

Abstract
Background
Clostridium pasteurianum as an emerging new microbial cell factory can produce both n-butanol (BuOH) and 1,3-propanediol (1,3-PDO), and the pattern of product formation changes significantly with the composition of the culture medium. Among others iron content in the medium was shown to strongly affect the products selectivity. However, the mechanism behind this metabolic regulation is still unclear. For a better understanding of such metabolic regulation and for process optimization, we carried out fermentation experiments under either iron excess or iron limitation conditions, and performed metabolic, stoichiometric and proteomic analyses.
Results
1,3-PDO is most effectively produced under iron limited condition (Fe−), whereas 1,3-PDO and BuOH were both produced under iron rich condition (Fe+). With increased iron availability the BuOH/1,3-PDO ratio increased significantly from 0.27 mol/mol (at Fe−) to 1.4 mol/mol (at Fe+). Additionally, hydrogen production was enhanced significantly under Fe+ condition. Proteomic analysis revealed differentiated expression of many proteins including several ones of the central carbon metabolic pathway. Among others, pyruvate: ferredoxin oxidoreductase, hydrogenases, and several electron transfer flavoproteins was found to be strongly up-regulated under Fe+ condition, pointing to their strong involvement in the regeneration of the oxidized form of ferredoxin, and consequently their influences on the product selectivity in C. pasteurianum. Of particular significance is the finding that H2 formation in C. pasteurianum is coupled to the ferredoxin-dependent butyryl-CoA dehydrogenase catalyzed reaction, which significantly affects the redox balance and thus the product selectivity.
Conclusions
The metabolic, stoichiometric and proteomic results clearly show the key roles of hydrogenases and ferredoxins dependent reactions in determining the internal redox balance and hence product selectivity. Not only the NADH pool but also the regulation of the ferredoxin pool could explain such product variation under different iron conditions.
Keywords: n-Butanol; 1,3-Propanediol; C. pasteurianum; Proteomics; Product selectivity; Metabolic analysis
Background
Clostridium pasteurianum is an emerging and promising microbial cell factory for the production of chemicals and fuels because of some unique features, e.g. utilization of a wide range of substrates [1–3], production of a wide spectrum of products [4, 5] and robust growth in simple media even under unsterile conditions [6]. Recently, C. pasteurianum was shown to accept electrons from the cathode by direct electron transfer [7], which make it an attractive candidate for new bioelectrical systems. Therefore, C. pasteurianum has received considerable interests for the production of chemicals and fuels such as 1,3-propanediol (1,3-PDO) and n-butanol (BuOH), which represents attractive bioprocesses for the use of renewable resources, like biodiesel-derived glycerol or glucose from biomass hydrolysates [8–14]. In such bioprocesses, several other fermentation products like gases (carbon dioxide and hydrogen), ethanol as well as acetic, butyric and lactic acid are produced [2, 4, 15], in addition to 1,3-PDO and BuOH. Even though the formation of organic acids is inevitable for the maintenance of the intracellular redox balance, it represents a loss of carbons at the expense of the target products. Moreover, the resulting product distribution, especially the selectivity of either 1,3-PDO or BuOH, is mainly influenced by the cultivation conditions and/or media supplements. For instances, several studies analyzed the effect of pH [16], inoculum conditions [17], supplementations of yeast extracts and ammonia [18], or acetic and butyric acid [2, 19], or phosphate and iron [18, 20]. Among others, iron seems to have extensive effects, since its absence lead to a strongly reduced BuOH formation [16, 20]. In real fermentation processes, especially under conditions more relevant to industrial applications, with raw substrates and high concentrations of products, the product selectivity and yield often strongly fluctuate and are hardly reproducible. The underlying mechanism(s) of selectivity and regulation of intracellular metabolic pathways are still unclear, even though a combined effect of many iron-related enzymes has been assumed [4]. Indeed, several iron containing enzymes are involved in clostridia metabolism, e.g. nitrogenases, ferredoxin coupled enzymes, and alcohol dehydrogenases. These enzymes play key roles in the maintenance of intracellular redox balance and a limited functionality of them, e.g. due to iron limitation, will be reflected by a metabolic shift and thus change of product selectivity.
In this work, the variations of product selectivity and the underlying mechanisms of pathway regulation in C. pasteurianum DSMZ 525 grown on glycerol under varied iron availability are studied with an integrated systems biology approach, particularly with stoichiometric, kinetic and proteomic analyses.
Methods
Microorganism, medium and cultivation
Clostridium pasteurianum DSMZ 525 was routinely maintained as cryoculture at −80 °C in Reinforced Clostridial Medium (RCM, Oxoid Deutschland GmbH) containing 20% (v/v) glycerin. The cryoculture was used for the pre-culture carried out in anaerobic bottles with RCM medium at 35 °C and pH 7 without shaking. The RCM contained 1 mg/L resazurin (7-hydroxy-10-oxidophenoxazin-10-ium-3-one) as a redox indicator for anaerobiosis and 2 g/L CaCO3 as pH-buffering agent. After 24 h this pre-culture was used as inoculum for bioreactor fermentation. The bioreactor medium contained the following ingredients in 1 L of distilled water (modified from [4]): glycerol, 80 g; yeast extract, 1 g; K2HPO4, 0.5 g; KH2PO4, 0.5 g; MgSO4·7H20, 0.2 g; (NH4)2SO4, 5 g; CaCl2 2H2O, 0.02 g; cysteine-HCl, 0.5 g; resarzurin, 0.005 g; trace element solution SL-7 (DSMZ), 2 mL. Iron concentrations were varied in the bioreactor medium as follows: Iron excess (Fe+) condition means the addition of 10 mg/L FeSO4·7H2O (2 mg Fe2+/L) into the medium and iron limitation (Fe−) condition means no iron addition. Iron originally present in the pre-culture (0.07 mg Fe2+/L) and those present in the yeast extract (up to 0.05 mg Fe2+/L) were the sole iron sources in the Fe− cultivations. Cultivations were run at 35 °C, pH 6 and 500 rpm agitation in a stirred tank bioreactor (Bioengineering) with a working volume of 1.2 L. During the fermentation pH was maintained at 6 with 5 M KOH. To achieve anaerobic condition prior to the inoculation, the autoclaved medium was sparged with sterile O2-free N2. The experiments were performed in duplicates. Total volume of the effluent fermentation gas was determined with a Milli-Gascounter (Dr.-Ing. Ritter Apparatebau GmbH & Co. KG), and its composition was measured with the mass spectrometer OmniStar 300 (Balzer Instruments/Pfeiffer Vacuum GmbH). The MS took samples in an interval of 0.5 mL/min for the concentration analysis of H2, CO2, O2, N2 and Ar.
Analytical methods and calculations
The optical density of cell suspension was measured turbidometrically at 600 nm and correlated with cell dry weight: biomass BM (g/L) = OD600 × 0.336. The specific growth rate µ (h−1) was determined from biomass data (smoothed using the software Origin 8.5.1 G SR1, OriginLab Corporation, Northampton, USA) according to Eq. 1, where x1 and x2 are the concentrations of biomass (g/L) at the times t1 and t2, respectively. The substrate and product titers in the supernatant were analyzed via HPLC equipped with a refractive index detector and an ultraviolet detector. HPLC was performed on an Aminex HPX-87H column (300 × 7.8 mm) at 60 °C, with 0.005 M H2SO4 as mobile phase at a flow rate of 0.6 mL/min.
For the measurement of 3-HPA, the method described by Oehmke and Zeng [21] was used, in which 3-HPA is converted into acrolein and the concentration of acrolein is determined spectrometrically by external calibration. Briefly, 100 µL of cell free culture supernatant were mixed with 200 µL of HCl (37%) and 50 µL of tryptophan solution in a cooled 96 well plate. The tryptophan solution consisted of 10 mM DL-tryptophan, 0.05 M HCl and 24 mM toluene. After 40 min incubation at 37 °C, the absorbance of the mixtures were determined with a Multiskan® Spectrum plate reader (Thermo Fisher Scientific) at 560 nm.
The yield coefficient (Y) for either product or substrate (i) was calculated according to Eq. 2. Based on the stoichiometric equations for glycerol utilization in C. pasteurianum [11], carbon and redox recovery were calculated according to Eqs. 3 and 4, respectively. Here C [−] is the number of carbon atoms in the products and substrate, c is the concentration of products in (mmol/L) and biomass (BM) in (g/L).
| 1 |
| 2 |
| 3 |
| 4 |
Comparative proteomic analysis
Samples for proteomics were taken in the exponential growth phase and stationary phase during parallel fermentations of C. pasteurianum DSMZ 525 under iron excess and iron limited conditions. The detailed methodical procedure for comparative proteomic analysis was previously described by Sabra et al. [11].
Results and discussion
Effects of iron availability on the growth and product formation of C. pasteurianum
Different concentrations of iron have been reported for the optimization of butanol or 1,3-PDO formation using C. pasteurianum [2, 18, 20]. Using a fractional factorial experimental design, Moon et al. used 60 mg/L FeSO4·7H2O for optimum butanol formation in C. pasteurianum in serum anaerobic bottle experiments, while no iron sulphate was supplemented for a better 1,3-PDO production [18]. In controlled bioreactor, we have found that 10 mg/L FeSO4·7H2O is enough to support a similar butanol productivities (0.9 g/L×h) and an almost doubled butanol concentration (21 g/L butanol) by the same strain [2]. Therefore, in the current investigation, 0 and 10 mg/L FeSO4·7H2O were chosen, respectively, to describe the growth and product formation under iron limited and iron excess conditions in our glycerol fermentation. The same pre-culture was used to inoculate two bioreactors containing the growth medium supplemented either with or without 10 mg/L FeSO4·7H2O (hereinafter termed as iron excess (Fe+) condition or iron limitation (Fe−) condition, respectively). One of the main differences observed was the relatively shorter lag phase under Fe− condition, which was accompanied by an early growth cessation (Fig. 1a). With excess iron in the medium, a higher biomass production with a maximum concentration of 5.1 ± 0.09 g/L and µmax of 0.31 ± 0.01 h−1 were reached, whereas under iron limitation condition only a biomass concentration of 3.2 ± 0.01 g/L and a µmax of 0.23 ± 0.01 h−1 could be achieved (Table 1). The cessation of growth under iron limitation was obviously not due to butanol toxicity, as the highest titer of BuOH reached did not exceed 3.7 g/L (Table 1), which was lower than the toxic concentration level of BuOH for C. pasteurianum (>5 g/L [11]). Depletion of the intracellular iron pool and/or the accumulation of 3-hydroxypropionaldehyde (3-HPA), a very toxic intermediate in the formation of 1,3-PDO [22, 23], may cause the relatively earlier growth cessation under Fe− condition. As shown in Fig. 1b, under Fe+ condition the 3-HPA concentration did not exceed 8 mg/L, whereas under Fe− condition up to 30 mg/L 3-HPA were produced. In this time range of relatively high concentrations of 3-HPA a growth cessation was observed. Indeed, it has been reported that the growth of vegetative cells of C. tyrobutyricum was completely inhibited at 38 mg/L externally added 3-HPA [24].
Fig. 1.
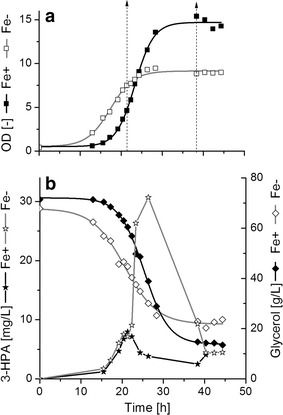
a Cell growth behavior and b 3-HPA formation and glycerol consumption in cultivation of C. pasteurianum DSMZ 525 under iron excess (Fe+) and iron limitation (Fe−) conditions. Arrows indicate time points of sampling for proteomic analysis
Table 1.
Product formation during the growth of C. pasteurianum DSMZ 525 under iron excess (Fe+) and iron limitation (Fe−) conditions. Fermentations were performed in duplicates
| Biomass | 1,3-PDO | BuOH | EtOH | Acetate | Butyrate | Lactate | Formate | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fe+ | Fe− | Fe+ | Fe− | Fe+ | Fe− | Fe+ | Fe− | Fe+ | Fe− | Fe+ | Fe− | Fe+ | Fe− | Fe+ | Fe− | |
| Final titer (g/L) | 5.1 ± 0.09 | 3.2 ± 0.01 | 9.4 ± 1.44 | 16.6 ± 2.26 | 12.3 ± 0.06 | 4.4 ± 0.70 | 1.0 ± 0.12 | 0.9 ± 0.60 | 0.8 ± 0.01 | 1.5 ± 0.02 | 1.7 ± 0.45 | 2.8 ± 0.13 | 0.1 ± 0.00 | 7.5 ± 2.58 | 0.5 ± 0.07 | 1.1 ± 0.11 |
| Production rate (g/L*h) at exponent-ial growth phase | 0.39 ± 0.00 | 0.16 ± 0.03 | 0.66 ± 0.17 | 0.74 ± 0.02 | 0.53 ± 0.11 | 0.16 ± 0.04 | 0.06 ± 0.02 | 0.03 ± 0.003 | 0.04 ± 0.004 | 0.07 ± 0.03 | 0.14 ± 0.02 | 0.13 ± 0.01 | 0.007 ± 0.003 | 0.28 ± 0.02 | 0.03 ± 0.01 | 0.04 ± 0.02 |
| Y S/X | Y P/X | |||||||||||||||
| Y i/X (g/g BM) | 9.6 ± 0.3 | 13.9 ± 1.2 | 1.16 ± 0.09 | 4.40 ± 0.52 | 1.88 ± 0.25 | 0.97 ± 0.13 | 0.15 ± 0.06 | 0.17 ± 0.13 | 0.10 ± 0.01 | 0.39 ± 0.09 | 0.39 ± 0.08 | 0.80 ± 0.17 | 0.02 ± 0.00 | 1.64 ± 0.37 | 0.08 ± 0.02 | 0.25 ± 0.05 |
Figure 2 shows the formation of fermentation products in C. pasteurianum DSMZ 525 under iron excess and iron limitation conditions. With higher iron concentration 12 g/L of BuOH and 11 g/L of 1,3-PDO were produced. In comparison, at iron limitation significantly less BuOH, i.e. 3.7 g/L, was formed, accompanied with the formation of 14.5 g/L of 1,3-PDO. In fact, the molar ratio of BuOH to 1,3-PDO decreased from 1.34 mol/mol under Fe+ condition to 0.27 mol/mol under Fe− condition. The yield of 1,3-PDO per biomass increased nearly 4 times from 1.16 ± 0.09 g/g at Fe+ to 4.4 ± 0.52 g/g at Fe−, whereas the specific yield of BuOH was halved from 1.88 ± 0.25 g/g at Fe+ to 0.97 ± 0.13 g/g at Fe−. Next to this, the acid formation changed remarkably, especially the lactate production, which was shown to increase significantly in the Fe− culture (Table 1). The specific lactate yield increased significantly from 0.02 ± 0.0 g/g in the Fe+ culture to 1.6 ± 0.4 g/g in the Fe− culture. Also acetate and butyrate yield increased under Fe− condition, but to less extent than that of lactate (Table 1). The reason(s) for these dramatic changes of metabolism are not clear yet, but of fundamental importance for the development of C. pasteurianum as an emerging microbial cell factory for the production of chemicals and fuels. Therefore in the following redox regulation and comparative proteomic analysis were performed.
Fig. 2.
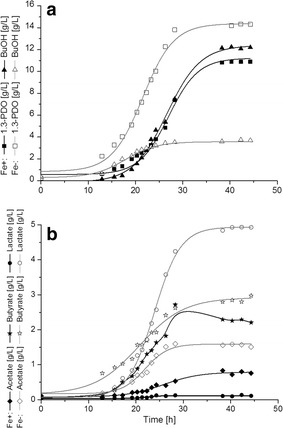
1,3-PDO and BuOH formation (a) and acid formation (b) in C. pasteurianum DSMZ 525 during glycerol fermentation under iron excess (Fe+) and iron limitation (Fe−) conditions
Redox regulation and H2 production in C. pasteurianum DSMZ 525
For the growth and metabolism of C. pasteurianum, particularly when growing on a more reduced substrate like glycerol, the maintenance of intracellular redox balance is crucial. The shift of metabolism under conditions of iron excess and limitation shown above is postulated to be strongly related to the redox regulation which is addressed below first from a stoichiometric point of view.
To check the stoichiometry and consistency of the fermentation data, fermentation balance analysis was first done. A very good consistency in carbon recovery was observed for the fermentations. The carbon recovered as fermentation products represented approximately 98% of the carbon source consumed. On the other hand, the calculated recovery of the reducing equivalents according to Eq. 4 reached 91% at Fe+ and 94% at Fe−, indicating a lower consistency in reducing equivalent recovery according to the assumed pathways of redox regulation. C. pasteurianum contains ferredoxin-dependent hydrogenases, which catalyze the re-oxidation of reduced ferredoxin with the formation of H2. Reduced ferredoxins are generally formed in the enzymatic step of forming acetyl-CoA from pyruvate catalyzed by pyruvate: ferredoxin oxidoreductase (PFOR). Hence, under the assumption that the formation of one mole acetyl-CoA from pyruvate is accompanied with the formation of one mole H2, the theoretical H2 production would be calculated according to Eq. 5, where q is the formation rate of each compound (mmol/g×h):
| 5 |
In repeated fermentations to those shown in Fig. 1 under similar conditions we measured the evolution of CO2 and H2 in effluent gas. The results are given in Fig. 3a. Under Fe+ conditions a cumulative amount of 452 mmol/L H2 and 399 mmol/L CO2 were produced, compared to 245 mmol/L H2 and 177 mmol/L CO2 produced under Fe− condition. Referred to the biomass formed, H2 and CO2 production increased significantly from 63 (±3.3) and 49 (±3.1) mmol/g biomass at Fe− condition to 91 (±1.9) and 82 (±3.9) mmol/g biomass at Fe+ condition, respectively. Interestingly, the theoretically calculated H2 production values were lower than the measured ones (Fig. 3b), particularly under Fe+ conditions. Obviously, the re-oxidation of reduced ferredoxin generated in the enzymatic step catalyzed by PFOR was not the only source of hydrogen formation. Similar behavior was also noticed previously in cultures of C. butyricum or Klebsiella pneumoniae [25, 26].
Fig. 3.
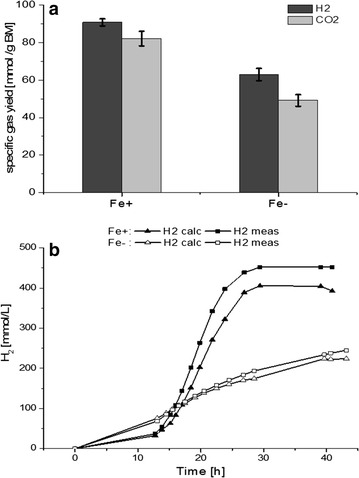
a Measured cumulative hydrogen and carbon dioxide production and b calculated (calc) vs. measured (meas) hydrogen production under iron excess (Fe+) and iron limitation (Fe−) conditions in C. pasteurianum DSMZ 525 cultures
It is known that butyryl-CoA is generally formed from crotonyl-CoA by the NADH dependent trans-2-enoyl-CoA reductase (Ter) enzyme (Eq. 6) [27].
| 6 |
But recently, Buckel and Thauer [28] proposed a new indirect route of H2 formation in C. pasteurianum from NADH and ferredoxin in two steps, catalyzed successively by the ferredoxin-dependent butyryl-CoA dehydrogenase/electron transferring flavoprotein complex (BCdH-ETF) (Eq. 7) and a hydrogenase (Eq. 8).
| 7 |
| 8 |
Since the measured H2 production values were significantly higher than the theoretically calculated ones based on Eqs. 7 and 8 (see Fig. 3b), it is reasonable to assume that in C. pasteurianum DSMZ 525, BCdH-ETF together with Ter is actively involved in the step of converting crotonyl-CoA to butyryl-CoA, giving rise to an additional source of H2 formation. Thus, with this new suggested butyryl-CoA formation route one mole NADH2 is additionally required for the formation of one mol butanol or one mol butyrate, accompanied with the formation of one mole more H2, in addition to the H2 formation counted in Eq. 5. Consequently, the calculation of reducing equivalent recovery should be modified as follows (Eq. 9), by also taking into account the difference of calculated and measured H2 values (c∆H2), representing the additionally consumed NADH2:
| 9 |
Using Eq. 9, a more satisfying reducing equivalent recovery of 105% under Fe+ condition and 100% under Fe− condition was obtained, giving a strong support for the involvement of the BCdH-ETF complex. Particularly, the results from Fe+ condition are in agreement with the corrected Eq. 9, where more BuOH and hydrogen were produced, and the deviation between the calculated and measured H2 was higher. However, this is in contrary to what was reported for C. acetobutylicum. For a more effective butanol production, a lower hydrogenase activity and hydrogen production was favored in C. acetobutylicum [29]. To shed more light on the mechanisms underlying the effect of iron on the regulation of glycerol metabolism in C. pasteurianum, comparative proteomic studies were performed as described below.
Comparative proteomic analysis of the iron effect
For proteomic analysis of the effects of iron concentration on the metabolism of C. pasteurianum, samples were taken from the two bioreactors in the exponential growth phase (termed as Fe+ early and Fe− early, respectively) and the stationary growth phase (Fe+ late and Fe− late, respectively) (Fig. 1). Each sample was analyzed in triplicates. After 2-DE separation of the intracellular proteins, protein spots showing statistically significant changes between Fe+ early and Fe− early, Fe+ late and Fe− late, Fe+ early and Fe+ late, as well as Fe− early and Fe− late were further identified by LC–MS/MS. Proteins which were identified as single protein present in a spot on the 2-D gels are summarized in Table 2 according to their functional categories and accession numbers, together with the information of their expression changes. The existence of more than one values of fold change for a single protein indicates that this protein appeared as multiple spots on the 2-D gels.
Table 2.
Proteins showing significant changed expression levels, compared between iron excess (Fe+) and iron limitation (Fe−) conditions, as well as between the exponential growth phase (early) and the stationary phase (late)
| Accession no. | Protein name | Cluster of orthologous groups (COG) | Conserved protein domain | Spot no. | Fold change iron-related | Fold change growth-related | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Early phase higher at | Late phase higher at | Fe+ higher at | Fe− higher at | |||||||||
| Fe− | Fe+ | Fe− | Fe+ | Early | Late | Early | Late | |||||
| Amino acid transport and metabolism | ||||||||||||
| F502_05097 | Amino peptidase 1 | COG1362 | Lap4 | 298 | 1.9 | 1.7 | ||||||
| F502_05412 | Carbamoyl phosphate synthase large subunit | COG0458 | CarB | 89 | 1.8 | 1.9 | ||||||
| F502_07028 | Cysteine synthase a | COG0031 | CysK | 449 | 1.8 | 2.1 | ||||||
| F502_17572 | Glutamine synthetase type III | COG3968 | GlnA3 | 187 | 1.6 | 1.9 | ||||||
| 190 | 1.6 | 1.6 | ||||||||||
| F502_18676 | Threonine synthase | COG0498 | ThrC | 312 | 1.6 | 2.2 | ||||||
| Carbohydrate transport and metabolism | ||||||||||||
| F502_03412 | Propanediol dehydratase small | COG4910 | PduE | 594 | 1.9 | 1.9 | ||||||
| F502_07638 | Subunit flavodoxin | COG0716 | FldA | 594 | ||||||||
| F502_03417 | Glycerol dehydratase reactivation factor large subunit | No COG | 220 | 2.1 | ||||||||
| F502_03937 | Glycogen synthase | COG0297 | GlgA | 329 | 2.7 | 2.3 | ||||||
| F502_06067 | Enolase | COG0148 | Eno | 695 | 4.1 | 4.0 | 2.0 | 2.0 | ||||
| 696 | 4.8 | 1.8 | 2.5 | |||||||||
| F502_06077 | Triosephosphate isomerase | COG0149 | TpiA | 491 | 1.5 | 2.6 | ||||||
| F502_06087 | Glyceraldehyde 3-phosphate dehydrogenase | COG0057 | GapA | 409 | 1.6 | 2.4 | ||||||
| F502_07098 | Glycoside hydrolase | COG1543 | 277 | 2.0 | 1.8 | |||||||
| F502_12758 | Dihydroxyacetone kinase | COG2376 | DAK1 | 158 | 1.7 | 2.0 | 1.7 | 2.1 | ||||
| 179 | 1.7 | 1.7 | 2.2 | 2.3 | ||||||||
| Cell cycle control/cell division | ||||||||||||
| F502_08238 | Cell division protein | COG3599 | DivIVA | 503 | 2.7 | 2.0 | ||||||
| Cell wall/membrane/envelope biogenesis | ||||||||||||
| F502_00655 | Peptidoglycan-binding protein | COG1388 | LysM | 233 | 7.9 | 6.1 | 1.6 | |||||
| F502_01965 | Spore coat protein Frelated protein | COG5577 | CotF | 549 | 1.5 | 2.1 | 1.6 | |||||
| Coenzyme transport and metabolism | ||||||||||||
| F502_07578 | Pyridoxal biosynthesis lyase | COG0214 | PdxS | 460 | 2.5 | 2.2 | ||||||
| Energy production and conversion | ||||||||||||
| F502_05017 | NifU-related domain containing protein | COG0822 | IscU | 519 | 0.4 | 2.4 | 0.6 | 1.6 | 1.3 | 0.8 | 0.9 | 1.1 |
| 520 | 3.7 | 2.9 | 1.9 | 1.5 | ||||||||
| F502_06282 | Electron transfer flavoprotein subunit alpha | COG2025 | FixB | 440 | 1.5 | |||||||
| F502_06287 | Electron transfer flavoprotein subunit alpha/beta-like protein | COG2086 | FixA | 487 | 1.7 | |||||||
| F502_06447 | Bifunctional acetaldehyde-CoA/alcohol dehydrogenase | COG1012 | AdhE | 119 | 2.1 | 3.7 | 1.6 | |||||
| 715 | 1.5 | 2.9 | 5.3 | |||||||||
| 717 | 1.6 | 3.0 | 4.7 | |||||||||
| F502_07493 | Nitroreductase | COG0778 | NfnB | 537 | 2.3 | 2.9 | 1.8 | |||||
| F502_07643 | Pyruvate: ferredoxin (flavodoxin) oxidoreductase. homodimeric | COG0674 | PorA | 70 | 2.4 | 2.4 | 1.5 | |||||
| 75 | 2.4 | 0.6 | 1.8 | |||||||||
| 76 | 2.7 | 1.9 | ||||||||||
| 77 | 1.5 | 2.3 | ||||||||||
| 87 | 2.0 | 2.8 | 1.5 | |||||||||
| 90 | 2.2 | 2.8 | 1.6 | 2.0 | ||||||||
| F502_07648 | Pyruvate: ferredoxin oxidoreductase | COG0674 | PorA | 40 | 2.3 | 1.6 | ||||||
| 57 | 2.3 | 2.1 | ||||||||||
| 58 | 1.5 | 2.4 | 2.1 | |||||||||
| 59 | 2.1 | 2.8 | ||||||||||
| F502_09238 | Rubredoxin/flavodoxin/oxidoreductase | COG0426 | NorV | 131 | 1.7 | 2.0 | ||||||
| 730 | 2.0 | 1.6 | ||||||||||
| F502_09488 | Hydratase (aconitase A) | COG1048 | AcnA | 156 | 2.4 | 2.5 | 1.9 | 2.0 | ||||
| F502_11871 | Butyrate kinase | COG3426 | Buk | 383 | 1.6 | 1.7 | ||||||
| F502_11976 | Pyruvate carboxylase | COG1038 | PycA | 71 | 1.5 | 2.7 | 1.7 | |||||
| F502_12091 | F0F1 ATP synthase subunit beta | COG0055 | AtpD | 645 | 2.1 | 1.9 | ||||||
| 646 | 2.2 | 2.4 | ||||||||||
| F502_12878 | Desulfo ferrodoxin | COG2033 | SorL | 611 | 2.2 | 2.8 | ||||||
| F502_13493 | Flavodoxin | COG0716 | FldA | 575 | 14.3 | 8.0 | 1.6 | |||||
| F502_14390 | [Fe] hydrogenase | COG4624 | Nar1 PurB | 303 | 2.1 | 2.5 | ||||||
| F502_04707 | Adenylosuccinate lyase | COG0015 | 303 | |||||||||
| F502_15080 | Rubrerythrin | COG1592 | YotD | 558 | 2.2 | 4.5 | 2.0 | |||||
| F502_16610 | Glycolate oxidase | COG0277 | GlcD | 315 | 1.8 | 2.3 | 1.9 | |||||
| F502_18287 | Hydrogenase-1 | COG1034 | NuoG | 223 | 4.5 | 1.6 | 2.3 | |||||
| 224 | 5.1 | 1.9 | 3.6 | |||||||||
| F502_18651 | NADP-dependent glyceraldehyde-3-phosphate dehydrogenase | COG1012 | AdhE | 736 | 2.7 | 1.5 | 2.1 | |||||
| F502_19556 | Formate acetyltransferase | COG1882 | PflD | 174 | 4.5 | 5.7 | ||||||
| 175 | 8.0 | 6.5 | ||||||||||
| 178 | 2.5 | 3.4 | ||||||||||
| 181 | 3.7 | 1.7 | 7.4 | |||||||||
| Function unknown/general function prediction only | ||||||||||||
| F502_02435 | Aldo/keto reductase | COG1453 | 362 | 2.6 | 2.9 | |||||||
| F502_05012 | Hypothetical protein (GGGtGRT protein) | No COG | 400 | 7.1 | 2.2 | 3.0 | ||||||
| F502_05962 | Hypothetical protein | No COG | 635 | 2.0 | 1.5 | |||||||
| F502_06682 | Hypothetical protein | COG2607 | 318 | 3.4 | 3.8 | 1.7 | 2.0 | |||||
| F502_15420 | Hypothetical protein | No COG | 597 | 1.7 | 2.8 | 1.5 | ||||||
| F502_16320 | Hypothetical protein | COG0393 | YbjQ | 637 | 2.3 | 2.0 | ||||||
| Lipid transport and metabolism | ||||||||||||
| F502_06297 | 3-Hydroxybutyryl-CoA dehydratase | COG1024 | CaiD | 472 | 2.1 | 2.2 | ||||||
| F502_10483 | Biotin carboxylase | COG0439 | AccC | 302 | 2.5 | 1.6 | 2.1 | 1.9 | ||||
| Nucleotide transport and metabolism | ||||||||||||
| F502_17300 | Bifunctional phosphoribosylaminoimidazolecarboxamide formyltransferase/IMP cyclohydrolase | COG0138 | PurH | 281 | 1.7 | 1.6 | ||||||
| Posttranslational modification/Protein turnover/Chaperones | ||||||||||||
| F502_03242 | Heat shock protein (molecular chaperone GrpE) | COG0576 | GrpE | 481 | 2.1 | 2.2 | ||||||
| F502_03247 | Molecular chaperone DnaK | COG0443 | Dank | 217 | 1.6 | 1.8 | ||||||
| F502_03987 | peptidase | COG1026 | Cym1 | 106 | 2.3 | 2.1 | ||||||
| 107 | 3.6 | 1.6 | 2.4 | |||||||||
| F502_05557 | ATP-dependent Clp protease ATP-binding subunit | COG0542 | ClpA | 165 | 1.8 | 2.1 | ||||||
| 168 | 1.9 | 1.7 | ||||||||||
| F502_06242 | Chaperonin | COG0459 | GroEL | 258 | 1.6 | 2.0 | ||||||
| F502_06247 | Co-chaperonin | COG0234 | GroES | 613 | 1.5 | 2.6 | ||||||
| F502_07608 | Thij/PfpI family protein | COG0693 | ThiJ | 553 | 1.8 | 1.6 | ||||||
| F502_10228 | Heat shock protein (molecular chaperone IbpA) | COG0071 | IbpA | 600 | 2.5 | 3.0 | 6.9 | |||||
| F502_15425 | Heat shock protein 90 | COG0326 | HtpG | 196 | 2.2 | 2.4 | ||||||
| 206 | 1.6 | 1.5 | ||||||||||
| F502_18446 | Clpb protein | COG0542 | ClpA | 138 | 1.8 | 1.5 | 2.8 | |||||
| 716 | 2.5 | 2.4 | 4.4 | |||||||||
| F502_18743 | ATPase with chaperone activity clpC. two ATP-binding domain protein | COG0542 | ClpA | 147 | 2.1 | 2.1 | 2.0 | 2.0 | ||||
| ClpA | 149 | 2.5 | 1.7 | 2.4 | 1.6 | |||||||
| Signal transduction/stress response/defense mechanism | ||||||||||||
| F502_04082 | GTP-binding protein | COG1217 | TypA | 47 | 1.7 | 1.8 | ||||||
| 148 | 1.7 | 2.4 | 2.1 | |||||||||
| F502_07703 | Chemotaxis histidine kinase. CheA (contains CheW-like adaptor domain) | COG0643 | CheA | 155 | 3.6 | 2.2 | 1.6 | |||||
| F502_10768 | Lipid hydroperoxide peroxidase | COG2077 | Tpx | 572 | 3.9 | 2.2 | 2.5 | |||||
| F502_13258 | CBS domain-containing protein | COG0517 | CBS | 606 | 4.5 | 4.8 | 1.7 | 1.9 | ||||
| F502_14770 | Serine protein kinase | COG2766 | PrkA | 193 | 4.7 | 1.5 | 3.4 | |||||
| 197 | 3.7 | 2.6 | 2.1 | |||||||||
| 200 | 6.2 | 1.9 | 3.6 | |||||||||
| 201 | 3.8 | 2.6 | ||||||||||
| F502_16565 | Nitrogen regulatory protein P-II |
COG0347 | GlnK | 638 | 3.1 | 1.8 | ||||||
| F502_17612 | Alkyl hydroperoxide reductase | COG0450 | AhpC | 567 | 5.3 | 2.9 | 2.4 | |||||
| F502_17637 | Spore coat protein | COG3546 | CotJC | 548 | 2.3 | 3.9 | 2.1 | |||||
| F502_18092 | Stage V sporulation protein T | COG2002 | AbrB | 544 | 6.1 | 4.5 | 1.7 | |||||
| Transcription/Defense mechanisms | ||||||||||||
| F502_12326 | Transcription accessory protein | COG2183 | Tex | 160 | 1.7 | 1.8 | 4.7 | 1.5 | ||||
| Translation/Ribosomal structure and biogenesis | ||||||||||||
| F502_04537 | 30S ribosomal protein S2 | COG0052 | RpsB | 497 | 2.1 | 2.1 | 2.7 | 2.7 | ||||
| 500 | 2.4 | 1.9 | ||||||||||
| F502_06817 | Ribosomal 5S rRNA E-loop binding protein Ctc/L25/TL5 | COG1825 | RplY | 504 | 3.5 | 3.9 | ||||||
| F502_12196 | Ribosome-associated protein Y (PSrp-1) | COG1544 | RaiA | 565 | 2.4 | 2.4 | ||||||
| F502_18808 | Elongation factor Tu | COG0050 | TufB | 327 | 1.7 | 1.5 | 1.5 | 1.4 | ||||
| F502_18833 | 50S ribosomal protein L1 | COG0081 | RplA | 681 | 2.4 | 1.5 | 2.0 | |||||
| F502_18948 | 50S ribosomal protein L5 | COG0094 | RplE | 752 | 2.1 | 1.8 | ||||||
| F502_18963 | 50S ribosomal protein L6 | COG0097 | RplF | 753 | 2.4 | 2.3 | ||||||
*1 COG: according to the annotation for C. pasteurianum DSMZ 525 by BioCyc database collection (http://www.biocyc.org/organism-summary?object=CPAS1262449)
*2 Conserved Protein Domain Family: according to the definition by NCBI Conserved Domains and Protein Classification (http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)
The pyruvate acetyl-CoA node: a focal point in the metabolism of C. pasteurianum
The conversion of pyruvate to acetyl-CoA linking glycolysis to TCA cycle is a fundamental metabolic step of living organisms in general. In anaerobes, pyruvate can be metabolized through a variety of pathways but it is often oxidized to CO2 and acetyl-CoA with the concomitant reduction of a low-potential redox protein, like ferredoxin or flavodoxin. The enzyme responsible for this oxidative decarboxylation of pyruvate in many anaerobic bacteria and archaea is pyruvate: ferredoxin oxidoreductase (PFOR). PFORs contain thiamin pyrophosphate (TPP) for the cleavage of carbon–carbon bonds next to a carbonyl group, as well as iron-sulfur clusters for electron transfer (see [30, 31] and references therein). For example, PFOR from C. pasteurianum W5 (ATCC 6013) was characterized to be an air-sensitive homodimer with each subunit containing eight iron atoms in two [4Fe–4S] clusters, for which pyruvate is the best substrate found among several α-ketoacids [30]. In the genome of C. pasteurianum DSMZ 525, three homologue enzymes of PFOR are present, namely two pyruvate ferredoxin oxidoreductases (F502_01955 and F502_07648), and one pyruvate:ferredoxin (flavodoxin) oxidoreductase (F502_07643) [32]. In this study, F502_07648 (termed as PFOR1) and F502_07643 (termed as PFOR2) were identified among the proteins showing significant expression changes. On the 2-D gels both PFORs appeared as a chain of spots (Fig. 4). Protein identification showed that the spots 70, 75, 76, 77, 87, 90 are pI isoforms of the homodimeric protein pyruvate: ferredoxin (flavodoxin) oxidoreductase (F502_07643), whereas the spots 40, 57, 58 and 59 are pI isoforms of pyruvate:ferredoxin oxidoreductase (F502_07648). F502_07643 and F502_07648 are homologous proteins with a sequence identity match of 66% and positive match of 81%. They have nearly identical molecular weights but the pI value of F502_07648 is more basic than that of F502_07643, which was also obvious on the 2-D gels. Although previous studies showed that under conditions of iron limitation many anaerobes synthesize flavodoxins as substitution of ferredoxins for many enzymatic reactions [28], all the isoforms of the two PFORs showed, in general, higher expression at iron excess than at iron limitation (Fig. 4). The expression patterns of the isoforms of PFOR1 were similar to each other, with the highest expression at Fe+ late. In contrast, the expression patterns of the isoforms of pyruvate:ferredoxin (flavodoxin) oxidoreductase (PFOR2) were different to each other. While the expression level of the spot 87 in the middle of the spot chain did not change between Fe+ early and Fe+ late, the more basic isoforms (spots 70 and 90) showed higher expression at Fe+ early, and the more acidic isoforms (spots 75, 76 and 77) were up-regulated at Fe+ late which was similar to the expression pattern of PFOR1. Therefore, based on the proteomic results alone, it is not clear whether these two PFORs function in synchronization or are differently regulated in response to iron availability. Furthermore, whether PFOR2 transfers the electrons generated during the decarboxylation reaction to a ferredoxin or flavodoxin remains elusive.
Fig. 4.
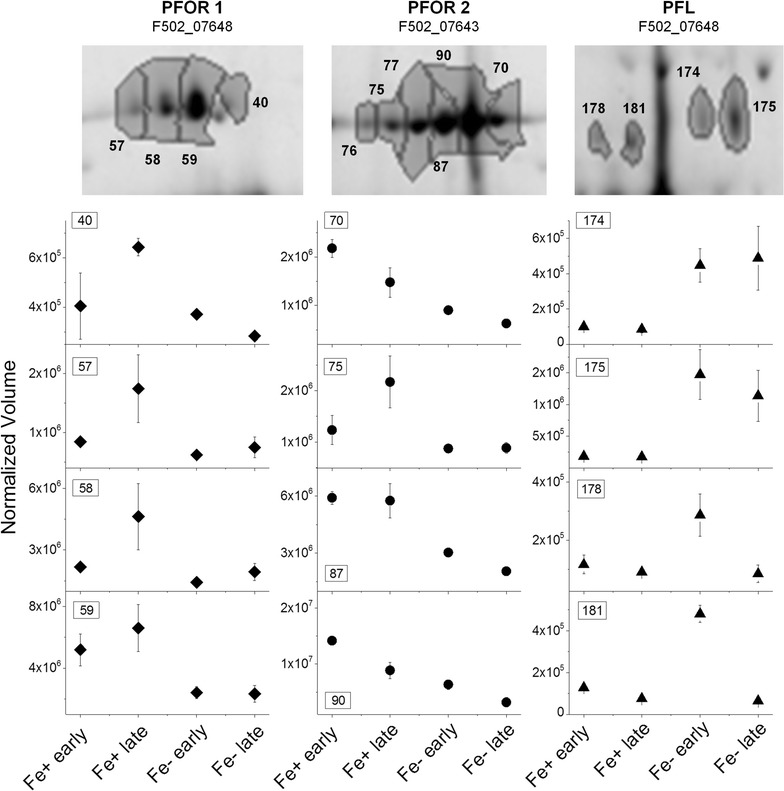
Expression patterns of enzymes catalyzing the conversion of pyruvate to acetyl-CoA under Fe+ and Fe− conditions at exponential growth phase (early) and stationary phase (late). PFOR 1 (F502_07648, pyruvate: ferredoxin oxidoreductase) and PFOR 2 [F502_07643, pyruvate:ferredoxin (flavodoxin) oxidoreductase] are up-regulated at Fe+ ; PFL (pyruvate formate lyase) is up-regulated at Fe−
Beside the ferredoxin (flavodoxin)-dependent PFORs, acetyl-CoA can be synthesized from pyruvate through the pyruvate formate-lyase (PFL) with the formation of formate. There are three genes (F502_19556, F502_15690, F502_15710) in the genome of C. pasteurianum DSMZ 525 being annotated to encoding enzymes functioning as pyruvate formate lyase (PFL). Only one of these PFLs (F502_19556, also named formate acetyltransferase) was unambiguously identified in four protein spots (Fig. 4). Interestingly, the molecular weight of the two acidic isoforms (spots 178 and 181) appeared lower than that of the two basic isoforms (spots 174 and 175). Compared to the iron excess condition, where the expression of PFL was nearly not detectable, all these four PFL isoforms were significantly but differently up-regulated under the iron limitation condition. While the two acidic isoforms showed 2.5 and 3.7 folds increased abundances only in the Fe− early sample, the expression of the two basic isoforms were up to eightfold strongly up-regulated in the Fe− early sample and about sixfold in Fe− late sample. Thus, under iron limitation, it was apparently favorable for C. pasteurianum to use the PFL-catalyzed reaction for the conversion of pyruvate to acetyl-CoA. Correspondingly, under this condition the formate yield was clearly higher than that under the Fe+ condition, namely 0.25 ± 0.05 g/g biomass in contrast to 0.08 ± 0.02 g/g biomass. Nevertheless, the expression levels of the two PFORs, especially the pyruvate:ferredoxin (flavodoxin) oxidoreductase (PFOR2), were visibly higher than that of PFL. Since protein synthesis is an energy-demanding process, cells usually do not produce useless enzymes in noticeable amounts. The presence of the two PFORs under Fe− condition may point to a fact that, in the absence of iron, the two PFORs, especially the pyruvate: ferredoxin (flavodoxin) oxidoreductase (PFOR2), probably use flavodoxin instead of ferredoxin as the electron acceptor. Indeed, the expression of a flavodoxin (F502_13493) was strongly up-regulated under iron limitation for 14.3 folds in the exponential growth phase and remained high even after entering the stationary phase (8.0 fold higher in Fe− late than in Fe+ late). However, whether or not the up-regulated expression of this flavodoxin was coupled to the functionality of the PFORs remains to be verified. In case it is, it did not help much in sustaining the production of butanol under the Fe− condition.
Regulation of the ferredoxin pool
For the proper function of PFORs, ferredoxin(red), which is generated in the PFOR-catalyzed pyruvate oxidative decarboxylation reaction, must be oxidized to regenerate ferredoxin(ox). C. pasteurianum DSMZ 525 possesses a big number of ferredoxins and the regeneration of ferredoxin(ox) can be achieved using different electron acceptors. The fact that the redox potentials of ferredoxins (−400 mV) are in the range of H2 electrodes (−414 mV, at pH 7) reveals that in most energy metabolisms where ferredoxins are active, H2 is also involved, either as substrate or as product. In general, nitrogenases and hydrogenases are the two enzyme classes capable of hydrogen production in Clostridia [33]. But Hallenbeck and Benemann [34] reported that hydrogenases are much more efficient, with more than 1000 times higher turnover than nitrogenases. Hydrogenases are divided into two main groups in Clostridia based on their metaollocenter composition, namely [NiFe] and [FeFe] hydrogenases [33]. In this study, the expression of hydrogenase-1 (F502_18287), which belongs to the [FeFe] group, was highly up-regulated under iron excess, showing up to fivefolds higher expression level in the exponential growth phase under the Fe+ condition compared to the Fe− condition. After entering the stationary phase the expression of hydrogenase-1 (H2-ase) was down-regulated for two to threefolds under the Fe+ condition, which could be possibly in response to a depletion of the intracellular iron pool in the Fe+ late sample required for this [FeFe]-hydrogenase. However, it was still nearly twofolds higher than its expression level under the Fe− condition. An additional [FeFe] hydrogenase (F502_14390) was also identified which showed expression regulations similar to that of the hydrogenase-1 (F502_18287). Nevertheless this hydrogenase did not appear as a spot containing only a single protein on the 2-D gels and therefore could not be quantified for comparison. The higher expression of hydrogenase-1 (F502_18287) coincided with the higher H2 production in the fermentation culture under iron excess and should have significantly contributed to the regeneration of ferredoxin(red) to ferredoxin(ox).
However, it is to notice that the regulation of hydrogenase-1 (F502_18287) expression is rather in agreement with that of the basic iso-forms of the ferredoxin (flavodoxin)-dependent PFOR2 than with the expression patterns of the ferredoxin-dependent PFOR1. Therefore, it is tempting to suggest that under the given experimental conditions the hydrogenase-1 catalyzed reaction should not be the only route of ferredoxin(ox) regeneration. The PFORs-catalyzed pyruvate oxidation to acetyl-CoA might be coupled with other but yet unknown ferredoxin(ox) regenerating reaction(s) catalyzed either by other unidentified hydrogenases (at least 5 genes in the genome of C. pasteurianum DSMZ 525 encode hydrogenases) or ferredoxin reductases. In addition, it has also been reported that PFOR can transfer the electrons generated in the decarboxylation reaction directly to protons to generate molecular hydrogen [35].
Within the cells of anaerobes including Clostridia, 90% of ferredoxins were reported to be present in reduced form, allowing them to serve as electron donors in different reactions [28]. In general, this is achieved in C. pasteurianum by the following three ferredoxin-dependent redox reactions: the oxidation of pyruvate to acetyl-CoA and CO2 (−500 mV), the oxidation of formate to CO2 (−430 mV, [36]) and the flavoprotein based electron bifurcation involved in the reduction of crotonyl-CoA to butyryl-CoA (Eq. 7). On the other hand, the oxidation of Fdred by NAD is excluded due to the absence of the ferredoxin: NAD oxidoreductase activity [28]. Therefore, hydrogen production via hydrogenase should be a main route of Fdox regeneration in C. pasteurianum DSMZ 525. Based on this assumption, we compared the hydrogen yield from glycerol between the Fe− and the Fe+ conditions. As shown in Fig. 5a, hydrogen yield decreased significantly from 0.75 mol/mol glycerol under Fe+ condition to 0.21 under Fe− condition, which was in agreement with the higher expression of hydrogenase-1 under the Fe+ condition.
Fig. 5.
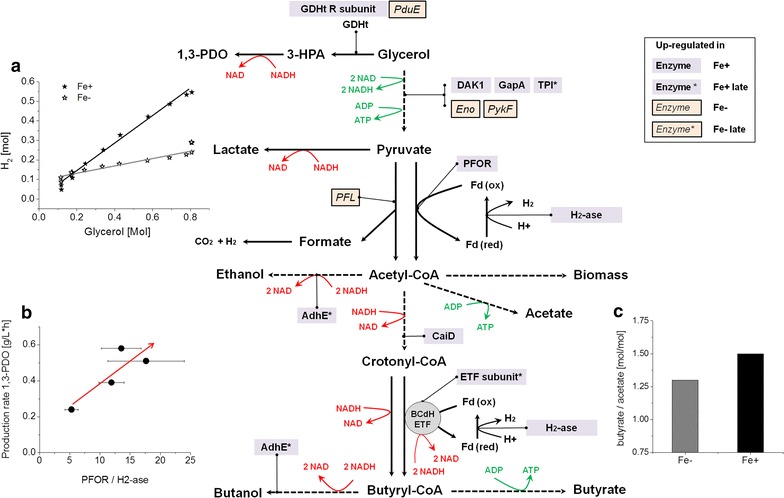
Revised metabolic pathway of glycerol bioconversion to 1,3-propanediol and n-butanol in C. pasteurianum. a Molar formation of H2 over consumed glycerol b 1,3-PDO productivity over the ratio PFOR/H2-ase c Molar ratio of butyrate to acetate under Fe+ and Fe− conditions
In addition, as described in the above redox balance analysis, a possible involvement of a ferredoxin-dependent butyryl-CoA dehydrogenase/electron transferring flavoprotein complex (BCdH-ETF) in H2 production was proposed. In the BCdH-ETF catalyzed reaction electron transfer flavoproteins (ETFs) are involved in the reduction of crotonyl-CoA to butyryl-CoA, coupled with ferredoxin(ox) reduction by bifurcating electrons from NADH (Fig. 5) [28, 37]. In this proteomic study, two ETFs, namely ETFs subunit alpha (F502_06282) and subunit alpha/beta-like protein (F502_06287), were identified among the most abundant proteins regardless of the iron availability; however, their abundances were slightly higher (1.5–1.7 folds) in the late phase of the Fe+ culture compared to the Fe− late condition. This might indicate a relative increase in the oxidized ferredoxin pool necessary to carry out the BCdH-ETF reaction and also contributed to the stronger H2 production in the late fermentation phase under Fe+ condition.
Nevertheless, not only the hydrogenase-1 but also the two PFORs were down-regulated under Fe− condition compared to that under Fe+ condition. Therefore, the relative changes of the expression levels of the two enzymes might be indicative of the overall Fdox regeneration state (Fig. 5). The expression levels shown as the protein spot intensities of both enzymes under Fe− and Fe+ conditions at the two time points were thus compared. As shown in Fig. 5b, the relative expression of PFOR to hydrogenase-1 (H2-ase) showed a positive correlation with the 1,3-PDO production rate. The decrease in the Fdox fraction under Fe− condition due to reduced H2-ase presence will decrease the Fdox coupled synthesis of butyryl-CoA catalyzed by the BCdH-ETF complex, a crucial step in butyrate and especially butanol biosynthesis. Moreover, the intermediate acetyl-CoA will be favorably channeled into the Fdox-independent acetate formation route than the Fdox-dependent butyrate formation route, as shown by the increase in the butyrate/acetate ratio from 1.3 at Fe− to 1.5 at Fe+ (Fig. 5c). Consequently, it seems that under Fe− condition, Fdox dependent conversion steps are reduced and the resulting free reducing power, usually needed for butanol formation, could be redirected for the sake of redox balance to the production of 1,3-PDO and lactate. Indeed, the overall yield of 1,3-PDO and lactate were much higher under iron limitation than under iron excess (Table 1). Nevertheless, lactate dehydrogenase catalyzing the conversion of pyruvate to lactate or 1,3-propanediol dehydrogenase catalyzing the formation of 1,3-propanediol from 3-HPA were not found among the proteins showing significant changes in expression level. Both enzymes are not involved in energy production but constitute the cell’s back-up for stabilizing an internal redox balance, and hence their constitutive production may be a mechanism to withstand sudden perturbations in the NADH/NAD ratio. Nevertheless, it should also bear in mind that higher or lower protein level does not always means higher or lower enzyme activity.
Glycerol conversion to 1,3-propanediol
In general the bioconversion of glycerol to 1,3-PDO takes place in two steps, catalyzed successively by glycerol dehydratase (GDHt) and 1,3-propanediol dehydrogenase (PDOR) (Fig. 5). It is known that glycerol dehydratase is the rate-limiting enzyme in this bioconversion. All the three subunit of GDHt encoded by pduC (F502_03402), pduD (F502_03407) and pduE (F502_03412) were identified but unfortunately not as single protein spots and, therefore, could not be quantified. Instead, the large subunit of glycerol dehydratase reactivating factor (GDHt reactivase, GDHtR) was identified in the spot containing this single protein (Fig. 6). GDHtR is a molecular chaperone participating in the reactivation of inactivated GDHt in the presence of ATP and Mg2+ [38, 39]. The expression pattern of GDHtR indicates rather a correlation of GDHtR expression to cell growth phase than to iron availability. Among the four samples compared by proteomics, the highest expression level of GDHtR was present in the iron excess culture in the middle exponential growth phase (Fe+ early) showing the highest specific growth rate (µ = 0.22). At this sampling time point, the culture under iron limitation (Fe− early) already entered late exponential growth phase with reduced specific growth rate (µ = 0.07), accompanied with lower GDHtR level. The GDHtR abundance was further reduced to merely detectable levels in the stationary phase (Fe+ late and Fe− late), where the production of 1,3-PDO stagnated. 1,3-propanediol dehydrogenase (PDOR), the responsible enzyme for the conversion of 3-HPA to 1,3-PDO, was one of the highly abundant proteins on the 2-D gels and did not show significant expression changes under the different conditions (data not shown).
Fig. 6.
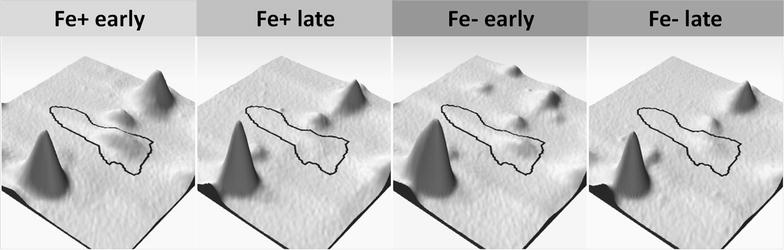
Expression pattern of the large subunit of glycerol dehydratase reactivating factor (GDHtR) under Fe+ and Fe− conditions as well as early and late sampling point
Conclusion
The iron content in the fermentation medium was shown to influence the product formation, especially the 1,3-propanediol and butanol distribution, in Clostridium pasteurianum DSMZ 525 grown on glycerol. Compared to the fermentation under iron limitation, it was shown that the butanol to 1,3-propanediol ratio increased almost fivefold in fermentation under iron excess. To better understand the effect of iron on the regulation of the cell metabolism, physiological and proteomic analyses were performed. Several enzymes like pyruvate: ferredoxin oxidoreductase (PFOR), hydrogenase and bifunctional acetaldehyde-CoA/alcohol dehydrogenase among others were found to be up-regulated under iron excess conditions. The differential expression of PFORs and a pyruvate formate lyase (PFL) in response to iron availability highlighted the impact of iron on the crucial step of the central carbon metabolism in C. pasteurianum. The importance of a hydrogenase in the regeneration of oxidized ferredoxin and therefore the maintaining of the redox balance was confirmed by its strong up-regulation under the iron excess condition. Beside the release of molecular hydrogen in the pyruvate to acetyl-CoA step catalyzed by PFORs, stoichiometric analysis showed a possible H2 production coupled to the reaction catalyzed by the ferredoxin-dependent butyryl-CoA dehydrogenase/electron transfer flavoprotein complex (BCdH-ETF). Indeed, proteomic analysis revealed the up-regulation of two electron transfer flavoproteins which may be involved in this metabolic conversion step. Since both 1,3-propanediol and butanol can be used as sink for NADH, we suggest that the ratio of oxidized ferredoxin to reduced ferredoxin in addition to the NADH availability contributes to the selectivity of the products.
Authors’ contributions
CG designed and performed the bioreactor experiments, interpreted the results and wrote the manuscript. WW carried out the proteomic studies, interpreted the results and wrote the manuscript. WS and TU helped in bioreactor experiments and results interpretation. APZ supervised the research. All authors read and approved the final manuscript.
Acknowledgements
The authors also like to thank Jan Bomnüter and Anna Gorte for their excellent assistance with the two dimensional electrophoresis of proteomic analysis.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Funding Information
This work was funded by from the European Union Seventh Framework Program (FP7/2007 2013) through the EuroBioRef project (Grant Agreement Nr. 241718). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Christin Groeger, Email: christin.groeger@tuhh.de.
Wei Wang, Email: wei.wang@tu-harburg.de.
Wael Sabra, Email: wael.sabra@tu-harburg.de.
Tyll Utesch, Email: tyll.utesch@tuhh.de.
An-Ping Zeng, Phone: +49 40 428 78 41 83, Email: aze@tuhh.de.
References
- 1.Kao WC, Lin DS, Cheng CL, Chen BY, Lin CY, Chang JS. Enhancing butanol production with Clostridium pasteurianum CH4 using sequential glucose–glycerol addition and simultaneous dual-substrate cultivation strategies. Bioresour Technol. 2013;135:324–330. doi: 10.1016/j.biortech.2012.09.108. [DOI] [PubMed] [Google Scholar]
- 2.Sabra W, Groeger C, Sharma PN, Zeng AP. Improved n-butanol production by a non-acetone producing Clostridium pasteurianum DSMZ 525 in mixed substrate fermentation. Appl Microbiol Biotechnol. 2014;98:4267–4276. doi: 10.1007/s00253-014-5588-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Schwarz KM, Grosse-Honebrink A, Derecka K, Rotta C, Zhang Y, Minton NP. Towards improved butanol production through targeted genetic modification of Clostridium pasteurianum. Metab Eng. 2017;40:124–137. doi: 10.1016/j.ymben.2017.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Biebl H. Fermentation of glycerol by Clostridium pasteurianum–batch and continuous culture studies. J Ind Microbiol Biotechnol. 2001;27:18–26. doi: 10.1038/sj.jim.7000155. [DOI] [PubMed] [Google Scholar]
- 5.Pyne ME, Sokolenko S, Liu X, Srirangan K, Bruder MR, Aucoin MG, Moo-Young M, Chung DA, Chou CP. Disruption of the reductive 1,3-propanediol pathway triggers production of 1,2-propanediol for sustained glycerol fermentation by Clostridium pasteurianum. Appl Environ Microbiol. 2016;82:5375–5388. doi: 10.1128/AEM.01354-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kaeding T, DaLuz J, Kube J, Zeng AP. Integrated study of fermentation and downstream processing in a miniplant significantly improved the microbial 1, 3-propanediol production from raw glycerol. Bioprocess Biosyst Eng. 2015;38(3):575–586. doi: 10.1007/s00449-014-1297-z. [DOI] [PubMed] [Google Scholar]
- 7.Choi O, Kim T, Woo HM, Um Y. Electricity-driven metabolic shift through direct electron uptake by electroactive heterotroph Clostridium pasteurianum. Sci Rep. 2014;4:6961. doi: 10.1038/srep06961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Groeger C, Sabra W, Zeng A-P. Study of in situ gas stripping and cellular metabolism for simultaneous production of 1,3-propanediol and n-butanol by Clostridium pasteurianum. Eng Life Sci. 2016;16:664–674. doi: 10.1002/elsc.201600058. [DOI] [Google Scholar]
- 9.Jensen TO, Kvist T, Mikkelsen MJ, Westermann P. Production of 1,3-PDO and butanol by a mutant strain of Clostridium pasteurianum with increased tolerance towards crude glycerol. AMB Express. 2012;2:44. doi: 10.1186/2191-0855-2-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lee CS, Aroua MK, Daud W, Cognet P, Peres-Lucchese Y, Fabre P, Reynes O, Latapie L. Conversion of bioglycerol into 1,3-propanediol via biological and chemical method. a review. Renew Sustain Energy reviews. 2015;42:963–972. doi: 10.1016/j.rser.2014.10.033. [DOI] [Google Scholar]
- 11.Sabra W, Wang W, Surandram S, Groeger C, Zeng AP. Fermentation of mixed substrates by Clostridium pasteurianum and its physiological, metabolic and proteomic characterizations. Microb Cell Fact. 2016;15:114. doi: 10.1186/s12934-016-0497-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Venkataramanan KP, Boatman JJ, Kurniawan Y, Taconi KA, Bothun GD, Scholz C. Impact of impurities in biodiesel-derived crude glycerol on the fermentation by Clostridium pasteurianum ATCC 6013. Appl Microbiol Biotechnol. 2012;93:1325–1335. doi: 10.1007/s00253-011-3766-5. [DOI] [PubMed] [Google Scholar]
- 13.Yadav S, Rawat G, Tripathi P, Saxena RK. Dual substrate strategy to enhance butanol production using high cell inoculum and its efficient recovery by pervaporation. Bioresour Technol. 2014;152:377–383. doi: 10.1016/j.biortech.2013.11.022. [DOI] [PubMed] [Google Scholar]
- 14.Pyne ME, Liu X, Moo-Young M, Chung DA, Chou CP. Genome-directed analysis of prophage excision, host defence systems, and central fermentative metabolism in Clostridium pasteurianum. Sci Rep. 2016;6:26228. doi: 10.1038/srep26228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Malaviya A, Jang YS, Lee SY. Continuous butanol production with reduced byproducts formation from glycerol by a hyper producing mutant of Clostridium pasteurianum. Appl Microbiol Biotechnol. 2012;93:1485–1494. doi: 10.1007/s00253-011-3629-0. [DOI] [PubMed] [Google Scholar]
- 16.Gallardo R, Alves M, Rodrigues LR. Influence of nutritional and operational parameters on the production of butanol or 1,3-propanediol from glycerol by a mutant Clostridium pasteurianum. New Biotechnol. 2017;34:59–67. doi: 10.1016/j.nbt.2016.03.002. [DOI] [PubMed] [Google Scholar]
- 17.Sarchami T, Johnson E, Rehmann L. Optimization of fermentation condition favoring butanol production from glycerol by Clostridium pasteurianum DSM 525. Bioresour Technol. 2016;208:73–80. doi: 10.1016/j.biortech.2016.02.062. [DOI] [PubMed] [Google Scholar]
- 18.Moon C, Lee CH, Sang BI, Um Y. Optimization of medium compositions favoring butanol and 1,3-propanediol production from glycerol by Clostridium pasteurianum. Bioresour Technol. 2011;102:10561–10568. doi: 10.1016/j.biortech.2011.08.094. [DOI] [PubMed] [Google Scholar]
- 19.Regestein L, Doerr EW, Staaden A, Rehmann L. Impact of butyric acid on butanol formation by Clostridium pasteurianum. Bioresour Technol. 2015;196:153–159. doi: 10.1016/j.biortech.2015.07.085. [DOI] [PubMed] [Google Scholar]
- 20.Dabrock B, Bahl H, Gottschalk G. Parameters Affecting Solvent Production by Clostridium pasteurianum. Appl Environ Microbiol. 1992;58:1233–1239. doi: 10.1128/aem.58.4.1233-1239.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Oehmke S, Zeng A-P. Recovery of biologically produced 3-hydroxypropionaldehyde and its dehydrated product acrolein. Eng Life Sci. 2015;15:133–139. doi: 10.1002/elsc.201300122. [DOI] [Google Scholar]
- 22.Abbad-Andaloussi S, Durr C, Raval G, Petitdemange H. Carbon and electron flow in Clostridium butyricum grown in chemostat culture on glycerol and on glucose. Microbiology. 1996;1432:1149–1158. doi: 10.1099/13500872-142-5-1149. [DOI] [PubMed] [Google Scholar]
- 23.Vollenweider S, Lacroix C. 3-hydroxypropionaldehyde: applications and perspectives of biotechnological production. Appl Microbiol Biotechnol. 2004;64:16–27. doi: 10.1007/s00253-003-1497-y. [DOI] [PubMed] [Google Scholar]
- 24.Avila M, Gomez-Torres N, Hernandez M, Garde S. Inhibitory activity of reuterin, nisin, lysozyme and nitrite against vegetative cells and spores of dairy-related Clostridium species. Int J Food Microbiol. 2014;172:70–75. doi: 10.1016/j.ijfoodmicro.2013.12.002. [DOI] [PubMed] [Google Scholar]
- 25.Zeng A-P. Pathway and kinetic analysis of 1,3-propanediol production from glycerol fermentation by Clostridium butyricum. Bioprocess Eng. 1996;14:169–175. doi: 10.1007/BF01464731. [DOI] [Google Scholar]
- 26.Zeng A-P, Biebl H, Schlieker H, Deckwer WD. Pathway analysis of glycerol fermentation by Klebsiella pneumoniae: regulation of reducing equivalent balance and product formation. Enzyme Microb Technol. 1993;15:770–779. doi: 10.1016/0141-0229(93)90008-P. [DOI] [Google Scholar]
- 27.Lan EI, Liao JC. Metabolic engineering of cyanobacteria for 1-butanol production from carbon dioxide. Metab Eng. 2011;13:353–363. doi: 10.1016/j.ymben.2011.04.004. [DOI] [PubMed] [Google Scholar]
- 28.Buckel W, Thauer RK. Energy conservation via electron bifurcating ferredoxin reduction and proton/Na(+) translocating ferredoxin oxidation. Biochim Biophys Acta. 2013;1827:94–113. doi: 10.1016/j.bbabio.2012.07.002. [DOI] [PubMed] [Google Scholar]
- 29.Kim BH, Bellows P, Datta R, Zeikus JG. Control of carbon and electron flow in Clostridium acetobutylicum fermentations: utilization of carbon monoxide to inhibit hydrogen production and to enhance butanol yields. Appl Environ Microbiol. 1984;48:764–770. doi: 10.1128/aem.48.4.764-770.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Moulis JM, Davasse V, Meyer J, Gaillard J. Molecular mechanism of pyruvate-ferredoxin oxidoreductases based on data obtained with the Clostridium pasteurianum enzyme. FEBS Lett. 1996;380:287–290. doi: 10.1016/0014-5793(96)00062-2. [DOI] [PubMed] [Google Scholar]
- 31.Charon MH, Volbeda A, Chabriere E, Pieulle L, Fontecilla-Camps JC. Structure and electron transfer mechanism of pyruvate: ferredoxin oxidoreductase. Curr Opin Struct Biol. 1999;9:663–669. doi: 10.1016/S0959-440X(99)00027-5. [DOI] [PubMed] [Google Scholar]
- 32.Summary of Clostridium pasteurianum, Strain DSM 525 = ATCC 6013, version 20.1. http://www.biocyc.org/organism-summary?object=CPAS1262449. Accessed 30 Nov 2016.
- 33.Calusinska M, Happe T, Joris B, Wilmotte A. The surprising diversity of clostridial hydrogenases: a comparative genomic perspective. Microbiology. 2010;156:1575–1588. doi: 10.1099/mic.0.032771-0. [DOI] [PubMed] [Google Scholar]
- 34.Hallenbeck PC, Benemann JR. Biological hydrogen production; fundamentals and limiting processes. Biohydrogen. 2002;27:1185–1193. [Google Scholar]
- 35.Menon S, Ragsdale SW. Unleashing hydrogenase activity in carbon monoxide dehydrogenase/acetyl-CoA synthase and pyruvate: ferredoxin oxidoreductase. Biochemistry. 1996;35:15814–15821. doi: 10.1021/bi9615598. [DOI] [PubMed] [Google Scholar]
- 36.Scherer PA, Thauer RK. Purification and properties of reduced ferredoxin: CO2 oxidoreductase from Clostridium pasteurianum, a molybdenum iron-sulfur-protein. Eur J Biochem. 1978;85:125–135. doi: 10.1111/j.1432-1033.1978.tb12220.x. [DOI] [PubMed] [Google Scholar]
- 37.Herrmann G, Jayamani E, Mai G, Buckel W. Energy conservation via electron-transferring flavoprotein in anaerobic bacteria. J Bacteriol. 2008;190:784–791. doi: 10.1128/JB.01422-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kajiura H, Mori K, Tobimatsu T, Toraya T. Characterization and mechanism of action of a reactivating factor for adenosylcobalamin-dependent glycerol dehydratase. J Biol Chem. 2001;276:36514–36519. doi: 10.1074/jbc.M105182200. [DOI] [PubMed] [Google Scholar]
- 39.Toraya T, Mori K. A reactivating factor for coenzyme B12-dependent diol dehydratase. J Biol Chem. 1999;274:3372–3377. doi: 10.1074/jbc.274.6.3372. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.