

Abstract
Background
A lesion-mimic mutant in rice (Oryza sativa L.), spotted leaf 5 (spl5), displays a disease-resistance-enhanced phenotype, indicating that SPL5 negatively regulates cell death and resistance responses. To understand the molecular mechanisms of SPL5 mutation-induced cell death and resistance responses, a proteomics-based approach was used to identify differentially accumulated proteins between the spl5 mutant and wild type (WT).
Results
Proteomic data from two-dimensional gel electrophoresis showed that 14 candidate proteins were significantly up- or down-regulated in the spl5 mutant compared with WT. These proteins are involved in diverse biological processes including pre-mRNA splicing, amino acid metabolism, photosynthesis, glycolysis, reactive oxygen species (ROS) metabolism, and defense responses. Two candidate proteins with a significant up-regulation in spl5 – APX7, a key ROS metabolism enzyme and Chia2a, a pathogenesis-related protein – were further analyzed by qPCR and enzyme activity assays. Consistent with the proteomic results, both transcript levels and enzyme activities of APX7 and Chia2a were significantly induced during the course of lesion formation in spl5 leaves.
Conclusions
Many functional proteins involving various metabolisms were likely to be responsible for the lesion formation of spl5 mutant. Generally, in spl5, the up-regulated proteins involve in defense response or PCD, and the down-regulated ones involve in amino acid metabolism and photosynthesis. These results may help to gain new insight into the molecular mechanism underlying spl5-induced cell death and disease resistance in plants.
Electronic supplementary material
The online version of this article (doi:10.1186/1939-8433-6-1) contains supplementary material, which is available to authorized users.
Keywords: Rice, Lesion mimic mutant, spl5, Proteomics, 2-DE
Background
In plants, one of the most common and effective defense responses to pathogen attack is the hypersensitive response (HR), which prevents further spread of pathogens to adjacent cells (Morel and Dangl 1997). Lesion mimic mutants (lmms), displaying HR-like lesions in the absence of pathogen attacks, have been identified from maize (Johal et al. 1995), Arabidopsis (Dietrich et al. 1994), barley (Wolter et al. 1993), and rice (Takahashi et al. 1999). Most lmms constitutively activate immune responses, including callose deposition, induction of Pathogenesis related (PR) genes, production of reactive oxygen species (ROS), and accumulation of phytoalexins (Staskawicz et al. 1995). Therefore, lmms are very useful genetic tools to dissect molecular mechanisms of programmed cell death (PCD) and defense responses in plants.
In rice, more than 43 lmms have been isolated, most of which display enhanced resistance to rice blast and/or bacterial blight pathogens (Takahashi et al. 1999; Yin et al. 2000; Mizobuchi et al. 2002; Jung et al. 2005; Mori et al. 2007; Wu et al. 2008; Qiao et al. 2010). So far, at least 11 lmms have been functionally characterized, including spl7 (Yamanouchi et al. 2002), spl11 (Zeng et al. 2004), Spl18 (Mori et al. 2007), spl28 (Qiao et al. 2010), sl (Fujiwara et al. 2010), ttm1 (Takahashi et al. 2007), rlin1 (Sun et al. 2011), NPR1 (Chern et al. 2005), lsd1 (Wang et al. 2005), acdr1 (Kim et al. 2009), and edr1 (Shen et al. 2011). Interestingly, these LMM genes encode different proteins with distinct functions. For example, SPL7 is a heat stress transcription factor (Yamanouchi et al. 2002); SPL11 is a E3 ubiquitin ligase (Zeng et al. 2004); SPL18, a acyltransferase (Mori et al. 2007); SPL28, a clathrin-associated adaptor protein complex 1 medium subunit 1 (Qiao et al. 2010). These findings indicate that numerous proteins with distinct functions in multiple signaling pathways and/or processes are involved to prevent inappropriate activation of PCD. Thus, lmms have helped to gain an in-depth insight into regulatory mechanisms of PCD and defense responses in plants.
Rice spotted leaf 5 (spl5) is a lmm with spontaneous HR-like lesions on its leaves, and broadly enhanced resistance to rice blast and bacterial blight pathogens (Yin et al. 2000; Mizobuchi et al. 2002). The spl5 gene was previously mapped into a 36.4-cM region on rice chromosome 7 (Iwata et al. 1978). Recently, we finely mapped and isolated spl5 by a map-based cloning, and surprisingly, it was found that the protein encoding by SPL5 gene (GeneBank accessioin: KC128660) shares a certain degree of homology with a human splicing factor 3b subunit 3 (SF3b3), one subunit of the SF3 protein complex involved in binding of U2 snRNP to the branch site in the splicing reaction of pre-mature RNAs (Chen et al. 2009, Chen et al. 2012). Therefore, it is likely that the SPL5 regulated cell death and resistance responses post-transcriptionally.
Two-dimensional gel electrophoresis (2-DE) is a most commonly used proteomics technology for monitoring global changes in protein levels in plants (Agrawal and Rakwal 2006). The comparative proteomics has been used to identify differentially expressed proteins between wild type (WT) rice and lmms (Takahashi et al. 2003; Tsunezuka et al. 2005; Jung et al. 2006; Kang et al. 2007; Kim et al. 2008). However, different defense-related proteins and metabolic enzymes were found to be differently accumulated during lesion formation in a lmm-specific manner or in different lmms. For example, two PR proteins (OsPR5 and OsPR10) and three ROS-scavenging enzyames [catalase (CAT), ascorbate peroxidae (APX), and superoxide dismutase (SOD)] were differentially expressed in the blm mutant (Jung et al. 2006); Peroxidase, thaumatin-like protein, probenazole-induced protein (PBZ1) were up-regulated in the spl1 mutant (Kim et al. 2008).
Here, we compared the protein profiles of spl5 mutant and WT by 2-DE and found that 14 proteins were differentially accumulated between WT and spl5. Among these 14 proteins, 7 were up-regulated and 7 were down-regulated, respectively, in spl5. The proteins up-regulated in spl5 are those involved in defense response or PCD, and the proteins down-regulated in spl5 involved in amino acid metabolism and photosynthesis. Interestingly, a clear correlation between levels of protein accumulation and levels of gene expression (or induction) was observed for the 7 up-regulated proteins in spl5. However, a corresponding correlation was not observed for the 7 down-regulated proteins in spl5. Together, our results may help to understand molecular mechanisms of lesion formation in spl5.
Results and discussion
2-DE analysis between WT and spl5 mutant
To compare protein expression profiles between WT and the spl5 mutant, total proteins extracted from fully developed leaves with lesions from spl5 and the corresponding leaves from WT were analyzed by 2-DE. After quantitative analysis, 14 spots with > 2-fold changes (p < 0.05) between spl5 and WT were identified (Figures 1 and 2). Compared to those in WT, seven proteins (spots 1, 5, 6, 8, 11, 13, and 14) were up-regulated and seven (spots 2, 3, 4, 7, 9, 10, and 12) were down-regulated, respectively, in spl5 mutant. Relative intensities and fold-changes of the spots differentially expressed between spl5 and WT were shown in Table 1.
Figure 1.

A silver-stained 2-DE gel of the proteins extracted from leaf blades of both WT and spl5 mutant. For IEF, 100 μg of total proteins was loaded onto pH 3-10 IPG strips (13 cm, nonlinear), and then transferred to 12.5% SDS-polyacrylamide gel for the second-dimensional electrophoresis. The protein gel was stained with silver nitrate solution. Quantitative analysis of digitized images was carried out using the Image Master software (Amersham, USA). Arrows indicate spots with more than 2-fold change in spl5 mutant compared to WT.
Figure 2.
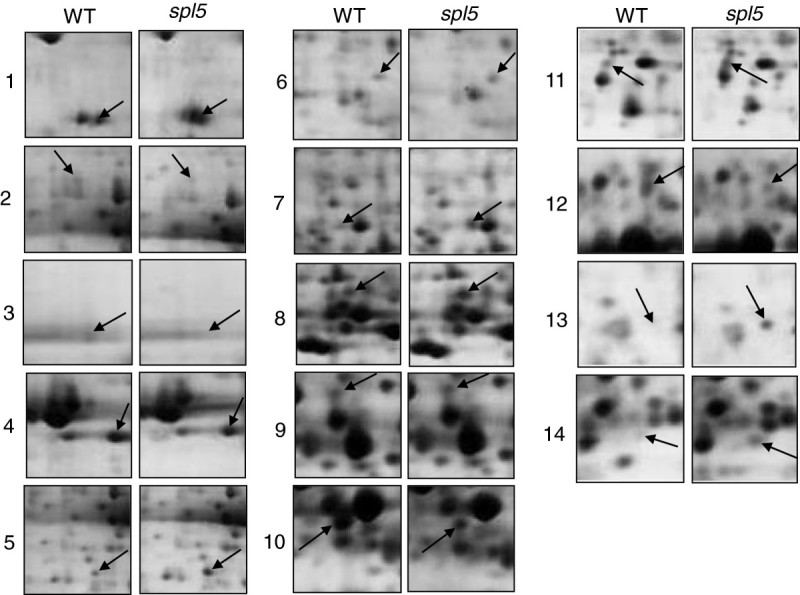
Magnified regions of 2-DE gels. Numbers at the left of images indicate protein spots showed by arrows in Figure 1 with significantly differential accumulation levels between spl5 and WT mutant in 2-DE gel analysis.
Table 1.
Identification of proteins differentially expressed between WT and spl5 mutant
| Function type | Spot id | Homologous protein | Score | Source | Accession | Coverage (%) | pI | MM (kDa) | Change fold | mRNA level |
|---|---|---|---|---|---|---|---|---|---|---|
| mRNA splicing | 2 | Thioredoxin-like protein 4B | 71 | O. sativa | gi|125576924 | 25 | 6.4 | 23.6 | −3.3 | - |
| Amino-acid metabolism | 4 | Alanine aminotransferase | 187 | O. sativa | gi|115470235 | 31 | 8 | 54.0 | −2.3 | - |
| 6 | Aspartate aminotransferase | 84 | O. sativa | gi|125541475 | 15 | 6.5 | 50.6 | 2.0 | up | |
| 12 | Cysteine synthase | 86 | O. sativa | gi|115489664 | 29 | 5.3 | 33.9 | −3.9 | - | |
| 5 | S-adenosylmethionine synthetase | 93 | O. sativa | gi|100801534 | 27 | 6.5 | 43.0 | 2.6 | up | |
| Photosynthesis | 3 | Rubisco large subunit | 130 | O. sativa | gi|115468792 | 33 | 8.5 | 48.4 | −2.7 | down |
| 9 | Rubisco activase | 74 | O. sativa | gi|1778414 | 28 | 5.4 | 48.1 | −2.0 | - | |
| 10 | Rubisco activase, chloroplast precursor | 174 | O. sativa | gi|108864712 | 44 | 5.1 | 36.7 | −3.1 | - | |
| Glycolysis | 8 | Glyceraldehyde-3-phosphate dehydrogenase | 78 | O. sativa | gi|115459078 | 24 | 7.3 | 36.9 | 3.1 | up |
| ROS metabolism | 11 | Glutathione S-transferase 14 | 88 | O. sativa | gi|46276327 | 33 | 6.5 | 30.8 | 2.1 | up |
| 13 | Ascorbate peroxidase 7 | 264 | O. sativa | gi|116310282 | 47 | 6.9 | 38.2 | 8.1 | up | |
| Defense-related | 14 | Chitinase Chia2a | 81 | O. sativa | gi|115483206 | 27 | 5.4 | 27.9 | 3.5 | up |
| Others | 1 | Retrotransposon Ty3-gypsy subclass | 68 | O. sativa | gi|77556153 | 16 | 4.7 | 29.1 | 3.0 | down |
| 7 | Nad-dependent formate dehydrogenase | 164 | O. sativa | gi|4760553 | 40 | 7.2 | 41.5 | −2.2 | - |
Spot ID refers to the spot identity as given in Figure 2; Accession, protein accession in GenBank (http://www.ncbi.nlm.nih.gov/); Coverage %, the percentage of sequence coverage; pI, experimental isoelectric points; MM, experimental molecular masses; Change fold, the expression change fold of protein level in spl5 mutant compared to WT; mRNA level, the expression change of mRNA level in spl5 mutant compared to WT.
Characterization of differentially expressed proteins in spl5 mutant
The 14 differentially expressed spots represent 14 annotated proteins, which could be categorized into seven functional classes including pre-mRNA splicing, amino acid metabolism, photosynthesis, glycolysis, ROS metabolism, defense-related, and other processes (Table 1). Interestingly, these proteins are mostly associated with cell death and defense responses in different organisms.
Pre-mRNA splicing protein
A protein component of RNA spliceosome, thioredoxin-like protein 4B (TXNL4B, Dim2, or DLP), was down-regulated in the spl5 mutant. In eukaryotes, the pre-mRNA splicing that removes intronic sequences is undertaken by the spliceosome, a macromolecular complex containing four snRNPs (U1, U2, U4/U6, and U5) and numerous auxiliary proteins (Kramer 1996). DLP functions in the cell nucleus and interacts with an U5 protein subunit of the spliceosome, and blocking DLP protein activity leads to insufficient pre-mRNA splicing (Sun et al. 2004).
Amino-acid metabolism enzymes
In differently expressed proteins, four enzymes including alanine aminotransferase (ALT; down), aspartate aminotransferase (AST; up), cysteine synthase (CSase; down), and S-adenosylmethionine synthetase (SAMS; up) (Table 1) are involved in amino acid transport and metabolism. SAMS catalyzes the biosynthesis of S-adenosylmethionine, which is a co-substrate for methylation reactions and serves as substrate for the synthesis of ethylene and the polyamines (Burstenbinder et al. 2010). Ethylene is an important hormone involved in plant responses to various stress situations (e.g. pathogen attack); and exposure of plants to ethylene can induce disease resistance (Geraats 2003). Polyamines can contribute to hydrogen peroxide (H2O2) formation in response to pathogen infections, which led to increased necrosis and resistance to disease (Marina et al. 2008; Moschou et al. 2009; Gonzalez et al. 2011). The up-regulation of SAMS was also found in the mutant cdr2 (Tsunezuka et al. 2005).
Photosynthesis proteins
Rubisco is a critical enzyme involved in photosynthetic CO2 assimilation and photorespiratory carbon oxidation. Rubisco is inactivated by ROS, and degraded during senescence and oxidative stresses (Ranjan et al. 2001; Sedigheh et al. 2011). The reduction of the Rubisco large subunit (Rubisco-L) in spl5 mutant might be caused by H2O2 over-accumulation (Chen et al. 2012). Moreover, in the present study we found that Rubisco activases (Rubisco-A), which catalyze Rubisco activation, were also down-regulated in spl5 mutant (Table 1). The reduction of Rubisco and/or Rubisco-A was also found in the lmms of spl1 (Kim et al. 2008), spl6 (Kang et al. 2007), crd2 (Tsunezuka et al. 2005), and blm (Jung et al. 2006), suggesting that the reduction in Rubisco and Rubisco-A accumulation is a shared phenomenon among lmms.
Glycolysis protein
A key enzyme of glycolysis, glyceraldehyde-3-phosphate dehydrogenase (GAPDH), was up-regulated in spl5 mutant (Table 1), as also observed in lmms spl1 (Kim et al. 2008) and cdr2 (Tsunezuka et al. 2005). GAPDH catalyzes the oxidation of dihydroxyacetone phosphate to glycerol-3-phosphate. More recently, GAPDH emerged as a multifunctional protein in several non-metabolic processes, namely a primary role in apoptosis. S-nitrosylated GAPDH initiates apoptotic cell death by nuclear translocation following Siah1 binding (Hara et al. 2005); GAPDH accumulates in mitochondria during apoptosis and induces the pro-apoptotic mitochondrial membrane permeabilization (Tarze et al. 2007); and it also mediates cell death by its nuclear translocation under oxidative stress (Nakajima et al. 2009). Additionally, it was reported that GLY1-encoded GAPDH plays an important role in plastidal oleic acid-mediated signaling and resistant signaling in Arabidopsis (Kachroo et al. 2004,2005; Chandra-Shekara et al. 2007; Xia et al. 2009). Compared to WT, Arabidopsis mutant gly1 is much more susceptible to pathogens, while plants with GLY 1 overexpression have enhanced resistance (Venugopal et al. 2009). Increased level of GAPDH protein in the spl1, spl5, and cdr2 mutants suggests that GAPDH may function in stimulating cell death and defense responses in rice.
ROS metabolism proteins
Two major enzymes of ROS-detoxification, glutathione S-transferase 14 (GST14) and APX7, were up-regulated in spl5 mutant (Table 1). ROS, such as superoxide anion (O2–) and H2O2, are toxic byproducts of aerobic metabolism (Mittler et al. 2004). Upon pathogen attack, ROS were immediately induced to kill the infected cells and also served as a signal to activate the defense response (Shigeoka et al. 2002). To avoid the oxidative damage to other cells in plants, the ROS must be scavenged by the antioxidant enzymes SOD, CAT, APX, or GST etc. H2O2 has been reported to up-regulate expression of APX (Lee et al. 1999; Morita et al. 1999). According to our previous results, H2O2 is over-accumulated in leaves of spl5 mutant (Chen et al. 2012). It is likely that the high level of H2O2 in this mutant induces the HR and increases the resistance to pathogens. Therefore, up-regulation of APX7 and GST14 might be responsible for scavenging excessive accumulation of ROS in spl5 mutant. However, inductions of APX7 and GST14 were apparently insufficient to detoxify the overproduction of ROS, which resulted in cell death in spl5 mutant.
Defense-related protein
A PR protein, Chia2a, was found to be differently expressed between WT and spl5 mutant (Table 1). Chia2a is a class II chitinase belonging to the PR-3 group (Muthukrishnan et al. 2001). Chitinase can break down glycosidic bonds in chitin, which is the main structural component of fungal cell walls and insect exoskeletons (Sela-Buurlage et al. 1993). The expression of the chitinase gene was significantly stimulated by fungi (Xu et al. 2008). Transgenic plants over-expressing chitinase gene showed enhanced resistance to fungal (Brogue et al. 1991; Dunsmuir et al. 1993; Oldach et al. 2001) and bacterial pathogens (Oldach et al. 2001). It is likely that the increased level of Chia2a is responsible, at least partially, for the enhanced resistance in spl5 mutant.
mRNA level of differentially expressed proteins in spl5 mutant
To assay the mRNA levels of 14 differentially expressed proteins in spl5 mutant, the semi-quantitative RT-PCRs were performed. We analyzed the expressions of these 14 genes in WT leaf blades and the three different parts of spl5 leaf blades, based on the degree of lesion formation: no lesion (NL), leaf area without any lesions; few lesions (FL), leaf area with 10–20% lesions; and many lesions (ML), leaf area with 70–80% lesions (Figure 3a). As shown in Figure 3b, in spl5 mutant, the expression of 6 genes were induced and 2 genes were suppressed, and 6 genes did not changed at mRNA level.
Figure 3.
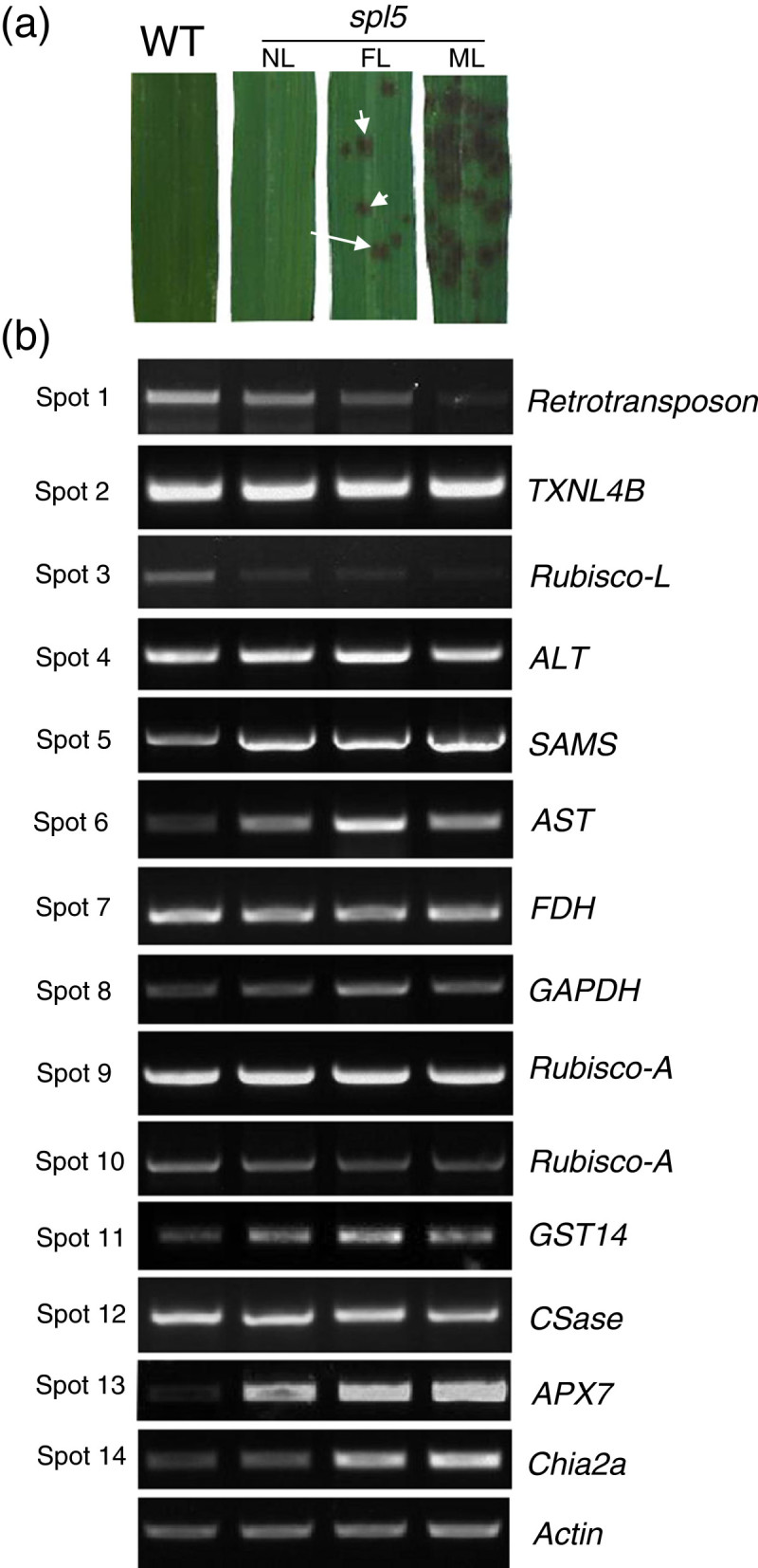
Gene expressions of candidate proteins by semi-quantitative RT-PCR. (a) Lesion-mimic phenotypes of spl5 mutants. WT (control), the leaf blades of WT; NL, FL and ML indicate no lesion, few lesions, and many lesions, respectively, in the leaf blades of spl5 mutant. Arrows indicate leaf lesions in spl5 mutant. (b) Semi-quantitative RT-PCR of 14-proteins’ genes in WT leaves and leaf parts with different lesions of spl5 mutant. The protein ID numbers are listed in the left of images, and the corresponding gene names are listed in the right of images. The Actin was used as a reference gene.
We compared the genes expression profile to our 2-DE data (Table 1). Most of the 7 up-regulated proteins (except spot 1) were transcriptionally induced in spl5 mutant. As expected, many of them (SAMS, GAPDH, GST14, APX7 and Chia2a) are defense- or PCD-related. It is likely that the SPL5 protein negatively regulates expression of these genes at transcriptional level. In contrast, the genes encoding down-regulated proteins (except spot 3) did not change significantly at the transcriptional level in spl5 mutant, and most of these proteins involve in amino-acid metabolism and photosynthesis. Based on the fact that the SPL5 gene encodes a subunit of splicing factor (Chen et al. 2012), it is likely that the genes encoding these down-regulated proteins might be controlled directly or indirectly by SPL5 through the post-transcriptional mRNA processing.
Further analysis of APX7 and Chia2a in spl5 mutant
Since the APX7 and Chia2a are directly involved in mediating PCD or defense responses, the gene expression of these proteins were further confirmed by qPCR. The qPCR results were similar to that of semi-quantitative RT-PCR (Figure 3b and Figure 4). The expression of APX7 increased in NL, FL, and ML of spl5 leaves, while the expression of Chia2a increased only in FL and ML but not in NL (Figure 4). The expression levels of these two genes were positively correlated with lesion numbers on the leaves of the spl5 mutant. Moreover, we further examined enzyme activities of APX and chitinase both in WT and spl5 mutant. As expected, APX and chitinase activities were proportional to the number of lesions on spl5 leaves (Figure 5).
Figure 4.
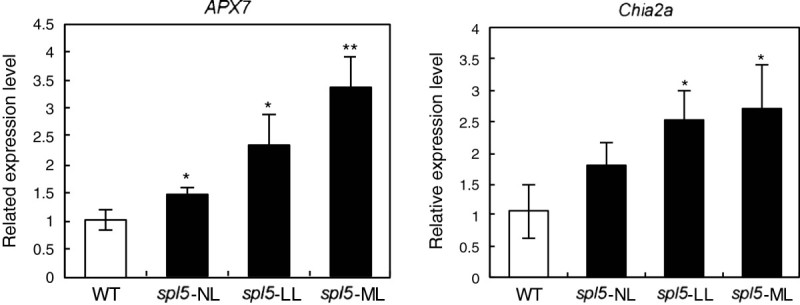
qPCR confirmation of gene expression for APX7 and Chia2a . qPCR results for APX7 and Chia2a in the leaf blades of WT (control) and leaf parts with different degree of lesions of spl5 mutant (see Figure 3a). Single and double asterisks indicate P < 0.05 and P < 0.01 (Student’s t-test), respectively.
Figure 5.
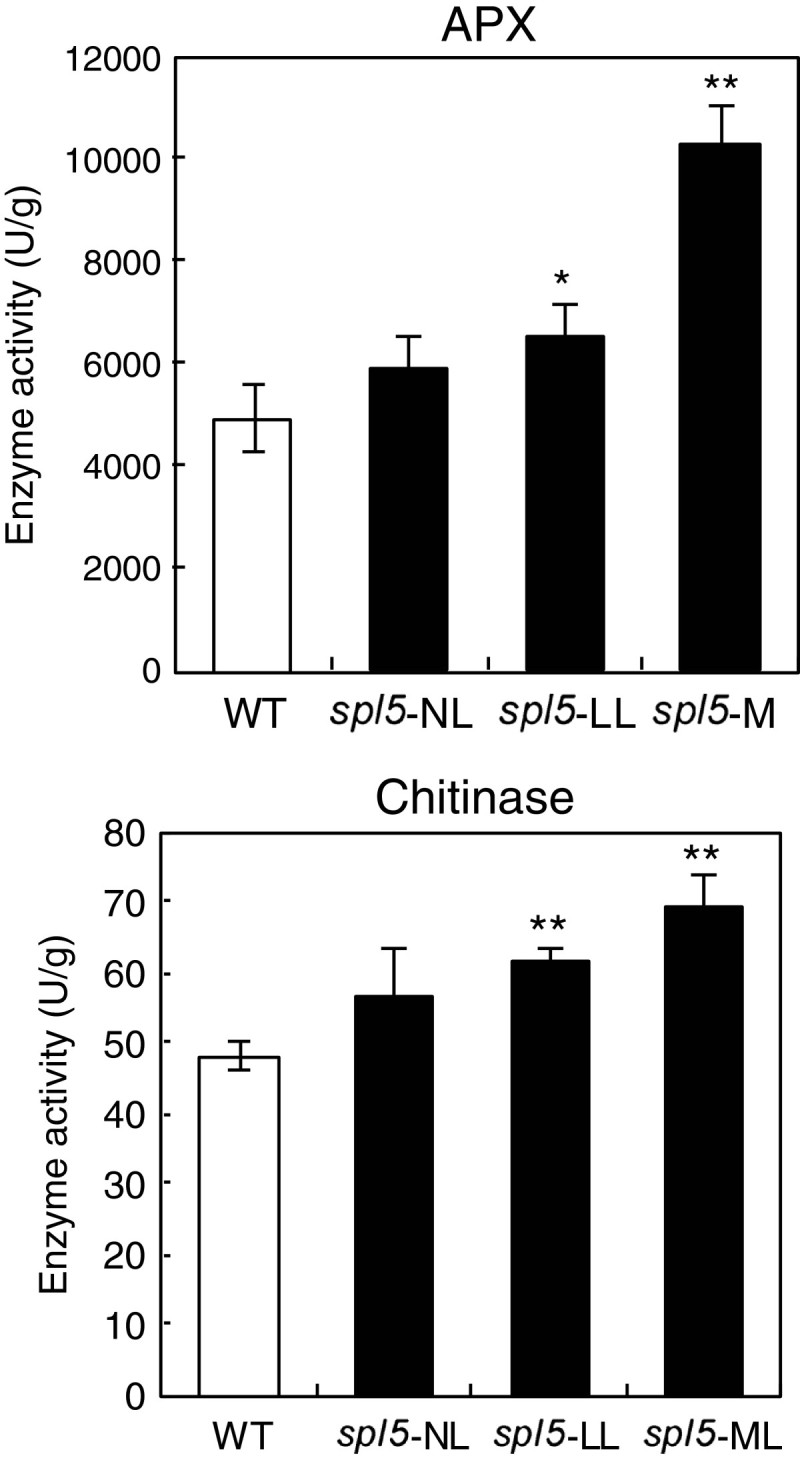
Enzyme activities of APX and Chitinase in spl5 mutant. Leaf parts with different number of lesions of spl5 mutant (see Figure 3a) were used to analyze enzyme activity: (a) APX and (b) chitinase. Single and double asterisks indicate P < 0.05 and P < 0.01 (Student’s t-test), respectively.
Conclusions
According to our 2-DE data and the proteomic results of other rice lmms (Takahashi et al. 2003; Tsunezuka et al. 2005; Jung et al. 2006; Kang et al. 2007; Kim et al. 2008), proteins involved in ROS scavenging, defense responses or cell death such as SOD (spl1, blm), CAT (spl6, blm), Peroxidase (spl1), APX (spl5, blm), GST (cdr2, spl5), were likely to be induced in lmms, whereas proteins involved in photosynthesis and glycolysis such as Rubisco (spl1, spl5, spl6, blm, crd2), Rubisco-A (spl5, spl6), and GAPDH (spl1, spl5, cdr2) were often deceased in lmms; defense-related protein like OsPR5 (spl1), OsPR10 (blm), PBZ1 (spl1, cdr2, blm), Chia2a (spl5) were significantly activated in lmms. This suggests that the differential accumulation of these proteins is common features for lmms.
Methods
Plant materials and growth conditions
An original spl5 mutant was screened from a γ-radiation-mutagenesis population of Norin8 (Oryza sativa L. ssp. japonica) by Iwata et al. (1978) in Japan. Zeng et al. (2003) crossed the spl5 mutant with WT Zhefu802 (a Chinese indica cultivar) through repeated backcrossing, and produced the mutant Zhefu802spl5/spl5 with spl5 lesion mimic phenotype as used in this study. The seeds of spl5 mutant and Zhefu802 were germinated in an incubator at 28°C and then incubated in nutrient solution (Yoshida et al. 1976) in a growth chamber at 28/24°C (day/night). The nutrient solution was maintained at pH 5.6 and refreshed each 5 d. The fully developed leaves of 60-days-old seedlings were collected from each plant and immediately frozen in liquid nitrogen and stored at −80°C.
Protein extraction
Total proteins were extracted from collected leaf samples of 10 plants and independently repeated three times. Leaf blades were ground in liquid nitrogen, and the tissue powder produced was immediately suspended in an extraction buffer containing 9.5 M urea, 4% w/v 3-[(3-cholamidopropyl) dimethylammonio]-1-propane-sulfonate (CHAPS), 65 mM dithiothreitol (DTT), and 2% v/v immobilized pH gradient (IPG) buffer pH 3–10. Crude homogenates were centrifuged at 4°C (9,000 × g for 30 min). The supernatants were precipitated by 10% TCA for 1 h at −20°C, followed by centrifugation at 13,000 × g for 30 min. The pellets were washed twice with cold acetone and allowed to air dry, and then resuspended with the extraction buffer and finally stored at −80°C. Protein contents were determined by the Bradford method (Bradford 1976) using a protein assay reagent (Bio-Rad, USA).
2-DE
Extracted proteins were analyzed by 2-DE and biologically repeated trice using different samples. For 2-DE, 100 and 500 μg of total proteins were loaded onto analytical and preparative gels, respectively. The Ettan IPGphor Isoelectric Focusing System (Amersham, USA) and pH 3–10 IPG strips (13 cm, nonlinear; Amersham) were used for isoelectric focusing (IEF). The IPG strips were rehydrated for 12 h in 250 μL of rehydration buffer containing the protein samples. IEF was performed in five steps: 30 V for 12 h, 500 V for 1 h, 1000 V for 1 h, 8000 V for 8 h, and 500 V for 4 h. The gel strips were equilibrated for 15 min in equilibration buffer [50 mM Tris–HCl (pH 8.8), 6 M urea, 2% sodium dodecyl sulfate (SDS), 30% glycerol, and 1% DTT]. This step was repeated using the same buffer with 4% iodoacetamide in place of 1% DTT. The strips were then subjected to the second-dimensional electrophoresis after transfer onto 12.5% SDS-polyacrylamide gels. Electrophoresis was performed using the Hofer SE 600 system (Amersham) at 15 mA per gel until the bromophenol blue reached the end of the gel. Both the proteins of WT and spl5 mutant were done 2-DE for 3 gels, respectively.
Gel staining and image analysis
After 2-DE, analytical gels were stained with ammoniacal silver nitrate based on the procedure described by Hochstrasser (1988), and preparative gels were stained with Coomassie Blue G250 (Bio-Rad). Resulting 2-D gels were scanned using an UMax Powerlook 2110XL Scanner (Amersham). The stained protein spots were detected using software Image Master (Amersham). After quantitative detection, the intensities of each spot were normalized by total valid spot intensity. The spots displaying significant changes were considered to be differentially expressed proteins. Expression differences per protein spot between the spl5 mutant and WT from 3 independent experiments were estimated by t-test (p < 0.05). Protein spots were selected based on the significant differences of spots quantities between the spl5 mutant and WT.
In-gel digestion
Protein spots were excised from preparative 2-DE gels and destained with 100 mM NH4HCO3 and 30% acetonitrile (ACN). After removing the destaining buffer, the gel pieces were lyophilized and rehydrated in 30 μL of 50 mM NH4HCO3 containing 50 ng of sequencing grade, modified trypsin (Promega, USA). After digestion overnight at 37°C, these peptides were extracted thrice with 0.1% trifluoroacetic acid (TFA) in 60% ACN, and extracts were pooled together and lyophilized. Peptide mixtures were redissolved in 0.1% TFA, desalted and concentrated using ZipTips from Millipore. Peptide solution (0.75 mL) was mixed with 0.75 mL of matrix [α-cyano-4-hydroxycinnamic acid (CHCA) in 30% ACN, 0.1% TFA] spotted on a target disk and allowed to air dry.
MALDI-TOF/TOF analysis and database searching
Mass spectra were acquired on a MALDI-TOF/TOF mass spectrometer, the Bruker-Daltonics AutoFlex TOF-TOF LIFT (Bruker, Germany). Protein database searching was performed with the MASCOT search engine (http://www.matrixscience.com) using monoisotopic peaks against the NCBI nonredundant protein database (http://www.ncbi.nlm.nih.gov/). The species selected was Oryza sativa.
Semi-quantitative RT-PCR and qPCR
Total RNAs of leaves were isolated by TRIzol Reagent (Invitrogen, USA). The first-strand synthesis of cDNAs was carried out by SuperScript® III First-Strand Synthesis System (Invitrogen) according to the manufacturer’s instruction. Semi-quantitative RT-PCR was performed for 25 cycles of 30 s at 94°C, 30 s at 60°C, and 1 min at 72°C. qPCR was performed in StepOne™ Real-Time PCR System (Applied Biosystems, USA) using the Fast SYBR Green Master Mix reagent (Applied Biosystems) by the manufacturer’s instructions, and the thermal cycle used was as follows: 95°C for 20 s; and 40 cycles of 95°C for 3 s, and 60°C for 30 s. OsRAc1 (GenBank accession: X16280), a rice constitutively expressed gene of Actin, was used as a standardization control, using the primer pair 5’-GGAACTGGTATGGTCAAGGC-3’ and 5’-AGTCTCATGGATACCCGCAG-3’ for semi--quantitative RT-PCR, 5’-TGGCATCTCTCAGCACATTCC-3’ and 5’-TGCACAATGGATGGGTCAGA-3’ for qPCR. Gene-specific primers of candidate genes for semi-quantitative PCR and qPCR are listed in Table 2 and Table 3, respectively. Independent biological repetitions of each experiment were performed three times.
Table 2.
Gene-specific primers for RT-PCR in this study
| aSpot id | bAccession | Forward primer (5’-3’) | Reverse primer (5’-3’) | c Tm (°C) |
|---|---|---|---|---|
| 1 | LOC_Os12g37540 | AACAAGGTAGGGATAGTTACTT | CCTTGTATGTGGGTTTTTTAGAA | 55 |
| 2 | AK067692 | AATGAAATCTTGCTTGCTGC | CTAAATCTTCTTGGGACATA | 60 |
| 3 | AK067692 | GCCTACTTCTTCACATTCAC | ATTTCATTACCTTCACGAGC | 55 |
| 4 | AK067732 | ACCCGCTTTATTCTGCTG | ATCCTTTTGACACAGTATGG | 60 |
| 5 | AK104875 | AGATGCTGTGCTTGATGCCT | CAATGACGAAGCGACCAGAT | 55 |
| 6 | AK105059 | CAAACAGGGTGAAGAGCCAG | CTCGCATTTAGCCAGGGACA | 62 |
| 7 | AK065872 | ACGCCGACAAGAATCCCAAC | CAATACGACCAGCCCCAACA | 60 |
| 8 | AK064960 | TTCATCACCACCGACTACAT | AACCCTCAACAATACCAAAC | 55 |
| 9 | AK060847 | GAAGCTGAAGAAGCAGGTGACATC | CGAAGACGAGCTCACACTGGAAG | 55 |
| 10 | AK104332 | ATGGGTGAATTCTGTGGTGAG | CCCTTCTTGATGATGTCTGCC | 58 |
| 11 | AK102889 | CTCGTTGCGGTAGTGCTGCT | AATGAAATCTTGCTTGCTGC | 55 |
| 12 | AK099598 | TGGCAGCGAAGACAAACAAC | CTGGAAGAGCACCGACGAAA | 62 |
| 13 | AK063934 | GCTTGAGATTTGATGTTGAG | GTCCTCTGCGTATTTTTCTG | 62 |
| 14 | AK070067 | CGACTTCTCCACCCTACTAT | ATGATGTTGGTGATGACGCC | 55 |
a Spot identity of protein in 2-D gel image; bmRMA accession of the corresponding protein in GenBank (http://www.ncbi.nlm.nih.gov/), except for spot 1 protein whose gene accession was only deposited in TIGR database (http://rice.plantbiology.msu.edu/index.shtml); cAnneal temperature of primer pair used in RT-PCR.
Table 3.
Gene-specific primers for qPCR in this study
| Gene name | Forward primer (5’-3’) | Reverse primer (5’-3’) |
|---|---|---|
| APX7 | ATACGCAGAGGACCAAGAAGCAT | CTACGAGCAAGATAAATAGCAGA |
| Chia2a | CCAACATCATCAACGGCGGCAT | TTGGGATACTACATCACTACAT |
Assay of enzyme activities
About 0.5–1 g of leaves were homogenized for activity assay of APX or chitinase, according to the methods previously described by Mishra et al. (1993) and Boller et al. (1983), respectively. Each experiment was independently repeated three times.
Acknowledgements
We thank Dr. Jianwei Pan for critical reading of this manuscript. This work was supported by the National Key Programs for Transgenic Crops (2011ZX08009-003), the National Natural Science Foundation of China (31171519, 31101130); the Natural Science Foundation of Zhejiang Province (Y3110234, Y3110230), the Welfare Technology Application Research Project of Zhejiang Province (2011C22005), and the Zhejiang Province Key Innovative Research Team Program (2010R50024).
Abbreviations
- 2-DE
Two-dimensional gel electrophoresis
- APX
Ascorbate peroxidase
- CAT
Catalase
- FL
Few lesions
- GAPDH
Glyceraldehyde-3-phosphate dehydrogenase
- GST
Glutathione S-transferase
- HR
Hypersensitive response
- IEF
Isoelectric focusing
- Lmm
Lesion mimic mutant
- ML
More lesions
- NL
No lesion
- PBZ1
Probenazole-induced protein
- PCD
Programmed cell death
- PR
Pathogenesis-related
- ROS
Reactive oxygen species
- SAMS
S-adenosylmethionine synthetase
- SF3b3
Splicing factor 3b subunit 3
- SOD
Superoxide dismutase
- spl
spotted leaf
- TXNL4B
Thioredoxin-like protein 4B
- WT
Wild-type.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
CX carried out the 2-DE analysis, protein alignments, and drafted the manuscript. FS designed PCR primers and carried out the RT-PCR, and participated in preparation of data. ZP carried out the assay of enzyme activities. GZ and LJ participated in the design of the study and performed the statistical analysis. QQ provided the plant materials of this study. MB conceived the study, and participated in its design and coordination and revised the manuscript. All authors read and approved the final manuscript.
Contributor Information
Xifeng Chen, Email: xfchen@zjnu.cn.
Shufang Fu, Email: fushufang@163.com.
Pinghua Zhang, Email: zph@zjnu.cn.
Zhimin Gu, Email: zmgu@zjnu.cn.
Jianzhong Liu, Email: jzliu@zjnu.cn.
Qian Qian, Email: qianqian188@hotmail.com.
Bojun Ma, Email: mbj@zjnu.cn.
References
- Agrawal GK, Rakwal R. Rice proteomics: a cornerstone for cereal food crop proteomes. Mass Spectrom Rev. 2006;25:1–53. doi: 10.1002/mas.20056. [DOI] [PubMed] [Google Scholar]
- Boller T, Gehri A, Mauch F, Vögeli U. Chitinase in bean leaves: induction by ethylene, purification, properties, and possible function. Planta. 1983;157:22–31. doi: 10.1007/BF00394536. [DOI] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Brogue K, Chet I, Holliday M, Cressman R, Biddle P, Knowlton S, Mauvais CJ, Broglie R. Transgenic plants with enhanced resistance to the fungal pathogen Rhizoctonia solani. Science. 1991;254:1194–1197. doi: 10.1126/science.254.5035.1194. [DOI] [PubMed] [Google Scholar]
- Burstenbinder K, Waduwara I, Schoor S, Moffatt BA, Wirtz M, Minocha SC, Oppermann Y, Bouchereau A, Hell R, Sauter M. Inhibition of 5’-methylthioadenosine metabolism in the yang cycle alters polyamine levels, and impairs seedling growth and reproduction in Arabidopsis. Plant J. 2010;62:977–988. doi: 10.1111/j.1365-313X.2010.04211.x. [DOI] [PubMed] [Google Scholar]
- Chandra-Shekara AC, Venugopal SC, Barman SR, Kachroo A, Kachroo P. Plastidial fatty acid levels regulate resistance gene-dependent defense signaling in Arabidopsis. Proc Natl Acad Sci USA. 2007;104:7277–7282. doi: 10.1073/pnas.0609259104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X, Pan J, Cheng J, Jiang G, Jin Y, Gu Z, Qian Q, Zhai W, Ma B. Fine genetic mapping and physical delimitation of the lesion mimic gene spotted leaf 5 (spl5) in rice (oryza sativa L.) Mol Breeding. 2009;24:387–395. doi: 10.1007/s11032-009-9299-2. [DOI] [Google Scholar]
- Chen X, Hao L, Pan J, Zheng X, Jiang G, Jin Y, Gu Z, Qian Q, Zhai W, Ma B. SPL5, a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice. Mol Breeding. 2012;30(2):939–949. doi: 10.1007/s11032-011-9677-4. [DOI] [Google Scholar]
- Chern M, Fitzgerald HA, Canlas PE, Navarre DA, Ronald PC. Overexpression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Mol Plant Microbe Interact. 2005;18:511–520. doi: 10.1094/MPMI-18-0511. [DOI] [PubMed] [Google Scholar]
- Dietrich RA, Delaney TP, Uknes SJ, Ward ER, Ryals JA, Dangl JL. Arabidopsis mutants simulating disease resistance response. Cell. 1994;77:565–577. doi: 10.1016/0092-8674(94)90218-6. [DOI] [PubMed] [Google Scholar]
- Dunsmuir P, Howie W, Newbigin E, Joe L, Penzes E. Resistance to rhizoctonia solani in transgenic tobacco. In: Nester EW, Verma DPS, editors. Advance in molecular genetics of plant-microbe interactions. Netherlands: Kluwer Academic Publishers; 1993. [Google Scholar]
- Fujiwara T, Maisonneuve S, Isshiki M, Mizutani M, Chen L, Wong HL, Kawasaki T, Shimamoto K. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J Biol Chem. 2010;285:11308–11313. doi: 10.1074/jbc.M109.091371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geraats BPJ. The role of ethylene perception in plant disease resistance. Netherlands: Utrecht University; 2003. [Google Scholar]
- Gonzalez ME, Marco F, Minguet EG, Carrasco-Sorli P, Blazquez MA, Carbonell J, Ruiz OA, Pieckenstain FL. Perturbation of spermine synthase gene expression and transcript profiling provide new insights on the role of the tetraamine spermine in Arabidopsis defense against pseudomonas viridiflava. Plant Physiol. 2011;156:2266–2277. doi: 10.1104/pp.110.171413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hara MR, Agrawal N, Kim SF, Cascio MB, Fujimuro M, Ozeki Y, Takahashi M, Cheah JH, Tankou SK, Hester LD, Ferris CD, Hayward SD, Snyder SH, Sawa A. S-nitrosylated GAPDH initiates apoptotic cell death by nuclear translocation following Siah1 binding. Nat Cell Biol. 2005;7:665–674. doi: 10.1038/ncb1268. [DOI] [PubMed] [Google Scholar]
- Hochstrasser DF, Patchornik A, Merril CR. Development of polyacrylamide gels that improve the separation of proteins and their detection by silver staining. Anal Biochem. 1988;173:412–423. doi: 10.1016/0003-2697(88)90208-4. [DOI] [PubMed] [Google Scholar]
- Iwata N, Omura T, Satoh H. Linkage studies in rice (oryza sativa L.) on some mutants for physiological leaf spots. J Fac Agr Kyushu Univ. 1978;22:243–251. [Google Scholar]
- Johal G, Hulbert S, Briggs S. Disease lesion mimic mutations of maize: a model for cell death in plants. Bioessays. 1995;17:685–692. doi: 10.1002/bies.950170805. [DOI] [Google Scholar]
- Jung YH, Lee JH, Agrawal GK, Rakwal R, Kim JA, Shim JK, Lee SK, Jeon JS, Koh HJ, Lee YH, Iwahashi H, Jwa NS. The rice (oryza sativa) blast lesion mimic mutant, blm, may confer resistance to blast pathogens by triggering multiple defense-associated signaling pathways. Plant Physiol Biochem. 2005;43:397–406. doi: 10.1016/j.plaphy.2005.03.002. [DOI] [PubMed] [Google Scholar]
- Jung YH, Rakwal R, Agrawal GK, Shibato J, Kim JA, Lee MO, Choi PK, Jung SH, Kim SH, Koh HJ, Yonekura M, Iwahashi H, Jwa NS. Differential expression of defense/stress-related marker proteins in leaves of a unique rice blast lesion mimic mutant (blm) J Proteome Res. 2006;5:2586–2598. doi: 10.1021/pr060092c. [DOI] [PubMed] [Google Scholar]
- Kachroo A, Venugopal SC, Lapchyk L, Falcone D, Hildebrand D, Kachroo P. Oleic acid levels regulated by glycerolipid metabolism modulate defense gene expression in Arabidopsis. Proc Natl Acad Sci USA. 2004;101:5152–5157. doi: 10.1073/pnas.0401315101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kachroo P, Venugopal SC, Navarre DA, Lapchyk L, Kachroo A. Role of salicylic acid and fatty acid desaturation pathways in ssi2-mediated signaling. Plant Physiol. 2005;139:1717–1735. doi: 10.1104/pp.105.071662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang SG, Matin MN, Bae H, Natarajan S. Proteome analysis and characterization of phenotypes of lesion mimic mutant spotted leaf 6 in rice. Proteomics. 2007;7:2447–2458. doi: 10.1002/pmic.200600961. [DOI] [PubMed] [Google Scholar]
- Kim ST, Kim SG, Kang YH, Wang Y, Kim JY, Yi N, Kim JK, Rakwal R, Koh HJ, Kang KY. Proteomics analysis of rice lesion mimic mutant (spl1) reveals tightly localized probenazole-induced protein (PBZ1) in cells undergoing programmed cell death. J Proteome Res. 2008;7:1750–1760. doi: 10.1021/pr700878t. [DOI] [PubMed] [Google Scholar]
- Kim JA, Cho K, Singh R, Jung YH, Jeong SH, Kim SH, Lee JE, Cho YS, Agrawal GK, Rakwal R, Tamogami S, Kersten B, Jeon JS, An G, Jwa NS. Rice OsACDR1 (Oryza sativa accelerated cell death and resistance 1) is a potential positive regulator of fungal disease resistance. Mol Cells. 2009;28:431–439. doi: 10.1007/s10059-009-0161-5. [DOI] [PubMed] [Google Scholar]
- Kramer A. The structure and function of proteins involved in mammalian pre-mRNA splicing. Annu Rev Biochem. 1996;65:367–409. doi: 10.1146/annurev.bi.65.070196.002055. [DOI] [PubMed] [Google Scholar]
- Lee SC, Kang BG, Oh SE. Induction of ascorbate peroxidase isoenzymes by ethylene and hydrogen peroxide during growth of cultured soybean cells. Molecular Cells. 1999;9:166–171. [PubMed] [Google Scholar]
- Marina M, Maiale SJ, Rossi FR, Romero MF, Rivas EI, Garriz A, Ruiz OA, Pieckenstain FL. Apoplastic polyamine oxidation plays different roles in local responses of tobacco to infection by the necrotrophic fungus sclerotinia sclerotiorum and the biotrophic bacterium pseudomonas viridiflava. Plant Physiol. 2008;147:2164–2178. doi: 10.1104/pp.108.122614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mishra NP, Mishra RK, Singhal GS. Changes in the activities of anti-oxidant enzymes during exposure of intact wheat leaves to strong visible light at different temperatures in the presence of protein synthesis inhibitors. Plant Physiol. 1993;102:903–910. doi: 10.1104/pp.102.3.903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mittler R, Vanderauwera S, Gollery M, Van Breusegem F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004;9:490–498. doi: 10.1016/j.tplants.2004.08.009. [DOI] [PubMed] [Google Scholar]
- Mizobuchi R, Hirabayashi H, Kaji R. Isolation and characterization of rice lesion-mimic mutants with enhanced resistance to rice blast and bacterial blight. Plant Sci. 2002;163:345–353. doi: 10.1016/S0168-9452(02)00134-6. [DOI] [Google Scholar]
- Morel JB, Dangl JL. The hypersensitive response and the induction of cell death in plants. Cell Death Differ. 1997;4:671–683. doi: 10.1038/sj.cdd.4400309. [DOI] [PubMed] [Google Scholar]
- Mori M, Tomita C, Sugimoto K, Hasegawa M, Hayashi N, Dubouzet JG, Ochiai H, Sekimoto H, Hirochika H, Kikuchi S. Isolation and molecular characterization of a spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol Biol. 2007;63:847–860. doi: 10.1007/s11103-006-9130-y. [DOI] [PubMed] [Google Scholar]
- Morita S, Kaminaka H, Masumura T, Tanaka K. Induction of rice cytosolic ascorbate peroxidase mRNA by oxidative stress; the involvement of hydrogen peroxide in oxidative stress signalling. Plant Cell Physiol. 1999;40:417–422. doi: 10.1093/oxfordjournals.pcp.a029557. [DOI] [Google Scholar]
- Moschou PN, Sarris PF, Skandalis N, Andriopoulou AH, Paschalidis KA, Panopoulos NJ, Roubelakis-Angelakis KA. Engineered polyamine catabolism preinduces tolerance of tobacco to bacteria and oomycetes. Plant Physiol. 2009;149:1970–1981. doi: 10.1104/pp.108.134932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muthukrishnan S, Liang G, Trick H, Gill B. Pathogenesis-related proteins and their genes in cereals. Plant Cell. 2001;64:93–114. [Google Scholar]
- Nakajima H, Amano W, Kubo T, Fukuhara A, Ihara H, Azuma YT, Tajima H, Inui T, Sawa A, Takeuchi T. Glyceraldehyde-3-phosphate dehydrogenase aggregate formation participates in oxidative stress-induced cell death. J Biol Chem. 2009;284:34331–34341. doi: 10.1074/jbc.M109.027698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oldach KH, Becker D, Lorz H. Heterologous expression of genes mediating enhanced fungal resistance in transgenic wheat. Mol Plant Microbe Interact. 2001;14:832–838. doi: 10.1094/MPMI.2001.14.7.832. [DOI] [PubMed] [Google Scholar]
- Qiao Y, Jiang W, Lee J, Park B, Choi MS, Piao R, Woo MO, Roh JH, Han L, Paek NC, Seo HS, Koh HJ. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit micro 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (oryza sativa) New Phytol. 2010;185:258–274. doi: 10.1111/j.1469-8137.2009.03047.x. [DOI] [PubMed] [Google Scholar]
- Ranjan R, Bohra S, Asija MJ. Plant senescence: physiological, biochemical and molecular aspects. India: Agrobios; 2001. [Google Scholar]
- Sedigheh HG, Mortazavian M, Norouzian D, Atyabi M, Akbarzadeh A, Hasanpoor K, Ghorbani M. Oxidative stress and leaf senescence. BMC Res Notes. 2011;4:477. doi: 10.1186/1756-0500-4-477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sela-Buurlage MB, Ponstein AS, Bres-Vloemans SA, Melchers LS, van den Elzen PJM, Comelissen BJC. Only specific tobacco (nicotiana tabacum) chitinases and [beta]-1,3-glucanase exhibit antifungal activity. Plant Physiol. 1993;101:857–863. doi: 10.1104/pp.101.3.857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen X, Liu H, Yuan B, Li X, Xu C, Wang S. OsEDR1 negatively regulates rice bacterial resistance via activation of ethylene biosynthesis. Plant Cell Environ. 2011;34:179–191. doi: 10.1111/j.1365-3040.2010.02219.x. [DOI] [PubMed] [Google Scholar]
- Shigeoka S, Ishikawa T, Tamoi M, Miyagawa Y, Takeda T, Yabuta Y, Yoshimura K. Regulation and function of ascorbate peroxidase isoenzymes. J Ex Bot. 2002;53:1305–1319. doi: 10.1093/jexbot/53.372.1305. [DOI] [PubMed] [Google Scholar]
- Staskawicz BJ, Ausubel FM, Baker BJ, Ellis JG, Jones JD. Molecular genetics of plant disease resistance. Science. 1995;268:661–667. doi: 10.1126/science.7732374. [DOI] [PubMed] [Google Scholar]
- Sun X, Zhang H, Wang D, Ma D, Shen Y, Shang Y. DLP, a novel Dim1 family protein implicated in pre-mRNA splicing and cell cycle progression. J Biol Chem. 2004;279:32839–32847. doi: 10.1074/jbc.M402522200. [DOI] [PubMed] [Google Scholar]
- Sun C, Liu L, Tang J, Lin A, Zhang F, Fang J, Zhang G, Chu C. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J Genet Genomics. 2011;38:29–37. doi: 10.1016/j.jcg.2010.12.001. [DOI] [PubMed] [Google Scholar]
- Takahashi A, Agrawal GK, Yamazaki M, Onosato K, Miyao A, Kawasaki T, Shimamoto K, Hirochika H. Rice Pti1a negatively regulates RAR1-dependent defense responses. Plant Cell. 2007;19:2940–2951. doi: 10.1105/tpc.106.047142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi A, Kawasaki T, Henmi K, Shi IK, Kodama O, Satoh H, Shimamoto K. Lesion mimic mutants of rice with alterations in early signaling events of defense. Plant J. 1999;17:535–545. doi: 10.1046/j.1365-313X.1999.00405.x. [DOI] [PubMed] [Google Scholar]
- Takahashi A, Kawasaki T, Wong HL, Suharsono U, Hirano H, Shimamoto K. Hyperphosphorylation of a mitochondrial protein, prohibitin, is induced by calyculin A in a rice lesion-mimic mutant cdr1. Plant Physiol. 2003;132:1861–1869. doi: 10.1104/pp.103.021733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tarze A, Deniaud A, Le Bras M, Maillier E, Molle D, Larochette N, Zamzami N, Jan G, Kroemer G, Brenner C. GAPDH, a novel regulator of the pro-apoptotic mitochondrial membrane permeabilization. Oncogene. 2007;26:2606–2620. doi: 10.1038/sj.onc.1210074. [DOI] [PubMed] [Google Scholar]
- Tsunezuka H, Fujiwara M, Kawasaki T, Shimamoto K. Proteome analysis of programmed cell death and defense signaling using the rice lesion mimic mutant cdr2. Mol Plant Microbe Interact. 2005;18:52–59. doi: 10.1094/MPMI-18-0052. [DOI] [PubMed] [Google Scholar]
- Venugopal SC, Chanda B, Vaillancourt L, Kachroo A, Kachroo P. The common metabolite glycerol-3-phosphate is a novel regulator of plant defense signaling. Plant Signal Behav. 2009;4:746–749. doi: 10.4161/psb.4.8.9111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang L, Pei Z, Tian Y, He C. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol Plant Microbe Interact. 2005;18:375–384. doi: 10.1094/MPMI-18-0375. [DOI] [PubMed] [Google Scholar]
- Wolter M, Hollricher K, Salamini F, Schulze-Lefert P. The mlo resistance alleles to powdery mildew infection in barley trigger a developmentally controlled defence mimic phenotype. Mol Gen Genet. 1993;239:122–128. doi: 10.1007/BF00281610. [DOI] [PubMed] [Google Scholar]
- Wu C, Bordeos A, Madamba MR, Baraoidan M, Ramos M, Wang GL, Leach JE, Leung H. Rice lesion mimic mutants with enhanced resistance to diseases. Mol Genet Genomics. 2008;279:605–619. doi: 10.1007/s00438-008-0337-2. [DOI] [PubMed] [Google Scholar]
- Xia Y, Gao Q, Yu K, Lapchyk L, Navarre DA, Hildebrand D, Kachroo A, Kachroo P. An intact cuticle in distal tissues is essential for the induction of systemic acquired resistance in plants. Cell Host Microbe. 2009;5:151–165. doi: 10.1016/j.chom.2009.01.001. [DOI] [PubMed] [Google Scholar]
- Xu X, Qin G, Tian S. Effect of microbial biocontrol agents on alleviating oxidative damage of peach fruit subjected to fungal pathogen. Int J Food Microbiol. 2008;126:153–158. doi: 10.1016/j.ijfoodmicro.2008.05.019. [DOI] [PubMed] [Google Scholar]
- Yamanouchi U, Yano M, Lin H, Ashikari M, Yamada K. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc Natl Acad Sci USA. 2002;99:7530–7535. doi: 10.1073/pnas.112209199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yin Z, Chen J, Zeng L, Goh M, Leung H, Khush GS, Wang G. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol Plant Microbe Interact. 2000;13:869–876. doi: 10.1094/MPMI.2000.13.8.869. [DOI] [PubMed] [Google Scholar]
- Yoshida S, Forno DA, Cook JH, Gomes KA. Routine procedure for growing rice plants in culture solution. Philippines, In: IRRI Laboratory manual for physiological studies of rice. Los Banos; 1976. [Google Scholar]
- Zeng D, Qian Q, Dong G, Zhu X, Dong F, Teng S, Guo L, Cao L, Cheng S, Xiong Z. Development of isogenic lines of morphological markers in indica rice. Acta Botanica Sinica. 2003;45:1116–1120. [Google Scholar]
- Zeng L, Qu S, Bordeos A, Yang C, Baraoidan M, Yan H, Xie Q, Nahm BH, Leung H, Wang G. Spotted leaf11, a negative regulator of plant cell death and defense, encodes a U-box/armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell. 2004;16:2795–2808. doi: 10.1105/tpc.104.025171. [DOI] [PMC free article] [PubMed] [Google Scholar]