

Abstract
Mortality associated with breast cancer is attributable to aggressive metastasis, to which TNFα plays a central orchestrating role. TNFα acts on breast tumor TNF receptors evoking the release of chemotactic proteins (e.g. MCP-1/CCL2). These proteins direct inward infiltration/migration of tumor-associated macrophages (TAMs), tumor-associated neutrophils (TANs), myeloid-derived suppressor cells (MDSCs), T-regulatory cells (Tregs), T helper IL-17-producing cells (Th17s), metastasis-associated macrophages (MAMs) and cancer-associated fibroblasts (CAFs). Tumor embedded infiltrates collectively enable immune evasion, tumor growth, angiogenesis, and metastasis. In the current study, we investigate the potential of apigenin, a known anti-inflammatory constituent of parsley, to downregulate TNFα mediated release of chemokines from human triple-negative cells (MDA-MB-231 cells). The results show that TNFα stimulation leads to large rise of CCL2, granulocyte macrophage colony-stimulating factor (GMCSF), IL-1α and IL-6, all suppressed by apigenin. While many aspects of the transcriptome for NFkB signaling were evaluated, the data show signaling patterns associated with CCL2 were blocked by apigenin and mediated through suppressed mRNA and protein synthesis of IKBKe. Moreover, the data show that the attenuation of CCL2 by apigenin in the presence TNFα paralleled the suppression of phosphorylated extracellular signal-regulated kinase 1 (ERK 1/ 2). In summary, the obtained findings suggest that there exists a TNFα evoked release of CCL2 and other LSP recruiting cytokines from human breast cancer cells, which can be attenuated by apigenin.
Introduction
Breast cancer is a leading cause of death in women worldwide, despite medical advances in surgical techniques, radiation and chemotherapy. There is a continued need to identify preventive and adjunctive chemosensitization agents in the management of breast cancer. Apigenin (4',5,7-trihydroxyflavone) is naturally occurring bioactive compound (NOBC) found in parsley, garlic, and celery. The biological relevance of apigenin in management of breast cancer is evidenced by the growing pool of research (in vitro and xenograft tumor models) demonstrating its capacity to inhibit tumor promoting enzymes [1,2], overcome chemoresistance [3,4] and/or multi-drug resistance [5], prevent angiogenesis [6] metastasis and lessen the growth of aggressive breast cancers [7]. There is also a plethora of research showing apigenin as a cytostatic agent with capacity to attenuate clonogenic survival in a wide range of breast cancer cells; MDA-MB-231 [8], SKBR3 [9,10], Hs578T, MCF-7 [11], MDA-MB-453 [12], MDA-MB-468 Cells [13,14], T47D [15] and BT-474 Cells [16]. Biological targets of apigenin relevant to tumor potential are known to involve MAPK/NFkappaB, phospho-JAK1/ STAT3 signaling [17], VEGF, aromatase, proteasomal processes, fatty acid synthase [18–21] and the Fas-associated protein with death domain (FADD) [22].
Despite surmounting evidentiary support for apigenin in tumor suppression, there is lack of research regarding its influence on the tumor microenvironment in response to pro-inflammatory cytokines, specifically TNFα. TNFα is present in high concentrations throughout the breast tumor/ stroma milieu, and upon activation of TNFα receptors, can trigger a powerful perpetual cascade of NF-κB activation [23,24]. epithelial-to-mesenchymal transition [25] and sustained release of diverse chemokines (i.e., CCL2/CCL5) [26]. Chemokines in turn ultimately trigger inward migration of a large number of leukocyte sub-populations (LSPs) bearing CCR2 / CCR5 receptors that embed into the tumor, and further drive tumor aggression and stem cell survival. [27,28] Tumor promoting LSPs are known to include tumor-associated macrophages (TAMs) [29], myeloid-derived suppressor cells (MDSCs) [30], tumor-associated neutrophils (TANs)[31,32], T-regulatory (Tregs) [33] metastasis-associated macrophages (MAMs), T helper IL-17-producing cells (Th17) cells and cancer-associated fibroblasts (CAFs) [34].
Thus, given the dynamic control of diverse chemokines as key trafficking molecules released by tumor cells in response to TNFα capable of driving LSP recruitment [35–38], here we investigated the effects of apigenin on TNFα mediated chemokine release in triple-negative breast cancer (TNBC) cells.
Results
A basic cell viability in MDA-MB-231 cells was performed to establish appropriate non-lethal working concentrations of apigenin and TNFα (Fig 1A) and (Fig 1B), respectively. Based on these baseline studies, we chose the concentrations of 40μM of apigenin and 40ng/ml of TNFα to carry out all subsequent studies. A semiquantitative analysis was performed to study the cytokine release pattern, in the presence of TNFα and +/- apigenin, using sandwich-based protein arrays from RayBiotech (Norcross, GA, USA). The data obtained show that TNFα caused significant upregulation of few specific cytokines. These cytokines included GM-CSF, CCL2, IL-1α, IL-6 (Fig 2A–2D) IL-8 and GRO CXCL1/CXCL2/CXCL3 a/b/g (Fig 3A–3D), some of which were downregulated by apigenin and subject to further validation by ELISA assays. First, the densitometry array values (reflected by INT/MM2) are shown for the effect of apigenin on TNFα mediated release of GM-CSF (Fig 4A), IL-6 (Fig 4B), CCL2 and IL-1α (Fig 4C). Results were then validated by independent ELISA, where the data paralleled between the findings in the arrays for both CCL2 and IL-1α (Fig 5A and 5B).
Fig 1.

The effect of apigenin (A) and TNFα (B) on cell viability of MDA-MB-231 cells at 5% CO2/Atm for 24 Hr. The data are presented as viability (% Ctrl), Mean ± S.E.M. n = 4. The significance of differences from the Ctrl and were determined by a one-way ANOVA, with a Tukey post hoc test. *p<0.05.
Fig 2.

(A-D). The release of cytokines as assessed using Human Cytokine Array C1000 membranes from MDA-MB-231 cells ± TNFα ± Apigenin. [A] Ctrls, [B] TNFα-treated (40ng/mL) [C] Apigenin (40μM) and [D] TNFα-treated (40ng/mL) + Apigenin (40μM) co-treated cells.
Fig 3.

(A-D). The release of cytokines as assessed using Human Cytokine Array C1000 membranes from MDA-MB-231 cells ± TNFα ± Apigenin. [A]Ctrls, [B]TNFα-treated (40ng/mL) [C]Apigenin (40μM) and [D]TNFα-treated (40ng/mL) + Apigenin (40μM) co-treated cells. The data present membrane spot densities (top) and manufacturer’s grid layout (bottom).
Fig 4.

The effect of apigenin on TNFα mediated changes in GM-CSF (A) IL-6 (B), CCL2 and IL-1α (C) in MDA-MB-231 cells. The data represent the densitometry values (expressed as intensity per square millimeter) (INT/MM2) per spot as Mean ± S.E.M. n = 3. The significance of differences between the Ctrl and TNFα groups and TNFα vs. TNFα + Apigenin were determined by a Students t-test *p<0.05.
Fig 5.

Effect of Apigenin on TNFα mediated changes in CCL2 (A) and IL-1α (B) in MDA-MB-231 cells. The data represent CCL (pg/ul) or IL-1α (O.D. @ 405nm) as Mean ± S.E.M. n = 3. The significance of differences between the Ctrl and TNFα groups and TNFα vs. TNFα + Apigenin were determined by a Students t-test *p<0.05.
To elucidate potential signaling changes associated with these findings, which were occurring at the transcriptome level, RT-PCR NF-κB arrays were conducted using Signaling Pathway H96 Predesigned 96-well panel for use with SYBR® Green.CAT Number: 10025558. Biorad (Hercules, CA, USA). The data show a lack of significant changes in all mRNAs involved with NF-κB transcriptome pathway, where IL-1α shows a distinct but non-significant change (Fig 6A and 6B and Table 1). The lack of data as reflected by this RT-PCR NF-κB array was surprising, but a limitation to this array was the absence of IKBK-epsilon which plays a pivotal role in driving tumor malignancy in both TNBC [39] and receptor positive breast cancer cells [40]. Therefore, in the next study, we investigate changes in IKBK-epsilon transcription determined by RT-PCR (Fig 7A) and protein expression using Western blot (Fig 7B). These findings validate IKBK-epsilon to be the major proponent of NF-κB signaling control involved with TNF-α mediated rise in CCL2 release and its reduction by apigenin. In the next study, we demonstrate additional influential effects of MAPK signaling on TNF-α mediated CCL2 release, focusing on changes in phospho-ERK protein, also downregulated by apigenin (Fig 8). These findings show that apigenin can alter TNF-α mediated breast cancer signaling pathways that control the release of CCL2, which in part involve IKBK-epsilon, and ERK, both essential biological events required for TAM infiltration and recruitment into solid breast tumor tissue.
Fig 6. NF-kB PCR microarray assessment on MDA-MB-231 cells ± TNFα ± Apigenin.
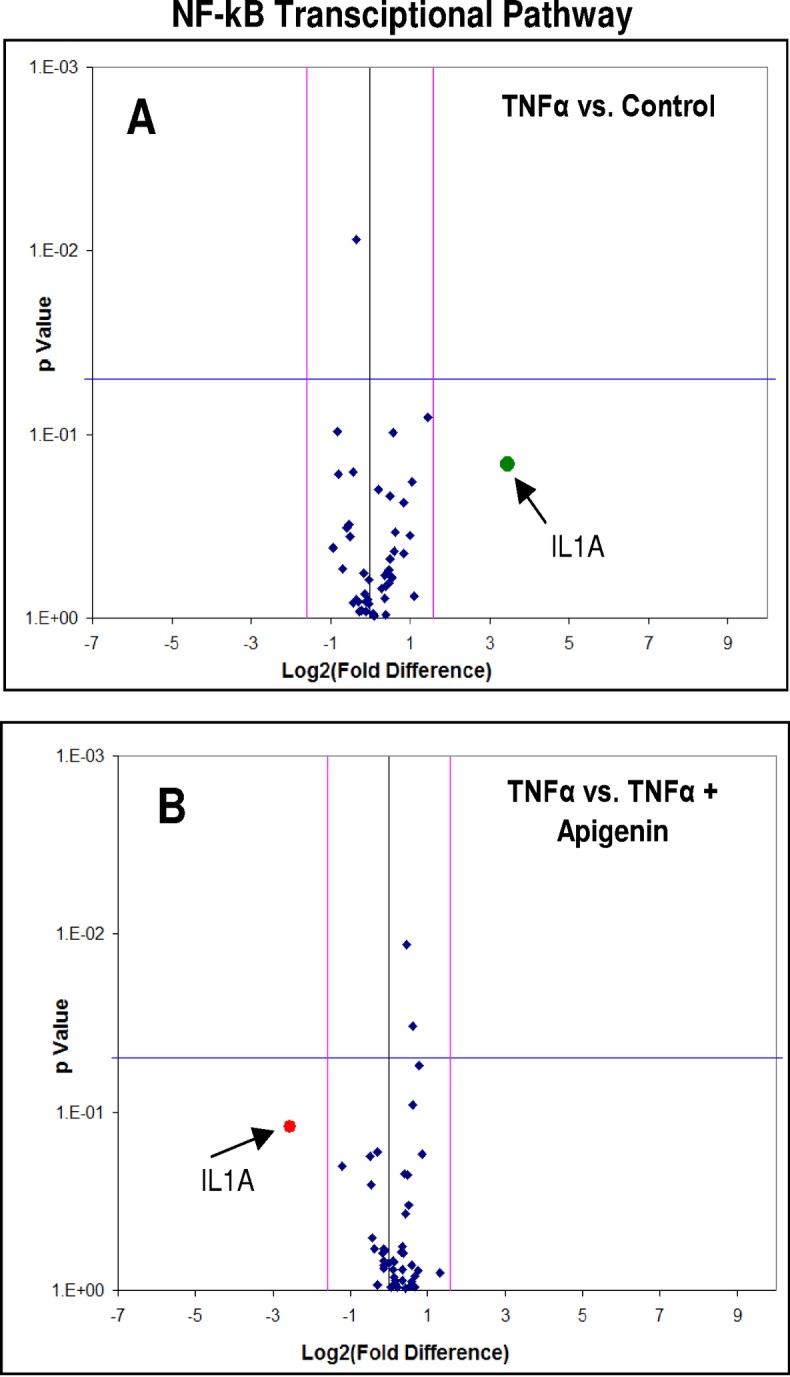
The data displays differential transcription for controls vs. TNFα (A) and TNFα vs. TNFα + Apigenin (B) in MDA-MB-231 cells treated for 24 hours. The data show no significant changes at P < .05 for either analysis, with non-significant differences for only IL-1α.
Table 1. NF-kB PCR microarray tabulated data on MDA-MB-231 cells ± TNFα ± Apigenin.
The data displays differential transcription as Log2 (Fc) for controls vs. TNFα (Left Panel) and TNFα vs. TNFα + Apigenin (Right Panel) in MDA-MB-231 cells treated for 24 hours. The data show no significant changes at P < .05 for either analysis, with non-significant differences for only IL-1α.
| TNF-alpha vs Controls | TNF-alpha vs TNF-alpha+Apigenin | ||||
|---|---|---|---|---|---|
| Symbol | Log2(Fc) | p Value | Symbol | Log2(Fc) | p Value |
| IL1A | 3.47 | 0.14 | IL1A | -2.555 | 0.121 |
| AKT1 | -0.03 | 0.83 | AKT1 | 0.088 | 0.769 |
| AKT2 | -0.36 | 0.01 | AKT2 | -0.122 | 0.6 |
| AKT3 | -0.27 | 0.92 | AKT3 | 0.415 | 0.975 |
| CD14 | -0.8 | 0.17 | CD14 | -0.482 | 0.176 |
| CD28 | -0.29 | 0.81 | CD28 | 0.168 | 0.923 |
| CHUK | 0.27 | 0.69 | CHUK | 0.655 | 0.833 |
| IKBKB | -0.07 | 0.79 | IKBKB | 0.851 | 0.173 |
| IKBKG | 0.84 | 0.24 | IKBKG | 0.375 | 0.619 |
| IL1R1 | 0.46 | 0.55 | IL1R1 | -0.019 | 0.705 |
| IL1RAP | 0.35 | 0.58 | IL1RAP | -0.169 | 0.616 |
| IRAK1 | -0.6 | 0.33 | IRAK1 | -0.445 | 0.507 |
| IRAK2 | 0.6 | 0.43 | IRAK2 | 0.515 | 0.334 |
| IRAK3 | 0.98 | 0.35 | IRAK3 | -1.222 | 0.201 |
| LBP | -0.29 | 0.81 | LBP | 0.168 | 0.923 |
| LTA | 1.12 | 0.76 | LTA | 0.488 | 0.225 |
| LTBR | 0.63 | 0.34 | LTBR | -0.372 | 0.584 |
| LY96 | 0.52 | 0.6 | LY96 | 0.755 | 0.776 |
| MAP3K1 | -0.44 | 0.82 | MAP3K1 | 0.138 | 0.846 |
| MYD88 | -0.52 | 0.36 | MYD88 | 0.421 | 0.371 |
| NKKB1 | 0.45 | 0.65 | NKKB1 | 0.328 | 0.609 |
| NFKB2 | 0.57 | 0.1 | NFKB2 | 0.778 | 0.055 |
| NFKBIA | 1.04 | 0.18 | NFKBIA | 0.105 | 0.687 |
| NFKBIB | 0.48 | 0.22 | NFKBIB | 0.625 | 0.092 |
| NFKBIE | 0.84 | 0.45 | NFKBIE | 0.348 | 0.656 |
| PRKCQ | -0.29 | 0.81 | PRKCQ | 0.168 | 0.923 |
| REL | 0.53 | 0.6 | REL | 0.665 | 0.956 |
| RELA | -0.43 | 0.16 | RELA | 0.605 | 0.033 |
| RELB | 1.45 | 0.08 | RELB | 0.395 | 0.222 |
| RIPK2 | 0.38 | 0.67 | RIPK2 | 0.598 | 0.72 |
| SUMO1 | 0.06 | 0.95 | SUMO1 | -0.299 | 0.927 |
| TBP | -0.13 | 0.74 | TBP | 0.558 | 0.946 |
| TLR4 | 0.09 | 0.98 | TLR4 | -0.149 | 0.587 |
| TNF | 0.37 | 0.95 | TNF | 0.215 | 0.955 |
| TNFRSF1A | -0.16 | 0.57 | TNFRSF1A | 0.118 | 0.692 |
| TNFRSF1B | 0.49 | 0.47 | TNFRSF1B | -0.465 | 0.255 |
| TRADD | -0.54 | 0.31 | TRADD | 0.045 | 0.959 |
| TRAF2 | 0.2 | 0.2 | TRAF2 | 0.438 | 0.012 |
| TRAF6 | 0.35 | 0.78 | TRAF6 | -0.129 | 0.728 |
| UBB | -0.82 | 0.1 | UBB | -0.295 | 0.167 |
| UBC | -0.12 | 0.92 | UBC | -0.135 | 0.685 |
Fig 7.

A. IKBKe transcription in MDA-MB-231 cells ± TNFα ± Apigenin. The data represent normalized expression and are expressed as the Mean ± S.E.M., n = 3. The significance of differences between the Ctrl and TNFα groups and TNFα vs. TNFα + Apigenin were determined by a Students t-test *p<0.05. Figure 7B. Protein expression of IKBKe/GAPDH with TNFα ± Apigenin at 24 hours.
Fig 8. Protein expression of ERK 1 2 (total and phosphorylated) and GAPDH in MDA-MB-231 cells ± TNFα ± Apigenin.

Discussion
The findings in this study describe another anti-cancer property of apigenin to add to the arsenal of existing research having shown its ability to attenuate tumor promoting enzymes [1,2], prevent chemoresistance/multi-drug resistance [3–5], halt angiogenesis [6] lessening VEGF, inhibiting aromatase, proteasomal processes and fatty acid synthase, [18–21] and an overall reduction in proliferation, migration, and invasion of aggressive breast cancer [7]. Here we demonstrate the ability of apigenin to block TNFα mediated release of pro-inflammatory cytokines which are themselves responsible for inward migration of leukocytes that drive tumor growth, metastatic invasion [27], stem cell survival and immune evasion [23].
TNFα evoked release of tumor-promoting chemokines such as CCL2 (MCP-1), CCL5 (RANTES) and CXCL8 (IL-8) [26] trigger infiltration of CCR2/CCR5 receptor bearing leukocyte sub-populations (LSPs) including TAMs [29], MDSCs [30], TANs[31,32], Tregs [33], MAMs and CAFs [34]. While all LSP subtypes enable a tumor permissive and the evasive immune environment [41], independently, each LSP can confer a unique independent effect. For example, the arrival of MDSCs leads to potent immunosuppression by disabling the normal function of T and NK cells [42]. Whereas, the arrival of TANS perpetuates a sustained release of CCL2, GM-CSF, CCL17 which correspond to elevated tumor size, microvascular invasion and inward migratory activity of the macrophages and Tregs [43]. Reported results on Tregs indicate a direct role in suppression of antitumor immunity by attenuating the T-cell-mediated tumor cell killing [33] and secreting TGF-beta1 [44,45]. Meanwhile, the collective activity of TANS and MDSCS both yield the perpetual release of CCL2 and M-CSF fostering synergistic infiltration of TAMS. TAMS, upon arrival, can then lead to even more release of promoting cytokines: TGF-beta, MMPs, VEGF, CM/M-CSF, CCL 17, IL-13 and CCL2 which secure pro-angiogenic tumor processes [46]. In aggressive malignancies, metastatic MAMs arrive at secondary sites and in turn recruit inflammatory CCR2 bearing monocytes by CCL2, which in themselves release CCL3 and express CCR1, amplifying tumor potential [47]. LSP tumor infiltration is a vastly complex topic, yet all of these studies suggest the importance of controlling the chemokines produced and released by tumor tissue.
The current study shows that apigenin can downregulate TNFα mediated release of CCL2, largely attributable to its ability to downregulate IKBKe. The importance of IKBKe in driving tumor malignancy potential in both TNBC [39] and receptor positive breast cancer cells are known [40]. Moreover, there is a correlation between concentration of IKBKe and activated JAK/SAT[39] TRAF2 [48] Akt/PI3K [49] phosphorylated p65-NFkB [40] and NF-kB activation [50]. Likewise, numerous studies show many of these signaling pathways are blocked by apigenin in tumor models [17] in particular NF-kB signaling [51,52] which controls the downstream release of TGF-b [53] and activity of MMP-9 [54], themselves involved with TAM/TANs [55–57] and Treg recruitment [44,45].
The capacity of apigenin to block CCL2 is important because this single event is not only in control of infiltrating/migratory activity of CAFs, TAMS, TANS, and MSCs into the tumor environment but enables its perpetual ongoing accelerated release of CCL2, CXCL8, CCL5, mediated by through NF-kB signaling [24]. There is sufficient evidence to show that initial release of chemokines such as CCL2, GM-CSF [58] mark a potentially irreversible turning point for tumor infiltration, immunological evasion, and tumor immune support [29,59,60]. Natural plant-derived chemicals that can downregulate CCL2 or act as CCR2 receptor antagonists [61] such as luteolin [62] esculetin [63] or EGCG [64,65] typically result in impaired migration, less proclivity for metastasis [66] enabling greater efficacy of immunotherapies and chemotherapy drugs [65].
In summary, these findings show that apigenin can block TNFα mediated release of CCL2 in a TNBC cell line. While the experimental evidence for the therapeutic application of apigenin in cancer treatment is growing, human clinical trials are lacking [67]. Future studies will be required to determine if apigenin can be used clinically to establish long-term remission in cancer patients.
Materials and methods
Cell line, chemicals, and reagents
Triple-negative human breast tumor (MDA-MB-231) cells were obtained from the American Type Culture Collection (Rockville, MD, USA). Dulbecco’s modified Eagle’s medium (DMEM), fetal bovine serum (FBS) and penicillin/streptomycin were all obtained from Invitrogen (Carlsbad, CA, USA). Recombinant human TNFα was purchased from RayBiotech (RayBiotech Inc., Norcross, GA, USA). Apigenin was purchased from Abcam (Cambridge, United Kingdom).
Cell culture
MDA-MB-231 cells were cultured in 75 cm2 or 175 cm2 flasks containing DMEM supplemented with 10% FBS and 1% 10,000 U/ml penicillin G sodium/10,000 μg/ml streptomycin sulfate. Cells were grown at 37°C with humidified 95% air and 5% CO2.
Cell viability assay
Alamar Blue cell viability assay was used to determine cytotoxicity. Viable cells are capable of reducing resazurin to resorufin, resulting in fluorescence changes. Briefly, 96-well plates were seeded with MDA-MB-231 cells at a density of 5×104cells/100 μl/well. Cells were treated without or with either apigenin (10 μM, 20 μM, 30 μM, 40 μM 50 uM, 60 μM 70 μM 80 μM 90 μM 100 μM) or TNFα (0.1, 1, 10, 20, 40, 80, 100 ng/ml) for 24 h at 37°C, 5% CO2. Alamar blue (0.1 mg/ml in HBSS) was added at 15% v/v to each well and incubated for 6–8 hrs. Quantitative analysis of dye conversion was measured on a microplate fluorometer–Model 7620-version 5.02 (Cambridge Technologies Inc, Watertown, MA, USA) set at 550/580 (excitation/ emission). The data were expressed as a percentage of live untreated controls.
Human adipokine array
Sandwich-based arrays purchased from RayBiotech (Norcross, GA, USA) consist of membranes with 62 different proteins in duplicate. Each experiment was carried out by manufacturer’s instructions. Briefly, antibody-coated array membranes were treated with 1 ml of medium from resting, apigenin-treated (40 μM), TNFα-treated (40 ng) and co-treated cells and incubated overnight at 4°C on a rocker/shaker. The medium was decanted, the membranes were washed with wash buffer and then incubated with 1 ml biotin-conjugated antibodies (overnight 4°C). The mixture of biotin-conjugated antibodies was removed, and membranes were incubated with horseradish peroxidase–conjugated streptavidin (2 h). After a final wash, membrane intensity was acquired using chemiluminescence and analyzed using Quantity One software (Biorad Laboratories, Hercules. CA, US). Densities were calculated by subtracting blanks values from each array, then calculating all values as a percentage of the positive control spots on each membrane multiplied by an arbitrary averaged control values across all arrays in one individual test set. The data are represented as Density (Intensity (INT)/MM2).
CCL2 and IL-1α detection by ELISA
Supernatants from resting and stimulated (24 h) MDA-MB-231 cells were collected and centrifuged at 100× g for 5 min at 4°C. Specific ELISAs were performed using MCP-1/ CCL2 ELISA kits (RayBiotech) following the manufacturer’s instructions. Briefly, 100 μl of supernatants from samples and standards were added to 96-well plates pre-coated with capture antibody. After incubation, 100 μl of prepared biotinylated antibody mixture was added to each well. After one hour, the mixture was decanted, and 100 μl streptavidin solution was placed in each well and incubated. Substrate reagent (100 μl) was then added to each well followed by the addition of 50 μl stop solution 30 min later. The plate was read at 450 nm using UV microplate reader.
RT-PCR and RT-RCR NF-kB signaling pathway
MDA-MB-231 cells were lysed with 1 ml Trizol reagent. Chloroform (0.2 ml) was added samples were vortexed, incubated at 15–30°C for 2–3 min and centrifuged at 10,000 x g for 15 min. at 2–8°C. The aqueous phase was transferred to a fresh tube, and the RNA precipitated by mixing 0.5 ml isopropyl alcohol. RNA was extracted and subject to iScript advanced reverse transcriptase (RT) to the reaction. The reverse transcription was performed for 30 min at 42°C and RT inactivation for 5 min. at 85°C. PCR reaction. The following components were mixed in a 0.5 ml PCR tube: 5.0 μl cDNA product, 10 μl Ss Advanced Universal SYBR®Green, 1.0 μl primer and 4 μl water. PCR was performed with 39 cycles of denaturation: 15 sec. at 95°C; annealing: 30 sec. at 60°C; and extension 60 sec. at 72°C using Bio-Rad CFX96 Real-Time System (Hercules, Ca, USA). cDNA synthesis and Real-Time PCR was performed using iScript Advanced synthesis kit / SsAdvanced Universal SYBR® Green according to manufacturer's instructions. RT-PCR for IKBKe was run normalized to GAPDH mRNA, and the normalized values for the NF-kB Signaling array are as specified in Fig 6A. The array used was the transcription—NF-kB Signaling Pathway H96 Predesigned 96-well panel for use with SYBR® Green (Bio-Rad, Hercules, CA).
Western blot ERK1/2 and IKBKe
Total cell protein concentrations from MDA-MB-231 cells treated with apigenin, with and without TNFα co-treatment, was determined using a modified Bio-Rad “DC” protein assay (Bio-Rad Laboratories, Hercules, CA, USA). Cell lysates were separated by electrophoresis on 10% SDS-polyacrylamide gels and then transferred to Immobilon-P PVDF membranes. Blots were blocked at 4°C overnight in 5% bovine serum albumin (Sigma, St. Louis, MO, USA) in Tris-buffered saline with 0.05% Tween 20 in PBS (PBST) and then incubated overnight at 4°C with mouse anti-human ERK1/2 and IKBKe affinity purified antibody (Cell Signaling, Danvers, Ma, USA). Membranes were washed with PBST and incubated overnight with anti-goat IgG-horseradish peroxidase (Santa Cruz Biotechnology, CA) in PBST overnight at 4°C. Protein loading was monitored in each gel lane by probing the membranes with anti-GAPDH antibodies (Santa Cruz, Ca, USA). Immunoblot images were obtained using a Flour-S Max Multimager (Bio-Rad Laboratories, Hercules, CA). Lane density data was acquired with Quantity One Software (Bio-Rad Laboratories, Hercules, and CA).
Statistical analysis
Statistical analysis was performed using GraphPad Prism (version 3.0; GraphPad Software Inc. San Diego, CA, USA) with the significance of the difference between the groups assessed using a one-way ANOVA, followed by Tukey post hoc means comparison test, two-way ANOVA or Student’s t-test.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This research was supported by the National Institute of Minority Health and Health Disparities of the National Institutes of Health through Grant Number G12 MD007582 and Grant Number P20 MD006738.
References
- 1.Chen SS, Corteling R, Stevanato L, Sinden J (2012) Polyphenols inhibit indoleamine 3,5-dioxygenase-1 enzymatic activity—a role of immunomodulation in chemoprevention. Discov Med 14: 327–333. [PubMed] [Google Scholar]
- 2.(2005) Common spice may slow Alzheimer's. Health News 11: 2. [PubMed] [Google Scholar]
- 3.Xu Y, Xin Y, Diao Y, Lu C, Fu J, et al. (2011) Synergistic effects of apigenin and paclitaxel on apoptosis of cancer cells. PLoS One 6: e29169 doi: 10.1371/journal.pone.0029169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ferrucci V, Boffa I, De Masi G, Zollo M (2016) Natural compounds for pediatric cancer treatment. Naunyn Schmiedebergs Arch Pharmacol 389: 131–149. doi: 10.1007/s00210-015-1191-5 [DOI] [PubMed] [Google Scholar]
- 5.Angelini A, Di Ilio C, Castellani ML, Conti P, Cuccurullo F (2010) Modulation of multidrug resistance p-glycoprotein activity by flavonoids and honokiol in human doxorubicin- resistant sarcoma cells (MES-SA/DX-5): implications for natural sedatives as chemosensitizing agents in cancer therapy. J Biol Regul Homeost Agents 24: 197–205. [PubMed] [Google Scholar]
- 6.Mafuvadze B, Benakanakere I, Hyder SM (2010) Apigenin blocks induction of vascular endothelial growth factor mRNA and protein in progestin-treated human breast cancer cells. Menopause 17: 1055–1063. doi: 10.1097/gme.0b013e3181dd052f [DOI] [PubMed] [Google Scholar]
- 7.Mafuvadze B, Liang Y, Besch-Williford C, Zhang X, Hyder SM (2012) Apigenin induces apoptosis and blocks growth of medroxyprogesterone acetate-dependent BT-474 xenograft tumors. Horm Cancer 3: 160–171. doi: 10.1007/s12672-012-0114-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tseng TH, Chien MH, Lin WL, Wen YC, Chow JM, et al. (2016) Inhibition of MDA-MB-231 breast cancer cell proliferation and tumor growth by apigenin through induction of G2/M arrest and histone H3 acetylation-mediated p21WAF1/CIP1 expression. Environ Toxicol. [DOI] [PubMed] [Google Scholar]
- 9.Seo HS, Ku JM, Choi HS, Woo JK, Jang BH, et al. (2015) Apigenin induces caspase-dependent apoptosis by inhibiting signal transducer and activator of transcription 3 signaling in HER2-overexpressing SKBR3 breast cancer cells. Mol Med Rep 12: 2977–2984. doi: 10.3892/mmr.2015.3698 [DOI] [PubMed] [Google Scholar]
- 10.Choi EJ, Kim GH (2009) Apigenin causes G(2)/M arrest associated with the modulation of p21(Cip1) and Cdc2 and activates p53-dependent apoptosis pathway in human breast cancer SK-BR-3 cells. J Nutr Biochem 20: 285–290. doi: 10.1016/j.jnutbio.2008.03.005 [DOI] [PubMed] [Google Scholar]
- 11.Lin CH, Chang CY, Lee KR, Lin HJ, Chen TH, et al. (2015) Flavones inhibit breast cancer proliferation through the Akt/FOXO3a signaling pathway. BMC Cancer 15: 958 doi: 10.1186/s12885-015-1965-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Choi EJ, Kim GH (2009) Apigenin Induces Apoptosis through a Mitochondria/Caspase-Pathway in Human Breast Cancer MDA-MB-453 Cells. J Clin Biochem Nutr 44: 260–265. doi: 10.3164/jcbn.08-230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Harrison ME, Power Coombs MR, Delaney LM, Hoskin DW (2014) Exposure of breast cancer cells to a subcytotoxic dose of apigenin causes growth inhibition, oxidative stress, and hypophosphorylation of Akt. Exp Mol Pathol 97: 211–217. doi: 10.1016/j.yexmp.2014.07.006 [DOI] [PubMed] [Google Scholar]
- 14.Seo HS, Ju JH, Jang K, Shin I (2011) Induction of apoptotic cell death by phytoestrogens by up-regulating the levels of phospho-p53 and p21 in normal and malignant estrogen receptor alpha-negative breast cells. Nutr Res 31: 139–146. doi: 10.1016/j.nutres.2011.01.011 [DOI] [PubMed] [Google Scholar]
- 15.Cao X, Liu B, Cao W, Zhang W, Zhang F, et al. (2013) Autophagy inhibition enhances apigenin-induced apoptosis in human breast cancer cells. Chin J Cancer Res 25: 212–222. doi: 10.3978/j.issn.1000-9604.2013.04.01 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Seo HS, Jo JK, Ku JM, Choi HS, Choi YK, et al. (2015) Induction of caspase-dependent extrinsic apoptosis by apigenin through inhibition of signal transducer and activator of transcription 3 (STAT3) signalling in HER2-overexpressing BT-474 breast cancer cells. Biosci Rep 35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Seo HS, Choi HS, Kim SR, Choi YK, Woo SM, et al. (2012) Apigenin induces apoptosis via extrinsic pathway, inducing p53 and inhibiting STAT3 and NFkappaB signaling in HER2-overexpressing breast cancer cells. Mol Cell Biochem 366: 319–334. doi: 10.1007/s11010-012-1310-2 [DOI] [PubMed] [Google Scholar]
- 18.Nabavi SM, Habtemariam S, Daglia M, Nabavi SF (2015) Apigenin and Breast Cancers: From Chemistry to Medicine. Anticancer Agents Med Chem 15: 728–735. [DOI] [PubMed] [Google Scholar]
- 19.van Meeuwen JA, Korthagen N, de Jong PC, Piersma AH, van den Berg M (2007) (Anti)estrogenic effects of phytochemicals on human primary mammary fibroblasts, MCF-7 cells and their co-culture. Toxicol Appl Pharmacol 221: 372–383. doi: 10.1016/j.taap.2007.03.016 [DOI] [PubMed] [Google Scholar]
- 20.Noolu B, Gogulothu R, Bhat M, Qadri SS, Reddy VS, et al. (2016) In Vivo Inhibition of Proteasome Activity and Tumour Growth by Murraya koenigii Leaf Extract in Breast Cancer Xenografts and by its active flavonoids in breast cancer cells. Anticancer Agents Med Chem. [DOI] [PubMed] [Google Scholar]
- 21.Chen D, Landis-Piwowar KR, Chen MS, Dou QP (2007) Inhibition of proteasome activity by the dietary flavonoid apigenin is associated with growth inhibition in cultured breast cancer cells and xenografts. Breast Cancer Res 9: R80 doi: 10.1186/bcr1797 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang QR, Yao XQ, Wen G, Fan Q, Li YJ, et al. (2011) Apigenin suppresses the growth of colorectal cancer xenografts via phosphorylation and up-regulated FADD expression. Oncol Lett 2: 43–47. doi: 10.3892/ol.2010.215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu D, Wang X, Chen Z (2016) Tumor Necrosis Factor-alpha, a Regulator and Therapeutic Agent on Breast Cancer. Curr Pharm Biotechnol 17: 486–494. [DOI] [PubMed] [Google Scholar]
- 24.Katanov C, Lerrer S, Liubomirski Y, Leider-Trejo L, Meshel T, et al. (2015) Regulation of the inflammatory profile of stromal cells in human breast cancer: prominent roles for TNF-alpha and the NF-kappaB pathway. Stem Cell Res Ther 6: 87 doi: 10.1186/s13287-015-0080-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Trivanovic D, Jaukovic A, Krstic J, Nikolic S, Okic Djordjevic I, et al. (2016) Inflammatory cytokines prime adipose tissue mesenchymal stem cells to enhance malignancy of MCF-7 breast cancer cells via transforming growth factor-beta1. IUBMB Life 68: 190–200. doi: 10.1002/iub.1473 [DOI] [PubMed] [Google Scholar]
- 26.Soria G, Ofri-Shahak M, Haas I, Yaal-Hahoshen N, Leider-Trejo L, et al. (2011) Inflammatory mediators in breast cancer: coordinated expression of TNFalpha & IL-1beta with CCL2 & CCL5 and effects on epithelial-to-mesenchymal transition. BMC Cancer 11: 130 doi: 10.1186/1471-2407-11-130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ben-Baruch A (2012) The Tumor-Promoting Flow of Cells Into, Within and Out of the Tumor Site: Regulation by the Inflammatory Axis of TNFalpha and Chemokines. Cancer Microenviron 5: 151–164. doi: 10.1007/s12307-011-0094-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Papi A, Storci G, Guarnieri T, De Carolis S, Bertoni S, et al. (2013) Peroxisome proliferator activated receptor-alpha/hypoxia inducible factor-1alpha interplay sustains carbonic anhydrase IX and apoliprotein E expression in breast cancer stem cells. PLoS One 8: e54968 doi: 10.1371/journal.pone.0054968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Steiner JL, Murphy EA (2012) Importance of chemokine (CC-motif) ligand 2 in breast cancer. Int J Biol Markers 27: e179–185. doi: 10.5301/JBM.2012.9345 [DOI] [PubMed] [Google Scholar]
- 30.Vrakas CN, O'Sullivan RM, Evans SE, Ingram DA, Jones CB, et al. (2015) The Measure of DAMPs and a role for S100A8 in recruiting suppressor cells in breast cancer lung metastasis. Immunol Invest 44: 174–188. doi: 10.3109/08820139.2014.952818 [DOI] [PubMed] [Google Scholar]
- 31.Tabaries S, Ouellet V, Hsu BE, Annis MG, Rose AA, et al. (2015) Granulocytic immune infiltrates are essential for the efficient formation of breast cancer liver metastases. Breast Cancer Res 17: 45 doi: 10.1186/s13058-015-0558-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ibrahim SA, Katara GK, Kulshrestha A, Jaiswal MK, Amin MA, et al. (2015) Breast cancer associated a2 isoform vacuolar ATPase immunomodulates neutrophils: potential role in tumor progression. Oncotarget 6: 33033–33045. doi: 10.18632/oncotarget.5439 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kindlund B, Sjoling A, Yakkala C, Adamsson J, Janzon A, et al. (2016) CD4 regulatory T cells in gastric cancer mucosa are proliferating and express high levels of IL-10 but little TGF-beta. Gastric Cancer. [DOI] [PubMed] [Google Scholar]
- 34.Mishra P, Banerjee D, Ben-Baruch A (2011) Chemokines at the crossroads of tumor-fibroblast interactions that promote malignancy. J Leukoc Biol 89: 31–39. doi: 10.1189/jlb.0310182 [DOI] [PubMed] [Google Scholar]
- 35.Ueno T, Toi M, Saji H, Muta M, Bando H, et al. (2000) Significance of macrophage chemoattractant protein-1 in macrophage recruitment, angiogenesis, and survival in human breast cancer. Clin Cancer Res 6: 3282–3289. [PubMed] [Google Scholar]
- 36.Chun E, Lavoie S, Michaud M, Gallini CA, Kim J, et al. (2015) CCL2 Promotes Colorectal Carcinogenesis by Enhancing Polymorphonuclear Myeloid-Derived Suppressor Cell Population and Function. Cell Rep 12: 244–257. doi: 10.1016/j.celrep.2015.06.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.McClellan JL, Davis JM, Steiner JL, Enos RT, Jung SH, et al. (2012) Linking tumor-associated macrophages, inflammation, and intestinal tumorigenesis: role of MCP-1. Am J Physiol Gastrointest Liver Physiol 303: G1087–1095. doi: 10.1152/ajpgi.00252.2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Qian BZ, Li J, Zhang H, Kitamura T, Zhang J, et al. (2011) CCL2 recruits inflammatory monocytes to facilitate breast-tumour metastasis. Nature 475: 222–225. doi: 10.1038/nature10138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Barbie TU, Alexe G, Aref AR, Li S, Zhu Z, et al. (2014) Targeting an IKBKE cytokine network impairs triple-negative breast cancer growth. J Clin Invest 124: 5411–5423. doi: 10.1172/JCI75661 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Jiang Z, Liu JC, Chung PE, Egan SE, Zacksenhaus E (2014) Targeting HER2(+) breast cancer: the TBK1/IKKepsilon axis. Oncoscience 1: 180–182. doi: 10.18632/oncoscience.18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Fujimoto H, Sangai T, Ishii G, Ikehara A, Nagashima T, et al. (2009) Stromal MCP-1 in mammary tumors induces tumor-associated macrophage infiltration and contributes to tumor progression. Int J Cancer 125: 1276–1284. doi: 10.1002/ijc.24378 [DOI] [PubMed] [Google Scholar]
- 42.Malek E, de Lima M, Letterio JJ, Kim BG, Finke JH, et al. (2016) Myeloid-derived suppressor cells: The green light for myeloma immune escape. Blood Rev. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhou SL, Zhou ZJ, Hu ZQ, Huang XW, Wang Z, et al. (2016) Tumor-Associated Neutrophils Recruit Macrophages and T-Regulatory Cells to Promote Progression of Hepatocellular Carcinoma and Resistance to Sorafenib. Gastroenterology. [DOI] [PubMed] [Google Scholar]
- 44.Wang Y, Liu T, Tang W, Deng B, Chen Y, et al. (2016) Hepatocellular Carcinoma Cells Induce Regulatory T Cells and Lead to Poor Prognosis via Production of Transforming Growth Factor-beta1. Cell Physiol Biochem 38: 306–318. doi: 10.1159/000438631 [DOI] [PubMed] [Google Scholar]
- 45.Sun L, Jin H, Li H (2016) GARP: a surface molecule of regulatory T cells that is involved in the regulatory function and TGF-beta releasing. Oncotarget. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rego SL, Helms RS, Dreau D (2014) Breast tumor cell TACE-shed MCSF promotes pro-angiogenic macrophages through NF-kappaB signaling. Angiogenesis 17: 573–585. doi: 10.1007/s10456-013-9405-2 [DOI] [PubMed] [Google Scholar]
- 47.Kitamura T, Qian BZ, Soong D, Cassetta L, Noy R, et al. (2015) CCL2-induced chemokine cascade promotes breast cancer metastasis by enhancing retention of metastasis-associated macrophages. J Exp Med 212: 1043–1059. doi: 10.1084/jem.20141836 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Shen RR, Zhou AY, Kim E, Lim E, Habelhah H, et al. (2012) IkappaB kinase epsilon phosphorylates TRAF2 to promote mammary epithelial cell transformation. Mol Cell Biol 32: 4756–4768. doi: 10.1128/MCB.00468-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Guo JP, Coppola D, Cheng JQ (2011) IKBKE protein activates Akt independent of phosphatidylinositol 3-kinase/PDK1/mTORC2 and the pleckstrin homology domain to sustain malignant transformation. J Biol Chem 286: 37389–37398. doi: 10.1074/jbc.M111.287433 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 50.Zhou AY, Shen RR, Kim E, Lock YJ, Xu M, et al. (2013) IKKepsilon-mediated tumorigenesis requires K63-linked polyubiquitination by a cIAP1/cIAP2/TRAF2 E3 ubiquitin ligase complex. Cell Rep 3: 724–733. doi: 10.1016/j.celrep.2013.01.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Shukla S, Shankar E, Fu P, MacLennan GT, Gupta S (2015) Suppression of NF-kappaB and NF-kappaB-Regulated Gene Expression by Apigenin through IkappaBalpha and IKK Pathway in TRAMP Mice. PLoS One 10: e0138710 doi: 10.1371/journal.pone.0138710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Shukla S, Kanwal R, Shankar E, Datt M, Chance MR, et al. (2015) Apigenin blocks IKKalpha activation and suppresses prostate cancer progression. Oncotarget 6: 31216–31232. doi: 10.18632/oncotarget.5157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Mirzoeva S, Franzen CA, Pelling JC (2014) Apigenin inhibits TGF-beta-induced VEGF expression in human prostate carcinoma cells via a Smad2/3- and Src-dependent mechanism. Mol Carcinog 53: 598–609. doi: 10.1002/mc.22005 [DOI] [PubMed] [Google Scholar]
- 54.Chunhua L, Donglan L, Xiuqiong F, Lihua Z, Qin F, et al. (2013) Apigenin up-regulates transgelin and inhibits invasion and migration of colorectal cancer through decreased phosphorylation of AKT. J Nutr Biochem 24: 1766–1775. doi: 10.1016/j.jnutbio.2013.03.006 [DOI] [PubMed] [Google Scholar]
- 55.Moses K, Brandau S (2016) Human neutrophils: Their role in cancer and relation to myeloid-derived suppressor cells. Semin Immunol. [DOI] [PubMed] [Google Scholar]
- 56.Yan C, Huo X, Wang S, Feng Y, Gong Z (2015) Stimulation of hepatocarcinogenesis by neutrophils upon induction of oncogenic kras expression in transgenic zebrafish. J Hepatol 63: 420–428. doi: 10.1016/j.jhep.2015.03.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Deryugina EI, Zajac E, Juncker-Jensen A, Kupriyanova TA, Welter L, et al. (2014) Tissue-infiltrating neutrophils constitute the major in vivo source of angiogenesis-inducing MMP-9 in the tumor microenvironment. Neoplasia 16: 771–788. doi: 10.1016/j.neo.2014.08.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Jehs T, Faber C, Juel HB, Bronkhorst IH, Jager MJ, et al. (2014) Inflammation-induced chemokine expression in uveal melanoma cell lines stimulates monocyte chemotaxis. Invest Ophthalmol Vis Sci 55: 5169–5175. doi: 10.1167/iovs.14-14394 [DOI] [PubMed] [Google Scholar]
- 59.Schmall A, Al-Tamari HM, Herold S, Kampschulte M, Weigert A, et al. (2015) Macrophage and cancer cell cross-talk via CCR2 and CX3CR1 is a fundamental mechanism driving lung cancer. Am J Respir Crit Care Med 191: 437–447. doi: 10.1164/rccm.201406-1137OC [DOI] [PubMed] [Google Scholar]
- 60.Hollmen M, Roudnicky F, Karaman S, Detmar M (2015) Characterization of macrophage—cancer cell crosstalk in estrogen receptor positive and triple-negative breast cancer. Sci Rep 5: 9188 doi: 10.1038/srep09188 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yumimoto K, Akiyoshi S, Ueo H, Sagara Y, Onoyama I, et al. (2015) F-box protein FBXW7 inhibits cancer metastasis in a non-cell-autonomous manner. J Clin Invest 125: 621–635. doi: 10.1172/JCI78782 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Choi HJ, Choi HJ, Chung TW, Ha KT (2016) Luteolin inhibits recruitment of monocytes and migration of Lewis lung carcinoma cells by suppressing chemokine (C-C motif) ligand 2 expression in tumor-associated macrophage. Biochem Biophys Res Commun 470: 101–106. doi: 10.1016/j.bbrc.2016.01.002 [DOI] [PubMed] [Google Scholar]
- 63.Kimura Y, Sumiyoshi M (2015) Antitumor and antimetastatic actions of dihydroxycoumarins (esculetin or fraxetin) through the inhibition of M2 macrophage differentiation in tumor-associated macrophages and/or G1 arrest in tumor cells. Eur J Pharmacol 746: 115–125. doi: 10.1016/j.ejphar.2014.10.048 [DOI] [PubMed] [Google Scholar]
- 64.Devaud C, John LB, Westwood JA, Yong CS, Beavis PA, et al. (2015) Cross-talk between tumors can affect responses to therapy. Oncoimmunology 4: e975572 doi: 10.4161/2162402X.2014.975572 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Zollo M, Di Dato V, Spano D, De Martino D, Liguori L, et al. (2012) Targeting monocyte chemotactic protein-1 synthesis with bindarit induces tumor regression in prostate and breast cancer animal models. Clin Exp Metastasis 29: 585–601. doi: 10.1007/s10585-012-9473-5 [DOI] [PubMed] [Google Scholar]
- 66.Jang JY, Lee JK, Jeon YK, Kim CW (2013) Exosome derived from epigallocatechin gallate treated breast cancer cells suppresses tumor growth by inhibiting tumor-associated macrophage infiltration and M2 polarization. BMC Cancer 13: 421 doi: 10.1186/1471-2407-13-421 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Shukla S, Gupta S (2010) Apigenin: a promising molecule for cancer prevention. Pharm Res 27: 962–978. doi: 10.1007/s11095-010-0089-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.