

Immune disturbances are a profound feature of chronic lymphocytic leukemia (CLL). From an early stage, immune defects contribute to infectious complications and autoimmune cytopenias.1 Moreover, the progressively immunosuppressive environment in CLL influences disease advancement and treatment efficacy.2 The mechanisms underlying these clinical phenomena encompass both the innate and adaptive immune system. At the crossroad of innate and adaptive immunity, natural killer (NK) cells exhibit diminished cytotoxicity, despite an increase in absolute number.3,4 NK cells are now recognized to be part of a larger group of innate lymphocytes, collectively referred to as innate lymphoid cells (ILCs).5 ILCs play a key role in the initial immune response, tumorsurveillance and tumorigenesis.6–8 The role of ILCs in CLL, however, has not yet been clarified. Hence, we decided to study the frequency and functionality of ILCs in CLL patients.
ILCs are a heterogeneous group of cells with lymphoid cell morphology without rearranged antigen receptors and, by definition, do not express lineage markers (Lin−).5 In addition to NK cells, the ILC family consists of three non-cytotoxic populations based on expression of transcription factors and cytokines, mirroring T helper cell subsets. ILC1s express T-bet and IFN-γ, ILC2s express GATA3 and RORα and secrete IL-5 and IL-13, whilst ILC3s express RORγt, IL-22 and IL-17, and are subdivided based on the expression of natural cytotoxicity receptor NKp44.5
Early evidence shows that ILCs can play a role in tumor development, chiefly by producing tumor-promoting cytokines and by supporting an immunosuppressive environment. In acute myeloid leukemia (AML), at diagnosis, ILCs are significantly dysregulated in terms of frequency, subtype composition and function.9,10 A hallmark feature of lymphoproliferative disorders is the intensive crosstalk between malignant lymphoid cells and other immune cells. To our knowledge, the role of ILCs in lymphoproliferative disorders, and particularly in the disturbed CLL immune system, has not been characterized. We studied ILCs in CLL patients to establish their frequency, distribution and functionality.
Peripheral blood samples from 21 CLL patients and 8 age-matched healthy controls (HCs) were analyzed (Online Supplementary Table). All patients were treatment-naïve and the median absolute leukocyte count (ALC) was 47.6×109 cells/L (17.5–179.3). The study was approved by the Medical Ethical Committee of the Academic Medical Center in Amsterdam, and all patients signed informed consent in accordance with the Declaration of Helsinki.
Total ILCs were defined as Lin−(CD1a−CD14−CD19−CD34−CD123−TCRαβ−TCRγδ-BDCA2-FcεR1−CD94−) CD127+CD161+ lymphocytes11 (Online Supplementary Figure S1A). Group 1 ILCs were defined as CRTH2−CD117−, group 2 ILCs as CRTH2+ and group 3 ILCs as CRTH2−CD117+, subdivided into NKp44+ and NKp44−. Cells were acquired on an LSRFortessa (BD Biosciences), and analyzed with FlowJo v10 for Mac (TreeStar). Statistical analyses were performed with t-tests, Pearson correlation analyses and Kruskal-Wallis test with Dunn’s multiple comparison test.
Total ILC counts in the peripheral blood were significantly increased in CLL patients in comparison to age-matched HCs (Figure 1A). Moreover, the ILC count correlated with the ALC in CLL patients, suggesting a rise in ILCs with disease progression (r2=0.477, P=0.023; Figure 1B). Accordingly, the ILC count correlated inversely with time to first treatment (r2=0,259, P=0.044; Figure 1C). The activation status of ILCs, as measured by CD69 expression, was similar in CLL patients and HCs (Online Supplementary Figure S1B). The absolute counts of both ILC1s and NKp44− ILC3s were significantly elevated in comparison to HCs (Figure 1D). NKp44+ ILC3s were virtually absent in the peripheral blood of both CLL patients and HCs. However, the proportion of each ILC subgroup in CLL patients resembled their respective fractions in the HCs (Figure 1E). ILC counts did not differ between M-CLL and U-CLL patients (Figure 1F). Similarly, ILC subtype distribution of M-CLL patients was equivalent to that of U-CLL (Online Supplementary Figure S1C).
Figure 1.
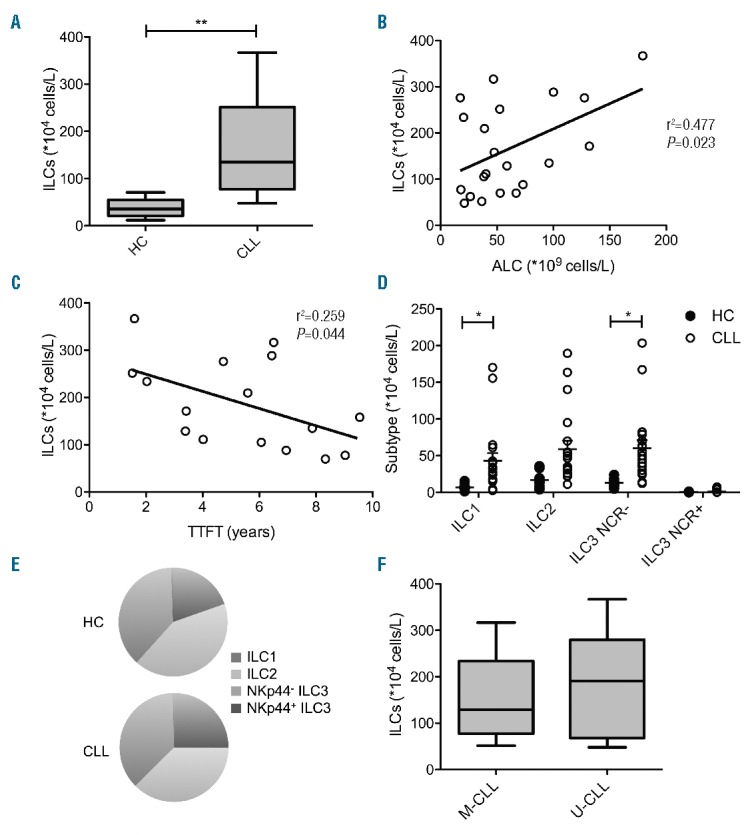
Frequency and subtype distribution of ILCs in untreated CLL patients and age-matched HCs. Cells were stained with FITC-conjugated antibodies against lineage markers (anti-CD1a, anti-CD11c, anti-CD14, anti-CD19, anti-CD34, anti-CD94, anti-CD123, anti-FcER1α), anti-CD69 APC Cy7 (Biolegend), anti-TCRαβ FITC, anti-TCRγδ FITC, anti-CRTH2 AF647, anti-CD3 V500 (BD Biosciences), anti-NKp44 PE, anti-CD117 PE-Cy5.5 and anti-CD127 PE-Cy7 (Beckman Coulter). (A) Absolute total ILC counts in HCs (N=8) and CLL patients (n=21). (B) Correlation between absolute ILC counts and ALC in CLL patients. (C) Correlation between absolute ILC counts and time to first treatment (TTFT) in CLL patients. Absolute counts (D) and percentage (E) of ILC1s, ILC2s, NKp44−ILC3s and NKp44+ ILC3s. (F) Absolute counts of ILCs in M-CLL (n=12) and U-CLL (n=9). Data are presented as mean with standard error of the mean (SEM). Statistically significant differences are presented as *P<0.05; **P<0.01.
Subsequently, we analyzed and compared cytokine production as a marker for ILC functionality. PBMCs from 6 CLL patients and 6 age-matched HCs, with comparable ILC subgroup distribution, were depleted from T and B cells using anti-CD3 and anti-CD19 magnetic microbeads (Miltenyi Biotec). Cells were stimulated with PMA (10 ng/mL; Sigma-Aldrich) and ionomycin (0.5 μg/mL; Sigma-Aldrich) for 4 hours in the presence of Brefeldin A (10 μg/mL; Invitrogen) and stained extracellularly to identify ILCs (Online Supplementary Figure S1D). Subsequently, cells were stained intracellularly for production of IFN-γ, TNF-α (group 1 ILCs), IL-13 (group 2 ILCs) and IL-22 (group 3 ILCs).
HC derived ILC1 cells produced robust levels of TNF-α. In contrast, TNF-α production in CLL patients was significantly reduced (Figure 2A), while IFN-γ production was comparable between CLL patients and HCs. The percentage of TNF-α producing ILCs correlated positively with the percentage of group 1 ILCs as measured in unstimulated samples (r2=0.736, P=0.029; Online Supplementary Figure S1E), as did IFN-γ producing ILCs (r2=0.669, P=0.047; Online Supplementary Figure S1F). In contrast, no differences were observed in type 2 (IL-13) and type 3 (IL-22) cytokine production (Figure 2A). These findings show strong induction of TNF-α in directly stimulated ILCs in healthy controls but defective TNF-α production in CLL ILCs.
Figure 2.
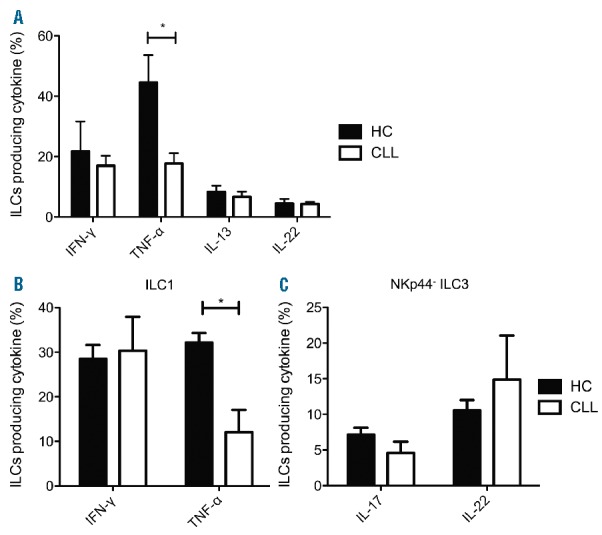
Cytokine production by ILCs in untreated CLL patients. (A) PBMCs depleted from T and B lymphocytes were stimulated with PMA/ionomycin for 4 hours and stained with anti-CD3 V500, FITC-conjugated anti-lineage, anti-CD127 PE-Cy7 and anti-CD161 BV711 CD161 to identify ILCs. Subsequently cells were stained with anti-IFN-γ BUV395, anti-TNF-α AF700, IL-13 APC (BD Biosciences) and IL-22 PE (eBioscience). Percentage of total ILCs of CLL patients (n=6) and age-matched HCs (n=6) producing IFN-γ, TNF-α, IL-13 and IL-22. (B–C) Cytokine production by ILC subgroups after ex vivo culture. Group 1 and NKp44− group 3 ILCs from CLL patients (n=4) and age-matched HCs (n=3) were FACS-sorted and cultured for 2 weeks. Subsequently, ILC1s were resorted, stimulated with PMA/ionomycin, IL-12 and IL-18 and then stained intracellularly with anti-IFN-γ BUV395 and anti-TNF-α AF700 (BD Biosciences). NKp44− ILC3s were stimulated with PMA/ionomycin, IL-1β and IL-23 and then stained intracellularly with anti-IL-17A AF700 and IL-22 PE (BD Biosciences). (B) Percentage of ILC1s producing IFN-γ and TNF-α after culture. (C) Percentage of NKp44− ILC3s producing IL-17A and IL-22 after culture. Data are presented as mean with standard error of the mean (SEM). Statistically significant differences are presented as *P<0.05.
Next we investigated whether this phenomenon remained present after in vitro activation of ILC subgroups. Using a BD Facs Aria cell sorter (BD Biosciences), ILC1s and NKp44− ILC3s were sorted from CD3/CD19-depleted PBMCs from 4 CLL patients and 3 age-matched HCs. Expansion of ILC2s did not succeed due to insufficient yield of ILC2 sorting. ILC1s and NKp44−ILC3s were cultured in Yssel’s medium supplemented with 1% human serum in the presence of irradiated HC PBMCs and JY-cells, phytohemagglutinin (PHA; 1 μg/mL) and IL-2 (100 U/mL) for 2 weeks and resorted afterwards, after which the purity for each sample was ≥95%. ILC1s were stimulated with IL-12, IL-18 (50 ng/ml, R&D Systems) and PMA/ionomycin as described above and subsequently stained for TNF-α and IFN-γ. NKp44− ILC3s were stimulated with IL-1β, IL-23 (50 ng/ml, R&D Systems) and PMA/ionomycin and subsequently stained for IL-17A and IL-22.
Again, ILC1s from CLL patients appeared to be functionally impaired when compared to HCs. In parallel with direct cytokine measurements in the total ILC pool, cultured ILC1s from CLL patients produced less TNF-α (Figure 2B). IFN-γ production by cultured ILC1s was unaffected. In contrast with ILC1s, production of IL-17A and IL-22 (Figure 2C) of NKp44-ILC3s was not affected upon culture.
Subsequently, we investigated whether ILC cytokine production was affected by CLL cells. PBMCs from HCs (n=3) were depleted from monocytes, T and B cells using anti-CD4, anti-CD8, anti-CD14 and anti-CD19 magnetic microbeads (Miltenyi Biotec). The enriched ILCs were cultured overnight with CLL cells (≥95% CD5+CD19+) obtained from 1 untreated donor in a 1:10 ratio or medium alone. During the final 4 hours, cells were stimulated with either PMA (10 ng/mL) and ionomycin (0.5 μg/mL) or medium in the presence of Brefeldin A (10 μg/mL). Cells were then stained extracellularly to identify ILCs and intracellularly for IFN-γ, TNF-α, IL-13 and IL-22.
CLL cells proved capable of stimulating HC ILCs, as measured by production of IFN-γ and TNF-α (Figure 3A). Stimulation with PMA/ionomycin enhanced IFN-γ and TNF-α production, although less when cells were cocultured with CLL cells (Figure 3B). IL-13 and IL-22 production by ILCs was not altered in the presence of CLL cells.
Figure 3.
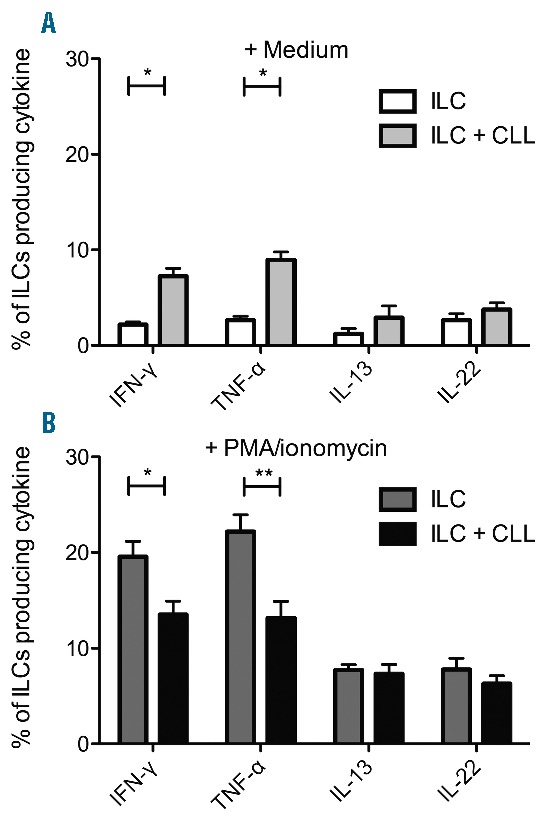
The effect of CLL cells on ILC cytokine production. PBMCs from HCs (n=3) depleted from monocytes, T and B cells were cocultured with CLL cells in a 1:10 ratio or medium alone overnight. (A) Brefeldin A was added during the final 4 hours of culture. Percentage of ILCs producing cytokines after coculture. (B) Cells were stimulated with PMA/ionomycin in the presence of Brefeldin A during the final 4 hours of culture. The percentage of ILCs producing cytokines after coculture.
In summary, we show for the first time that peripheral blood ILCs are altered in terms of number and function in untreated CLL patients. The increase in ILC counts is progressively seen in patients with higher lymphocyte counts and shorter TTFT, suggesting a disease-related process. Moreover, our findings reveal distorted ILC1 function as TNF-α production is disturbed, which persists upon ex vivo expansion. Interestingly, we show that CLL cells are capable of stimulating ILCs upon coculture, inducing IFN-γ and TNF-α production. CLL cells have been reported to produce IL-12,12,13 a cytokine to which ILC1 in particular are responsive. When ILCs are stimulated with PMA/ionomycin, however, the presence of CLL cells averted further enhancement of IFN-γ and TNF-α production. Thus, ILC1s that have been in the presence of CLL cells, either in vivo or in vitro, appear less responsive to stimulatory signals. Whether this decreased responsiveness of group 1 ILCs reflects an immunosuppressive effect of the CLL cells, or results from competition for nutrients and cytokines between ILCs and CLL cells remains to be clarified.
These observations are similar to what has been observed for NK cells in CLL patients. NK cells are expanded early on during CLL, while their numbers correlate with prognosis.4 Moreover, in CLL patients, NK cells have impaired cytolytic activity14 and cytokine production.15 Previous studies have identified both soluble and cell-cell mechanisms by which CLL cells may reduce NK cell function.
Taken together, we demonstrate that in patients with CLL, ILC counts are increased and the function of type 1 ILCs is changed, similar to what has previously been demonstrated for NK cells. These observations may represent a bystander effect or may reflect functional involvement of ILCs in CLL pathobiology. The extent to which ILCs affect disease progression and therapeutic response is yet to be identified.
Supplementary Material
Footnotes
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Forconi F, Moss P. Perturbation of the normal immune system in patients with CLL. Blood. 2015;126(5):573–581. [DOI] [PubMed] [Google Scholar]
- 2.Riches JC, Gribben JG. Understanding the immunodeficiency in chronic lymphocytic leukemia: potential clinical implications. Hematol Oncol Clin North Am. 2013;27(2):207–235. [DOI] [PubMed] [Google Scholar]
- 3.Wild J, Schmiedel BJ, Maurer A, et al. Neutralization of (NK-cell-derived) B-cell activating factor by Belimumab restores sensitivity of chronic lymphoid leukemia cells to direct and Rituximab-induced NK lysis. Leukemia. 2015;29(8):1676–1683. [DOI] [PubMed] [Google Scholar]
- 4.Huergo-Zapico L, Acebes-Huerta A, Gonzalez-Rodriguez AP, et al. Expansion of NK cells and reduction of NKG2D expression in chronic lymphocytic leukemia. Correlation with progressive disease. PLoS One. 2014;9(10):e108326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Artis D, Spits H. The biology of innate lymphoid cells. Nature. 2015;517(7534):293–301. [DOI] [PubMed] [Google Scholar]
- 6.Eisenring M, vom Berg J, Kristiansen G, Saller E, Becher B. IL-12 initiates tumor rejection via lymphoid tissue-inducer cells bearing the natural cytotoxicity receptor NKp46. Nat Immunol. 2010; 11(11):1030–1038. [DOI] [PubMed] [Google Scholar]
- 7.Kirchberger S, Royston DJ, Boulard O, et al. Innate lymphoid cells sustain colon cancer through production of interleukin-22 in a mouse model. J Exp Med. 2013;210(5):917–931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Chan IH, Jain R, Tessmer MS, et al. Interleukin-23 is sufficient to induce rapid de novo gut tumorigenesis, independent of carcinogens, through activation of innate lymphoid cells. Mucosal Immunol. 2014;7(4):842–856. [DOI] [PubMed] [Google Scholar]
- 9.Trabanelli S, Curti A, Lecciso M, et al. CD127+ innate lymphoid cells are dysregulated in treatment naive acute myeloid leukemia patients at diagnosis. Haematologica. 2015;100(7):e257–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Munneke JM, Bjorklund AT, Mjosberg JM, et al. Activated innate lymphoid cells are associated with a reduced susceptibility to graft-versus-host disease. Blood. 2014;124(5):812–821. [DOI] [PubMed] [Google Scholar]
- 11.Hazenberg MD, Spits H. Human innate lymphoid cells. Blood. 2014; 124(5):700–709. [DOI] [PubMed] [Google Scholar]
- 12.Hoogendoorn M, Wolbers JO, Smit WM, et al. Generation of B-cell chronic lymphocytic leukemia (B-CLL)-reactive T-cell lines and clones from HLA class I-matched donors using modified B-CLL cells as stimulators: implications for adoptive immunotherapy. Leukemia. 2004;18(7):1278–1287. [DOI] [PubMed] [Google Scholar]
- 13.Burgler S, Gimeno A, Parente-Ribes A, et al. Chronic lymphocytic leukemia cells express CD38 in response to Th1 cell-derived IFN-gamma by a T-bet-dependent mechanism. J Immunol. 2015; 194(2):827–835. [DOI] [PubMed] [Google Scholar]
- 14.Ziegler HW, Kay NE, Zarling JM. Deficiency of natural killer cell activity in patients with chronic lymphocytic leukemia. Int J Cancer. 1981;27(3):321–327. [DOI] [PubMed] [Google Scholar]
- 15.Eskelund CW, Nederby L, Thysen AH, et al. Interleukin-21 and rituximab enhance NK cell functionality in patients with B-cell chronic lymphocytic leukaemia. Leuk Res. 2011;35(7):914–920. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.