

Summary
Cementum was first demonstrated by microscopy, about 180 years ago. Since then the biology of cementum has been investigated by the most advanced techniques and equipment at that time in various fields of dental sciences. A great deal of data on cementum histology have been accumulated. These data have been obtained from not only human, but also non-human animals, in particular, rodents such as the mouse and rat. Although many dental histologists have reviewed histology of human cementum, some descriptions are questionable, probably due to incorrect comparison of human and rodent cementum. This review was designed to introduce current histology of human cementum, i.e. its structure, function, and development and to re-examine the most questionable and controversial conclusions made in previous reports.
Keywords: Acellular extrinsic fiber cementum, Cellular intrinsic fiber cementum, Cellular mixed stratified cementum, Extrinsic fibers, Intrinsic fibers, Human cementum
1. Introduction
Cementum, or root cementum, is a mineralized tissue covering the entire root surface. According to Denton [1], cementum was first demonstrated microscopically by Fraenkel and Raschkow (1835) and Retzius (1836), and has since become a part of general knowledge in dentistry. Cementum exists fundamentally in mammalian teeth, which fit into alveolar sockets of alveolar bone, and functions as a tooth-supporting device in concert with the periodontal principal fibers and alveolar bone. The cementum covering the enamel surface also exists in several non-human animals such as the horse, sheep, rabbit and guinea pig. This type of cementum is termed “coronal cementum” and distinct from root cementum. Cementum is often referred to as a bone-like tissue. Cementum, however, is avascular, does not undergo dynamic remodeling, and increases in thickness throughout life. On these points, cementum is markedly different from bone.
Knowledge of cementum histology has been accumulated with the advancement of investigating techniques and equipment. Most of the data, however, have been obtained from rodents such as the mouse and rat. Hence, previous reports on histology of human cementum have contained some questionable descriptions, probably due to incorrect comparison of human and rodent cementum. The purpose of this review is to introduce current histology of human cementum and to re-examine the most questionable and controversial conclusions made in previous reports.
2. Classification of cementum
Cementum has historically been classified into cellular and acellular cementum by inclusion or non-inclusion of cementocytes. Generally, acellular cementum is thin and covers the cervical root, whereas thick cellular cementum covers the apical root (Fig. 1) Cementum contains two types of fibers, i.e. extrinsic (Sharpey's) fibers which are embedded ends of the principal fibers and intrinsic fibers which are fibers of cementum proper. It is believed that the extrinsic fibers are secreted by fibroblasts and partly cementoblasts and that the intrinsic fibers are secreted by only cementoblasts. Jones [2] added these fibers to items of cementum classification, and Schroeder [3] established the current classification, which is now widely used in the dental field. In accordance with this classification, three major types of cementum are distinguishable [2], [3], [4], [5]. “Acellular extrinsic fiber cementum (AEFC)” contains densely packed extrinsic fibers and no cementocytes (Fig. 2). AEFC corresponds to classical acellular cementum. “Cellular intrinsic fiber cementum (CIFC)” contains intrinsic fibers and cementocytes. “Cellular mixed stratified cementum (CMSC)” corresponds to classical cellular cementum. Typical CMSC is partitioned by intensely hematoxylin-stainable lines or incremental lines. The individual partitioned cementum is CIFC, and occasionally AEFC (Fig. 3A–D). Namely, CMSC represents the whole of cellular cementum composed of stratified CIFC and AEFC. Cellular cementum with both intrinsic and extrinsic fibers is often found within CMSC (Fig. 3D). This type of cementum is not distinctively classified and is regarded as a sub-variety of CIFC in the current classification [5]. This review will deal with the three major types.
Figure 1.

Full view of a mandibular molar (left) and a maxillary incisor (right). Thin AEFC and thick CMSC cover cervical and apical roots, respectively. CMSC is much thicker in the molar than in the incisor. Hematoxylin-stained ground section. Bar 5 mm.
Figure 2.
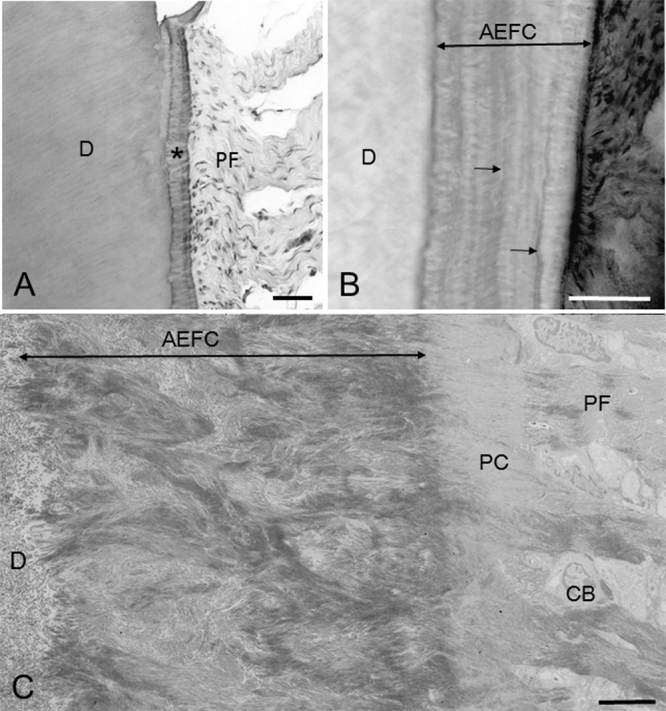
Light (A, B) and transmission electron (C) micrographs showing AEFC. (A) Principal fibers (PF) enter AEFC (*) as extrinsic fibers. D, dentin. Hematoxylin and eosin-stained paraffin section. Bar 50 μm. (B) Extrinsic fibers are observed as white lines in AEFC. Extrinsic fibers change their orientation at the intensely stainable incremental lines (arrows). D, dentin. Hematoxylin-stained ground section. Bar 50 μm. (C) Extrinsic fibers are branching and anastomosing. D, dentin; CB, cementoblasts; PC, precementum; PF, principal fibers. Bar 5 μm.
Figure 3.

(A–D) Micrographs showing structural variety of CMSC in hematoxylin-stained ground sections. CMSC is partitioned by many, intensely stainable incremental lines. RD, root dentin. Bars 100 μm (A–C), 50 μm (D). (A) CMSC consists of stratified CIFC. (B) AEFC (*) is present as the first formed cementum of CMSC. (C) AEFC (*) intervenes between CIFC. (D) Magnification of extrinsic fiber-rich (*), -poor (**), and -free CIFC (***). (E, F) Transmission electron micrographs showing extrinsic (EF) and intrinsic fibers (IF) in CIFC. Bars 5 μm. (E) Extrinsic fibers are branching and anastomosing. Intrinsic fibers fill the space between extrinsic fibers. (F) Intrinsic fibers encircle extrinsic fibers or meander among them in a tangential section parallel to cementum surface.
3. Structure of cementum
3.1. Acellular extrinsic fiber cementum (AEFC)
Generally, AEFC covers cervical root surfaces in both permanent and deciduous teeth. The covering range is different among types of teeth; 60–90% of the total root length in single-rooted teeth, and cervical half to one third in multi-rooted teeth (Fig. 1). Its thickness, which increases with age, ranges from 50 to 200 μm [3], [4].
AEFC contains collagen fibers and non-collagenous proteins as organic matrices, both of which are fully mineralized. All collagen fibers belong to extrinsic fibers connecting to the principal fibers in the periodontal ligament [2], [3], [4]. The extrinsic fibers are densely packed and arranged nearly perpendicularly to the root surface (Fig. 2A, B). The diameter of extrinsic fibers is roughly calculated at 3–6 μm [3]. They are not simple straight fiber bundles, and show branching and anastomosing (Fig. 2C). The structural correlation of the extrinsic fibers and dentin matrix fibers at the cemento-dentinal junction has been often disputed. A majority of previous studies revealed interdigitation of extrinsic fibers and dentin matrix fibers and thus suggested that cementum and dentin were firmly united by the fiber interdigitation and mineralization [4], [6], [7], [8], [9]. In contrast, some investigators found a fibril-poor, non-collagenous protein-rich layer at the cemento-dentinal junction and proposed that the proteins served as an adhesive of cementum and dentin [10], [11], [12].
Thick AEFC shows several incremental lines which are intensely stained with hematoxylin (Fig. 2B). The incremental lines are highly mineralized [13] and are thus regarded as resting lines, formed in resting phases during intermittent AEFC formation. In many cases, the extrinsic fiber orientation changes at the incremental lines. The change is considered to correlate with relative positional changes of tooth and alveolar bone during tooth eruption [3]. Unmineralized cementum is seen as thin precementum or cementoid on AEFC.
3.2. Cellular intrinsic fiber cementum (CIFC)
As mentioned above, CIFC is usually present as a component of CMSC [3], [4], [5] (Fig. 3A–C). The structure of CIFC will be described in detail in the following section.
3.3. Cellular mixed stratified cementum (CMSC)
CMSC is predominantly seen in the interradicular and apical regions of roots (Figure 1, Figure 3A–C). According to Schroeder [3], in middle-aged individuals, its maximum thickness ranges between 400 and 600 μm in incisors, around 500 μm in canines, between 300 and 1000 μm in premolars, and between 700 and 1500 μm in molars. In molars, CMSC often covers apical two thirds of the roots (Fig. 1). CMSC usually consists of stratified CIFC, with each CIFC being demarcated by intensely hematoxylin-stainable incremental lines (Fig. 3A–D). Occasionally, AEFC also appears as a component of CMSC (Fig. 3B, C). CIFC often contains extrinsic fibers as well as intrinsic fibers, and the density of extrinsic fibers varies among individual CIFC. Hence, various types of CIFC, i.e. extrinsic fiber-rich CIFC, extrinsic fiber-poor CIFC, and extrinsic fiber-free CIFC are subdivided on the basis of the density of extrinsic fibers (Fig. 3D). In extrinsic fiber-containing CIFC, the extrinsic fibers, like those in AEFC, show branching and anastomosing, and are encircled by the intrinsic fibers [3], [7], [14] (Fig. 3E, F). The diameter of extrinsic fibers is under 10 μm and thicker than those extrinsic fibers in AEFC. The extrinsic fibers of CIFC often contain an unmineralized central core, surrounded by a highly mineralized cortical part [3], [7]. It is presently unknown whether all extrinsic fibers in CIFC have such a core.
In hematoxylin-stained sections an alternation of darkly and faintly stainable lines, both of which are about 2.5 μm thick, is seen within extrinsic fiber-poor and -free CIFC [14], [15], [16], [17], [18] (Fig. 4A). For convenience in description, these lines will be referred to as “lamellae” in this review. Scanning electron microscopy revealed that two types of lamellae, i.e. lamellae with longitudinally and near-longitudinally cut fibrils and those lamellae with transversely and near-transversely cut fibrils stratified alternately and created the alternating lamellae [16], [17], [18] (Fig. 4B). The alternating lamellae are very similar to those in compact bone [19], [20], [21]. In compact bone a twisted plywood model has been proposed to explain the alternating lamellae [19]. According to this model, all collagen fibrils run parallel in a given plane, and their direction rotates from plane to plane. A 180° rotation of fibril arrays corresponds to a period. This periodic change in fibril array orientation appears as the alternating lamellae when observed in histological sections. The alternating lamellae in CIFC conform to the twisted plywood model [16], [17], [18] (Fig. 4B). In a single CIFC the alternating lamellae are more distinct on the periodontal ligament side than on the dentin side (Fig. 4A). In other words, the lamellae are more obvious on the dentin side of an incremental line, and less obvious on the periodontal ligament side. How the periodic rotation of intrinsic fibers is created and how the structural disproportion appears will be discussed later.
Figure 4.
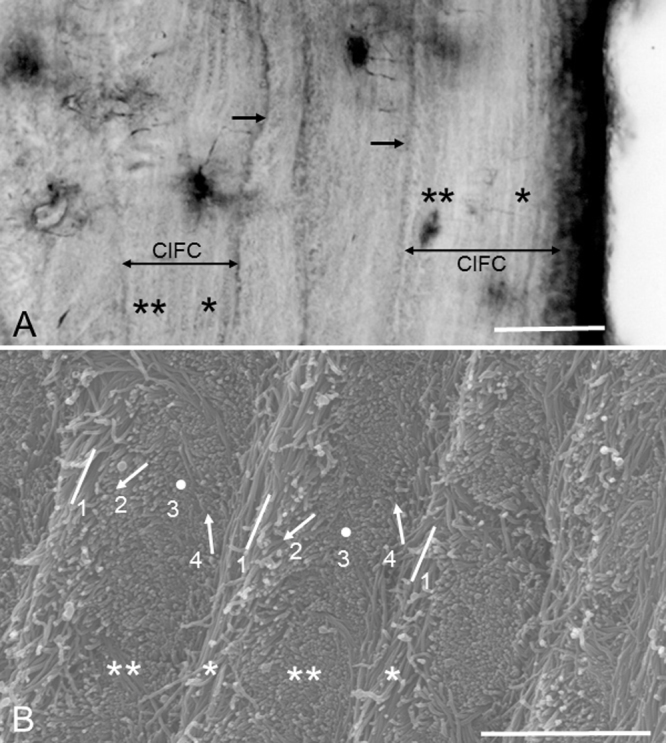
(A) Magnification of extrinsic fiber-free CIFC partitioned by incremental lines (arrows). In CIFC an alternation of darkly and faintly stainable lamellae is obvious on the periodontal ligament side (*) and non-obvious on the dentin side (**). Bar 30 μm. Hematoxylin-stained ground section. (B) Scanning electron micrograph showing the alternating lamellae. The specimen (a mandibular molar) has been treated by 10% NaOH maceration method to observe individual collagen fibrils clearly. Two types of lamellae, i.e. lamellae of longitudinally and near-longitudinally cut fibril arrays (*) and lamellae of transversely and near-transversely cut fibril arrays (**) create the alternating lamellae. Four types of fibril arrays are roughly recognized: 1, longitudinally cut fibril arrays; 2, obliquely cut fibril arrays facing downward; 3, transversely cut fibril arrays; 4, obliquely cut fibril arrays facing upward. As traced from left to right, the fibril arrays appear to rotate clockwise. Bar 3 μm.
3.4. Other varieties of cementum
3.4.1. Acellular afibrillar cementum (AAC)
AAC consists of a mineralized matrix containing neither collagen fibers nor cementocytes. AAC is found as isolated patches or as the most cervical part of AEFC on enamel just coronal to the cemento-enamel junction [3], [7], [8]. Its composition of non-collagenous proteins is very similar to that of AEFC. However, its function and origin have not yet been determined. Furthermore, it is still unknown whether AAC is an essential tissue for the tooth. Three possible origins have been proposed [7]: (1) Connective tissue-derived cells produce AAC. If this is true, connective tissue cells must replace reduced enamel epithelium. (2) AAC is an epithelial product, deposited by the inner enamel epithelial cells which are about to become Hertwig's epithelial root sheath. (3) AAC is a mere precipitate derived from tissue fluid or serum.
3.4.2. Intermediate cementum
The intermediate cementum has confused dental histologists since it first appeared in the literature and its origin is still controversial in the dental histological field. Hence, the intermediate cementum will be re-examined here.
In 1927, Bencze [22] first used the term “intermediate cementum (intermediäre Zementscicht)” to indicate a narrow part containing cellular elements and/or lacunae between dentin and CMSC (Fig. 5A, B). Earlier, Hopewell-Smith [23] had found a homogeneous layer between AEFC and the granular layer of Tomes (Fig. 5C). The homogeneous layer is now referred to as the hyaline layer of Hopewell-Smith. On the basis of elaborate histological observations, it is now established that the two tissues are homologous. The structure in question is referred to as the hyaline layer of Hopewell-Smith in the AEFC region and as the intermediate cementum in the CMSC region [3]. With regard to its origin, the two investigators proposed different views. Hopewell-Smith described the homogenous layer as a peripheral part of dentin. In contrast, Bencze [22] insisted that the intermediate cementum was a part of cementum, although he had earlier assumed it to be a part of dentin. The origin of the intermediate cementum has been a focus of many dental histologists. In histological sections where the cemento-dentinal junction is clearly differentiated, the intermediate cementum exists on the dentin side of the junction and there is no boundary between the intermediate cementum and dentin (Fig. 5A–C). This fact proves that the intermediate cementum is definitely a part of dentin. Nevertheless some investigators [24] have still emphasized that the intermediate cementum is not dentin, and further, that it is a particular tissue produced by Hertwig's epithelial root sheath.
Figure 5.

(A–C) Light micrographs showing the apical roots covered with CMSC (A, B) and cervical root covered with AEFC (C) in hematoxylin-stained ground sections. Bars 100 μm. (A, B) The intermediate cementum (*) contains lacunae (small arrows) and exists on the dentin side of the cemento-dentinal junction (large arrow). Arrowheads in (B) indicate the granular layer of Tomes. (C) The hyaline layer of Hopewell-Smith (*) is present between the granular layer of Tomes (arrowheads) and cemento-dentinal junction (arrow).
The investigators who studied the origin of the intermediate cementum can be divided into two groups. The first group believes that it is a part of dentin. Owens [25], [26] observed the intermediate cementum in human premolars and molars and found the continuity of dentinal tubules between the intermediate cementum and dentin. Hence, he concluded that the intermediate cementum was a surface layer of dentin (= mantle dentin or a part of mantle dentin). Schroeder [3] and Yamamoto [10], [27] agreed with this conclusion. In addition, Owens [25], [26] and subsequently, Kawasaki [28] elucidated that the hyaline layer of Hopewell-Smith started to be mineralized later than the deep part of dentin (= circumpulpal dentin). Afterwards, this finding became a significant key for another hypothesis of the second group.
The second group believes that the intermediate cementum is not a part of dentin, but an enameloid-like tissue produced by the epithelial root sheath. This hypothesis will be referred to as an epithelial origin hypothesis in this section. In brief, the epithelial origin hypothesis is based on the following logic [29], [30], [31]. (1) The cemento-dentinal junction or the innermost cementum layer in rodent teeth corresponds to the intermediate cementum in human teeth. (2) The cemento-dentinal junction is an epithelial sheath product in rodent teeth. (3) Hence, the intermediate cementum in human teeth is of epithelial sheath origin. Regarding the premise (1), approximately 1 μm thick superficial dentin layer shows delayed mineralization in initial acellular cementogenesis of rat molars [27], [32], [33]. Thereafter, an intensely hematoxylin-stainable, PAS-positive matrix appears in the superficial dentin layer and forms the cemento-dentinal junction [27], [32], [33], [34], [35]. In addition, many epithelial sheath cells are embedded near the cemento-dentinal junction in rat cellular cementogenesis [36], [37]. The cemento-dentinal junction of human teeth, like that of rat teeth, is intensely hematoxylin-stainable and PAS-positive (10). Nevertheless, based on the delayed mineralization and cell inclusion, the second group considers the cemento-dentinal junction in rat teeth to correspond to the intermediate cementum in human teeth. The superficial dentin layer in rat molars is a very narrow, fibril-poor tissue and obviously different from the mantle dentin which consists of densely packed dentin matrix fibers [27], [32], [33], [34], [35]. Further, it is established that the epithelial sheath cells are never embedded in the cementum in human cementogenesis [3], [4], [6], [7], [8], [9]. Hence, the premise (1) of the epithelial origin hypothesis is lacking supportive evidence. The premise (2) is still controversial, whether the hematoxylin-stainable matrix is a cementoblast product or epithelial sheath cell product is not yet determined. Even though the matrix is an epithelial product, this does not relate to the origin of the intermediate cementum. For these reasons, the epithelial origin hypothesis is questionable, based on uncertain premises.
Fujita [38] regarded the intermediate cementum as a peripheral part of dentin and added, “Most of the lacunae in the intermediate cementum are considered to be swollen dentinal tubules, and thus the cellular elements would be swollen processes of odontoblasts. If the lacunae contain cell bodies, they would be those of odontoblasts which have failed to retreat.” Actually, Yamamoto [27] found that the lacunae in question were definitely continuous with dentinal tubules. Hence, to date Fujita's view [38] appears most reasonable. In conclusion in this section, the intermediate cementum is a part of dentin, not cementum, and this term may be reconsidered to avoid unnecessary confusion.
4. Composition of organic matrix
There have been no investigations which have detected the chemical composition separately in individual cementum types. The following description is not for a particular type of cementum.
4.1. Collagens
The organic matrix of cementum consists predominantly of collagens. In bovine cementum type I collagen accounts for more than 90% of the organic matrix and type III collagen approximately 5% [39]. In human cementum type I collagen appears to be the only collagen type [40]. The collagens form cross-striated fibrils in cementum and induce biological mineralization as a scaffold for the mineral crystals during mineralization, and maintain the structural integrity of cementum after mineralization.
4.2. Non-collagenous proteins
Major non-collagenous proteins are bone sialoprotein and osteopontin [7], [8]. They play important roles in the mineralization process, binding collagen fibrils and hydroxyapatite. After mineralization, they serve to maintain structural integrity of cementum. By immunohistochemistry AEFC contains the two glycoproteins more densely than CIFC. Their high density in AEFC is probably associated with its slower formation speed.
Proteoglycans are complex macromolecules composed of a core protein to which glycosaminoglycans are covalently attached. By immunohistochemistry dermatan sulfate, chondroitin sulfate, and keratan sulfate are detected as glycosaminoglycans. Regarding the types of proteoglycans, versican as large proteoglycan and decorin, biglycan, and lumican as small proteoglycans are detected. They exist exclusively in cellular cementum [41], [42]. It is proposed that proteoglycans inhibit mineralization of collagen fibrils by possessing specific sites on collagen fibrils normally destined to be filled with hydroxyapatite. Thus the proteoglycan content becomes lower after mineralization [41].
The presence of enamel-related proteins as constituents of cementum is still controversial. Although some investigators [43], [44] proposed that these proteins in cementum have cementoblast-inducing activity, this view is still presently unconfirmed.
Great efforts have been made, to identify unique cementum-specific marker proteins. Cementum-derived growth factor, cementum attachment protein, and cementum protein-23 had been the candidates, but were all later found in tissues other than cementum [45]. To date, marker proteins specific to only cementum are not yet discovered.
5. Function of cementum
The main function of cementum is tooth support or tooth anchorage together with the principal fibers and alveolar bone. AEFC is therefore the most suitable cementum for tooth support [3], [4], [5], [7]. The function of CIFC is more complicated. Extrinsic fiber-poor and -free CIFC do not appear to contribute to tooth support. Instead their function is adaptation, i.e. reshaping the root surface during tooth movement and compensating for crown wear [2], [3], [4], [5]. Such CIFC also appears as reparative cementum which fills resorbed root surfaces. As described previously, extrinsic fiber-poor and -free CIFC have the alternating lamellae, based on the twisted plywood structure. In compact bone, the structure is considered to resist stresses from various directions [46]. In the same way, the alternating lamellae in CIFC may function to resist multi-directional masticatory stresses. In contrast, extrinsic fiber-rich CIFC may serve as tooth support more than adaptation. When the adaptation is required, extrinsic fiber-poor or -free CIFC forms patch-wise on applicable portions. In contrast, when tooth anchorage is required, extrinsic fiber-rich CIFC or AEFC forms.
CMSC is generally thicker in molars than in anterior teeth. The reason may be deduced simply; the thickness of CMSC is parallel to the masticatory stress loaded on the tooth. However, it is often found that impacted or pre-functional molars, like fully functioning molars, possess thick CMSC. In relation to the CMSC thickness, Schroeder [3] commented, “CMSC distribution and thickness of a particular tooth may reflect its past history of eruption rather than masticatory function. Pre- and post-eruptive tooth movements may thus, necessitate CMSC deposition.” This view has not yet been established, but appears reasonable.
6. Development of cementum
Due to ethical issues, there have been no reports which investigate human cementogenesis precisely. The following section is fundamentally based on previous reports [3], [4], [6], [7], [8], [9], [47], in which developing human premolars were observed from apical to cervical regions to follow cementogenesis. In some portions, findings from rat cementogenesis will be used supplementally and to a minimum.
6.1. Acellular extrinsic fiber cementum (AEFC)
Premolars with roots developed to 50–60% of their presumable final length were examined [4], [6], [7]. At the forming root tip Hertwig's epithelial root sheath proliferates and grows apically, inducing dental papilla cells to differentiate into odontoblasts. Where the initial dentin mineralization starts, the external and internal surface of dentin is not yet mineralized, and thus unmineralized dentin, i.e. predentin, depicts a V-shape (Fig. 6, section 1). In premolars the distance between mineralized dentin tip and predentin tip (= forming root tip) is about 50 μm. Cervically, the epithelial sheath separates from the root surface and disintegrates into cell clusters, i.e. epithelial cell rests of Malassez. Cementoblasts with fibroblast-like shape and collagen fibrils subsequently appear on the exposed dentin surface. Ends of the fibrils intermingles with predentin matrix fibers. The fibrils aggregate into short bundles, which are arranged in parallel and oriented almost perpendicularly to the root surface (Fig. 6, section 1). At this point the fibril bundles are not yet connected with periodontal ligament fibers which are arranged parallel to the root surface. Hence, in the strict sense the fibril bundles are not extrinsic fibers. In accordance with previous reports [4], [6], [7], they will be referred to as “fiber fringe” in this section. From findings of human and rat AEFC formation [4], [48], [49], the fibril-aggregating or fringe-generating mechanism is presumed as follows. Cementoblasts as well as fibroblasts extend wing- or plate-like processes and surround the immature fibril bundles. They then form tubular compartments by connecting the processes in concert with other cementoblasts (Fig. 7), and add collagen fibrils linearly and laterally to the immature fibril bundles within the compartments. As a result, the fiber fringe is generated on the root surface.
Figure 6.

Schematic diagram depicting AEFC genesis. Section 1: Cementoblasts appear and start to form fiber fringe on the unmineralized dentin. Periodontal ligament fibers are arranged in parallel with the root surface. Section 2: Fiber fringe with maximum density is established. Dentin mineralization reaches the base of fiber fringe and progresses into the fringe. Section 3: Fiber fringe elongates and begins to connect with periodontal ligament fibers. Section 4: The tooth anchorage system, or principal fiber-extrinsic fiber linkage, is established. CB, cementoblasts; ERM, epithelial cell rests of Malassez; FF, fiber fringe; HERS, Hertwig's epithelial root sheath; MD, mineralized dentin; PLF, periodontal ligament fibers; UMD, unmineralized dentin.
Modified from Schroeder [4].
Figure 7.

(A, B) Cementoblasts on established AEFC in rat molars by transmission (A) and scanning electron microscopy (B). Bars 30 μm. (A) Cementoblasts encircle principal fibers with cytoplasmic processes in a tangential section through cementum surface. (B) The specimen has been treated by KOH-collagenase method and thereby collagen fibers and interfibrillar matrix are removed and only cementoblasts can be selectively observed. Cementoblasts are viewed from the cementum side. Cementoblasts form cylindrical compartments with wing-like processes.
The mineralization of the external dentin proceeds outward and eventually reaches the base of fiber fringe. This point is about 300 μm distant from the root tip in premolars. Then the fiber fringe begins to be mineralized and can be recognized as the AEFC matrix (Fig. 6, section 2). At the same time bone sialoprotein and osteopontin, secreted from cementoblasts, become detectable in the cementum. Mineralized spherules also emerge as isolated patches in the vicinity of the fiber fringe. This means that early mineralization of the fiber fringe starts from two sites. After the two mineralization fronts coalesce, mineralization proceeds outward with additional AEFC formation.
More cervically, AEFC increases in thickness and the fiber fringe becomes more elongated. The periodontal ligament fibers develop further and change their arrangement to form principal fibers (Fig. 6, section 3). In premolars, when the cervical AEFC reaches about 15–20 μm thickness or the tooth is about to start occlusion, the fiber fringe becomes continuous with the principal fibers [4], [7], [9]. At this point the fiber fringe is organized into extrinsic fibers and the tooth anchorage system is established in the cervical region (Fig. 6, section 4). Thereafter, cementoblasts concentrate on secretion of non-collagenous proteins to induce further AEFC mineralization and strengthen the anchorage system. The mechanism by which the fiber fringe connects to the principal fibers is still unknown. As described previously, cementum increases in thickness with age. According to Sequeira et al. [50], the growth rate of cervical AEFC is approximately 2.9 μm/year in maxillary first premolars and approximately 1.5 μm/year in mandibular second premolars.
6.2. Cellular mixed stratified cementum (CMSC)
CIFC usually forms as the first cementum in CMSC genesis [3], [4], [5], [6], [7] (Fig. 3A–C). It has been widely believed that occlusal stimuli trigger CISC genesis. However, Bosshardt and Selvig [7] claimed, “It seems that the initiation of CIFC genesis does not depend on stimuli transmitted by masticatory forces and that influence by pressure may reduce the rate of matrix deposition.” The trigger for CIFC deposition is still unclear.
The development of extrinsic fiber-free CIFC will be here described for the initial genesis of CMSC. Following findings were obtained from premolars with roots developed to 75% of their presumable final length [4], [6], [7]. At the forming root tip the CIFC deposition starts almost concurrently with dentin formation. Cementoblasts appear on the predentin immediately after Hertwig's epithelial sheath detaches from it (Fig. 8, section 1). The cementoblasts assemble densely and begin to secrete collagen fibrils in various directions. These fibrils are therefore randomly arranged in the initial CIFC and intermingle with predentin matrix fibers. As collagen fibrils are densely accumulated, cementoblasts show concaves on the whole cell surface, which compartmentalize the newly formed fibril bundles. In accordance with Bosshardt and Schroeder [47], these cementoblasts are producing CIFC matrix in a multipolar mode causing rapid matrix deposition. Some cementoblasts are embedded as cementocytes in the rapidly growing matrix. At this point the CIFC matrix is still unmineralized, meaning it is still precementum (Fig. 8, section 1). The external dentin is mineralized much faster than in AEFC genesis. After the dentin mineralization extends to CIFC (approximately 100–200 μm from the root tip), cementoblasts shift their matrix-producing mode from multipolar to unipolar, and produce CIFC matrix slowly only in parallel with root surface. At the same time, bone sialoprotein and osteopontin become detectable in the mineralized CIFC. A narrow precementum containing numerous mineralized spherules appears on the CIFC surface (Fig. 8, section 2).
Figure 8.

Schematic diagram depicting the formation of extrinsic fiber-free CIFC as the initial CMSC genesis. Section 1: Cementoblasts appear and produce the cementum matrix rapidly in a multipolar mode on the unmineralized dentin. Section 2: Dentin mineralization reaches the cementum matrix and progresses into it. Cementoblasts produce cementum matrix slowly in a unipolar mode. Unmineralized cementum matrix is recognized as precementum. CB/m, cementoblasts with multipolar matrix production; CB/u, cementoblasts with unipolar matrix production; CM, cementum matrix; ERM, epithelial cell rests of Malassez; HERS, Hertwig's epithelial root sheath; MCM, mineralized cementum matrix; MD, mineralized dentin; UMD, unmineralized dentin; PC, precementum.
Modified from Schroeder [4].
After the initial CIFC is established on the root surface remote from the root tip, CIFC and AEFC form in unpredictable order with resting phases. As described previously, the surrounding environment or requirement (adaptation and tooth anchorage) determines the type of cementum. In other words, according to the ratio of the two requirements, various types of cementum, i.e. AEFC, extrinsic fiber-rich CIFC, extrinsic fiber-poor CIFC, and extrinsic fiber-free CIFC, are generated. At the same time, the matrix-producing modes may also be determined to control the cementogenesis speed [4]. When the cementum deposition shifts from extrinsic fiber-rich CIFC to extrinsic fiber-poor and -free CIFC, all or part of the extrinsic fibers lose continuity with the principal fibers.
In a series of studies of human and rat cementogenesis, two morphologically different types of cementoblasts were found in the advanced CIFC formation [16], [17], [18]. The first type was found on the extrinsic fiber-poor or -free CIFC, and second type on the extrinsic fiber-rich CIFC. The first type had long finger-like processes and was suggested to secrete intrinsic fibers in close and parallel association with the processes. The cementoblasts of this type extended the processes together in the same direction on the CIFC surface where the alternating lamellae were generating [16], [17], [18] (Fig. 9). These findings lead to the following conclusion: the cementoblasts control the intrinsic fiber arrangement with the finger-like processes. They move the processes synchronously and periodically and cause a periodic change in intrinsic fiber arrangement. This dynamic sequence results in the alternating lamellae. Osteoblasts were also recently suggested to create the alternating lamellae in a similar fashion in rat compact bone [51]. The structural disproportion of the lamellae has been interpreted as follows: whenever cementoblasts start to deposit cementum matrix again after a resting phase, the cells cannot yet organize intrinsic fibers sufficiently. The intrinsic fibers are accordingly poorly organized. As the cementoblasts recovers fiber-organizing activity, the intrinsic fibers become well-organized to create the alternating lamellae [15], [16], [17], [18]. Cementum increases in degree of mineralization toward the periodontal ligament side between adjoining incremental lines [52], which suggests that cementoblasts recover mineralization-inducing activity as well.
Figure 9.

(A) Transmission electron micrographs showing the CIFC surface where alternating lamellae are generating. Flat cementoblasts cover a lamella (*) of transversely and near-transversely cut fibrils. Bar 2 μm. (B) Magnification of the boxed area in (A). A long, thin process (large arrow) is in a close and parallel association with longitudinally cut fibrils (small arrows). Bar 2 μm.
The second type had wing-like and finger-like processes. The cementoblasts connected the wing-like processes to form tubular compartments surrounding the principal fibers. The finger-like processes, arranged in parallel with the cementum surface, were formed on the cementum-facing side of the wing-like processes. Intrinsic fibers emerged in close and parallel association with the finger-like processes (Fig. 10). From these findings the following was suggested (Fig. 11): when the cementoblasts move away from the cementum or are embedded in the cementum, these cells retract the wing-like processes, dividing them into finger-like processes. At the same time they secrete fibrils along the finger-like processes to fill the spaces around the principal fibers. As a result, the intrinsic fibers encircle the extrinsic fibers in extrinsic fiber-rich CIFC [18], [53], [54].
Figure 10.
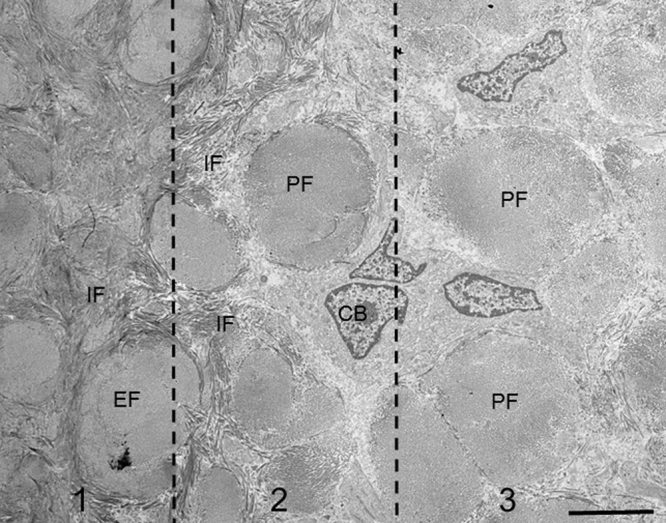
Transmission electron micrograph showing the extrinsic fiber-rich CIFC surface in a tangential section through the cementum surface. Three sections are divided. Section 1 indicates the interior of cementum, where intrinsic fibers (IF) surround extrinsic fibers (EF). Section 2 indicates the cementum surface. Intrinsic fibers and cementoblasts (CB) surround principal fibers. Section 3 indicates an area slightly distant from the cementum surface. Cementoblasts (CB) surrounds principal fibers with cytoplasmic processes. Only a few or no intrinsic fibers are seen around principal fibers. Bar 5 μm.
Figure 11.
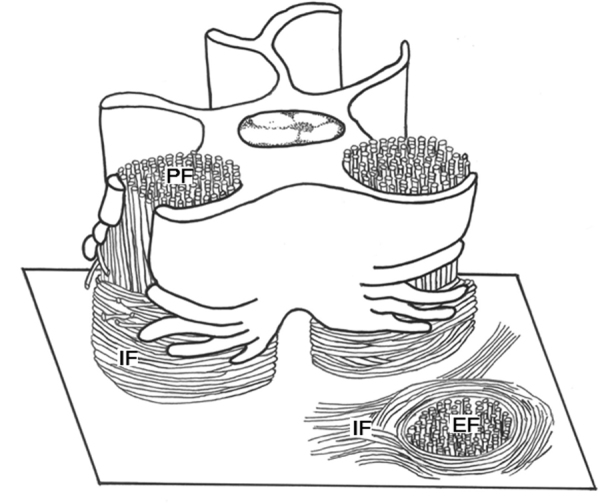
Schematic diagram depicting how cementoblasts produce intrinsic fibers around principal fibers on extrinsic fiber-rich CIFC. Cementoblasts surround principal fibers (PF) in cylindrical compartments with wing-like processes. With further cementogenesis, they retract the wing-like processes and divide them into finger-like processes. At the same time they secrete intrinsic fibers (IF) along the finger-like processes. As a result, the intrinsic fibers encircle the extrinsic fibers (EF) in the cementum.
Modified from Yamamoto et al. [54].
6.3. Origin of cementoblasts
Cementoblasts have long been believed to derive from mesenchymal dental follicle [3], [45], [55], [56]. This concept will be referred to as a classical mesenchymal hypothesis in this section [45]. In the 1980s and 1990s a different hypothesis has arisen from in vitro and in vivo experiments using rats and mice, namely that some epithelial sheath cells transdifferentiate into cementoblasts by epithelial-mesenchymal transition (EMT). This hypothesis will be referred to as an alternative epithelial hypothesis [45]. The alternative epithelial hypothesis has now almost surpassed the classical mesenchymal hypothesis, because the alternative epithelial hypothesis can explain why epithelial sheath cells decrease in number during epithelial sheath disintegration [56], [57], [58], [59]. EMT is a normal phenomenon in embryos in several developing organs, e.g. in palatogenesis [60], [61]. However, EMT of epithelial sheath during cementogenesis is not yet determined.
The alternative epithelial hypothesis has been first proposed in an in vitro study by Thomas [62]. He found that cultured epithelial sheath cells changed shape into mesenchymal cell phenotype and co-expressed vimentin and keratin, which are markers of mesenchymal and epithelial cells, respectively. Based on the fact that epithelial cells undergoing EMT co-express vimentin and keratin [60], [61], Thomas [62] proposed that the cultured epithelial sheath cells transformed into mesenchymal cementoblasts, and further that epithelial sheath cells similarly transformed into cementoblasts through EMT during in vivo normal cementogenesis. Subsequently, findings supportive of the alternative epithelial hypothesis have been accumulated in in vitro [63], [64], [65] and in vivo studies [66], [67], [68]. It is likely that cultured epithelial sheath cells undergo EMT under special conditions facilitating EMT. However, results of the in vivo studies are questionable, because in these studies cementoblasts and epithelial cells were probably misidentified. In the in vivo studies, which used rat and mouse teeth [66], [67], [68], some cementoblasts and cementocytes were keratin-immunoreactive, and thus these cells were regarded as the epithelial sheath cells which were undergoing EMT at that exact moment. However, in rat and mouse cementogenesis [36], [37], [56], [57], it is established that epithelial cell rests of Malassez coexist with true cementoblasts on the cementum surface and that many epithelial sheath cells are embedded in the cellular cementum. Therefore, in the in vivo studies epithelial cell rests and embedded epithelial cells were probably misidentified as cementoblasts and cementocytes, respectively. Findings supportive of the classical mesenchymal hypothesis have also accumulated in in vivo studies using mouse and rat teeth [56], [58], [59], [69], [70], [71]. In these studies any epithelial sheath-derived cells did not express mesenchymal or cementoblast characteristics, e.g. immunoreactivity for vimentin, bone sialoprotein, and osteopontin. Taken together, to date there is no evidence to support the alternative epithelial hypothesis in in vivo cementogenesis.
The epithelial cells decrease in number during epithelial sheath disintegration when observed in histological sections [56], [57], [58], [59]. If EMT does not occur, how can the cell number reduction be explained? Apoptosis may be another possible cause for the cell number reduction, but it has been shown that apoptosis occurs in epithelial cell rests of Malassez and epithelial cells embedded in cementum, but does not in the disintegrating epithelial sheath [59], [72], [73]. Recent studies [56], [58], [59] have suggested that the cell number reduction may be related to the growth rate and shape of the epithelial sheath (Fig. 12). In brief, the epithelial sheath bends toward the dental papilla and forms a tapered cylinder. During root formation the epithelial sheath maintains the tapered shape and proliferates only at the apical end. In contrast, the root grows quite fast and almost straight. In this situation, the discrepancy in surface area could be generated between the epithelial sheath and the growing root. As a result, the epithelial sheath is stretched out and covers only small portions of the root surface. Accordingly, the cell numbers are observed to reduce in histological sections, although the actual cell number is unchanged.
Figure 12.

Schematic diagrams depicting the disintegration of Hertwig's epithelial root sheath during root development. (A) Hertwig's epithelial root sheath (HERS) bends inside toward dental pulp (DP) at the tip of developing root (R). (B) Three-dimensionally, the epithelial sheath forms a tapered cylinder. (C) Root formation proceeds almost straight, whereas the epithelial sheath maintains the tapered shape. As a result, due to the discrepancy in surface area, the epithelial sheath is stretched out and fragmented into epithelial cell rests of Malassez.
Modified from Yamamoto et al. [58].
In concluding this section, the classical mesenchymal hypothesis is still convincing, namely cementoblasts derive from dental follicle and no epithelial sheath cells transform into cementoblasts.
Conflict of interest
None.
References
- 1.Denton G.B. The discovery of cementum. J Dent Res. 1939;18:239. [Google Scholar]
- 2.Jones S.J. Cement. In: Osborn J.W., editor. Dental anatomy and embryology. Blackwell; Oxford: 1981. pp. 193–205. [Google Scholar]
- 3.Schroeder H.E. Cementum. In: Schroeder H.E., editor. The periodontium. Springer; Berlin: 1986. pp. 23–127. [Google Scholar]
- 4.Schroeder H.E. Biological problems of regenerative cementogenesis: synthesis and attachment of collagenous matrices on growing and established root surface. Int Rev Cytol. 1992;142:1–59. doi: 10.1016/s0074-7696(08)62074-4. [DOI] [PubMed] [Google Scholar]
- 5.Schroeder H.E. Human cellular mixed stratified cementum: a tissue with alternating layers of acellular extrinsic- and cellular intrinsic fiber cementum. Schweiz Monatsschr Zahnmed. 1993;103:550–560. [PubMed] [Google Scholar]
- 6.Bosshardt D.D., Schroeder H.E. Cementogenesis reviewed. A comparison between human premolars and rodent molars. Anat Rec. 1996;245:267–292. doi: 10.1002/(SICI)1097-0185(199606)245:2<267::AID-AR12>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 7.Bosshardt D.D., Selvig K.A. Dental cementum: the dynamic tissue covering of the root. Periodontology 2000. 1997;13:41–75. doi: 10.1111/j.1600-0757.1997.tb00095.x. [DOI] [PubMed] [Google Scholar]
- 8.Bosshardt D.D., Zalzal S., Mckee M.C., Nanci A. Developmental appearance and distribution of bone sialoprotein and osteopontin in human and rat cementum. Anat Rec. 1998;250:13–33. doi: 10.1002/(SICI)1097-0185(199801)250:1<13::AID-AR3>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 9.Nanci A., Bosshardt D.D. Structure of periodontal tissues in health and disease. Periodontology 2000. 2006;40:11–28. doi: 10.1111/j.1600-0757.2005.00141.x. [DOI] [PubMed] [Google Scholar]
- 10.Yamamoto T., Domon T., Takahashi S., Islam N., Suzuki R., Wakita M. The structure and function of the cemento-dentinal junction in human teeth. J Periodontal Res. 1999;34:261–268. doi: 10.1111/j.1600-0765.1999.tb02252.x. [DOI] [PubMed] [Google Scholar]
- 11.Yamamoto T., Domon T., Takahashi S., Islam M.N., Suzuki R. The fibrous structure of the cemento-dentinal junction in human molars shown by scanning electron microscopy combined with NaOH-maceration. J Periodontal Res. 2000;35:59–64. doi: 10.1034/j.1600-0765.2000.035002059.x. [DOI] [PubMed] [Google Scholar]
- 12.Yamamoto T., Domon T., Takahashi S., Islam M.N., Suzuki R. The fibrillar structure of the cemento-dentinal junction in different kinds of human teeth. J Periodontal Res. 2001;36:317–321. doi: 10.1034/j.1600-0765.2001.360507.x. [DOI] [PubMed] [Google Scholar]
- 13.Fujii Y. Mineralization pattern of human cementum. Shigaku. 1969;57:206–228. [in Japanese with English abstract] [PubMed] [Google Scholar]
- 14.Yamamoto T., Domon T., Takahashi S., Islam M.D., Suzuki R., Wakita M. The regulation of fiber arrangement in advanced cellular cementogenesis of human teeth. J Periodontal Res. 1998;33:83–90. doi: 10.1111/j.1600-0765.1998.tb02296.x. [DOI] [PubMed] [Google Scholar]
- 15.Chen M. Observations on the structure of matrix fibers in human cementum. J Stomtol Soc Jpn. 1987;54:635–675. doi: 10.5357/koubyou.54.635. [in Japanese with English abstract] [DOI] [PubMed] [Google Scholar]
- 16.Yamamoto T., Domon T., Takahashi S., Wakita M. Formation of alternate lamellar pattern in the advanced cellular cementogenesis in human teeth. Anat Embryol. 1997;196:115–121. doi: 10.1007/s004290050084. [DOI] [PubMed] [Google Scholar]
- 17.Yamamoto T., Domon T., Takahashi S., Islam N., Suzuki R. Twisted plywood structure of an alternating lamellar pattern in cellular cementum of human teeth. Anat Embryol. 2000;202:25–30. doi: 10.1007/pl00008241. [DOI] [PubMed] [Google Scholar]
- 18.Yamamoto T., Li M., Liu Z., Guo Y., Hasegawa T., Masuki H. Histological review of the human cellular cementum with special reference to an alternating lamellar pattern. Odontology. 2010;98:102–109. doi: 10.1007/s10266-010-0134-3. [DOI] [PubMed] [Google Scholar]
- 19.Giraud-Guille M.M. Twisted plywood architecture of collagen fibrils in human compact bone osteons. Calcif Tissue Int. 1988;42:167–180. doi: 10.1007/BF02556330. [DOI] [PubMed] [Google Scholar]
- 20.Ziv V., Sabanay I., Arad T., Traub W., Weiner S. Transitional structures in lamellar bone. Microsc Res Tech. 1996;33:203–213. doi: 10.1002/(SICI)1097-0029(19960201)33:2<203::AID-JEMT10>3.0.CO;2-Y. [DOI] [PubMed] [Google Scholar]
- 21.Weiner S., Traub W., Wagner H.D. Lamellar bone: structure-function relations. J Struct Biol. 1999;126:241–255. doi: 10.1006/jsbi.1999.4107. [DOI] [PubMed] [Google Scholar]
- 22.Bencze L. Befunde an der Dentinzementgrenze. Z Stomatol. 1927;25:877–896. [Google Scholar]
- 23.Hopewell-Smith A. Concerning human cementum. J Dent Res. 1920;2:59–76. [Google Scholar]
- 24.Avery J.K., Chiergo D.J., Jr. Cementum. In: Avery J.K., Chiergo D.J. Jr., editors. Essential of oral histology and embryology. A clinical approach. 3rd ed. Mosby Inc.; St. Louis: 2006. pp. 137–144. [Google Scholar]
- 25.Owens P.D.A. Light microscopic observations on the formation of the layer of Hopewell-Smith in human teeth. Arch Oral Biol. 1972;17:1785–1788. doi: 10.1016/0003-9969(72)90243-9. [DOI] [PubMed] [Google Scholar]
- 26.Owens P.D.A. The root surface in human teeth: a microradiographic study. J Anat. 1976;122:389–401. [PMC free article] [PubMed] [Google Scholar]
- 27.Yamamoto T. The structure and origin of Hopewell-Smith Hyaline layer and intermediate cementum in human and rat teeth. J Jpn Soc Periodontol. 2000;42:298–306. [in Japanese with English abstract] [Google Scholar]
- 28.Kawasaki K. On the configuration of incremental lines in human dentine as revealed by tetracycline labelling. J Anat. 1975;119:61–66. [PMC free article] [PubMed] [Google Scholar]
- 29.Lindskog S. Formation of intermediate cementum. I. Early mineralization of aprismatic enamel and intermediate cementum in monkey. J Craniofac Genet Dev Biol. 1982;2:147–160. [PubMed] [Google Scholar]
- 30.Lindskog S. Formation of intermediate cementum. II. A scanning electron microscopic study of the epithelial root sheath of Hertwig in monkey. J Craniofac Genet Dev Biol. 1982;2:161–169. [PubMed] [Google Scholar]
- 31.Lindskog S., Hammarström L. Formation of intermediate cementum. III. 3H-tryptophan and 3H-proline uptake into the epithelial root sheath of Hertwig in vitro. J Craniofac Genet Dev Biol. 1982;2:171–177. [PubMed] [Google Scholar]
- 32.Owens P.D.A. A light and electron microscopic study of the early stages of root surface formation in molar teeth in the rat. Arch Oral Biol. 1980;24:901–907. doi: 10.1016/0003-9969(79)90215-2. [DOI] [PubMed] [Google Scholar]
- 33.Yamamoto T., Wakita M. Initial attachment of principal fibers to the root dentin surface in rat molars. J Periodontal Res. 1990;25:113–119. doi: 10.1111/j.1600-0765.1990.tb00901.x. [DOI] [PubMed] [Google Scholar]
- 34.Paynter K.J., Pudy G. A study of the structure, chemical nature, and development of cementum in the rat. Anat Rec. 1958;131:233–251. doi: 10.1002/ar.1091310207. [DOI] [PubMed] [Google Scholar]
- 35.Yamamoto T. The innermost layer of cementum in rat molars: its ultrastructure, development, and calcification. Arch Histol Jpn. 1986;49:459–481. doi: 10.1679/aohc.49.459. [DOI] [PubMed] [Google Scholar]
- 36.Lester K.S. The incorporation of epithelial cells by cementum. J Ultrastruct Res. 1969;27:63–87. [PubMed] [Google Scholar]
- 37.Lester K.S. The unusual nature of root formation in molar teeth of the laboratory rat. J Ultrastruct Res. 1969;28:481–506. doi: 10.1016/s0022-5320(69)80035-3. [DOI] [PubMed] [Google Scholar]
- 38.Fujita K. Cementum. In: Fujita K., editor. Histology of tooth. 1st ed. Ishiyaku Shuppan; Tokyo: 1983. pp. 133–146. [Google Scholar]
- 39.Birkedal-Hansen H., Butler W.T., Taylor R.E. Proteins of the periodontium. Characterization of the insoluble collagens of bovine dental cementum. Calcif Tissue Res. 1977;23:39–44. doi: 10.1007/BF02012764. [DOI] [PubMed] [Google Scholar]
- 40.Christner P., Robinson P., Clark C.C. A preliminary characterization of human cementum collagen. Calcif Tissue Res. 1977;23:147–150. doi: 10.1007/BF02012780. [DOI] [PubMed] [Google Scholar]
- 41.Ababneh K.T., Hall R.C., Embry G. Immunolocalization of glycosaminoglycans in aging, healthy and periodontally diseased human cementum. Arch Oral Biol. 1998;43:235–246. doi: 10.1016/s0003-9969(98)00001-6. [DOI] [PubMed] [Google Scholar]
- 42.Ababneh K.T., Hall R.C., Embery G. The proteoglycans of human cementum: immunohistochemical localization in healthy, periodontally involved and aging teeth. J Periodontal Res. 1999;34:87–96. doi: 10.1111/j.1600-0765.1999.tb02227.x. [DOI] [PubMed] [Google Scholar]
- 43.Hammarström L. Origins of cementum. Oral Dis. 1996;2:63–69. doi: 10.1111/j.1601-0825.1996.tb00205.x. [DOI] [PubMed] [Google Scholar]
- 44.Hammarström L. Enamel-matrix, cementum development and regeneration. J Clin Periodontol. 1997;24:658–668. doi: 10.1111/j.1600-051x.1997.tb00247.x. [DOI] [PubMed] [Google Scholar]
- 45.Foster B.L., Popowics T.E., Fong H.K., Somerman M.J. Advances in defining regulators of cementum development and periodontal regeneration. Curr Top Dev Biol. 2007;78:47–126. doi: 10.1016/S0070-2153(06)78003-6. [DOI] [PubMed] [Google Scholar]
- 46.Ascenzi A., Bonnuci E. The compressive properties of single osteons. Anat Rec. 1968;161:377–392. doi: 10.1002/ar.1091610309. [DOI] [PubMed] [Google Scholar]
- 47.Bosshardt D.D., Schroeder H.E. Evidence for rapid multipolar and slow unipolar production of human cellular and acellular cementum matrix with intrinsic fibers. J Clin Periodontol. 1990;17:663–668. [PubMed] [Google Scholar]
- 48.Yamamoto T., Wakita M. Bundle formation of principal fibers in rat molars. J Periodontal Res. 1992;27:20–27. doi: 10.1111/j.1600-0765.1992.tb02081.x. [DOI] [PubMed] [Google Scholar]
- 49.Yamamoto T., Domon T., Takahashi S., Islam N., Suzuki R., Wakita M. The structure and function of periodontal ligament cells in acellular cementum in rat molars. Ann Anat. 1998;180:519–522. doi: 10.1016/S0940-9602(98)80058-7. [DOI] [PubMed] [Google Scholar]
- 50.Sequeira P., Bosshardt D.D., Schroeder H.E. Growth of acellular extrinsic fiber cementum (AEFC) and density of inserting fibers in human premolars of adolescents. J Periodontal Res. 1992;27:134–142. doi: 10.1111/j.1600-0765.1992.tb01815.x. [DOI] [PubMed] [Google Scholar]
- 51.Yamamoto T., Hasegawa T., Sasaki M., Hongo H., Tabata C., Liu Z. Structure and formation of the twisted plywood pattern of collagen fibrils in rat lamellar bone. J Electron Microsc. 2012;61:113–121. doi: 10.1093/jmicro/dfs033. [DOI] [PubMed] [Google Scholar]
- 52.Matsuo A. Study of the lamellar structure of cementum, its degree of mineralization and fibrous components by light and scanning electron microscopy and by contact microradiography. Higashi Nippon Dent J. 1993;12:193–217. [in Japanese with English abstract] [Google Scholar]
- 53.Yamamoto T., Hinrichsen K.V. The development of cellular cementum in rat molars, with special reference to the fiber arrangement. Anat Embryol. 1993;188:537–549. doi: 10.1007/BF00187009. [DOI] [PubMed] [Google Scholar]
- 54.Yamamoto T., Domon T., Takahashi S., Wakita M. Cellular cementogenesis in rat molars: the role of cementoblasts in the deposition of intrinsic matrix fibers of cementum proper. Anat Embryol. 1996;193:495–500. doi: 10.1007/BF00185880. [DOI] [PubMed] [Google Scholar]
- 55.Cho M.I., Garant P.R. Development and general structure of the periodontium. Periodontology 2000. 2000;24:9–27. doi: 10.1034/j.1600-0757.2000.2240102.x. [DOI] [PubMed] [Google Scholar]
- 56.Diekwisch T.G.H. The developmental biology of cementum. Int J Dev Biol. 2001;45:695–706. [PubMed] [Google Scholar]
- 57.Luan X., Ito Y., Diekwisch T.G.H. Evolution and development of Hertwig's epithelial root sheath. Dev Dyn. 2006;235:1167–1180. doi: 10.1002/dvdy.20674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Yamamoto T., Yamamoto T., Yamada T., Hasegawa T., Hongo H., Oda K. Hertwig's epithelial root sheath cell behavior during initial acellular cementogenesis in rat molars. Histochem Cell Biol. 2014;142:489–496. doi: 10.1007/s00418-014-1230-1. [DOI] [PubMed] [Google Scholar]
- 59.Yamamoto T., Yamada T., Yamamoto T., Hasegawa T., Hongo H., Oda K. Hertwig's epithelial root sheath fate during initial cellular cementogenesis in rat molars. Acta Histochem Cytochem. 2015;48:95–101. doi: 10.1267/ahc.15006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fitchett J.E., Hay E.D. Medial edge transform to mesenchyme after embryonic palatal shelves fuse. Dev Biol. 1989;131:455–474. doi: 10.1016/s0012-1606(89)80017-x. [DOI] [PubMed] [Google Scholar]
- 61.Gibbins J.R., Manthey A., Tazawa Y.M., Scott B., Bloch-Zupan A., Hunter N. Midline fusion in the formation of secondary palate anticipated by upregulation of keratinK5/6 and localized expression of vimentin mRNA in medial edge epithelium. Int J Dev Biol. 1999;43:237–244. [PubMed] [Google Scholar]
- 62.Thomas H.F. Root formation. Int J Dev Biol. 1995;39:231–237. [PubMed] [Google Scholar]
- 63.Zeichner-David M., Oishi K., Su Z., Zakartchenco V., Chen L.S., Arzate H. Role of Hertwig's epithelial root sheath cells in tooth root development. Dev Dyn. 2003;228:651–663. doi: 10.1002/dvdy.10404. [DOI] [PubMed] [Google Scholar]
- 64.Sonoyama W., Seo B.M., Yamaza T., Shi S. Human Hertwig's epithelial root sheath cells play crucial roles in cementum formation. J Dent Res. 2007;86:594–599. doi: 10.1177/154405910708600703. [DOI] [PubMed] [Google Scholar]
- 65.Akimoto T., Fujiwara N., Kagiya T., Otsu K., Ishizeki K., Harada H. Establishment of Hertwig's epithelial root sheath cell line from cells involved in epithelial-mesenchymal transition. Biochem Biophys Res Commun. 2011;404:308–312. doi: 10.1016/j.bbrc.2010.11.112. [DOI] [PubMed] [Google Scholar]
- 66.Webb P.P., Moxham B.J., Benjamin M., Ralphs J.R. Changing expression of intermediate filaments in fibroblasts and cementoblasts of the developing periodontal ligament of the rat molar teeth. J Anat. 1996;188:529–539. [PMC free article] [PubMed] [Google Scholar]
- 67.Lézot F., Davideau J.-L., Thomas B., Sharpe P., Forest N. Epithelial Dlx-2 homeogene expression and cementogenesis. J Histochem Cytochem. 2000;48:277–283. doi: 10.1177/002215540004800213. [DOI] [PubMed] [Google Scholar]
- 68.Huang X., Bringas P., Jr., Slavkin H.C., Chai Y. Fate of HERS during tooth root development. Dev Biol. 2009;334:22–30. doi: 10.1016/j.ydbio.2009.06.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Hirata A., Nakamura H. Localization of perlecan and heparanase in Hertwig's epithelial root sheath during root formation in mouse molars. J Histochem Cytochem. 2006;54:1105–1113. doi: 10.1369/jhc.5A6883.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Yamamoto T., Domon T., Takahashi S., Anjuman K.A.Y., Fukushima C., Wakita M. Mineralization process during acellular cementogenesis in rat molars: a histochemical and immunohistochemical study using fresh-frozen sections. Histochem Cell Biol. 2007;127:303–311. doi: 10.1007/s00418-006-0242-x. [DOI] [PubMed] [Google Scholar]
- 71.Yamamoto T., Takahashi S. Hertwig's epithelial root sheath cells do not transform into cementoblasts in rat molar cementogenesis. Ann Anat. 2009;191:547–555. doi: 10.1016/j.aanat.2009.07.004. [DOI] [PubMed] [Google Scholar]
- 72.Kaneko H., Hashimoto S., Enokiya Y., Ogiuchi H., Shimono M. Cell proliferation and death of Hertwig's epithelial root sheath in the rat. Cell Tissue Res. 1999;298:95–103. doi: 10.1007/s004419900061. [DOI] [PubMed] [Google Scholar]
- 73.Cerri P.S., Freymüller E., Katchburian E. Apoptosis in the early developing periodontium of rat molars. Anat Rec. 2000;255:136–144. doi: 10.1002/(SICI)1097-0185(20000201)258:2<136::AID-AR3>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]