

Abstract
We investigated problem solving abilities of capuchin monkeys via the “floating object problem,” a task in which the subject must use creative problem solving to retrieve a favoured food item from the bottom of a clear tube. Some great apes have solved this problem by adding water to raise the object to a level at which it can be easily grabbed. We presented seven capuchins with the task over eight trials (four “dry” and four “wet”). None of the subjects solved the task, indicating that no capuchin demonstrated insightful problem solving under these experimental conditions. We then investigated whether capuchins would emulate a solution to the task. Seven subjects observed a human model solve the problem by pouring water from a cup into the tube, which brought the object to the top of the tube, allowing the subject to retrieve it. Subjects were then allowed to interact freely with an unfilled tube containing the object in the presence of water and objects that could be used to solve the task. While most subjects were unable to solve the task after viewing a demonstrator solve it, one subject did so, but in a unique way. Our results are consistent with some previous results in great ape species, and indicate that capuchins do not spontaneously solve the floating object problem via insight.
Keywords: problem solving, Sapajus, Cebus, emulation, insight
While the use of tools to produce novel solutions to problems has become a popular topic reported in a variety of species in the comparative and cognitive literature, less is known of the initial development of these novel solutions. Are they arrived at via trial and error learning or in a manner familiar to readers who have experienced an ‘aha’ moment, i.e., a sudden realization of the solution to a problem? The latter is referred to as insightful problem solving, the “sudden production of a new adaptive response not arrived at by trial behaviour” (Thorpe 1963), and has been studied in a variety of tasks with rats (Tolman and Honzik 1930), corvids (Bird and Emery 2009; Heinrich 2000), and great apes (Köhler 1931). Results of these and other studies suggest that numerous animal species are capable of insightful problem solving.
Recently, a task called the “floating peanut problem” or “floating object task” was developed to examine the insightful use of water as a tool by great apes. Some captive great ape populations solved the floating object task at high rates: for example, all five orangutans tested by Mendes et al. (2007) were able to retrieve a peanut by adding water to a fixed vertical tube in which the peanut was placed. However, other captive populations performed relatively poorly on this task, with low rates of spontaneous solution; these include the chimpanzees, gorillas, and orangutans tested by Hanus et al. (2011). Children's solution rates of this task depend on their age, with low rates of solution at age 4 (Nielsen 2013) increasing to high, but not universal solution at age 8 (Hanus et al. 2011).
Because of their close phylogenetic relationship with humans, great apes have often been the focus of research into the cognitive capabilities of nonhuman primates. However, capuchin monkeys (Sapajus/Cebus spp. (Lynch Alfaro et al. 2012)) share several cognitive characteristics with humans and other great apes. Capuchins have a large relative brain size as adults (Rilling and Insel 1999) and are one of the few New World monkey species that have been documented using tools in the wild (e.g., hammer and anvil nut cracking (Izawa and Mizuno 1977), “sponge-dipping” (Phillips 1998) and grooming with sticks (Visalberghi 1990)). Capuchins have direct cortico-motoneuronal connections; the dense projection into the lateral and dorsal motor nuclei that innervate the intrinsic hand muscles contributes to their high manual dexterity (Lemon 2008). Capuchins also have well-developed parietal areas 2 and 5 – regions that receive input from the hand (Padberg et al. 2007). This sensorimotor integration enables capuchins to use pseudo-opposition between the side of the index finger and tip of the thumb, providing precision handling and individual digit control to grasp and manipulate objects and tools (Fragaszy 1998; Phillips 1998; Spinozzi et al. 2007). Different groups of wild capuchins, like chimpanzees and orangutans (van Schaik et al. 2003; Whiten et al. 1999), have differing suites of behavioral traditions (Panger et al. 2002; Perry et al. 2003) but it is unknown as to whether these traditions were initially arrived at via insight or trial and error. Capuchins have been observed to engage in social learning and are able learn from both conspecifics (Bonnie and De Waal 2007; Dindo et al. 2008; Fredman and Whiten 2008) and human demonstrators(Custance et al. 1999; Fredman and Whiten 2008). Because of these shared behavioural and cognitive characteristics, it is important to examine the performance of capuchins on tasks which are usually given to great apes, as they may well be able to solve these tasks.
As performance on the floating object task has not been previously investigated in capuchin monkeys we presented this task to captive animals with two goals: (i) to determine whether they would insightfully solve the task (Experiment 1), and (ii) failing this, to determine whether they would solve it after viewing a demonstration of the solution by a human experimenter (Experiment 2).
Experiment 1 – Insightful Problem Solving
The goal of the problem was for the subject to get an object (a preferred food item – a miniature marshmallow) out of the tube. Water (available via a Lix-it and a small plastic child's pool filled with water), straw, and other manipulanda were available in the enclosure to aid problem solving. While we did not expect the capuchins to solve this problem in the same way as great apes have (i.e., by gathering water in the mouth and spitting it into the tube), we reasoned capuchins might use their dextrous hands and objects in the enclosure to get water into the tube.
Method
Subjects
Seven capuchin monkeys (Cebus apella or Sapajus apella, (Lynch Alfaro et al. 2012)) were used in this study, including four females and three males ranging in age from 2 to 27 years (M = 11.38 years). These capuchins previously participated in studies of curiosity and motor skill (Phillips et al. 2012; Phillips and Thompson 2013).
The monkeys were socially housed at the Southwest National Primate Research Center (Texas Biomedical Research Institute, San Antonio, Texas) in an indoor/outdoor enclosure equipped with perches, swings, PVC tubes and joints, a Jolly Teaser ball, and fresh browse. During the summer, when this study was conducted, a small children's plastic pool containing water was in the outdoor portion of the enclosure. New World Monkey chow and water were available ad libitum; fresh fruit or nuts were provided once a day. This research (both Experiments) complied with the protocol approved by the IACUC at Texas Biomedical Research Institute, adhered to the Federal requirements governing research with nonhuman primates in the United States of America, and adhered to the American Society of Primatologists Principles for the Ethical Treatment of Nonhuman Primates.
Procedure
Each monkey was temporarily separated from the social group for testing. As the group enclosure had indoor and outdoor components, this allowed for one individual to be isolated for testing outdoors while the rest of the group was indoors. The monkeys who were in the indoor enclosure did not have visual access to the outdoor enclosure. All monkeys were habituated to this temporary separation before testing began.
The testing procedure was similar to that used by Mendes et al. (2007). A clear, Plexiglas tube (12.5 cm long, 2 cm diameter) was vertically attached to the outside of the enclosure with metal clips. The bottom of the tube was closed (forming a water tight vial in which to place the object and water depending on the trial). Monkeys were initially given four “dry” trials, in which no water was in the tube when it was presented to them. In “wet” trials, the tube was one-quarter filled with water before the trial began. The monkey was allowed into the outdoor enclosure once the experiment was set up. The Experimenter, who was positioned outside the enclosure, gained the attention of the subject with the marshmallow and then dropped it into the tube. The marshmallow either was at the bottom of the tube (dry trials) or floated in the water (wet trials), but was still unreachable. Trials lasted 10 minutes, but if the monkey showed interest in the task, then the time was extended by 5 minutes, up to two times, for a possible total time of 20 minutes per trial. At the end of the trial the monkey rejoined the social group. We recorded whether the marshmallow was retrieved and all occurrences of the behavior the subject demonstrated while interacting with the apparatus. Each monkey received a total of four dry trials and then four wet trials with a one-week inter-trial interval in each condition. The dry trials give the least information about possible solutions, while the presence of water in the tube in wet trials gives more information (see Hanus et al. 2011, Experiment 2). Dry trials serve as a baseline; if individuals solve the problem in the wet but not the dry condition, it may be the presence of water that spurs them to discover a solution. In this study, dry trials were given before the wet trials (to ensure that the sample size for evaluating the baseline solution rate was sufficient).
Results
All seven capuchins were interested in the tube and the marshmallow across most trials, as evidenced by behaviour such as putting their fingers into the tube, biting the top, and slapping the top of the tube with their hand. Monkeys infrequently used objects in the enclosure such as PVC or straw in their manipulative actions on the tube. However, no monkey added water to the tube or retrieved the marshmallow from the tube on any trial.
All monkeys displayed manipulative behaviour toward the tube during all four dry trials. Across the dry trials, the mean frequency of all actions directed toward the tube changed significantly over time (repeated measures ANOVA, F(3, 18) = 3.48, p = 0.038; see Figure 1). Post-hoc tests revealed that from Trial 2 to Trial 4, capuchins directed significantly fewer manipulative actions toward the tube (mean difference = 11.14, SE = 4.15 and p = 0.048).
Figure 1.
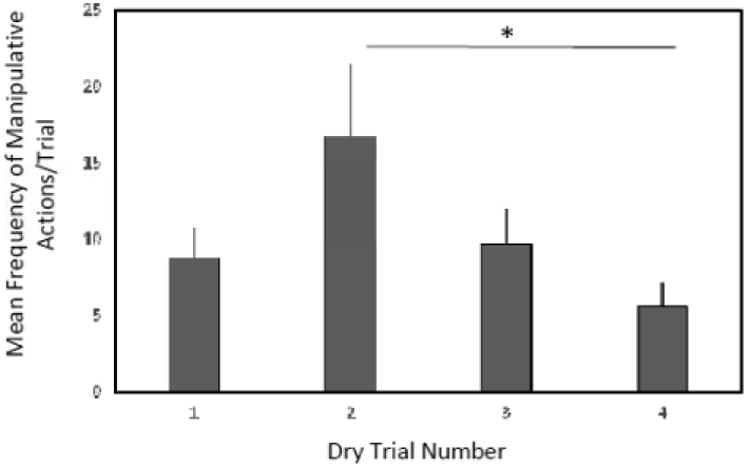
Mean frequency of manipulative actions performed during the Dry Trials in Experiment 1. Capuchins performed significantly fewer manipulative actions from Trial 2 to Trial 4 (p < 0.05).
The frequency of manipulative actions directed toward the tube did not significantly differ across the wet trials (repeated measures ANOVA, F(3,12) = 1.24, p = 0.34; see Figure 2). Three monkeys did not interact with the tube on the fourth trial.
Figure 2.
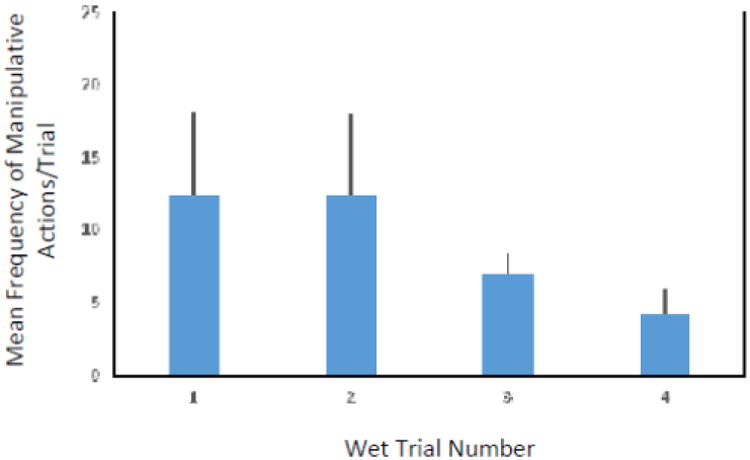
Mean frequency of manipulative actions performed during the Wet Trials in Experiment 1.
Discussion
The capuchins could have solved this task in a couple of different ways. One solution would have been to use a piece of PVC in the enclosure (such as T-joint or Jolly Teaser ball) to transfer water from the pool or Lix-it to the tube in order to float the marshmallow up the tube and into a reachable position. No monkey engaged in this behaviour during either dry or wet trials. A second solution (and perhaps more probable given capuchins' well-documented abilities to engage in tool use) would have been to use straw browse to spear the marshmallow. No monkey attempted to use straw or other objects in the enclosure as a tool to retrieve the marshmallow. Thus, none of the capuchins demonstrated insightful problem solving on the floating object task under these experimental conditions.
Experiment 2 – Emulation
In addition to its utility in testing insightful problem solving, the floating object task has been used to explore social learning. Social demonstrations may consist of a model engaging in a full or partial solution to the task. Social demonstration of the solution to this problem has been shown to spur some (but not all) individuals to solve the task (children (Nielsen 2013) and chimpanzees (Tennie et al. 2010)). When solving this task after seeing a demonstration of the solution, a learner may copy the end result but use unique, undemonstrated actions (engaging in emulation) or copy both the actions of and results obtained by the demonstrator (engaging in imitation) (Horner and Whiten 2005; Whiten and Ham 1992). For example, after seeing a human use a separate container to pour water into the tube, some chimpanzees spit water into the tube (Tennie et al. 2010); this is emulation. Use of a container to pour water into the tube would constitute imitation.
Capuchin monkeys engage in social learning and are able to learn from both capuchin (Bonnie and De Waal 2007; Dindo et al. 2008; Fredman and Whiten 2008) and human (Fredman and Whiten 2008) demonstrators. The next experiment thus investigated whether capuchins could solve the floating object problem after viewing a demonstration of the solution by a human experimenter.
Method
Subjects
All seven of the subjects from Experiment 1 participated in Experiment 2.
Procedure
We temporarily separated each monkey from the social group during testing similarly as in Experiment 1. Testing occurred in the outdoor portion of the enclosure. The outdoor enclosure had numerous enrichment items and objects, including perches, swings, PVC tubes and joints, a Jolly Teaser ball, fresh browse, and plastic cups.
Two clear, Plexiglas tubes (each 12.5 cm long, 2 cm diameter) were vertically attached using metal clips to the outside of the enclosure. The tubes were approximately 10 cm apart. The bottoms of the tubes were closed. The monkey was allowed into the outdoor enclosure once the experiment was set up. The human Demonstrator was positioned outside the enclosure, gained the attention of the monkey and dropped a marshmallow into one of the tubes. Monkeys then observed the human solve the task by pouring water from a cup into the tube, which brought the marshmallow to the top of the tube. The monkey was allowed to grab and consume the marshmallow. After the demonstration, retrieval, and consumption of the marshmallow, the Demonstrator regained the attention of the monkey and dropped a marshmallow into the unfilled tube. The capuchin was allowed to interact freely with the tube.
Trials lasted 10 minutes, but if the monkey showed interest in the task, then the time was extended by 5 minutes, up to two times, for a possible total time of 20 minutes. At the end of the trial the monkey rejoined the social group. We recorded whether the marshmallow was retrieved and the behaviour the subject demonstrated while interacting with the apparatus. Each monkey received four trials with the human Demonstrator with a one-week inter-trial interval. The Demonstrator was constant across all trials.
Results
All monkeys were interested in the tube and the marshmallow on all trials. After observing the Demonstrator pour water in the tube, they engaged in similar behaviour as they did in Experiment 1: they placed their fingers into the tube, bit the top of the tube, tried to insert large objects into the tube, and slapped the top of the tube with their hand. The mean frequency of manipulative actions toward the tube was the highest on Trial 2 (largely due to the increased activity by one monkey), and declined thereafter. Two monkeys did not interact with the tube on Trial 4. There was no significant difference in the frequency of manipulative actions directed toward the tube across the four trials (F(3, 12) = 0.796, p = 0.52; see Figure 3).
Figure 3.
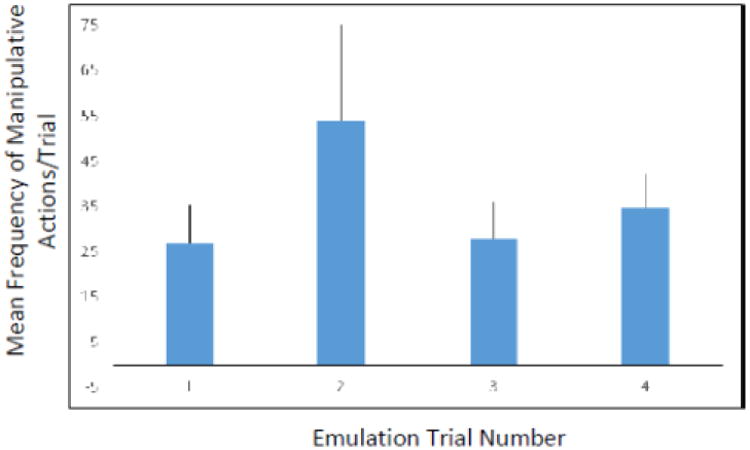
Mean frequency of manipulative actions performed during the Emulation trials in Experiment 2.
Six of the capuchins never solved the problem. The seventh capuchin also did not solve the problem on the first three trials. However, during the fourth trial, one capuchin solved the problem in a unique manner that did not involve adding water to the tube. The monkey, a subadult female, retrieved a wad of straw browse and pushed the wad all the way into the tube such that the straw was touching the bottom of the tube. She then slowly pulled out the wad of straw, to which the marshmallow was adhered, and retrieved the marshmallow. She performed this behaviour once, successfully.
General Discussion
Capuchins did not solve the floating object problem via insight alone. This is similar to results previously obtained with great apes; in one study, none of 31 chimpanzees solved the problem in the baseline condition (Tennie et al. 2010), and in another study, 5 of 58 great apes solved the problem in the baseline condition (Hanus et al. 2011). The results from these experiments contrast with an earlier report of high task solution rates among captive orangutans (Mendes et al. 2007).
After a solution using water to displace the object was demonstrated by a human Demonstrator, one out of seven capuchins solved the task using an object. This low solution rate can be compared to the proportion of chimpanzees who solved the problem upon seeing the solution demonstrated (Tennie et al. 2010); eight of 27 (30%) were able to solve the task.
In Experiment 1, capuchins were initially interested in the task, as evidenced by their manipulative actions directed toward the tube, including biting, slapping and insertion of their fingers in unsuccessful attempts to retrieve the marshmallow. Despite the failure of these actions to retrieve the marshmallow, no subjects attempted alternative behaviour. Instead, the capuchins lost interest in the task as evidenced by a decrease in tube-directed behavior in trial 4 for the dry trials. No capuchin attempted to use any of the objects in the enclosure as a tool to aid in retrieving the marshmallow. In some ways this was surprising, as they had previously engaged in a dipping tool-use task (Phillips and Thompson 2013).
Although early studies found little evidence of social learning or imitation in capuchins (reviewed in Fragaszy & Visalberghi (2004)), recent studies have presented a more complex picture. Several publications have reported evidence of possible emulative behavior in capuchins. This includes both human-reared (Custance et al. 1999; Fredman and Whiten 2008) and mother-reared (Dindo et al. 2008; Fredman and Whiten 2008) individuals. Tasks on which capuchins have been found to emulate include problem boxes (Custance et al. 1999; Dindo et al. 2008) and a problem box-tool combination (Fredman and Whiten 2008). It is clear that at least some capuchins are able to engage in moderate fidelity copying. The reasons the capuchins generally failed to do so here may include physical (difficulty of getting water into the tube, distance from water source) or cognitive (lack of understanding of relationship of objects) barriers.
Additionally, functional fixedness via individual learning (Hanus et al. 2011) may explain the observed behaviour. Individuals may have been cognitively blocked from applying their previous experience with using objects in this different context. That is, they may be unable to see past how an object has been previously used, to use the object in a new way. This was seen in the study with chimpanzees and the floating peanut task (Hanus et al. 2011); after provision of a novel water dispenser chimpanzees were more likely to solve the problem. The subadult female capuchin in Experiment 2 was able to overcome potential cognitive blocking to solve the problem in an unusual way by using straw. When previously tested on a dipping tool-use task (Phillips and Thompson 2013), capuchins (including this female) solved the problem by selecting a single piece of straw, and using fine motor skill to insert it into a small-diameter hole to retrieve applesauce from a tube. In solving the current floating object problem, the successful subadult female capuchin inserted a wad of straw into the tube (almost as if to use the wad as a sponge), pushing it down to the bottom of the tube before retrieving the wad slowly.
Acknowledgments
Funding: This study was partialy funded by the National Institutes of Health (R15NS070717).
Footnotes
Conflict of Interest: The authors declare that they have no conflict of interest.
Ethical Approval: All applicable international, national, and institutional guidelines for the care and use of animals were followed. All procedures performed in the study involving animals were in accordance with the ethical standards of the institution or practice at which the studies were conducted.
References
- Bird CD, Emery NJ. Insightful problem solving and creative tool modification by captive nontool-using rooks. Proceedings of the National Academy of Sciences USA. 2009;106:10370–10375. doi: 10.1073/pnas.0901008106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonnie KE, De Waal FBM. Copying without rewards: socially influenced foraging decisions among broan capuchin monkeys. Animal Cognition. 2007;10:283–292. doi: 10.1007/s10071-006-0069-9. [DOI] [PubMed] [Google Scholar]
- Custance D, Whiten A, Fredman T. Social learning of an artificial fruit task in capuchin monkeys (Cebus apella) Journal of Comparative Psychology. 1999;113:12–23. [Google Scholar]
- Dindo M, Thierry B, Whiten A. Social diffusion of novel foraging methods in brown capuchin monkeys (Cebus apella) Proceedings of the Royal Society B. 2008;275:187–193. doi: 10.1098/rspb.2007.1318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fragaszy D, Visalberghi E. Socially biased learning in monkeys. Learning and Behavior. 2004;32:24–35. doi: 10.3758/bf03196004. [DOI] [PubMed] [Google Scholar]
- Fragaszy DM. How non-human primates use their hands. In: Connolly KJ, editor. The Psychobiology of the Hand. Mac Keith; London: 1998. pp. 77–96. [Google Scholar]
- Fredman T, Whiten A. Observational learning from tool using models by human-reared and mother-reared capuchin monkeys (Cebus apella) Animal Cognition. 2008;11:295–309. doi: 10.1007/s10071-007-0117-0. [DOI] [PubMed] [Google Scholar]
- Hanus D, Mendes N, Tennie C, Call J. Comparing the performances of apes (Gorilla gorilla, Pan troglodytes, Pongo pygmaeus) and human children (Homo sapiens) in the floating peanut task. PLoS One. 2011;6:e19555. doi: 10.1371/journal.pone.0019555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heinrich B. Testing insight in ravens. In: Heyes C, Huber L, editors. The Evolution of Cognition. MIT Press; Cambridge, MA: 2000. pp. 289–305. [Google Scholar]
- Horner V, Whiten A. Causal knowledge and imitation/emulation switching in chimpanzees (Pan troglodytes) and children (Homo sapiens) Animal Cognition. 2005;8:164–181. doi: 10.1007/s10071-004-0239-6. [DOI] [PubMed] [Google Scholar]
- Izawa K, Mizuno A. Palm-fruit cracking behavior of wild black-capped capuchins (Cebus apella) Primates. 1977;18:773–792. [Google Scholar]
- Köhler W. The Mentality of Apes. 2. Kegan Paul, Trench, Trubner & Co., Ltd; London: 1931. [Google Scholar]
- Lemon RN. Descending pathways in motor control. Annual Review of Neuroscience. 2008;31:195–218. doi: 10.1146/annurev.neuro.31.060407.125547. [DOI] [PubMed] [Google Scholar]
- Lynch Alfaro JW, de Sousa e Silva J, Jr, Rylands AB. How different are robust and gracile capuchin monkeys? An argument for the use of Sapajus and Cebus. American Journal of Primatology. 2012;74:273–286. doi: 10.1002/ajp.22007. [DOI] [PubMed] [Google Scholar]
- Mendes N, Hanus D, Call J. Raising the level: orangutans use water as a tool. Biology Letters. 2007;22:453–455. doi: 10.1098/rsbl.2007.0198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen M. Young children's imitative and innovative behaviour on the floating object task. Infant and Child Development. 2013;22:44–52. [Google Scholar]
- Padberg J, et al. Parallel evolution of cortical areas involved in skilled hand use. J Neurosci. 2007;27:10106–10115. doi: 10.1523/JNEUROSCI.2632-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panger MA, Perry S, Rose L, Gros-Louis J, Vogel E, MacKinnon KC, Baker M. Cross-site differences in foraging behaviour of white-faced capuchins (Cebus capucinus) American Journal of Primatology. 2002;119:52–66. doi: 10.1002/ajpa.10103. [DOI] [PubMed] [Google Scholar]
- Perry S, et al. Social conventions in wild white-faced capuchin monkeys: evidence for traditions in a neotropical primate. Current Anthropology. 2003;44:241–268. [Google Scholar]
- Phillips KA. Tool use in wild capuchin monkeys (Cebus albifrons trinitatis) American Journal of Primatology. 1998;46:259–261. doi: 10.1002/(SICI)1098-2345(1998)46:3<259::AID-AJP6>3.0.CO;2-R. [DOI] [PubMed] [Google Scholar]
- Phillips KA, Subiaul F, Sherwood CC. Curious monkeys have greater gray matter density in the precuneus. Neuroscience Letters. 2012;518:172–175. doi: 10.1016/j.neulet.2012.05.004. [DOI] [PubMed] [Google Scholar]
- Phillips KA, Thompson CR. Hand preference for tool-use in capuchin monkeys (Cebus apella) is associated with asymmetry of the primary motor cortex. American Journal of Primatology. 2013;75:435–440. doi: 10.1002/ajp.22079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rilling JK, Insel T. The primate neocortex in comparative perspective using magnetic resonance imaging. Journal of Human Evolution. 1999;37:191–223. doi: 10.1006/jhev.1999.0313. [DOI] [PubMed] [Google Scholar]
- Spinozzi G, Lagana T, Truppa V. Hand use by tufted capuchins (Cebus apella) to extract a small ood item from a tube: digit movements, hand preference, and performance. American Journal of Primatology. 2007;69:336–352. doi: 10.1002/ajp.20352. [DOI] [PubMed] [Google Scholar]
- Tennie C, Call J, Tomasello M. Evidence for emulation in chimpanzees in social settings using the floating peanut task. PLoS One. 2010;5:e10544. doi: 10.1371/journal.pone.0010544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorpe WH. Learning and Instinct in Animals. 2. Methuen & Co.; London: 1963. [Google Scholar]
- Tolman EC, Honzik CH. “Insight” in rats. Vol. 4. University of California Publications in Psychology; 1930. pp. 215–232. [Google Scholar]
- van Schaik CP, et al. Orangutan cultures and the evolution of material culture. Science. 2003;299:102–105. doi: 10.1126/science.1078004. [DOI] [PubMed] [Google Scholar]
- Visalberghi E. Tool use in Cebus. Folia Primatologica. 1990;54:146–154. doi: 10.1159/000156438. [DOI] [PubMed] [Google Scholar]
- Whiten A, et al. Cultures in chimpanzees. Nature. 1999;399:682–685. doi: 10.1038/21415. [DOI] [PubMed] [Google Scholar]
- Whiten A, Ham R. On the nature and evolution of imitation in the animal kingdom: reappraisal of a centry of research. Advances in the Study of Behavior. 1992;21:239–283. [Google Scholar]