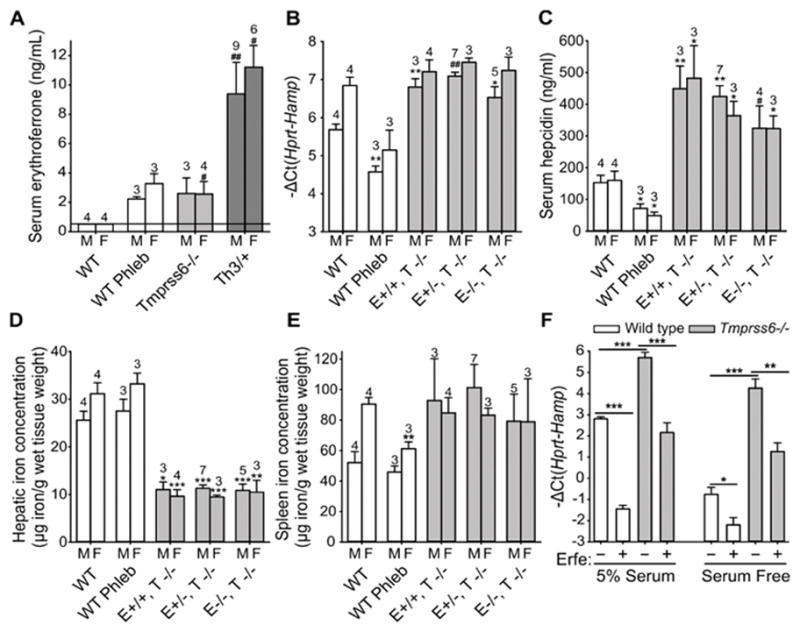
(A) Serum ERFE concentration was elevated in Tmprss6−/− mice similarly to WT (wild-type) mice 24 hours after phlebotomy, but was lower than in thalassemic mice (Th3/+). (B,C) Neither liver hepcidin mRNA expression nor serum hepcidin were significantly different in Erfe−/− Tmprss6−/− (E−/−,T−/−) mice compared to Erfe+/+ Tmprss6−/− (E+/+,T−/−) or Erfe+/− Tmprss6−/− (E+/−,T−/−) mice. (D) Liver iron content was similarly lower in Tmprss6−/− mice compared to WT mice regardless of the Erfe genotype whereas spleen iron content (E) was comparable to those of WT mice. (A–E) The number of mice studied is shown for each group. Data shown are means ± SEM. Statistical analysis was performed compared to WT controls of the same gender, using the two-tailed Student t-test (***P<.001, **P<.01, *P<.05) or Mann-Whitney test (##P<.01, #P<.05). M=male, F=female. (F) Hepcidin mRNA expression was strongly reduced in primary hepatocytes from WT and Tmprss6−/− mice 16 hours after treatment with conditioned medium containing ERFE (final concentration 500 ng/ml of recombinant ERFE), both in 5% serum or serum-free conditions. Data shown are means ± SEM of three independent experiments. Statistical analysis was performed using two-tailed Student t-test (***P<.001, **P<.01, *P<.05).
Table 1. Hematological parameters are unchanged by ERFE ablation in Tmprss6−/− mice.
RBC was increased while hemoglobin, hematocrit and MCV were decreased in Tmprss6−/− mice compared to WT mice at 6 weeks of age. Ablation of Erfe in Tmprss6−/− mice did not result in any change in hematological parameters compared to Tmprss6−/− animals at 6 weeks of age (two-tailed Student t-test comparison of Erfe−/− Tmprss6−/− and Erfe+/− Tmprss6−/− to Erfe+/+ Tmprss6−/−). None of the CBC parameters were statistically different by gender or by genotype when comparing Erfe−/− Tmprss6−/− and Erfe+/− Tmprss6−/− to Erfe+/+ Tmprss6−/− by two-way ANOVA. Data shown are means ± SEM and were compared for each group to control WT mice by two-tailed Student t-test (n=6 to 16 per group, gender combined).
| N | RBC (M/μL) | HB (g/dL) | HCT (%) | MCV (fL) | |
|---|---|---|---|---|---|
| Males | |||||
| WT | 4 | 9.4 ± 0.2 | 16.3 ± 0.5 | 45.7 ± 1.2 | 48.6 ± 0.2 |
| WT phlebotomized | 3 | 8.0** ± 0.1 | 13.9*± 0.4 | 38.7**± 0.4 | 48.3 ± 0.7 |
| Erfe +/+ Tmprss6 −/− | 5 | 10.3 ± 0.4 | 8.4*** ± 0.3 | 34.9***± 1.2 | 34.0** ± 0.4 |
| Erfe +/− Tmprss6 −/− | 12 | 10.7* ± 0.3 | 8.9*** ± 0.2 | 35.7*** ± 0.7 | 33.5** ± 0.4 |
| Erfe −/− Tmprss6 −/− | 5 | 10.8* ± 0.5 | 9.0*** ± 0.4 | 36.6* ± 1.3 | 34.0*** ± 0.2 |
| Females | |||||
| WT | 4 | 9.2 ± 0.1 | 16.3 ± 0.4 | 44.3 ± 0.6 | 48.1 ± 0.2 |
| WT phlebotomized | 3 | 6.8*** ± 0.3 | 11.8*** ± 0.4 | 33.3*** ± 1.1 | 48.6 ± 0.2 |
| Erfe +/+ Tmprss6 −/− | 5 | 11.4** ± 0.4 | 9.4*** ± 0.3 | 38.6** ± 1.1 | 33.9*** ± 0.2 |
| Erfe +/− Tmprss6 −/− | 4 | 10.2* ± 0.4 | 8.7*** ± 0.5 | 34.8*** ± 1.1 | 34.0*** ± 0.4 |
| Erfe −/− Tmprss6 −/− | 6 | 11.3*** ± 0.3 | 9.7** ± 0.2 | 37.6*** ± 0.9 | 33.3*** ± 0.1 |
P < .001,
P < .01,
P < .05.
To determine whether ERFE requires TMPRSS6 to regulate hepcidin production, we treated freshly isolated hepatocytes from WT or Tmprss6−/− mice with conditioned medium from cells expressing recombinant ERFE or from control cells. Consistent with stimulated BMP/Smad signaling, baseline hepcidin expression was higher in Tmprss6−/− hepatocytes compared to WT hepatocytes in serum-containing and serum-free conditions (Figure 1F). Hepcidin mRNA expression was reduced more than 12-fold in both WT and Tmprss6−/− hepatocytes after treatment with ERFE in serum-containing media. Similarly, treatment of hepatocytes with ERFE in serum-free media (i.e. in the absence of any soluble matriptase-2) resulted in a 2.5 and 5-fold decrease in hepcidin expression in WT and Tmprss6−/− hepatocytes, respectively. These results demonstrate in vitro that ERFE represses hepcidin independently of TMPRSS6. Interestingly, Id1 mRNA expression (Supplemental Figure 3A) was also reduced in primary mouse hepatocytes from WT and Tmprss6−/− mice after treatment with ERFE in serum-containing and serum-free conditions. Similarly, Atoh8 and Smad7 mRNA expression (Supplemental Figure 3B, C) was decreased in ERFE-treated WT and Tmprss6−/− hepatocytes in serum-containing conditions. In serum-free conditions, the decrease in Atoh8 and Smad7 mRNA expression was observed only in ERFE-treated Tmprss6−/− hepatocytes but not in WT hepatocytes. This suggests a potential crosstalk between BMP and ERFE signaling that is independent of TMPRSS6.
The reason for the impairment of erythropoietic suppression of hepcidin in patients and mice with IRIDA may lie in the intense stimulation of hepcidin by the BMP pathway in vivo, opposing the effect of ERFE. In accordance with the findings by Nai and colleagues, we previously demonstrated that hepcidin suppression after phlebotomy is blunted in iron-overloaded WT mice and Tfr2−/− mice where the BMP pathway is strongly stimulated by iron2. These observations indicate that physiologic erythroid regulators (including ERFE) act additively to BMP-signaling and have a smaller relative effect on hepcidin production when the BMP/Smad signaling is highly activated.
The relatively weak effect of erythroid suppressors of hepcidin in IRIDA must be reconciled with the dominant effect of erythroid suppressors of hepcidin despite the iron-overload stimulated BMP signaling in patients with β-thalassemia and other disorders with ineffective erythropoiesis. Here, three kinds of differences may be relevant. First, the concentrations of ERFE (and perhaps other erythroid suppressors) may be higher in conditions with ineffective erythropoiesis than in IRIDA, likely because of the greater number of ERFE-secreting erythroblasts in ineffective erythropoiesis. Secondly, in anemias other than IRIDA, erythropoietin stimulation may increase the production of matriptase 2 through a posttranslational mechanism, potentiating the suppressive effect of erythroferrone on hepcidin. Thirdly, there are important species differences in the observed effect of erythroid suppressors of hepcidin in the mouse models of ineffective erythropoiesis vs. human patients. Young β-thalassemic mice exhibit suppressed hepcidin, but progressively increase hepcidin with age to levels comparable to those of adult WT mice as they become iron overloaded, despite elevated ERFE production6. In contrast, adult human patients with β-thalassemia continue to have decreased to undetectable levels of hepcidin despite severe iron overload. Human studies will be needed to quantitate the effects of erythroid suppressors of hepcidin production.
Supplementary Material
Acknowledgments
The authors thank Jodie Babitt (Massachusetts General Hospital) for providing the Tmprss6−/− mice and Xin Du, Justin Chapman, and Marc Nasoff of Silarus Therapeutics for providing the ERFE assay.
Sources of funding—This research was supported by the NIH grant R01 DK 065029 to T.G., the ASH scholar award and ANR-16-ACHN-0002-01 to L.K. and the NHLBI award F31HL129760 and NIH grant T32 GM007185 to S.A.
Disclosures—E.N and T.G. are shareholders and scientific advisors of Intrinsic LifeSciences, Merganser Biotech, and Silarus Therapeutics. T.G. is a consultant to Keryx Biopharmaceuticals.
Footnotes
AUTHOR CONTRIBUTIONS:
S.A. performed experiments, analyzed the data, and edited the manuscript, L.K. designed and performed the experiments, analyzed the data and wrote the paper, E.N. and T.G. supervised the project and wrote the paper.
REFERENCE LIST
- 1.Kautz L, Jung G, Nemeth E, Ganz T. Erythroferrone contributes to recovery from anemia of inflammation. Blood. 2014;124(16):2569–2574. doi: 10.1182/blood-2014-06-584607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kautz L, Jung G, Valore EV, Rivella S, Nemeth E, Ganz T. Identification of erythroferrone as an erythroid regulator of iron metabolism. NatGenet. 2014;46(7):678–684. doi: 10.1038/ng.2996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.De Falco L, Sanchez M, Silvestri L, et al. Iron refractory iron deficiency anemia. Haematologica. 2013;98(6):845–853. doi: 10.3324/haematol.2012.075515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Nicolas G, Deschemin JC, Ramsay AJ, et al. Is EPO therapy able to correct iron deficiency anaemia caused by matriptase-2 deficiency? Br J Haematol. 2011;152(4):498–500. doi: 10.1111/j.1365-2141.2010.08473.x. [DOI] [PubMed] [Google Scholar]
- 5.Nai A, Rubio A, Campanella A, et al. Limiting hepatic Bmp-Smad signaling by matriptase-2 is required for erythropoietin-mediated hepcidin suppression in mice. Blood. 2016;127(19):2327–2336. doi: 10.1182/blood-2015-11-681494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kautz L, Jung G, Du X, et al. Erythroferrone contributes to hepcidin suppression and iron overload in a mouse model of beta-thalassemia. Blood. 2015;126(17):2031–2037. doi: 10.1182/blood-2015-07-658419. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.