

Abstract
Purpose:
Recent studies have identified multilocus imprinting disturbances (MLIDs) in a subset of patients with imprinting diseases (IDs) caused by epimutations. We examined MLIDs in patients with Temple syndrome (TS14) and Kagami-Ogata syndrome (KOS14).
Methods:
We studied four TS14 patients (patients 1–4) and five KOS14 patients (patients 5–9) with epimutations. We performed HumanMethylation450 BeadChip (HM450k) analysis for 43 differentially methylated regions (DMRs) (753 CpG sites) and pyrosequencing for 12 DMRs (62 CpG sites) using leukocyte genomic DNA (Leu-gDNA) of patients 1–9, and performed HM450k analysis for 43 DMRs (a slightly different set of 753 CpG sites) using buccal cell gDNA (Buc-gDNA) of patients 1, 3, and 4. We also performed mutation analysis for six causative and candidate genes for MLIDs and quantitative expression analysis using immortalized lymphocytes in MLID-positive patients.
Results:
Methylation analysis showed hypermethylated ZDBF2-DMR and ZNF597/NAA60-DMR, hypomethylated ZNF597-DMR in both Leu-gDNA and Buc-gDNA, and hypomethylated PPIEL-DMR in Buc-gDNA of patient 1, and hypermethylated GNAS-A/B-DMR in Leu-gDNA of patient 3. No mutations were detected in the six genes for MLIDs. Expression patterns of ZDBF2, ZNF597, and GNAS-A/B were consistent with the identified MLIDs.
Conclusion:
This study indicates the presence of MLIDs in TS14 patients but not in KOS14 patients.
Genet Med 19 4, 476–482.
Keywords: epimutation, imprinting, Kagami-Ogata syndrome, multilocus imprinting disturbance, Temple syndrome
Introduction
The human chromosome 14q32.2 imprinted region harbors paternally and maternally expressed genes, together with the germ line–derived primary DLK1-MEG3 intergenic differentially methylated region (IG-DMR) and the postfertilization-derived secondary MEG3-DMR that function as the imprinting control centers in the placenta and the body, respectively.1 Consistent with this, maternal uniparental disomy 14 (UPD(14)mat) and loss-of-methylation (LOM)-type epimutations and microdeletions affecting the paternally derived imprinted region lead to Temple syndrome (TS14, OMIM 616222), which is associated with pre- and postnatal growth deficiency, hypotonia, small hands, and precocious puberty.2 Similarly, paternal uniparental disomy 14 (UPD(14)pat) and gain-of-methylation (GOM)-type epimutations and microdeletions involving the maternally inherited imprinted region result in Kagami-Ogata syndrome (KOS14, OMIM 608149), which is characterized by unique facial features; a small, bell-shaped thorax with a coat-hanger appearance of the ribs; abdominal-wall defects; placentomegaly; and polyhydramnios.1
Recent studies have identified multilocus imprinting disturbances (MLIDs) in a subset of patients with imprinting diseases (IDs) caused by epimutations.3 Indeed, although the frequency of MLIDs is variable among studies, MLIDs have been detected in approximately 50% of patients with transient neonatal diabetes mellitus caused by LOM of the PLAGL1-DMR, approximately 12% of patients with Beckwith-Wiedemann syndrome caused by LOM of the Kv-DMR or GOM of the H19-DMR, 8–10% of patients with Silver-Russell syndrome (SRS) caused by LOM of the H19-DMR, and 8–10% of patients with pseudohypoparathyroidism type 1b caused by LOM of the GNAS-A/B-DMR, as well as in a single patient with Angelman syndrome caused by LOM of the SNRPN-DMR.3 In addition, although ID-related DMRs have usually been examined to detect MLIDs by conventional methylation analyses including pyrosequencing, genome-wide MLID analysis has become possible by array-based methods.4 The underlying factors for the MLIDs remain to be clarified in most patients, but the causative genes for MLIDs (ZFP57, NLRP2, and NLRP5), as well as candidate genes for MLIDs (NLRP7, KHDC3L, and TRIM28), have been identified in a few patients with MLIDs.4,5
MLIDs have been poorly studied in patients with TS14 and KOS14, primarily because of their rarity. To our knowledge, TS14 has been identified in 58 patients, including 10 patients with epimutations of the IG-DMR and the MEG3-DMR, and KOS14 has been identified in 57 patients, including 7 patients with epimutations of the two DMRs.1,2,6,7,8,9 Thus, we examined MLIDs in TS14 and KOS14 caused by epimutations.
Materials and Methods
Ethical approval
This study was approved by the Institutional Review Board Committee at the National Center for Child Health and Development and was performed after obtaining written informed consent to publish the clinical and molecular information.
Patients
This study consisted of four TS14 patients with LOM-type epimutations (patients 1–4) and five KOS14 patients with GOM-type epimutations (patients 5–9) (Table 1). LOMs/GOMs were confirmed by the conventional methylation analysis for IG-DMR and MEG3-DMR using leukocyte genomic DNA (Leu-gDNA). UPD(14)mat/pat was ruled out by microsatellite analysis using parental Leu-gDNA, and microdeletions were excluded by fluorescence in situ hybridization analysis of the two DMRs and a custom-built array comparative genomic hybridization analysis for the 14q32.2 imprinted region.1,10
Table 1. Methylation indices (%) for CpG dinucleotides determined by pyrosequencing analysis for leukocyte genomic DNA samples.
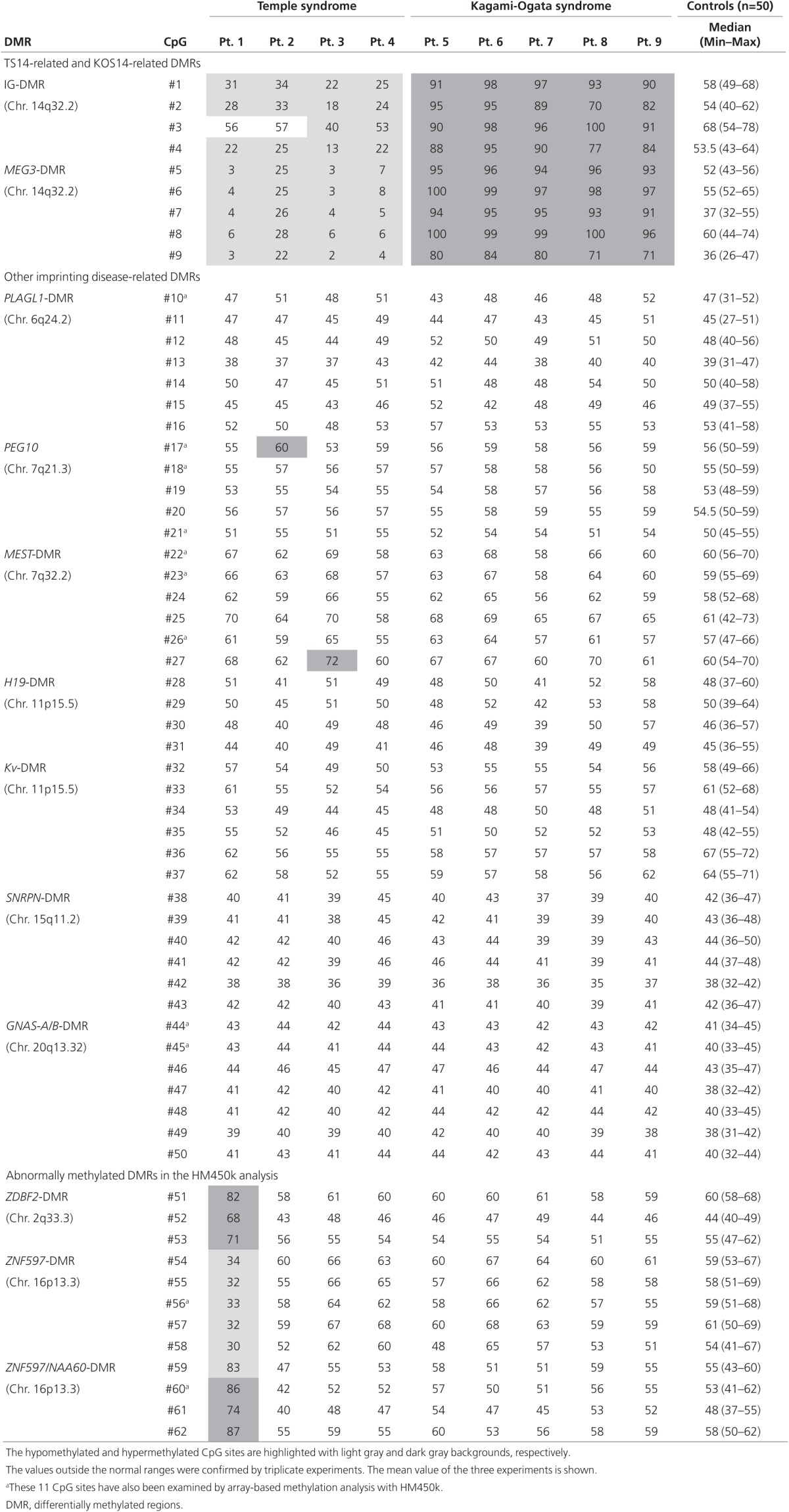
Detailed phenotypes are shown in Supplementary Tables S1 and S2 online. In brief, patient 1 exhibited Prader-Willi syndrome (PWS)-like phenotype in the absence of GOM at the SNRPN-DMR, and patients 2–4 manifested SRS-compatible phenotype in the absence of LOM at the H19-DMR and UPD(7)mat. Prenatal growth failure was absent from patient 1 and present in patients 2–4, whereas postnatal growth failure was present in patients 1–4. KOS14 patients were identified by the typical phenotype.
Molecular studies
We conducted methylation, mutation, and expression analyses. Detailed methods are described in the Supplementary Methods online. In brief, methylation analysis was performed by array-based HumanMethylation450 BeadChip (HM450k) (Illumina, San Diego, CA) analysis for 753 CpG sites on 43 DMRs using Leu-gDNA of patients 1–9 and for a slightly different set of 753 CpG sites on 43 DMRs using epithelial buccal cell gDNA (Buc-gDNA) of patients 1, 3, and 4 (Supplementary Table S3 online), and by pyrosequencing analysis for 50 CpG sites on nine ID-related DMRs and for 12 CpG sites on three DMRs that were assessed to be abnormally methylated by the HM450k analysis (Table 1) using Leu-gDNA of patients 1–9 (for primer information, see Supplementary Table S4 online). Mutation analysis was performed for the causative and candidate genes for MLIDs (ZFP57, NLRP2, NLRP7, KHDC3L, NLRP5, and TRIM28)4,5 by whole-exome sequencing using Leu-gDNA of MLID-positive patients and their mothers. Expression analysis was performed by quantitative real-time polymerase chain reaction for imprinted genes regulated by the abnormally methylated DMRs using mRNA samples extracted from immortalized lymphocytes derived from MLID-positive patients. We utilized the nomenclature for DMRs and imprinted genes proposed by the European Network of Human Congenital Imprinting Disorders (http://www.imprinting-disorders.eu/).
Results
Methylation analysis
MLID was identified in patients 1 and 3 with TS14 (Figure 1a, Supplementary Table S3 online, and Table 1) (actual methylation levels of each patient and the average ± 3 SD values of control subjects obtained by HM450k analysis are shown in Supplementary Tables S5 and S6 online). In patient 1, GOM at ZDBF2-DMR, LOM at ZNF597-DMR, and GOM at ZNF597/NAA60-DMR were identified by HM450k analysis for Leu-gDNA and Buc-gDNA, with more obvious aberrant methylation patterns in Buc-gDNA than in Leu-gDNA, and by pyrosequencing analysis for Leu-gDNA. In addition, LOM at PPIEL-DMR was found only in Buc-gDNA by HM450k analysis. In patient 3, GOM at GNAS-A/B-DMR was detected only in Leu-gDNA by HM450k analysis, but not by pyrosequencing analysis.
Figure 1.

Methylation and expression analyses. (a) Heat map indicating the Δβ values for 753 probes (CpG sites) examined by HumanMethylation450 BeadChip using Leu-gDNA (Leu.) and Buc-gDNA (Buc.). The methylation levels of CpG sites are classified into nine categories based on Δβ values. The open rectangles indicate CpG sites with no signal intensities. A single row indicates a single probe (CpG site). Green rectangles represent 11 CpG sites that were also examined by pyrosequencing (Table 1). (b) Quantitative real-time polymerase chain reaction analysis using immortalized lymphocytes. Shown are relative mRNA expression levels for ZDBF2, ZNF597, and GNAS-A/B against GAPDH (mean ± SE). The expression studies were performed three times for each sample. Statistical significance between patients and controls was examined by the Mann-Whitney U-test.
No other MLID was identified. Although pyrosequencing analysis showed slight GOMs at the #17 CpG site within PEG10-DMR in patient 2 and the #27 CpG site within MEST-DMR in patient 3 (Table 1), other CpG sites within these DMRs were normal. Similarly, although HM450k analysis revealed several abnormally methylated CpG sites (Figure 1a, Supplementary Table S3 online), they accounted for <20% of CpGs within corresponding DMRs.
Of the 11 CpG sites examined by HM450k and pyrosequencing analyses, discordant methylation data were obtained for the #17 CpG site in patient 2 and for the #44 and #45 CpG sites in patient 3. Furthermore, HM450k analysis indicated (i) more severe LOMs at MEG3-DMR of Leu-gDNA in patient 1 than in patients 2–4; (ii) more severe LOMs at MEG3-DMR in Buc-gDNA than in Leu-gDNA of patients 3 and 4; (iii) invariable GOMs and LOMs at MEG8-DMR in TS14 and KOS14 patients, respectively; (iv) normal methylation patterns at H19-DMR and SNRPN-DMR in Buc-gDNA of patients 1, 3, and 4, as well as in Leu-gDNA of patients 1–9; and (v) hypermethylated GPR1-AS-DMR in Leu-gDNA of patients 1–9 and control subjects (Supplementary Table S5 online).
Mutation analysis
No pathogenic mutation was identified in the six genes of MLID-positive patients 1 and 3 and their mothers.
Expression analysis
Expression analysis revealed significantly increased ZDBF2 expression and decreased ZNF597 expressions in patient 1 and significantly decreased GNAS-A/B expression in patient 3 (Figure 1b).
Discussion
We performed genome-wide MLID analysis for four TS14 and five KOS14 patients with epimutations. MLIDs were found in Leu-gDNA and Buc-gDNA of patient 1 and in Leu-gDNA of patient 3 with TS14. In particular, the MLIDs identified in Leu-gDNA (the GOMs at ZDBF2-DMR and ZNF597/NAA60-DMR in patient 1 and the GOM at GNAS-A/B-DMR in patient 3) would be consistent with the expression data using immortalized lymphocytes. Indeed, under normal conditions, ZDBF2 is expressed from a paternally derived chromosome with methylated ZDBF2-DMR,11 ZNF597 is expressed from a maternally inherited chromosome with unmethylated ZNF597/NAA60-DMR,12 and GNAS-A/B is expressed from a paternally derived chromosome with unmethylated GNAS-A/B-DMR.13 By contrast, because MLID was absent from KOS14 patients, the GOM at IG-DMR and MEG3-DMR may occur as an isolated epimutation.
Several findings are notable with regard to the identified MLIDs. First, the degrees of GOMs at ZBDF2-DMR and ZNF597/NAA60-DMR in patient 1 differed between Leu-gDNA and Buc-gDNA, and the LOM at PPIEL-DMR in patient 1 and GOM at GNAS-A/B-DMR in patient 3 were found only in Buc-gDNA and Leu-gDNA, respectively. The extents of LOMs at MEG3-DMR also differed between Buc-gDNA and Leu-gDNA of patients 3 and 4. These findings imply the occurrence of somatic mosaicism caused by defective maintenance of methylation patterns at these DMRs during development.14 This notion assumes that the associated GOM-type dominant MLIDs in patients 1 and 3 with TS14 have taken place independently of the LOM-type epimutations at MEG3-DMR. Consistent with this, no pathologic mutation was identified in the six genes for MLIDs. However, the possibility of a mutation in a hitherto-unknown gene for MLIDs has not been excluded formally, and further studies are required to elucidate the underlying factors for the development of MLIDs. Second, GOM at GNAS-A/B-DMR in Leu-gDNA of patient 3 was identified by HM450k analysis but not by pyrosequencing analysis, with discordant results for the same #44 and #45 CpG sites (Table 1). This would be due primarily to the differences in the control subjects and the normal ranges (average ± 3 SD range for HM450k analysis as used in the previous study,4 and the minimum–maximum range for pyrosequencing analysis as used in our previous study10) between the two analyses. Age-, sex-, and race-specific normal ranges would need to be established to permit data comparison among different studies and different methods. Finally, MLIDs in patient 1 were detected in ID-unrelated nonclassic DMRs. Recent array-based studies have also detected MLIDs in such DMRs.5 Thus, application of array-based analysis in epimutation-positive patients, including those who have been studied by conventional methylation analyses only, would facilitate the identification of MLIDs affecting such DMRs.
Several additional matters are also worth pointing out. First, MEG8-DMR exhibited methylation patterns opposite to those of IG-DMR and MEG3-DMR, as previously reported.15,16 This exemplifies the complexity of the methylation patterns of DMRs within a given imprinted region. Second, GPR1-AS-DMR was hypermethylated in Leu-gDNA of all patients and control subjects. Thus, although GPR1-AS-DMR remains a maternally methylated DMR and appears to control the methylation pattern of the paternally methylated secondary ZDBF2-DMR in the placenta,11,15,17 it is unlikely that such an interaction underlies in the development of GOM at ZDBF2-DMR in Leu-gDNA and Buc-gDNA of patient 1. Third, LOM at ZNF597-DMR co-existed with GOM at ZNF597/NAA60-DMR in Leu-gDNA of patient 1. This is consistent with the maternally methylated ZNF597-DMR functioning as an upstream regulator for the methylation pattern of the secondary paternally methylated ZNF597/NAA60-DMR.12,15
Clinical features of patients 1 and 3 appear to be within the phenotypic spectrum of TS14, despite the presence of MLIDs. Indeed, TS14 patients have been identified primarily by PWS- or SRS-like phenotypes.2 However, it is known that UPD(16)mat with GOM of ZNF597-DMR and excessive ZNF597 expression is associated with pre- and postnatal growth failure,18 as is UPD(20)mat with GOM at GNAS-A/B-DMR and no GNAS-A/B expression.19 Thus, it may be possible that the LOM of ZNF597-DMR is relevant to the normal fetal growth in patient 1, and that the GOM at GNAS-A/B -DMR is involved in the growth failure in patient 3. In addition, because ZDBF2 is expressed in the fetal mouse,11 the GOM of ZDBF2-DMR also may have played a certain role in the growth pattern of patient 1. Furthermore, although it remains unknown why TS14 patients show PWS- or SRS-like features, more severe LOMs at MEG3-DMR in patient 1 than in patients 2–4 might be involved in the phenotypic difference. In addition, although the methylation patterns of the PWS- and SRS-related DMRs were normal in the examined tissues of patients 1–4, there might be abnormally methylated DMR(s) hidden in the critical tissues.20
In summary, we identified MLIDs in two of four patients with TS14 and none of five patients with KOS14 with epimutations. Further studies will permit a better assessment of the frequency of MLIDs and the phenotypic consequences of MLIDs in patients with IDs.
Disclosure
The authors declare no conflict of interest.
Acknowledgments
We are grateful to all of the patients and their parents for their cooperation. We thank Toshiro Nagai, Rika Kosaki, Seiji Mizuno, Yasuhiro Naiki, Kenji Kurosawa, Goro Sasaki, Kouji Masumoto, Yumiko Komatsu, Akiko Yamamoto, and Takashi Imamura for providing us with clinical data and materials for molecular studies. We also thank Hiromi Kamura, Tamae Tanji, and Akihiro Umezawa for their support regarding molecular analyses. This work was supported by grants from the Japan Society for the Promotion of Science (JSPS) (A-25253023 and 15K15096), the National Center for Child Health and Development (25-10 and 28-6), the Japan Agency for Medical Research and Development (AMED) (16ek0109067h0003, 16ek0109030h0003, and 16ek0109141h0002), the Takeda Science Foundation, and the Daiichi Sankyo Foundation.
Supplementary Material
References
- Ogata T, Kagami M. Kagami-Ogata syndrome: a clinically recognizable upd(14)pat and related disorder affecting the chromosome 14q32.2 imprinted region. J Hum Genet 2016;61:87–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ioannides Y, Lokulo-Sodipe K, Mackay DJ, Davies JH, Temple IK. Temple syndrome: improving the recognition of an underdiagnosed chromosome 14 imprinting disorder: an analysis of 51 published cases. J Med Genet 2014;51:495–501. [DOI] [PubMed] [Google Scholar]
- Mackay DJ, Eggermann T, Buiting K, et al. Multilocus methylation defects in imprinting disorders. Biomol Concepts 2015;6:47–57. [DOI] [PubMed] [Google Scholar]
- Court F, Martin-Trujillo A, Romanelli V, et al. Genome-wide allelic methylation analysis reveals disease-specific susceptibility to multiple methylation defects in imprinting syndromes. Hum Mutat 2013;34:595–602. [DOI] [PubMed] [Google Scholar]
- Docherty LE, Rezwan FI, Poole RL, et al. Mutations in NLRP5 are associated with reproductive wastage and multilocus imprinting disorders in humans. Nat Commun 2015;6:8086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenfeld JA, Fox JE, Descartes M, et al. Clinical features associated with copy number variations of the 14q32 imprinted gene cluster. Am J Med Genet A 2015;167A:345–353. [DOI] [PubMed] [Google Scholar]
- Corsello G, Salzano E, Vecchio D, et al. Paternal uniparental disomy chromosome 14-like syndrome due a maternal de novo 160 kb deletion at the 14q32.2 region not encompassing the IG- and the MEG3-DMRs: Patient report and genotype-phenotype correlation. Am J Med Genet A 2015;167A:3130–3138. [DOI] [PubMed] [Google Scholar]
- Briggs TA, Lokulo-Sodipe K, Chandler KE, Mackay DJ, Temple IK. Temple syndrome as a result of isolated hypomethylation of the 14q32 imprinted DLK1/MEG3 region. Am J Med Genet A 2016;170A:170–175. [DOI] [PubMed] [Google Scholar]
- Sachwitz J, Strobl-Wildemann G, Fekete G, et al. Examinations of maternal uniparental disomy and epimutations for chromosomes 6, 14, 16 and 20 in Silver-Russell syndrome-like phenotypes. BMC Med Genet 2016;17:20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kagami M, Mizuno S, Matsubara K, et al. Epimutations of the IG-DMR and the MEG3-DMR at the 14q32.2 imprinted region in two patients with Silver-Russell Syndrome-compatible phenotype. Eur J Hum Genet 2015;23:1062–1067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi H, Yamada K, Morita S, et al. Identification of the mouse paternally expressed imprinted gene Zdbf2 on chromosome 1 and its imprinted human homolog ZDBF2 on chromosome 2. Genomics 2009;93:461–472. [DOI] [PubMed] [Google Scholar]
- Nakabayashi K, Trujillo AM, Tayama C, et al. Methylation screening of reciprocal genome-wide UPDs identifies novel human-specific imprinted genes. Hum Mol Genet 2011;20:3188–3197. [DOI] [PubMed] [Google Scholar]
- Maupetit-Méhouas S, Azzi S, Steunou V, et al. Simultaneous hyper- and hypomethylation at imprinted loci in a subset of patients with GNAS epimutations underlies a complex and different mechanism of multilocus methylation defect in pseudohypoparathyroidism type 1b. Hum Mutat 2013;34:1172–1180. [DOI] [PubMed] [Google Scholar]
- Azzi S, Blaise A, Steunou V, et al. Complex tissue-specific epigenotypes in Russell-Silver Syndrome associated with 11p15 ICR1 hypomethylation. Hum Mutat 2014;35:1211–1220. [DOI] [PubMed] [Google Scholar]
- Court F, Tayama C, Romanelli V, et al. Genome-wide parent-of-origin DNA methylation analysis reveals the intricacies of human imprinting and suggests a germline methylation-independent mechanism of establishment. Genome Res 2014;24:554–569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bens S, Kolarova J, Gillessen-Kaesbach G, et al. The differentially methylated region of MEG8 is hypermethylated in patients with Temple syndrome. Epigenomics 2015;7:1089–1097. [DOI] [PubMed] [Google Scholar]
- Sanchez-Delgado M, Martin-Trujillo A, Tayama C, et al. Absence of maternal methylation in biparental hydatidiform moles from women with NLRP7 maternal-effect mutations reveals widespread placenta-specific imprinting. PLoS Genet 2015;11:e1005644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azzi S, Salem J, Thibaud N, et al. A prospective study validating a clinical scoring system and demonstrating phenotypical-genotypical correlations in Silver-Russell syndrome. J Med Genet 2015;52:446–453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mulchandani S, Bhoj EJ, Luo M, et al. Maternal uniparental disomy of chromosome 20: a novel imprinting disorder of growth failure. Genet Med 2016;18:309–315. [DOI] [PubMed] [Google Scholar]
- Alders M, Maas SM, Kadouch DJ, et al. Methylation analysis in tongue tissue of BWS patients identifies the (EPI)genetic cause in 3 patients with normal methylation levels in blood. Eur J Med Genet 2014;57:293–297. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.