

Abstract
Background
Autonomic dysfunction is implicated in a range of psychological conditions, including depression and anxiety. The fragile X mental retardation-1 (FMR1) premutation is a common genetic mutation that affects ~1:150 women and is associated with psychological vulnerability. This study examined cardiac indicators of autonomic function among women with the FMR1 premutation and control women as potential biomarkers for psychological risk that may be linked to FMR1.
Methods
Baseline inter-beat interval and respiratory sinus arrhythmia (a measure of parasympathetic vagal tone) were measured in 35 women with the FMR1 premutation and 28 controls. The women completed anxiety and depression questionnaires. FMR1 genetic indices (i.e., CGG repeat, quantitative FMRP, FMR1 mRNA, activation ratio) were obtained for the premutation group.
Results
Respiratory sinus arrhythmia was reduced in the FMR1 premutation group relative to controls. While depression symptoms were associated with reduced respiratory sinus arrhythmia among control women, these variables were unrelated in the FMR1 premutation. Elevated FMR1 mRNA was associated with higher respiratory sinus arrhythmia.
Conclusions
Women with the FMR1 premutation demonstrated autonomic dysregulation characterized by reduced vagal tone. Unlike patterns observed in the general population and in study controls, vagal activity and depression symptoms were decoupled in women with the FMR1 premutation, suggesting independence between autonomic regulation and psychopathological symptoms that is atypical and potentially specific to the FMR1 premutation. The association between vagal tone and mRNA suggests that molecular variation associated with FMR1 plays a role in autonomic regulation.
Keywords: Fragile X carriers, Vagal tone, Heart rate, Physiological arousal, FMRP, FMR1 mRNA
Background
The autonomic nervous system plays a fundamental role in health. Working in conjunction with other stress regulation systems, such as the hypothalamic-pituitary-adrenal axis and the immune system, the autonomic nervous system promotes adaptability to life stressors while helping the body maintain a well-controlled, functional physiological state [1]. Optimally, the sympathetic (“fight or flight”) and parasympathetic (“rest and digest”) branches of the autonomic nervous system work together in a coordinated and often antagonistic fashion to effectively respond to internal and external demands. When the dynamic interplay between the sympathetic and parasympathetic nervous systems is functioning well, the autonomic system serves a broad protective role, boosting the immune system, shielding against cardiovascular disease, and warding away psychopathology [2]. Conversely, dysfunction of the autonomic nervous system is associated with vulnerability to a host of physical and mental health disorders.
The integrity of the autonomic system can be assessed objectively and non-invasively through peripheral measures of cardiac activity. The heart is innervated by the vagal nerve, which provides a pathway for brain-heart communication via connections in the brainstem and the sinoatrial node of the heart. The measurement of inter-beat interval (IBI; or the time between consecutive heart beats) provides an estimate of general arousal level influenced by both sympathetic and parasympathetic branches of the autonomic system [3]. A specific index of parasympathetic activity can be obtained by measuring heart rate variability patterns, which index parasympathetic influences on the heart via the vagal nerve [4]. Vagal tone can be estimated through descriptive measures of heart rate variability, as well as through the quantification of respiratory sinus arrhythmia (RSA), a measure of variability in the rise and fall of heart rate that occurs with respiration (see [5], for review).
Cardiac autonomic dysregulation in mood and anxiety disorders
Converging empirical and theoretical evidence supports cardiac autonomic indices as objective, non-invasive markers for mood and anxiety disorders (e.g., [6–9]). Dampened vagal tone is well documented in a major depressive disorder [10–13], and vagal level has also been shown to correlate with the continuous distribution of depression symptoms in non-clinical samples (e.g., [10, 14–16]). A number of reports demonstrate that depressed individuals with low vagal tone have greater symptom severity [17] and are less likely to recover or demonstrate symptomatic improvement [18, 19], and successful treatment for depression corresponds with vagal increases [20–22], albeit with some mixed findings (e.g., [23, 24]). Reduced cardiac vagal tone is also thought to represent a physiological pathway leading to anxiety. Low vagal tone relates to anxiety symptoms in non-clinical groups [25–27], and low vagal tone has been documented extensively among individuals with anxiety disorders, including populations affected by generalized anxiety disorder, panic disorder, and post-traumatic stress disorder (see [8, 9] for review).
Psychophysiological theories of vagal tone, such as the Polyvagal Theory [28, 29] and the Neurovisceral Integration Model [30, 31], support the integral role of the parasympathetic vagal system in emotional expression and regulation, accounting for reduced vagal level in mood and anxiety disorders—clinical conditions characterized by impaired emotional regulation [32]. A large body of literature suggests a mechanistic role of vagal tone in emotional regulation, coping, and social engagement [33, 34]. According to theory, vagal control is one component of a larger central autonomic network that serves to regulate defensive social behavior. The parasympathetic vagal system, in conjunction with other mechanisms, works to inhibit sympatho-excitatory threat circuits. When the vagal system is hypoactive, the body remains in a state of hypermobilization and defense, which increases “allostatic load,” or wear and tear to the bodily system over time [35]. The ability to inhibit threat circuits via the vagus is compromised in disorders of impaired emotional regulation, such as anxiety and depression (see [36]).
Cardiac indices as biomarkers for psychological risk
The identification of biomarkers holds promise for furthering the prevention and treatment for complex mental health conditions such as anxiety and depression. Biomarkers, or measurable, endogenous traits that mark either the risk or manifestation of psychiatric illness [37], allow clinical groups to be deconstructed at the biological level, thus yielding information relevant to (1) the development of treatments targeted towards core mechanisms rather than symptoms, (2) the stratification of biological subgroups who are mostly likely to respond to targeted interventions, and (3) the identification of individuals who are most at risk, perhaps even before the onset of clinical symptoms [38]. A number of prior studies have put forth cardiac indicators of autonomic dysfunction as potentially useful biomarkers for anxiety and depression (e.g., [39, 40]) given that they co-occur with the clinical presentation of mood and anxiety disorders, are associated with symptoms in non-clinical samples, represent heritable and stable traits, are quantitative, and can be measured non-invasively and relatively quickly [41–44]. Thus, the study of cardiac activity in relation to depression and anxiety may prove useful in understanding the biological bases of these mental health conditions.
The FMR1 premutation as a genetic model for psychological risk
Studying cardiac function within high-risk genetic groups can inform the intercorrelation between psychophysiological traits and unique genetic profiles. In this regard, the fragile X mental retardation-1 (FMR1) premutation represents a particularly promising condition for study. This genetic condition is linked with significant psychological risk and may hold promise for uncovering genetic determinants for autonomic alterations. The FMR1 premutation occurs when the trinucleotide (CGG) sequence on the FMR1 gene of the X chromosome expands to 55–200 repeats [45]. This mutation is characterized by excess production of FMR1 messenger RNA (mRNA), which causes neuronal toxicity [46–48]. The FMR1 premutation expansion is highly prevalent, occurring in approximately 1 in 113–250 females and 1 in 250–810 males depending on ethnicity and world region [49–53]. Individuals with the FMR1 premutation are at risk for passing the mutated gene to their children, which may undergo further expansions as it is transmitted through generations, increasing the severity of the disease. Risk for generational expansion is related to genetic factors such as CGG repeat size and the number of AGG anchors, as well as environmental factors such as maternal age [54, 55]. When the expansion extends beyond 200 CGG repeats, the gene becomes inactivated by methylation and fragile X syndrome results, a neurodevelopmental disorder affecting approximately 1 in 5000 individuals [56] that is associated with intellectual disability and autism spectrum disorder [57]. The present study focuses on women with the FMR1 premutation, who show a well-documented psychological profile characterized by risk for depression and anxiety disorders. Research focused on women—and in particular mothers—with the FMR1 premutation is important given that the FMR1 premutation phenotype is associated with negative outcomes both for the affected individual as well as for their children with fragile X syndrome (e.g., [58, 59]).
Women with the FMR1 premutation, referred to as “carriers” of fragile X, were once thought to be clinically unaffected; however, new evidence clearly supports clinical involvement in this group [60]. This includes risk for fragile X-specific conditions such as fragile X-associated primary ovarian insufficiency [61] and fragile X-associated tremor/ataxia syndrome (FXTAS), a late-onset neurodegenerative movement disorder characterized by tremors, gait ataxia, peripheral neuropathy, executive dysfunction, and cognitive decline that affects about 16% of women with the premutation [62]. A subset of women with the premutation may also present with certain cognitive deficits related to executive functioning, working memory [63], and symptoms of attention deficit-hyperactivity disorder [64], which may worsen with age [65, 66]. Social difficulties have also been documented in females with the FMR1 premutation, such as social-language deficits [58, 67] and elevated rates of autism spectrum disorder [68]. Finally, a higher rate of immune-mediated disorders, sleep apnea, hypertension, migraines, and seizures has also been observed in individuals with the premutation (reviewed in [69]).
Psychological risk in the FMR1 premutation
Elevated rates of mood and anxiety disorders are one of the earliest and most consistently documented features of the FMR1 premutation phenotype, with the risk for these conditions increasing significantly over time during adulthood [70]. Reported lifetime rates of major depressive disorder range from 12 to 54% in females with the premutation [70–75]. Lifetime rates of any anxiety disorder ranges from 25 to 47% [72, 74, 76]. This includes elevated lifetime rates of panic disorder [74, 75], social phobia [72, 74], and post-traumatic stress disorder [74]; although, findings vary somewhat depending on sample characteristics and diagnostic instruments. Reported rates for current occurrence range from 5 to 13% for major depressive disorder [70, 75] and 13 to 50% for anxiety disorders [75–77].
Psychological risk in women with the FMR1 premutation likely has a multifactorial basis, with both FMR1 gene dysfunction and environmental factors, such as child-related challenges, mechanistically implicated in an additive or interactive manner. Chronic stressors associated with raising a child with a developmental disorder, such as elevated child problem behaviors, are linked to increased likelihood of anxiety disorders and major depression in women with the FMR1 premutation [75, 78]. Yet, mental health problems in women with the FMR1 premutation often proceed the birth of their child with fragile X syndrome [75] and women with the FMR1 premutation who do not have a child affected by fragile X syndrome also show increased rates of psychological disorders [79], supporting genetic contributors to psychopathological risk that are independent of child-related stressors. Studies have begun to characterize specific FMR1 genetic markers associated with psychiatric symptoms. A number of reports have documented that risk for depression in the FMR1 premutation is highest among individuals with CGG repeat length within the midsize range [70, 75, 80, 81]. Seltzer et al. [81] also detected CGG-dependent sensitivity to the environmental context, where women with midsize CGG repeat length and above-average life stress showed greater vulnerability for depression and anxiety compared to women with higher or lower repeat lengths, whereas women with midsize CGG repeats and below-average life stress were the most resilient to depression and anxiety.
Psychological vulnerability may also be related to increased FMR1 mRNA expression, which is present at up to eightfold normal levels and increases linearly with CGG repeat size in the premutation [82, 83]. Elevated mRNA levels were found to be associated with increased psychological symptoms and reduced amygdala activation in males with the premutation, and these associations were present even among male carriers without FXTAS, suggesting that the impact of mRNA toxicity is not exclusive to FXTAS [84, 85]. Levels of mRNA are also linked with the age of depression onset in individuals with the FMR1 premutation, consistent with the hypothesis that mRNA toxicity builds over time, contributing to vulnerability [86]. Females may be more protected from mRNA toxicity, due to the presence of the second X chromosome. In females, a high activation ratio, or a high proportion of cells carrying the normal allele on the active X chromosome, can dilute the effects of the premutation allele [87] and has been associated with less severe clinical effects, such as lower parenting stress [88] and more typical patterns of cortisol stress responses [89]. Higher levels of mRNA are correlated with self-reported anxiety symptoms among women with the premutation, but only when the sample was restricted to women with an activation ratio of less than 0.5 [84].
Slightly reduced levels of fragile X mental retardation protein (FMRP) have also been reported among individuals with the FMR1 premutation, particularly among individuals with high CGG repeats [90, 91]. FMRP is an mRNA-binding protein that regulates the translation of about one-third of the proteins in the pre- and post-synaptic proteomes, supporting its critical role in synaptic plasticity and the development and maintenance of neuronal circuits [92]. Its absence is thought to underlie the neurobehavioral impairments seen in the full mutation [93]. Yet, the phenotypic impact of reduced FMRP in the FMR1 premutation is less clear. Until recently, FMRP level has been measured indirectly (e.g., by counting the percent of FMRP-positive lymphocytes, see [94]), limiting the ability to capture subtle variation in protein expression. No relationships have been detected between the percentage of lymphocytes staining positive for FMRP and psychological symptoms in males or females with the FMR1 premutation [84]. However, new technological advances, such as the approach used in the present study, allow for quantitative measurement of actual FMRP levels in the blood and may lead to a new wave of discoveries into the role of FMRP in the clinical profile of the FMR1 premutation. For instance, studies using quantitative FMRP have revealed relationships with neurobehavioral profiles, such as preliminary evidence of FMRP-mediated blunted amygdala responses that are associated with deficient social information processing in men [85].
Despite the documented links between FMR1-related variation and depression in the FMR1 premutation, a number of studies have failed to detect molecular genetic correlates of anxiety in this group [70, 72, 77, 84, 86, 95, 96] and extant findings suggest a complex, multifactorial, epigenetic basis to anxiety symptom expression. Anxiety in women with the premutation has been linked with environmental factors such as child problem behaviors [75], with the impact of the stress of raising a child with fragile X syndrome moderated by variation on CRHR1, a gene involved in cortisol regulation [97]. Epigenetic changes associated with abnormal methylation have also been implicated, with one study showing that methylation of the CpG10-12 sites located at the FMR1 intron 1 boundary predicted social anxiety with 92% sensitivity in women with the FMR1 premutation [98]. Finally, anxiety in women with the premutation has been linked with neuroanatomical changes, specifically, with reduced hippocampal volume associated with elevated mRNA [99].
Autonomic function in the FMR1 premutation
Given the elevated risk for depression and anxiety in the FMR1 premutation and documented associations with FMR1-associated genetic mechanisms, the FMR1 premutation may represent a “portal” condition that can yield important information on the molecular genetic basis for autonomic alterations relevant to both individuals with and without FMR1 mutations. This work may also inform prevention and treatment efforts specific to the FMR1 premutation. The lack of useful biomarkers represents a critical barrier to targeted treatment for this group, given the incomplete penetrance of associated clinical effects. Should cardiac indicators account for inter-individual variability in psychological risk within this population, they may prove useful in identifying vulnerable subgroups who may benefit from targeted prevention efforts.
Although cardiac autonomic dysregulation is a robust, well-documented feature of fragile X syndrome (see [100], for review), no studies have examined cardiac autonomic integrity in the FMR1 premutation. Yet, the clinical effects of the FMR1 premutation are highly suggestive of autonomic impairment, such as increased rates of thyroid disorders, fibromyalgia, and hypertension—all conditions associated with autonomic dysfunction [101]. Moreover, symptoms consistent with autonomic dysfunction are common in FXTAS such as impotence, bowel and bladder incontinence, hypertension, and syncope [102]. Neuropathological involvement in the autonomic ganglion of the heart and autonomic neurons of the spinal cord has also been detected in postmortem studies of individuals with FXTAS [103, 104]. In the only study to date that employed direct measures of autonomic function in the premutation, Hessl and colleagues [105] detected dampened sympathetic reactivity to a social greeting task among a sample of 12 men with the premutation using measures of electrodermal response. No associations were detected between sympathetic activation and psychological symptoms or FMR1 molecular measures (CGG repeat size and mRNA); although, conclusions were preliminary given the small sample.
The present study
Further investigation of autonomic nervous system activity among individuals with the FMR1 premutation will help identify biophysiological pathways rooted in FMR1 gene dysfunction, shedding light on biomarkers that may be linked with clinical impairment in this group. This work has implications for the identification of at-risk individuals based on specific biological markers and the potential to shift treatment efforts away from symptom-based approaches to target specific underlying mechanisms. In sum, investigations into cardiac indicators of autonomic function may provide insight into the intermediate functions of the FMR1 gene that are coupled with psychological risk. The present study addressed the following questions:
Do cardiac markers of autonomic function (i.e., IBI and RSA) differ between women with the FMR1 premutation and control women at baseline? It was hypothesized that women with the FMR1 premutation would have elevated general arousal and reduced vagal tone when compared to controls, mirroring the physiological profile seen in the full mutation.
Are cardiac markers of autonomic function related to symptoms of depression and anxiety among women with the FMR1 premutation and control women? It was hypothesized that low baseline vagal tone and high general arousal would relate to increased psychological symptoms in both groups.
Are cardiac markers of autonomic function associated with FMR1-related genetic variation in women with the FMR1 premutation? Given the lack of prior research in this area, this aim was considered exploratory and specific hypotheses regarding gene-autonomic relationships were not made.
Methods
Participants
Participants included 35 women with the FMR1 premutation and 28 control women who were enrolled in a larger study of the social-language phenotype of women with the FMR1 premutation. Inclusionary criteria for the broader study specified that all participants were native speakers of English, were mothers, and did not have an intellectual disability (i.e., IQ composite >80 on the Kaufman Brief Intelligence Test-II; [106]). Women who were pregnant were excluded from the study to control for pregnancy-related physiological changes (e.g., [107]). The women with the FMR1 premutation were recruited through their children, who were participating in developmental studies of children with fragile X syndrome (PI’s: Abbeduto, Roberts). Genetic status of the women with the FMR1 premutation was confirmed through blood tests collected through this study (n = 31) or via medical records. The premutation was defined as an allele ranging from 55 to 200 CGG repeats on FMR1. Although it was beyond the scope of the present study to conduct genetic testing on control participants, 61% of controls completed genetic testing to rule out the FMR1 premutation through dual enrollment in a related study. Control women had no known family history of fragile X-associated conditions and were mothers of typically developing children (i.e., children who had not been diagnosed or treated for any type of developmental delay or disorder, per participant report). Additionally, control women were excluded from the study if their child scored above the cut-off for autism spectrum disorder on the Social Communication Questionnaire [108]. Recruitment of controls was focused in the local community using flyers, social media, and word of mouth.
Descriptive and demographic information is presented in Table 1. The groups did not differ significantly on age, IQ, race, or household income. A higher proportion of women with the FMR1 premutation were using psychotropic medications compared to the control women (48 vs 15%, p = 0.008). While the presence FXTAS was not an exclusionary criteria, none of the women reported a clinical diagnosis of FXTAS. The groups did not differ in self-reported functional symptoms of tremor measured with the Tremor Disability Questionnaire [109], p = 0.508. Information on menopause status was also collected from the women with the FMR1 premutation, as autonomic changes are observed among postmenopausal women (e.g., [110]) and the FMR1 premutation is linked with early menopause [111]. Fifty-eight percent of the women in the FMR1 premutation group had completed menopause, defined here as the cessation of menses for >1 year. Finally, the Parenting Stress Inventory-4 [112] was administered, given the reported relationships between parenting stress and maternal psychological health in other disability groups (e.g., [113]). Parenting stress was significantly elevated in the FMR1 premutation group (p = 0.001).
Table 1.
Group characteristics
| Variable | Group | ||
|---|---|---|---|
| Women with the FMR1 Premutation (n = 35) |
Control Women (n = 28) |
Test of group differences (p value) | |
| Age in years | |||
| M (SD) | 44.31(8.63) | 41.70 (9.34) | 0.251 |
| Range | 25.53–60.94 | 28.72–65.23 | |
| IQa | |||
| M (SD) | 104.26 (11.90) | 104.57 (11.46) | 0.928 |
| Range | 81.00–130.00 | 83.00–135.00 | |
| Race | 0.181 | ||
| Caucasian | 94% | 85% | |
| African American | 3% | 15% | |
| American Indian | 3% | – | |
| Household Income | |||
| <20k | 9% | 12% | 0.164 |
| 21–40k | 12% | 7% | |
| 41–80k | 33% | 35% | |
| 81–120k | 12% | 30% | |
| >121k | 34% | 11% | |
| Medication use | |||
| Atypical antipsychotics | 3% | – | 0.008* |
| Classical antipsychotics | 3% | – | |
| Antidepressants | 48% | 15% | |
| Mood stabilizers | 7% | – | |
| Anti-anxiety | 10% | – | |
| Stimulants | 3% | 4% | |
| Total stress percentileb | 0.001* | ||
| M (SD) | 62.12 (22.87) | 34.64 (25.63) | |
| Range | 4.00–96.00 | 1.00–88.00 | |
| Tremor Disability Scorec | 0.508 | ||
| M (SD) | 4.30 (15.29) | 2.06 (4.15) | |
| Range | 0–77.42 | 0–12.90 | |
Procedures
Assessments took place in a university laboratory setting. Baseline cardiac activity was the first assessment activity completed after consent was obtained. To control for the potential influences of circadian rhythm, assessments were conducted in the morning (generally starting at 9:00 a.m.). Participants were asked to refrain from drinking coffee for at least 1 h prior to the assessment. Procedures were approved by the Institutional Review Board of the University of South Carolina.
Measures
Cardiac autonomic activity
Cardiac activity was sampled during a 5-min baseline context where participants viewed a video of ocean waves that was designed for meditation and relaxation. Participants were instructed to “sit back and try to relax.” Data were analyzed from the final 3 min of viewing, which allowed additional time for participants to “settle into” the task. Cardiac data were collected with an Actiwave Cardio monitor (CamNtech Ltd., Cambridge, UK), which samples activity via two electrodes placed on the participant’s chest and internally records the ECG signal. Data were sampled at a rate of 1024 Hz. The IBI series was extracted from the ECG signal using QRSTool [114] with a threshold detection method. CardioEdit software (Brain-Body Center, University of Illinois at Chicago) was then used to edit artifacts and arrhythmias (<5%). Mean values for RSA and IBI were then extracted using CardioBatch software (Brain-Body Center, University of Illinois at Chicago). Briefly, CardioBatch samples sequential heart periods in 250 ms epochs and uses a 21-point moving polynomial algorithm to de-trend the data [115, 116]. The data are then bandpass filtered to extract variance associated with spontaneous breathing parameters (0.12–0.40 Hz), and RSA is estimated by transforming the variance to its natural logarithm. RSA and IBI were measured from 30 s epochs and then averaged for a total mean across the 3-min baseline period.
Depression symptom severity
Participants completed the Beck Depression Inventory-II [117], which is a 21-item questionnaire measuring self-reported symptoms of depression occurring over the last 2 weeks. Items are designed to reflect the defining symptoms of major depressive disorder as outlined in the Diagnostic and Statistical Manual for Mental Health Disorders [118] and are tallied to create a continuous index of depression symptom severity. The Beck depression inventory-II (BDI-II) demonstrates high test-retest reliability, internal consistency, and validity estimates (e.g., [119–121]). Nine women with the FMR1 premutation obtained a score of 14 or higher on the BDI-II, which is considered indicative of clinical depression; no control women scored within this range.
Anxiety symptom severity
The Beck Anxiety Inventory [122] measured self-reported generalized anxiety symptoms occurring over the past week. This 21-item questionnaire provides a total score reflecting anxiety symptom severity, aligning with the criteria outlined in the Diagnostic and Statistical Manual for Mental Health Disorders [118]. The Beck anxiety inventory (BAI) has high internal consistency, adequate test-retest reliability, and evidence supporting convergent and discriminant validity [123, 124]. Scores above 9 are considered indicative of clinically significant anxiety; 12 women with the FMR1 premutation and 3 control women scored above this cut-off.
FMR1 molecular measures
Genomic DNA was isolated from peripheral blood lymphocytes using standard methods (Qiagen, Valencia, CA). CGG repeat length was determined using polymerase chain reaction (PCR) and Southern Blot, as previously described [125, 126]. Activation ratio, or the percent of cells carrying the normal allele on the active X chromosome, was measured using an Alpha Innotech FluorChem 8800 Image Detection System [87]. Total RNA was isolated from 3 mL of blood collected in PAXgene® tubes. To determine the relative expression levels of the FMR1 gene, qRT-PCR amplification was carried out on total RNA using custom-designed Taqman gene expression assays, for both the validated target FMR1 gene and the reference genes (β-glucoronidase) in a 7900 Sequence detector (Applied Biosystems, Foster City, CA) as detailed in [87]. A quantitative index of FMRP was obtained by using a capture Luminex-based immunoassay to determine the amount of FMRP in peripheral blood lymphocytes (expressed in pg/ug of total lysate). This assay has been shown to have high accuracy with dried blood spots, peripheral lymphocytes, brain, and other human tissues [127, 128].
Data analysis
Analyses were conducted in SAS 9.4 [129]. The data were first examined for normality. Skewedness was detected for several variables; the Box-Cox transformation [130] was applied to find the optimal normalizing transformation for IBI (λ = −0.50), depression symptoms on the BDI-II (λ = 0), anxiety symptoms on the BAI (λ = −0.25), CGG repeat length (λ = 0), activation ratio (λ = 1.50), and mRNA (λ = −1.50); the data were transformed accordingly. The remaining variables were normally distributed and did not require transformation. Transformed values were used in all analyses. Descriptive statistics were computed and are presented in Tables 2 and 3. To explore potential confounds related to menopause status, t tests examined differences in the cardiac indices between the subgroups of pre- and postmenopausal women. Mean RSA and IBI did not differ by menopause status in the women with the FMR1 premutation (p’s >0.172). Information on menopause status was not available for the control participants.
Table 2.
Descriptive statistics
| Variable | Group | |
|---|---|---|
| FMR1 premutation | Control | |
| IBI (untransformed) M (SD), range |
816.20 (131.11), 540.27–1135.33 | 791.58 (137.04), 577.93–1193.69 |
| IBI (transformed) M (SD), range |
1.93 (0.01), 1.91–1.94 | 1.93 (0.01), 1.91–1.94 |
| RSA M (SD), range |
4.82 (1.44), 1.78–7.64 | 5.56 (0.97), 3.24–7.26 |
| BDI-II (untransformed) M (SD), range |
10.97 (7.84), 0–33.00 | 4.07 (3.66), 0–13.00 |
| BDI-II (transformed) M (SD), range |
2.56 (0.58), 1.39–3.61 | 1.99 (0.44), 1.38–2.83 |
| BAI (untransformed) M (SD), range |
7.96 (6.79), 0–24.00 | 3.72 (5.29), 0–23.00 |
| BAI (transformed) M (SD), range |
1.73 (0.35), 1.17–2.26 | 1.49 (0.29), 1.17–2.25 |
IBI inter-beat interval, RSA respiratory sinus arrhythmia, BDI-II Beck Depression Inventory, BAI Beck Anxiety Inventory
Table 3.
Descriptive statistics: FMR1 molecular measures in the FMR1 premutation group
| Variable | M (SD), range |
|---|---|
| CGG repeat length (untransformed) | 95.81 (17.42), 64–147 |
| CGG repeat length (transformed) | 4.54 (0.18), 4.16–4.99 |
| Quantitative FMRP | 9.16 (3.99), 2.81–18.44 |
| Activation ratio (untransformed) | 0.60 (0.18), 0.10–0.90 |
| Activation ratio (transformed) | −0.35 (0.13), −0.65 to −1.00 |
| Messenger RNA (untransformed) | 0.77 (0.19), 0.49–1.24 |
| Messenger RNA (transformed) | 0.68 (0.01), 0.68–0.70 |
To test the first research question, general linear regression models tested group as a predictor of IBI and RSA. Covariates in the models included age, medication use (captured as the total number of psychotropic medications used), and parenting stress level (indexed by the total stress percentile on the Parenting Stress Inventory-4 [112]); these variables have been shown to influence cardiac functioning in prior work [27, 131–133]. Cohen’s d effect sizes were computed for group differences [134]. In general, effect sizes of 0.32 or less are interpreted as “small,” 0.33–0.55 “medium,” and 0.56–1.20 “large” [135]. Then, a series of general linear models tested each of the cardiac variables, group, and their interaction as predictors of depression and anxiety symptoms, after controlling for age, medication use, and parenting stress level. False discovery was controlled by adjusting at the level of the model F test using the Benjamini-Hochberg correction procedure [136]. Interaction contrasts were estimated to determine the effect of the cardiac predictor on psychological symptoms at each level of group. Partial eta squared (η 2 p) effect sizes were computed. In general, values of η 2 p at 0.01, 0.06, and 0.14 are considered “small,” “medium,” and “large,” respectively [134].
Finally, exploratory Pearson correlations were conducted between the cardiac variables and the FMR1 molecular variables within the FMR1 premutation group. Significant correlations were followed with more sophisticated general linear models testing the molecular genetic variable as a predictor of the cardiac outcome, controlling for age, medication use, and parenting stress level. Because of the exploratory nature of this aim, we did not attempt to adjust for multiple comparisons in these analyses. Regression models including quadratic and cubic terms were also conducted to test for non-linear associations with CGG expansion size, considering recent reports of curvilinear associations with CGG repeat length (e.g., [75, 80]).
Results
Descriptive statistics
Means, standard deviations, and ranges for the cardiac indices and psychological symptoms are presented in Table 2. t tests indicated significant group differences for these variables, with the women with the FMR1 premutation presenting with higher levels of both depression symptoms (t [58.40] = 4.22, p < 0.001) and anxiety symptoms (t [53.83] = 2.74, p = 0.007). Table 3 presents the descriptive statistics of the FMR1 genetic data within the FMR1 premutation group.
Group comparisons on cardiac indicators
The combined effects of group, age, medication use, and parenting stress level accounted for significant variability in RSA, F (1, 51) = 2.84, p = 0.033, R 2 = 0.18. Group accounted for significant variability in RSA, with the women with the FMR1 premutation exhibiting lower RSA than controls, F (1, 51) = 4.17, p = 0.046. Cohen’s d effect size was 0.54, consistent with a medium effect. The combined effects group, age, medication use, and parenting stress level did not account for significant variability in IBI, F (1, 51) = 2.54, p = 0.051, and R 2 = 0.17. Cohen’s d for the group differences in IBI was 0.05, which is consistent with a small effect size. Regression coefficients are presented in Table 4, and group comparisons are presented in Fig. 1.
Table 4.
Regression coefficients depicting group membership as a predictor of cardiac autonomic indices
| Effect | B | SE | t | p | R 2 |
|---|---|---|---|---|---|
| Coefficients: RSA model | |||||
| Intercept | 5.87 | 0.88 | 6.70 | <0.001* | 0.18 |
| Groupa | −0.75 | 0.37 | −2.04 | 0.046* | |
| Age | −0.01 | 0.02 | −0.44 | 0.659 | |
| Medication use | −0.33 | 0.23 | −1.48 | 0.144 | |
| Parenting stress | <0.01 | 0.01 | 0.27 | 0.785 | |
| Coefficients: IBI model | |||||
| Intercept | 1.92 | <0.01 | 466.47 | <0.001* | 0.17 |
| Groupa | <0.01 | <0.01 | 0.16 | 0.876 | |
| Age | <0.01 | <0.01 | 2.92 | 0.005* | |
| Medication use | <0.01 | <0.01 | −1.11 | 0.272 | |
| Parenting stress | <0.01 | <0.01 | 0.47 | 0.641 | |
aThe control group was set as the reference category
*p < 0.05
Fig. 1.
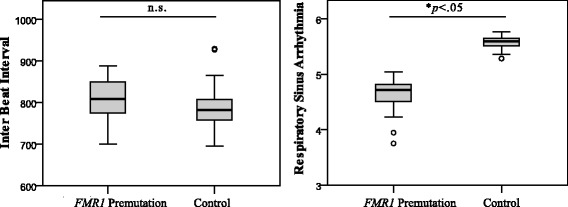
Group comparisons on respiratory sinus arrhythmia and inter-beat interval. Note: Figures present model-adjusted values, controlling for age, medication use, and parenting stress level. Untransformed IBI values are depicted for graphical representation. Boxes indicate data between the 25th and 75th percentile, with the horizontal bar reflecting the median (whiskers = the highest and lowest cases within the interquartile range; open circles = outliers, defined as cases falling greater than 1.5 times outside the interquartile range)
Relationship between cardiac activity and symptoms of anxiety and depression
A significant effect was detected for the overall model testing RSA as a predictor of depression symptoms (F [6, 48] = 9.82, p = 0.004, R 2 = 0.55). After controlling for age, medication use, and parenting stress level, the main effect for group was statistically significant, F [1, 48] = 21.39, p < 0.001, η 2 p = 0.31. A significant group-by-RSA interaction term was also detected (F [1, 48] = 7.83, p = 0.007, with a η 2 p effect size of 0.14 consistent with a large effect. Regression coefficients are presented in Table 5. Interaction contrasts confirmed that the effect of RSA on depression symptom severity differed by group; among the control women, decreased RSA was significantly associated with elevated depression symptoms with a medium-to-large effect (F [1, 48] = 6.40, p = 0.015, η 2 p = 0.12), whereas the association between RSA and depression symptoms was not statistically significant in the women with the FMR1 premutation with a small effect size (F [1, 48] = 1.83, p = 0.182, η 2 p = 0.04), see Fig. 2. The remaining models testing the cardiac variables as predictors of depression and anxiety symptoms did not indicate a significant effect of RSA, IBI, or their interactions with group on the psychological outcomes (see Tables 5 and 6).
Table 5.
Regression coefficients testing RSA as a predictor of depression and anxiety symptom severity
| Effect | β | SE | t | p | R 2 |
|---|---|---|---|---|---|
| Coefficients: Depression Symptom Severity Model | |||||
| Intercept | 3.64 | 0.60 | 6.10 | <0.001* | 0.55 |
| RSA | −0.24 | 0.09 | −2.53 | 0.015* | |
| Groupa | −1.34 | 0.62 | −2.16 | 0.036* | |
| Group x RSA | 0.32 | 0.11 | 2.85 | 0.007* | |
| Age | −0.12 | 0.01 | −2.27 | 0.028* | |
| Medication Use | 0.06 | 0.09 | 0.72 | 0.477 | |
| Parenting Stress | 0.01 | <0.01 | 3.85 | <0.001* | |
| Coefficients: Anxiety Symptom Severity Model | |||||
| Intercept | 2.04 | 0.42 | 4.87 | <0.001* | 0.41 |
| RSA | −0.05 | 0.07 | −0.78 | 0.437 | |
| Groupa | −0.23 | 0.43 | −0.52 | 0.604 | |
| Group × RSA | 0.07 | 0.08 | 0.88 | 0.384 | |
| Age | −0.01 | <0.01 | −2.21 | 0.032* | |
| Medication use | 0.04 | 0.06 | 0.58 | 0.564 | |
| Parenting stress | <0.01 | <0.01 | 3.06 | 0.004* | |
aThe control group was set as the reference category
*p < 0.05
Fig. 2.
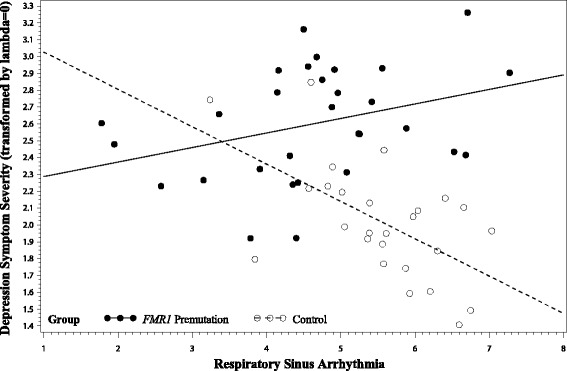
Differential associations between respiratory sinus arrhythmia and depression symptom severity across groups. Note: Model-adjusted values are depicted, controlling for age, medication use, and parenting stress level
Table 6.
Regression coefficients testing IBI as a predictor of depression and anxiety symptom severity
| Effect | β | SE | t | p | R 2 |
|---|---|---|---|---|---|
| Coefficients: Depression Symptom Severity Model | |||||
| Intercept | 17.76 | 35.52 | 0.50 | 0.619 | 0.48 |
| IBI | −7.96 | 18.49 | −0.42 | 0.669 | |
| Groupa | −31.06 | 43.19 | −0.72 | 0.476 | |
| Group × IBI | 16.30 | 22.39 | 0.73 | 0.470 | |
| Age | −0.02 | 0.01 | −2.11 | 0.040* | |
| Medication use | 0.04 | 0.09 | 0.40 | 0.691 | |
| Parenting stress | 0.01 | <0.01 | 3.46 | 0.001* | |
| Coefficients: Anxiety Symptom Severity Model | |||||
| Intercept | 15.92 | 22.38 | 0.71 | 0.481 | 0.43 |
| IBI | −7.33 | 11.65 | −0.63 | 0.532 | |
| Groupa | −34.14 | 27.09 | −1.26 | 0.214 | |
| Group × IBI | 17.77 | 14.04 | 1.27 | 0.212 | |
| Age | −0.01 | 0.01 | −2.13 | 0.039* | |
| Medication use | 0.04 | 0.06 | 0.61 | 0.542 | |
| Parenting stress | 0.01 | <0.01 | 3.10 | 0.003* | |
aThe control group was set as the reference category
*p < 0.05
Relationship between cardiac autonomic activity and FMR1 molecular variation
Exploratory Pearson correlations between the genetic and cardiac variables within the FMR1 premutation group are presented in Table 7. Elevated mRNA was correlated with higher RSA (r = 0.51, p = 0.009). CGG repeat length was also positively correlated with RSA (r = 0.57, p < 0.001). Significant correlations were followed with general linear models including age, medication use, and parenting stress level as covariates. After including for these covariates, mRNA remained a significant predictor of RSA, F (1, 18) = 4.88, p = 0.040, with a η 2 p of 0.21 which is consistent with a large effect, see Fig. 3. The general linear model testing CGG repeat length as a predictor of RSA did not show a significant effect of CGG repeat size after controlling for age, medication use, and parenting stress level; F (1, 22) = 2.51, p = 0.128, and η 2 p = 0.10. Regression coefficients are presented in Table 8. Finally, general linear regression models including quadratic and cubic terms were run to test for non-linear CGG effects, with no significant non-linear CGG effects detected.
Table 7.
Genetic correlations with the cardiac activity in women with the FMR1 premutation
| CGG repeat length | Quantitative FMRP | Messenger RNA | Activation ratio | |
|---|---|---|---|---|
| IBI | 0.14 | 0.26 | 0.17 | −0.01 |
| RSA | 0.57** | −0.17 | 0.51** | .0.07 |
IBI inter-beat interval, RSA respiratory sinus arrhythmia
**p < 0.01
Fig. 3.
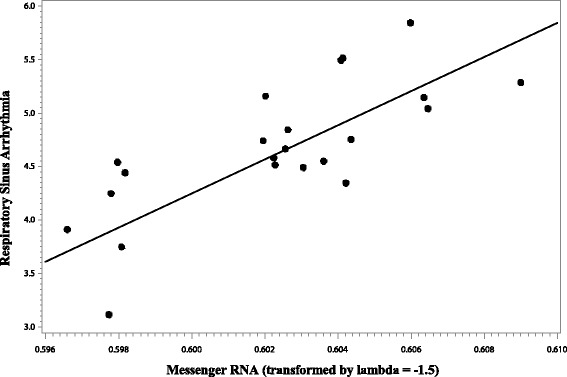
Association between FMR1 messenger RNA and respiratory sinus arrhythmia in women with the FMR1 premutation. Note: Model-adjusted values are presented, controlling for age, medication use, and parenting stress level
Table 8.
Regression coefficients testing FMR1 mRNA and CGG repeat length as predictors of RSA
| Effect | β | SE | t | p | R 2 |
|---|---|---|---|---|---|
| Coefficients: FMR1 mRNA predicting RSA | 0.29 | ||||
| Intercept | −112.07 | 53.06 | −2.11 | 0.049* | |
| mRNA | 191.70 | 86.79 | 2.21 | 0.040* | |
| Age | 0.04 | 0.06 | 0.78 | 0.444 | |
| Medication use | −0.01 | 0.01 | −0.69 | 0.497 | |
| Parenting stress | −0.35 | 0.30 | −1.17 | 0.256 | |
| Coefficients: CGG repeat length predicting RSA | |||||
| Intercept | −5.74 | 7.76 | −0.74 | 0.458 | 0.20 |
| CGG repeat | 2.52 | 1.59 | 1.58 | 0.128 | |
| Age | −0.02 | 0.04 | −0.42 | 0.679 | |
| Medication use | <0.01 | 0.01 | 0.14 | 0.891 | |
| Parenting stress | −0.59 | 0.30 | −1.98 | 0.060 | |
*p < 0.05
Discussion
Women with the FMR1 premutation are at substantially increased risk for depression and anxiety disorders, which are conditions associated with autonomic dysregulation in the general population. Given its single-gene basis, the FMR1 premutation may serve as a foothold to inform the genetic background for autonomic aberrations. This is the first study to examine cardiac autonomic function in women with the FMR1 premutation and its psychological and genetic correlates. Vagal tone was significantly depressed among the women with the FMR1 premutation, supporting impaired parasympathetic function in this group. Unlike the patterns observed in study controls and the general population, vagal tone and depression symptoms were unrelated in women with the FMR1 premutation, suggesting that the parasympathetic system is not serving its normal emotional regulatory functions in this group. Elevated FMR1 mRNA, which is typically associated with neuronal toxicity, was correlated with higher (i.e., “better”) vagal tone among women with the FMR1 premutation. Results underscore the need for additional research to delineate the clinical correlates and predictive utility of autonomic markers in this high-risk group and their relationship with FMR1-related mechanisms.
Group comparisons on cardiac indices of autonomic function
This study provides the first evidence of reduced vagal tone in women with the FMR1 premutation, which could not be accounted for by elevated parenting stress or increased use of psychotropic medications. Dampened vagal tone is thought to indicate inflexibility of psychophysiological resources that regulate affective information processing [137]. A large body of literature documents a supporting role of the vagus in emotional regulation and pro-social behavior. Adults with high vagal tone show greater self-regulatory capacity [138], better regulation of negative facial expressions [139, 140], increased perceived social support [141], and increased feelings of social integration and acceptance [142]. Vagal tone has also been shown to moderate the impact of negative life experiences, acting as a buffer to shield at risk individuals from negative emotional and physical consequences [143–145]. Moreover, an “upwards spiral” reciprocal causality effect has been suggested, where high vagal tone supports psychological well-being, which in turn promotes further vagal increases [146]. The finding of dampened vagal activity among women with the FMR1 premutation suggests that these individuals may lack the physiological resources that are needed to support optimal social-adaptive outcomes. Blunted vagal tone may be a factor in the elevated risk for emotional and physical health conditions seen in this group. Additional research is needed to determine the utility of vagal tone in predicting individual differences in clinical risk. Penetrance is not complete in the FMR1 premutation and the identification of a biomarker that can account for phenotypic variability would contribute significantly to prevention and treatment efforts.
Differential relationships between cardiac activity and psychological symptoms across groups
Vagal activity was not associated with depression symptoms in the women with the premutation, although, this relationship was observed in study controls and has been documented in the general population [14] and among individuals with clinically diagnosed mood disorders [10]. A similar decoupling between vagal tone and anxiety symptoms was observed in the FMR1 premutation, which is contrary to a wealth of evidence supporting a link between vagal regulation and anxiety symptoms in other groups [8]. Together, these findings suggest that parasympathetic control of the heart via the vagal nerve is not only suboptimal (i.e., reduced in level) but also dysfunctional (i.e., not serving its normal functions) in women with the FMR1 premutation. In other populations, the vagus is thought to play a mechanistic role in psychological vulnerability; when vagal tone is reduced, the body is unable to maintain an adaptive physiological state that promotes social engagement, leading to increased risk for emotional regulatory disorders [29]. Here, we found that vagal activity and psychological risk were not correlated in the FMR1 premutation, despite the fact that vagal tone was reduced and psychological symptoms were increased. This may suggest different mechanistic underpinnings in the FMR1 premutation and is consistent with distinct symptom profiles seen in this group (e.g., women with the premutation have a lower likelihood of recurrent major depressive episodes than women in the general population; Roberts et al., [75]). Future work incorporating measures of vagal reactivity may clarify relationships. The present study only included measures of tonic vagal activity, and some work suggests that task-related vagal modulation may be a more robust marker for depression than are baseline levels [39]. Future research may also investigate relationships among individuals who meet clinical thresholds for depression and anxiety, as opposed to investigating continuous symptom presentation across affected and unaffected individuals, as was done here, or among individuals with lifetime histories of depression and anxiety as opposed to current symptomatology.
Relationship between FMR1 molecular variation and cardiac activity
It is unexpected that elevated FMR1 mRNA was associated with higher (i.e., “better”) vagal levels within the FMR1 premutation group because mRNA is thought to be toxic to the neural system. So, why was elevated mRNA linked with superior vagal functioning in this sample? Undetected non-linear effects might explain this association, which would be consistent with evidence of CGG-dependent curvilinear risk patterns (i.e., [70, 75, 80, 81]) and the suggestion that maximal mRNA toxicity may occur within the mid-premutation range [80, 147]. However, an undetected curvilinear relationship seems unlikely, as the statistical tests and scatterplot distribution both suggest a linear association. The unexpected mRNA association underscores the complexity in untangling gene-brain-behavior relationships. FMR1 mRNA toxicity is thought to involve the sequestration of other RNA binding proteins, which prevents the proteins of other genes from carrying out their normal functions [69]. Thus, FMR1 does not function in isolation and the mechanisms by which FMR1 variation leads to autonomic dysfunction are not straightforward. It is possible that the relationship between mRNA and vagal tone is driven by background gene dysfunction caused by protein sequestration associated with elevated mRNA. More research is needed to understand the inter-correlations between FMR1 mRNA and other FMR1 and non-FMR1 mechanisms and their collective role in autonomic regulation.
Potential sex effects should also be considered, as much of our understanding of the functions of FMR1 mRNA comes from work involving males with the premutation (e.g., [103, 148, 149]). The functions of mRNA and its neurodegenerative consequences may differ across males and females, which is consistent with evidence that FXTAS is less prevalent among females and characterized by less white matter disease, reduced brain atrophy, fewer astrocytic inclusions, and lower likelihood for dementia when compared to males [150–152]. Sex differences may be partially accounted for by random X inactivation in females, but the influence of sex-specific hormonal patterns must also be considered and has not yet been characterized. Longitudinal work will also be an informative next step, particularly given that FMR1 mRNA gain-of-function is hypothesized to represent a degenerative, rather than developmental, mechanism, with toxicity building over time [153]. It should also be noted that mRNA levels were measured from peripheral blood lymphocytes and therefore might not necessarily reflect expression levels in relevant brain regions.
Findings did not support a relationship between cardiac activity and quantitatively measured FMRP levels in women with the premutation. No other studies have examined FMRP-autonomic relationships in the premutation, but these results are consistent with prior reports failing to detect a relationship between cardiac activity and the percent of lymphocytes staining positive for FMRP in males with the full mutation [154, 155]. One study did document a relationship between FMRP and vagal activity in females with the full mutation [155]; although, the significance of these findings are unclear as the association was only present when vagal tone was indexed using descriptive measures of heart rate variability but not when respiratory sinus arrhythmia was used, which is considered to be a more accurate measure of vagal tone [156, 157]. Overall, more research including larger samples is needed to determine whether FMRP is implicated in autonomic dysregulation in fragile X conditions.
Summary and directions
There are a number of future directions of this work. First, follow-up studies including more diverse samples and testing gender effects are needed. This study was limited by a relatively small sample, which may have reduced statistical power. Considering that the a priori power calculations for the larger study were based on by a different set of questions and assumptions, we reported effect sizes when possible to provide insight into the strength of the detected relationships. Given the novelty of the research question addressing the relationships between cardiac function and FMR1 molecular variation, this aim was considered exploratory and we did not attempt to correct for multiple comparisons. The exploratory associations detected here may be used to generate follow-up studies including more focused hypotheses. Follow-up work may also include more comprehensive investigation of associations with menopause, given that ~20% of women with the FMR1 premutation experience fragile X-associated primary ovarian insufficiency, and some research suggests changes in autonomic function following menopause (e.g., [110]). Finally, genotyping was not conducted on all controls and we cannot definitively rule out the presence of atypical CGG repeat numbers in this group, which could attenuate group differences.
It should also be noted that the premutation group consisted of mothers who had a child affected by fragile X syndrome, and results may not generalize to premutation carriers who do not have an affected child. While we covaried for parenting stress levels in our models, future work may more comprehensively examine the potential moderating role of parenting stress on the patterns observed here. Interactions with environmental factors such as social support should also be considered in future work, in light of evidence suggesting an “upwards spiral” reciprocal causality effect, where vagal tone and feelings of social connectedness reciprocally and prospectively predict one another [146]. Women with the FMR1 premutation report increased aloof personality traits [67] and heightened interpersonal sensitivity [95], which may interact with the vagal system. Vagal tone may also be important for understanding the family environment, as women with the premutation are particularly susceptible to parenting stress [158], and low vagal tone is thought to magnify sensitivity to psychosocial stressors [159]. Furthermore, vagal tone has been shown to moderate the parenting behaviors of shy-anxious mothers, influencing child outcomes [160]. Recent work shows that disruption of other allostatic systems, such as the neuroendocrine system, directly impacts maternal responsivity in mothers who carry the FMR1 premutation [161]. Adopting a biobehavioral approach may be invaluable in parsing out the complex, multi-dimensional influences on individual and family risk factors in this population.
It should also be acknowledged that the autonomic system is one of the many bodily stress regulatory systems, and a multisystem approach is needed to account for how interactions and coordination across systems may influence findings. For instance, hypothalamic-pituitary-adrenal (HPA) axis function of the neuroendocrine system is blunted in women with the FMR1 premutation and is related to FMR1 variation [81, 89]. Some evidence suggests that the vagus plays an inhibitory role in the regulation of other allostatic systems, including the neuroendocrine system, with individuals with low vagal tone showing poor post-stress recovery of cardiovascular, neuroendocrine, and immune markers [162]. Better understanding of how these interacting systems function together will be important for developing targeted treatments.
Conclusions
In summary, the present study provides evidence that autonomic dysfunction extends to the premutation, highlighting autonomic dysregulation as a hallmark feature associated with defects on FMR1. Associations between FMR1-related variation and cardiac activity were detected, which sheds light on genetic determinants of autonomic alterations relevant to FMR1-associated conditions and the general population as well. Despite the elevated depression and anxiety symptoms, we observed independence between psychological symptoms and the autonomic system dysfunction in women with the FMR1 premutation group. This suggests that cardiac indices may have limited utility as biomarkers for anxiety and depression in this group. Yet, there is little understanding of the clinical consequences of autonomic dysregulation in this group and future studies may identify cardiac indices as useful markers for other clinical phenotypes associated with FMR1 gene dysfunction, such as FXTAS. The identification of biomarkers for clinical risk in the FMR1 premutation may improve early identification, tailored treatment, prevention, and the ability to predict which individuals are most at risk for late-onset symptom presentation. This study represents a first step in that direction.
Acknowledgements
We would like to thank the women who participated in this study.
Funding
This research was supported by the National Institutes of Health (F32DC013934, PI: Klusek; R01MH090194, PI: Roberts; R01HD024356, PI: Abbeduto; R01HD02274, PI: Tassone), the Research Participant Registry Core of the Carolina Institute for Developmental Disabilities (P30HD03110), and the IDDRC Administrative Core (U54HD079125).
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Authors’ contributions
JK conceived the study and lead the data collection, analysis, and interpretation. JER provided guidance on study design, data collection, and interpretation. GL, TA, WTB, and FT contributed to the collection and interpretation of the genetic data. JK drafted the manuscript. All authors contributed to the interpretation of the results and critical revising of the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Informed consent was obtained from all participants. All procedures were approved by the Institutional Review Board of the University of South Carolina.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- BAI
Beck anxiety inventory
- BDI-II
Beck depression inventory-II
- FMR1
Fragile X mental retardation-1
- FMRP
Fragile X mental retardation protein
- FXTAS
Fragile X-associated tremor/ataxia syndrome
- IBI
Inter-beat interval
- mRNA
Messenger RNA
- RSA
Respiratory sinus arrhythmia
Contributor Information
Jessica Klusek, Phone: (803) 777-5049, Email: klusek@mailbox.sc.edu.
Giuseppe LaFauci, Email: giuseppe.x.lafauci@opwdd.ny.gov.
Tatyana Adayev, Email: tatyana.adayev@opwdd.ny.gov.
W. Ted Brown, Email: ted.brown@opwdd.ny.gov.
Flora Tassone, Email: ftassone@ucdavis.edu.
Jane E. Roberts, Email: jane.roberts@sc.edu
References
- 1.McEwen BS. Stress, adaptation, and disease: allostasis and allostatic load. Ann N Y Acad Sci. 1998;840:33–44. doi: 10.1111/j.1749-6632.1998.tb09546.x. [DOI] [PubMed] [Google Scholar]
- 2.Lovallo WR. Stress and health: Biological and psychological interactions. 3. Tousand Oaks: Sage Publications; 2015. [Google Scholar]
- 3.Jänig W, McLachlan EM. Autonomic Failure: A Textbook of Clinical Disorders of the Autonomic Nervous System Oxford. 2013. Neurobiology of the autonomic nervous system; pp. 21–34. [Google Scholar]
- 4.Shaffer F, McCraty R, Zerr CL. A healthy heart is not a metronome: an integrative review of the heart’s anatomy and heart rate variability. Front Psychol. 2014;5:1040. [DOI] [PMC free article] [PubMed]
- 5.Grossman P, Taylor EW. Toward understanding respiratory sinus arrhythmia: relations to cardiac vagal tone, evolution and biobehavioral functions. Biol Psychol. 2007;74:263–85. doi: 10.1016/j.biopsycho.2005.11.014. [DOI] [PubMed] [Google Scholar]
- 6.Appelhans BM, Luecken LJ. Heart rate variability as an index of regulated emotional responding. Rev Gen Psychol. 2006;10:229.
- 7.Nardelli M, Valenza G, Cristea IA, Gentili C, Cotet C, David D, Lanata A, Scilingo EP. Characterizing psychological dimensions in non-pathological subjects through autonomic nervous system dynamics. Front Comput Neurosci. 2015;9:37. doi: 10.3389/fncom.2015.00037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Friedman BH. An autonomic flexibility-neurovisceral integration model of anxiety and cardiac vagal tone. Biol Psychol. 2007;74:185–99. doi: 10.1016/j.biopsycho.2005.08.009. [DOI] [PubMed] [Google Scholar]
- 9.Gorman JM, Sloan RP. Heart rate variability in depressive and anxiety disorders. Am Heart J. 2000;140:S77–83. doi: 10.1067/mhj.2000.109981. [DOI] [PubMed] [Google Scholar]
- 10.Carney RM, Blumenthal JA, Stein PK, Watkins L, Catellier D, Berkman LF, Czajkowski SM, O’Connor C, Stone PH, Freedland KE. Depression, heart rate variability, and acute myocardial infarction. Circulation. 2001;104:2024–8. doi: 10.1161/hc4201.097834. [DOI] [PubMed] [Google Scholar]
- 11.Kemp AH, Quintana DS, Felmingham KL, Matthews S, Jelinek HF. Depression, comorbid anxiety disorders, and heart rate variability in physically healthy, unmedicated patients: implications for cardiovascular risk. PLoS One. 2012;7:e30777. doi: 10.1371/journal.pone.0030777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stapelberg NJ, Hamilton-Craig I, Neumann DL, Shum DHK, McConnell H. Mind and heart: heart rate variability in major depressive disorder and coronary heart disease—a review and recommendations. Aust N Z J Psychiatry. 2012;46:946–57. doi: 10.1177/0004867412444624. [DOI] [PubMed] [Google Scholar]
- 13.Koenig J, Kemp AH, Beauchaine TP, Thayer JF, Kaess M. Depression and resting state heart rate variability in children and adolescents—a systematic review and meta-analysis. Clin Psychol Rev. 2016;46:136–50. doi: 10.1016/j.cpr.2016.04.013. [DOI] [PubMed] [Google Scholar]
- 14.Hughes JW, Stoney CM. Depressed mood is related to high-frequency heart rate variability during stressors. Psychosom Med. 2000;62:796–803. doi: 10.1097/00006842-200011000-00009. [DOI] [PubMed] [Google Scholar]
- 15.Vazquez L, Blood JD, Wu J, Chaplin TM, Hommer RE, Rutherford HJV, Potenza MN, Mayes LC, Crowley MJ. High frequency heart-rate variability predicts adolescent depressive symptoms, particularly anhedonia, across one year. J Affect Disord. 2016;196:243–7. doi: 10.1016/j.jad.2016.02.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Koval P, Ogrinz B, Kuppens P, Van den Bergh O, Tuerlinckx F, Sütterlin S. Affective instability in daily life is predicted by resting heart rate variability. PLoS One. 2013;8:e81536. doi: 10.1371/journal.pone.0081536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Agelink MW, Boz C, Ullrich H, Andrich J. Relationship between major depression and heart rate variability: clinical consequences and implications for antidepressive treatment. Psychiatry Res. 2002;113:139–49. doi: 10.1016/S0165-1781(02)00225-1. [DOI] [PubMed] [Google Scholar]
- 18.Agelink MW, Klimke A, Cordes J, Sanner D, Kavuk I, Malessa R, Klieser E, Baumann B. A functional-structural model to understand cardiac autonomic nervous system (ANS) dysregulation in affective illness and to elucidate the ANS effects of antidepressive treatment. Eur J Med Res. 2004;9:37–50. [PubMed] [Google Scholar]
- 19.Chambers AS, Allen JJ. Vagal tone as an indicator of treatment response in major depression. Psychophysiology. 2002;39:861–4. doi: 10.1111/1469-8986.3960861. [DOI] [PubMed] [Google Scholar]
- 20.Balogh S, Fitzpatrick DF, Hendricks SE, Paige SR. Increases in heart rate variability with successful treatment in patients with major depressive disorder. Psychopharmacol Bull. 1993;29:201–6. [PubMed] [Google Scholar]
- 21.Carney RM, Freedland KE, Stein PK, Skala JA, Hoffman P, Jaffe AS. Change in heart rate and heart rate variability during treatment for depression in patients with coronary heart disease. Psychosom Med. 2000;62:639–47. doi: 10.1097/00006842-200009000-00007. [DOI] [PubMed] [Google Scholar]
- 22.de Guevara MS, Schauffele SI, Nicola-Siri LC, Fahrer RD, Ortiz-Fragola E, Martinez-Martinez JA, Cardinali DP, Guinjoan SM. Worsening of depressive symptoms 6 months after an acute coronary event in older adults is associated with impairment of cardiac autonomic function. J Affect Disord. 2004;80:257–62. doi: 10.1016/S0165-0327(03)00105-8. [DOI] [PubMed] [Google Scholar]
- 23.Royster EB, Trimble LM, Cotsonis G, Schmotzer B, Manatunga A, Rushing NN, Pagnoni G, Auyeung SF, Brown AR, Schoenbeck J, et al. Changes in heart rate variability of depressed patients after electroconvulsive therapy. Cardiovasc Psychiatry Neurol. 2012;2012:794043. doi: 10.1155/2012/794043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Karpyak VM, Rasmussen KG, Hammill SC, Mrazek DA. Changes in heart rate variability in response to treatment with electroconvulsive therapy. J ECT. 2004;20:81–8. doi: 10.1097/00124509-200406000-00002. [DOI] [PubMed] [Google Scholar]
- 25.Miu AC, Heilman RM, Miclea M. Reduced heart rate variability and vagal tone in anxiety: trait versus state, and the effects of autogenic training. Auton Neurosci. 2009;145:99–103. doi: 10.1016/j.autneu.2008.11.010. [DOI] [PubMed] [Google Scholar]
- 26.Watkins LL, Grossman P, Krishnan R, Sherwood A. Anxiety and vagal control of heart rate. Psychosom Med. 1998;60:498–502. doi: 10.1097/00006842-199807000-00018. [DOI] [PubMed] [Google Scholar]
- 27.Brosschot JF, Van Dijk E, Thayer JF. Daily worry is related to low heart rate variability during waking and the subsequent nocturnal sleep period. Int J Psychophysiol. 2007;63:39–47. doi: 10.1016/j.ijpsycho.2006.07.016. [DOI] [PubMed] [Google Scholar]
- 28.Porges SW. The polyvagal theory: phylogenetic contributions to social behavior. Physiol Behav. 2003;79:503–13. doi: 10.1016/S0031-9384(03)00156-2. [DOI] [PubMed] [Google Scholar]
- 29.Porges SW. The polyvagal perspective. Biol Psychol. 2007;74:116–43. doi: 10.1016/j.biopsycho.2006.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Thayer JF, Lane RD. A model of neurovisceral integration in emotion regulation and dysregulation. J Affect Disord. 2000;61:201–16. doi: 10.1016/S0165-0327(00)00338-4. [DOI] [PubMed] [Google Scholar]
- 31.Thayer JF, Lane RD. Claude Bernard and the heart–brain connection: further elaboration of a model of neurovisceral integration. Neurosci Biobehav Rev. 2009;33:81–8. doi: 10.1016/j.neubiorev.2008.08.004. [DOI] [PubMed] [Google Scholar]
- 32.Campbell-Sills L, Barlow DH. Incorporating emotion regulation into conceptualizations and treatments of anxiety and mood disorders. In: Gross JJ, editor. Handbook of Emotion Regulation. New York: Guilford Press; 2007. pp. 542–59. [Google Scholar]
- 33.Thayer JF, Hansen AL, Saus-Rose E, Johnsen BH. Heart rate variability, prefrontal neural function, and cognitive performance: the neurovisceral integration perspective on self-regulation, adaptation, and health. Ann Behav Med. 2009;37:141–53. doi: 10.1007/s12160-009-9101-z. [DOI] [PubMed] [Google Scholar]
- 34.Porges SW, Furman SA. The early development of the autonomic nervous system provides a neural platform for social behavior: a polyvagal perspective. Infant Child Dev. 2011;20:106–18. doi: 10.1002/icd.688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.McEwen BS, Wingfield JC. The concept of allostasis in biology and biomedicine. Horm Behav. 2003;43:2–15. doi: 10.1016/S0018-506X(02)00024-7. [DOI] [PubMed] [Google Scholar]
- 36.Thayer JF, Brosschot JF. Psychosomatics and psychopathology: looking up and down from the brain. Psychoneuroendocrinology. 2005;30:1050–8. doi: 10.1016/j.psyneuen.2005.04.014. [DOI] [PubMed] [Google Scholar]
- 37.Beauchaine TP. The role of biomarkers and endophenotypes in prevention and treatment of psychopathological disorders. Biomark Med. 2009;3:1–3. doi: 10.2217/17520363.3.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Beauchaine TP, Neuhaus E, Brenner SL, Gatzke-Kopp L. Ten good reasons to consider biological processes in prevention and intervention research. Dev Psychopathol. 2008;20:745. doi: 10.1017/S0954579408000369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yaroslavsky I, Rottenberg J, Kovacs M. Atypical patterns of respiratory sinus arrhythmia index an endophenotype for depression. Dev Psychopathol. 2014;26:1337–52. doi: 10.1017/S0954579414001060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Beauchaine TP. Respiratory sinus arrhythmia: a transdiagnostic biomarker of emotion dysregulation and psychopathology. Curr Opin Psychol. 2015;3:43–7. doi: 10.1016/j.copsyc.2015.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Boomsma DI, Plomin R. Heart rate and behavior of twins. Merrill-Palmer Q (1986) 1986;32:141–51. [Google Scholar]
- 42.Doussard-Roosevelt JA, Montgomery LA, Porges SW. Short-term stability of physiological measures in kindergarten children: respiratory sinus arrhythmia, heart period, and cortisol. Dev Psychobiol. 2003;43:230–42. doi: 10.1002/dev.10136. [DOI] [PubMed] [Google Scholar]
- 43.Fracasso MP, Porges SW, Lamb ME, Rosenberg AA. Cardiac activity in infancy: reliability and stability of individual differences. Infant Behav Dev. 1994;17:277–84. doi: 10.1016/0163-6383(94)90006-X. [DOI] [Google Scholar]
- 44.Kupper NH, Willemsen G, van den Berg M, de Boer D, Posthuma D, Boomsma DI, de Geus EJ. Heritability of ambulatory heart rate variability. Circulation. 2004;110:2792–6. doi: 10.1161/01.CIR.0000146334.96820.6E. [DOI] [PubMed] [Google Scholar]
- 45.Maddalena A, Richards CS, McGinniss MJ, Brothman A, Desnick RJ, Grier RE, Hirsch B, Jacky P, McDowell GA, Popovich B. Technical standards and guidelines for fragile X: the first of a series of disease-specific supplements to the Standards and Guidelines for Clinical Genetics Laboratories of the American College of Medical Genetics. Genet Med. 2001;3:200–5. doi: 10.1097/00125817-200105000-00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hagerman PJ, Hagerman RJ. Fragile X-associated tremor/ataxia syndrome. Ann N Y Acad Sci. 2015;1338:58–70. doi: 10.1111/nyas.12693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Oh SY, He F, Krans A, Frazer M, Taylor JP, Paulson HL, Todd PK. RAN translation at CGG repeats induces ubiquitin proteasome system impairment in models of fragile X-associated tremor ataxia syndrome. Hum Mol Genet. 2015;24:4317–26. doi: 10.1093/hmg/ddv165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Todd PK, Oh SY, Krans A, He F, Sellier C, Frazer M, Renoux AJ, Chen K-c, Scaglione KM, Basrur V. CGG repeat-associated translation mediates neurodegeneration in fragile X tremor ataxia syndrome. Neuron. 2013;78:440–55. doi: 10.1016/j.neuron.2013.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fernandez-Carvajal I, Walichiewicz P, Xiaosen X, Pan R, Hagerman PJ, Tassone F. Screening for expanded alleles of the FMR1 gene in blood spots from newborn males in a Spanish population. J Mol Diagn. 2009;11:324–9. doi: 10.2353/jmoldx.2009.080173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hantash FM, Goos DM, Crossley B, Anderson B, Zhang K, Sun W, Strom CM. FMR1 premutation carrier frequency in patients undergoing routine population-based carrier screening: Insights into the prevalence of fragile X syndrome, fragile X-associated tremor/ataxia syndrome, and fragile X-associated primary ovarian insufficiency in the United States. Genet Med. 2011;13:39–45. doi: 10.1097/GIM.0b013e3181fa9fad. [DOI] [PubMed] [Google Scholar]
- 51.Seltzer MM, Baker MW, Hong J, Maenner M, Greenberg J, Mandel D. Prevalence of CGG expansions of the FMR1 gene in a US population-based sample. Am J Med Genet B Neuropsychiatr Genet. 2012;159B:589–97. doi: 10.1002/ajmg.b.32065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Tassone F, Iong KP, Tong T-H, Lo J, Gane LW, Berry-Kravis E, Nguyen D, Mu LY, Laffin J, Bailey DB. FMR1 CGG allele size and prevalence ascertained through newborn screening in the United States. Genome Med. 2012;4:100. doi: 10.1186/gm401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Toledano-Alhadef H, Basel-Vanagaite L, Magal N, Davidov B, Ehrlich S, Drasinover V, Taub E, Halpern GJ, Ginott N, Shohat M. Fragile-X carrier screening and the prevalence of the premutation and full-mutation carriers in Israel. Am J Hum Genet. 2001;69:351–60. doi: 10.1086/321974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Yrigollen CM, Martorell L, Durbin-Johnson B, Naudo M, Genoves J, Murgia A, Polli R, Zhou L, Barbouth D, Rupchock A, et al. AGG interruptions and maternal age affect FMR1 CGG repeat allele stability during transmission. J Neurodev Disord. 2014;6:1–12. doi: 10.1186/1866-1955-6-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nolin SL, Glicksman A, Ersalesi N, Dobkin C, Brown WT, Cao R, Blatt E, Sah S, Latham GJ, Hadd AG. Fragile X full mutation expansions are inhibited by one or more AGG interruptions in premutation carriers. Genet Med. 2014;17:358–64. doi: 10.1038/gim.2014.106. [DOI] [PubMed] [Google Scholar]
- 56.Coffee B, Keith K, Albizua I, Malone T, Mowrey J, Sherman SL, Warren ST. Incidence of fragile X syndrome by newborn screening for methylated FMR1 DNA. Am J Hum Genet. 2009;85:503–14. doi: 10.1016/j.ajhg.2009.09.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Hagerman RJ, Hagerman PJ. Fragile X syndrome: Diagnosis, treatment, and research. Taylor & Francis US; 2002.
- 58.Klusek J, McGrath SE, Abbeduto L, Roberts JE. Pragmatic language features of mothers with the FMR1 premutation are associated with the language outcomes of adolescents and young adults with fragile X syndrome. J Speech Lang Hear Res. 2016;59:49–61. doi: 10.1044/2015_JSLHR-L-15-0102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wheeler A, Hatton D, Reichardt A, Bailey D. Correlates of maternal behaviours in mothers of children with fragile X syndrome. J Intellect Disabil Res. 2007;51:447–62. doi: 10.1111/j.1365-2788.2006.00896.x. [DOI] [PubMed] [Google Scholar]
- 60.Tassone F, Hagerman PJ, Hagerman RJ. Fragile X premutation. J Neurodev Disord. 2014;6:1–4. doi: 10.1186/1866-1955-6-22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Allingham-Hawkins DJ, Babul-Hirji R, Chitayat D, Holden JJA, Yang KT, Lee C, Hudson R, Gorwill H, Nolin SL, Glicksman A, et al. Fragile X premutation is a significant risk factor for premature ovarian failure: the international collaborative POF in fragile X study—preliminary data. Am J Med Genet. 1999;83:322–5. doi: 10.1002/(SICI)1096-8628(19990402)83:4<322::AID-AJMG17>3.0.CO;2-B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rodriguez-Revenga L, Madrigal I, Pagonabarraga J, Xuncla M, Badenas C, Kulisevsky J, Gomez B, Mila M. Penetrance of FMR1 premutation associated pathologies in fragile X syndrome families. Eur J Hum Genet. 2009;17:1359–62. doi: 10.1038/ejhg.2009.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Shelton AL, Cornish K, Kraan C, Georgiou-Karistianis N, Metcalfe SA, Bradshaw JL, Hocking DR, Archibald AD, Cohen J, Trollor JN. Exploring inhibitory deficits in female premutation carriers of fragile X syndrome: through eye movements. Brain Cogn. 2014;85:201–8. doi: 10.1016/j.bandc.2013.12.006. [DOI] [PubMed] [Google Scholar]
- 64.Kraan CM, Hocking DR, Georgiou‐Karistianis N, Metcalfe SA, Archibald AD, Fielding J, Trollor J, Bradshaw JL, Cohen J, Cornish KM. Impaired response inhibition is associated with self‐reported symptoms of depression, anxiety, and ADHD in female FMR1 premutation carriers. Am J Med Genet B Neuropsychiatr Genet. 2014;165:41–51. doi: 10.1002/ajmg.b.32203. [DOI] [PubMed] [Google Scholar]
- 65.Goodrich-Hunsaker NJ, Wong LM, McLennan Y, Srivastava S, Tassone F, Harvey D, Rivera SM, Simon TJ. Young adult female fragile X premutation carriers show age- and genetically-modulated cognitiveimpairments. Brain Cogn. 2011;75:255-60. [DOI] [PMC free article] [PubMed]
- 66.Goodrich-Hunsaker NJ, Wong LM, McLennan Y, Tassone F, Harvey D, Rivera SM, Simon TJ. Adult female fragile X premutation carriers exhibit age- and CGG repeat length-related impairments on an attentionally-based enumeration task. Front Hum Neurosci. 2011;5:63. [DOI] [PMC free article] [PubMed]
- 67.Losh M, Klusek J, Martin GE, Sideris J, Parlier M, Piven J. Defining genetically meaninful language and personality traits in relatives of individuals with fragile X syndrome and relatives of individuals with autism. Am J Med Genet B Neuropsychiatr Genet. 2012;159B:660–8. doi: 10.1002/ajmg.b.32070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Clifford S, Dissanayake C, Bui QM, Huggins R, Taylor AK, Loesch DZ. Autism spectrum phenotype in males and females with fragile X full mutation and premutation. J Autism Dev Disord. 2007;37:738–47. doi: 10.1007/s10803-006-0205-z. [DOI] [PubMed] [Google Scholar]
- 69.Hagerman R, Hagerman P. Advances in clinical and molecular understanding of the FMR1 premutation and fragile X-associated tremor/ataxia syndrome. Lancet Neurol. 2013;12:786–98. doi: 10.1016/S1474-4422(13)70125-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Roberts JE, Tonnsen BL, McCary LM, Ford AL, Golden RN, Bailey DB. Trajectory and Predictors of Depression and Anxiety Disorders in Mothers with the FMR1 Premutation. Biol Psychiatry. 2016;79:85. [DOI] [PMC free article] [PubMed]
- 71.Thompson NM, Rogeness GA, McClure E, Clayton R, Johnson C. Influence of depression on cognitive functioning in fragile X females. Psychiatric Res. 1996;64:97–104. doi: 10.1016/0165-1781(96)02785-0. [DOI] [PubMed] [Google Scholar]
- 72.Franke P, Leboyer M, Gansicke M, Weiffenbacj O. Genotype-phenotype relationship in female carriers of the premutation and full mutation of FMR-1. Psychiatry Res. 1998;90:113–27. doi: 10.1016/S0165-1781(98)00055-9. [DOI] [PubMed] [Google Scholar]
- 73.Reiss A, Freund L, Abrams MT, Boehm C, Kazazian H. Neurobehavioral effects of the fragile X premutation in adult women: a controlled study. Am J Hum Genet. 1993;52:884–94. [PMC free article] [PubMed] [Google Scholar]
- 74.Bourgeois JA, Seritan AL, Casillas EM, Hessl D, Schneider A, Yang Y, Kaur I, Cogswell JB, Nguyen DV, Hagerman RJ. Lifetime prevalence of mood and anxiety disorders in fragile X premutation carriers. J Clin Psychiatry. 2011;72:175–82. doi: 10.4088/JCP.09m05407blu. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Roberts JE, Bailey DB, Mankowski J, Ford A, Weisenfeld LA, Heath TM, Golden RN. Mood and anxiety disorders in females with the FMR1 premutation. Am J Med Genet B Neuropsychiatr Genet. 2009;150B:130–9. doi: 10.1002/ajmg.b.30786. [DOI] [PubMed] [Google Scholar]
- 76.Hunter JE, Rohr JK, Sherman SL. Co-occurring diagnoses among FMR1 premutation allele carriers. Clin Genet. 2010;77:374–81. doi: 10.1111/j.1399-0004.2009.01317.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cordeiro L, Abucayan F, Hagerman R, Tassone F, Hessl D. Anxiety disorders in fragile X premutation carriers: preliminary characterization of probands and non-probands. Intractable Rare Dis Res. 2015;4:123–30. doi: 10.5582/irdr.2015.01029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Roberts JE, Tonnsen BL, McCary LM, Ford AL, Golden RN, Bailey DB. Trajectory and predictors of depression and anxiety disorders in mothers with the FMR1 premutation. Biol Psychiatry. 2016;79:850–7. doi: 10.1016/j.biopsych.2015.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Franke P, Maier W, Hautzinger M, Weiffenbach O, Gänsicke M, Iwers B, Poustka F, Schwab SG, Froster U. Fragile-X carrier females: evidence for a distinct psychopathological phenotype? Am J Med Genet A. 1996;64:334–9. doi: 10.1002/(SICI)1096-8628(19960809)64:2<334::AID-AJMG20>3.0.CO;2-F. [DOI] [PubMed] [Google Scholar]
- 80.Loesch D, Bui M, Hammersley E, Schneider A, Storey E, Stimpson P, Burgess T, Francis D, Slater H, Tassone F. Psychological status in female carriers of premutation FMR1 allele showing a complex relationship with the size of CGG expansion. Clin Genet. 2015;87:173–8. doi: 10.1111/cge.12347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Seltzer MM, Barker ET, Greenberg JS, Hong J, Coe C, Almeida D. Differential sensitivity to life stress in FMR1 premutation carrier mothers of children with fragile X syndrome. Health Psychol. 2012;31:612–22. doi: 10.1037/a0026528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Allen EG, He W, Yadav-Shah M, Sherman SL. A study of the distributional characteristics of FMR1 transcript levels in 238 individuals. Hum Genet. 2004;114:439–47. doi: 10.1007/s00439-004-1086-x. [DOI] [PubMed] [Google Scholar]
- 83.Tassone F, Beilina A, Carosi C, Albertosi S, Bagni C, Li L, Glover K, Bentley D, Hagerman PJ. Elevated FMR1 mRNA in premutation carriers is due to increased transcription. RNA. 2007;13:555–62. doi: 10.1261/rna.280807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Hessl D, Tassone F, Loesch DZ, Berry-Kravis E, Leehey MA, Gane LW, Barbato I, Rice C, Gould E, Hall DA, et al. Abnormal elevation of FMR1 mRNA is associated with psychological symptoms in individuals with the fragile X premutation. Am J Med Genet B Neuropsychiatr Genet. 2005;139:115–21. doi: 10.1002/ajmg.b.30241. [DOI] [PubMed] [Google Scholar]
- 85.Hessl D, Wang JM, Schneider A, Koldewyn K, Le L, Iwahashi C, Cheung K, Tassone F, Hagerman PJ, Rivera SM. Decreased FMRP expression underlies amygdala dysfunction in carriers of the fragile X premutation. Biol Psychiatry. 2011;70:859–65. doi: 10.1016/j.biopsych.2011.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Seritan AL, Bourgeois JA, Schneider A, Mu Y, Hagerman RJ, Nguyen DV. Ages of onset of mood and anxiety disorders in fragile X premutation carriers. Curr Psychiatr Rev. 2013;9:65–71. doi: 10.2174/157340013805289662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Tassone F, Hagerman RJ, Taylor AK, Gane LW, Godfrey TE, Hagerman PJ. Elevated levels of FMR1 messenger RNA in carrier males: a new mechanism of involvement in the fragile X syndrome. Am J Hum Genet. 2000;66:6–15. doi: 10.1086/302720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Tonnsen BL, Cornish KM, Wheeler AC, Roberts JE. Maternal predictors of anxiety risk in young males with fragile X. Am J Med Genet B: Neuropsychiatr Genet. 2014;165B:299-409. [DOI] [PMC free article] [PubMed]
- 89.Hartley SL, Seltzer MM, Hong J, Greenberg JS, Smith L, Almeida D, Coe C, Abbeduto L. Cortisol response to behavior problems in FMR1 premutation mothers of adolescents and adults with fragile X syndrome: a diathesis-stress model. Int J Behav Dev. 2012;36:53–61. doi: 10.1177/0165025411406857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kenneson A, Zhang F, Hagedorn CH, Warren ST. Reduced FMRP and increased FMR1 transcription is proportionally associated with CGG repeat number in intermediate-length and premutation carriers. Hum Mol Genet. 2001;10:1449–54. doi: 10.1093/hmg/10.14.1449. [DOI] [PubMed] [Google Scholar]
- 91.Primerano B, Tassone F, Hagerman RJ, Hagerman P, Amaldi F, Bagni C. Reduced FMR1 mRNA translation efficiency in fragile X patients with premutations. RNA. 2002;8:1482–8. [PMC free article] [PubMed] [Google Scholar]
- 92.Darnell JC, Van Driesche SJ, Zhang C, Hung KY, Mele A, Fraser CE, Stone EF, Chen C, Fak JJ, Chi SW, et al. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism. Cell. 2011;146:247–61. doi: 10.1016/j.cell.2011.06.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Penagarikano O, Mulle JG, Warren ST. The pathophysiology of fragile x syndrome. Annu Rev Genomics Hum Genet. 2007;8:109–29. doi: 10.1146/annurev.genom.8.080706.092249. [DOI] [PubMed] [Google Scholar]
- 94.Schutzius G, Bleckmann D, Kapps-Fouthier S, di Giorgio F, Gerhartz B, Weiss A. A quantitative homogeneous assay for fragile X mental retardation 1 protein. J Neurodev Disord. 2013;5:8. doi: 10.1186/1866-1955-5-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Johnston C, Eliez S, Dyer-Friedman J, Hessl D, Glaser B, Blasey C, Taylor A, Reiss A. Neurobehavioral phenotype in carriers of the fragile X premutation. Am J Med Genet. 2001;103:314–9. doi: 10.1002/ajmg.1561. [DOI] [PubMed] [Google Scholar]
- 96.Hunter JE, Allen EG, Abramowitz A, Rusin M, Leslie M, Novak G, Hamilton D, Shubeck L, Charen K, Sherman SL. Investigation of phenotypes associated with mood and anxiety among male and female fragile X premutation carriers. Behav Genet. 2008;38:493–502. doi: 10.1007/s10519-008-9214-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Hunter JE, Leslie M, Novak G, Hamilton D, Shubeck L, Charen K, Abramowitz A, Epstein MP, Lori A, Binder E, et al. Depression and anxiety symptoms among women who carry the FMR1 premutation: impact of raising a child with fragile X syndrome is moderated by CRHR1 polymorphisms. Am J Med Genet B Neuropsychiatr Genet. 2012;0:549–59. doi: 10.1002/ajmg.b.32061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Cornish KM, Kraan CM, Bui QM, Bellgrove MA, Metcalfe SA, Trollor JN, Hocking DR, Slater HR, Inaba Y, Li X. Novel methylation markers of the dysexecutive-psychiatric phenotype in FMR1 premutation women. Neurology. 2015;84:1631–8. doi: 10.1212/WNL.0000000000001496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Adams PE, Adams JS, Nguyen DV, Hessl S, Brunberg JA, Tassone S, et al. Psychological symptoms correlate with reduced hippocampal volume in fragile X premutation carriers. Am J Med Genet. 2010;153B:775–85. doi: 10.1002/ajmg.b.31046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Klusek J, Roberts JE, Losh M. Cardiac autonomic regulation in autism and fragile X syndrome: a review. Psychol Bull. 2015;141:141–75. doi: 10.1037/a0038237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Coffey SM, Cook K, Tartaglia N, Tassone F, Nguyen DV, Pan R, Bronsky HE, Yuhas J, Borodyanskaya M, Grigsby J, et al. Expanded clinical phenotype of women with the FMR1 premutation. Am J Med Genet A. 2008;146:1009–16. doi: 10.1002/ajmg.a.32060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Jacquemont S, Hagerman RJ, Leehey M, Grigsby J, Zhang L, Brunberg JA, Greco C, Des Portes V, Jardini T, Levine R, et al. Fragile X premutation tremor/ataxia syndrome: molecular, clinical, and neuroimaging correlates. Am J Hum Genet. 2003;72:869–78. doi: 10.1086/374321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Greco CM, Berman RF, Martin RM, Tassone F, Schwartz PH, Chang A, Trapp BD, Iwahashi C, Brunberg J, Grigsby J, et al. Neuropathology of fragile X-associated tremor/ataxia syndrome (FXTAS) Brain. 2005;129:243–55. doi: 10.1093/brain/awh683. [DOI] [PubMed] [Google Scholar]
- 104.Gokden M, Al-Hinti JT, Harik SI. Peripheral nervous system pathology in fragile X tremor/ataxia syndrome (FXTAS) Neuropathology. 2009;29:280–4. doi: 10.1111/j.1440-1789.2008.00948.x. [DOI] [PubMed] [Google Scholar]
- 105.Hessl D, Rivera S, Koldewyn K, Cordeiro L, Adams J, Tassone F, Hagerman PJ, Hagerman RJ. Amygdala dysfunction in men with the fragile X premutation. Brain. 2007;130:404–16. doi: 10.1093/brain/awl338. [DOI] [PubMed] [Google Scholar]
- 106.Kaufman AS, Kaufman NL. Kaufman Brief Intelligence Test. 2. Los Angeles: Pearson Assessments; 2004. [Google Scholar]
- 107.DiPietro JA, Costigan KA, Gurewitsch ED. Maternal psychophysiological change during the second half of gestation. Biol Psychol. 2005;69:23–38. doi: 10.1016/j.biopsycho.2004.11.003. [DOI] [PubMed] [Google Scholar]
- 108.Rutter M, Bailey A, Lord C. SCQ: The Social Communication Questionnaire. Los Angeles: Western Psychological Services; 2003. [Google Scholar]
- 109.Louis ED, Barnes LF, Wendt KJ, Albert SM, Pullman SL, Yu Q, Schneier FR. Validity and test-retest reliability of a disability questionnaire for essential tremor. Mov Disord. 2000;15:516–23. doi: 10.1002/1531-8257(200005)15:3<516::AID-MDS1015>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 110.Moodithaya SS, Avadhany ST. Comparison of cardiac autonomic activity between pre and post menopausal women using heart rate variability. Indian J Physiol Pharmacol. 2009;53:227–34. [PubMed] [Google Scholar]
- 111.Sullivan AK, Marcus M, Epstein MP, Allen EG, Anido AE, Paquin JJ, Yadav-Shah M, Sherman SL. Association of FMR1 repeat size with ovarian dysfunction. Hum Reprod. 2005;20:402–12. doi: 10.1093/humrep/deh635. [DOI] [PubMed] [Google Scholar]
- 112.Abidin RR. Parenting Stress Index, Fourth Edition Short Form (PSI-4-SF) Lutz: PAR, Inc; 2013. [Google Scholar]
- 113.Tomeny TS. Parenting stress as an indirect pathway to mental health concerns among mothers of children with autism spectrum disorder. Autism. 2016. doi:10.1177/1362361316655322. [DOI] [PubMed]
- 114.Allen JJ, Chambers AS, Towers DN. The many metrics of cardiac chronotropy: a pragmatic primer and a brief comparison of metrics. Biol Psychol. 2007;74:243–62. doi: 10.1016/j.biopsycho.2006.08.005. [DOI] [PubMed] [Google Scholar]
- 115.Porges SW, Bohrer RE. Analyses of periodic processes in psychophysiological research. In: Cacioppo JT, Tassinary LG, editors. Principles of Psychophysiology: Physical, Social, and Inferential Elements. New York: Cambridge University Press; 1990. pp. 708–53. [Google Scholar]
- 116.Porges SW. Method and apparatus for evaluating rhythmic oscillations in aperiodic physiological response systems. (States U ed., vol. 4520944; 1985.
- 117.Beck AT, Steer RA, Brown GK. Beck Depression Inventory-II. San Antonio: Psychological Corporation; 1996. p. b9. [Google Scholar]
- 118.American Psychological Association . Diagnostic and Statistical Manual of Mental Disorders (DSM-IV) 4. Washington: American Psychological Association; 1994. [Google Scholar]
- 119.Storch EA, Roberti JW, Roth DA. Factor structure, concurrent validity, and internal consistency of the beck depression inventory—second edition in a sample of college students. Depress Anxiety. 2004;19:187–9. doi: 10.1002/da.20002. [DOI] [PubMed] [Google Scholar]
- 120.Sprinkle SD, Lurie D, Insko SL, Atkinson G, Jones GL, Logan AR, Bissada NN. Criterion validity, severity cut scores, and test-retest reliability of the Beck Depression Inventory-II in a university counseling center sample. J Couns Psychol. 2002;49:381. doi: 10.1037/0022-0167.49.3.381. [DOI] [Google Scholar]
- 121.Osman A, Kopper BA, Barrios F, Gutierrez PM, Bagge CL. Reliability and validity of the Beck Depression Inventory-II with adolescent psychiatric inpatients. Psychol Assess. 2004;16:120. doi: 10.1037/1040-3590.16.2.120. [DOI] [PubMed] [Google Scholar]
- 122.Beck AT, Steer RA. Manual for the Beck Anxiety Scale. San Antonio: Pyschological Corportation; 1990. [Google Scholar]
- 123.Fydrich T, Dowdall D, Chambless DL. Reliability and validity of the Beck Anxiety Inventory. J Anxiety Disord. 1992;6:55–61. doi: 10.1016/0887-6185(92)90026-4. [DOI] [Google Scholar]
- 124.de Beurs E, Wilson KA, Chambless DL, Goldstein AJ, Feske U. Convergent and divergent validity of the Beck Anxiety Inventory for patients with panic disorder and agoraphobia. Depress Anxiety. 1997;6:140–6. doi: 10.1002/(SICI)1520-6394(1997)6:4<140::AID-DA2>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 125.Tassone F, Pan R, Amiri K, Taylor AK, Hagerman PJ. A rapid polymerase chain reaction-based screening method for identification of all expanded alleles of the fragile X (FMR1) gene in newborn and high-risk populations. J Mol Diagn. 2008;10:43–9. doi: 10.2353/jmoldx.2008.070073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Filipovic-Sadic S, Sah S, Chen L, Krosting J, Sekinger E, Zhang W, Hagerman PJ, Stenzel TT, Hadd AG, Latham GJ, Tassone F. A novel FMR1 PCR method for the routine detection of low abundance expanded alleles and full mutations in fragile X syndrome. Clin Chem. 2010;56:399–408. doi: 10.1373/clinchem.2009.136101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.LaFauci G, Adayev T, Kascsak R, Kascsak R, Nolin S, Mehta P, Brown WT, Dobkin C. Fragile X screening by quantification of FMRP in dried blood spots by a Luminex immunoassay. J Mol Diagn. 2013;15:508–17. doi: 10.1016/j.jmoldx.2013.02.006. [DOI] [PubMed] [Google Scholar]
- 128.Adayev T, LaFauci G, Dobkin C, Caggana M, Wiley V, Field M, Wotton T, Kascsak R, Nolin SL, Glicksman A. Fragile X protein in newborn dried blood spots. BMC Med Genet. 2014;15:119. doi: 10.1186/s12881-014-0119-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.SAS Institute: SAS Institute version 9.4. Cary NC; 2013.
- 130.Box GE, Cox DR. An analysis of transformations. J R Stat Soc Ser B (Methodological). 1964;26:211–52.
- 131.O’Brien P, Oyebode F. Psychotropic medication and the heart. Adv Psychiatr Treat. 2003;9:414–23. doi: 10.1192/apt.9.6.414. [DOI] [Google Scholar]
- 132.Das S, O’Keefe JH. Behavioral cardiology: recognizing and addressing the profound impact of psychosocial stress on cardiovascular health. Curr Atheroscler Rep. 2006;8:111–8. doi: 10.1007/s11883-006-0048-2. [DOI] [PubMed] [Google Scholar]
- 133.De Meersman RE, Stein PK. Vagal modulation and aging. Biol Psychol. 2007;74:165–73. doi: 10.1016/j.biopsycho.2006.04.008. [DOI] [PubMed] [Google Scholar]
- 134.Cohen J. Statistical power analysis for the behavioral sciences. Hillsdale: L. Erlbaum Associates; 1988. [Google Scholar]
- 135.Lipsey MW. Design sensitivity: Statistical power for experimental research. Newbury Park: Sage; 1990.
- 136.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc Ser B Methodol. 1995;57:289–300. [Google Scholar]
- 137.Thayer JF, Ahs F, Fredrikson M, Sollers JJ, Wager TD. A meta-analysis of heart rate variability and neuroimaging studies: implications for heart rate variability as a marker of stress and health. Neurosci Biobehav Rev. 2012;36:747–56. doi: 10.1016/j.neubiorev.2011.11.009. [DOI] [PubMed] [Google Scholar]
- 138.Segerstrom SC, Nes LS. Heart rate variability reflects self-regulatory strength, effort, and fatigue. Psychol Sci. 2007;18:275–81. doi: 10.1111/j.1467-9280.2007.01888.x. [DOI] [PubMed] [Google Scholar]
- 139.Demaree HA, Robinson JL, Everhart E, Schmeichel BJ. Resting RSA is associated with natural and self-regulated responses to negative emotional stimuli. Brain Cogn. 2004;56:14–24. doi: 10.1016/j.bandc.2004.05.001. [DOI] [PubMed] [Google Scholar]
- 140.Kettunen J, Ravaja N, Naatanen P, Keltikangas-Jarvinen L. The relationship of respiratory sinus arrhythmia to the co-activation of autonomic and facial responses during the Rorschach test. Psychophysiology. 2000;37:242–50. doi: 10.1111/1469-8986.3720242. [DOI] [PubMed] [Google Scholar]
- 141.Schwerdtfeger AR, Schlagert H. The conjoined effect of naturalistic perceived available support and enacted support on cardiovascular reactivity during a laboratory stressor. Ann Behav Med. 2011;42:64–78. doi: 10.1007/s12160-011-9272-2. [DOI] [PubMed] [Google Scholar]
- 142.Geisler FCM, Kubiak T, Siewert K, Weber H. Cardiac vagal tone is associated with social engagement and self-regulation. Biol Psychol. 2013;93:279–86. doi: 10.1016/j.biopsycho.2013.02.013. [DOI] [PubMed] [Google Scholar]
- 143.El-Sheikh M, Harger J, Whitson SM. Exposure to interparental conflict and children’s adjustment and physical health: the moderating role of vagal tone. Child Dev. 2001;72:1617–36. doi: 10.1111/1467-8624.00369. [DOI] [PubMed] [Google Scholar]
- 144.Leary A, Katz LF. Coparenting, family-level processes, and peer outcomes: the moderating role of vagal tone. Dev Psychopathol. 2004;16:593–608. doi: 10.1017/S0954579404004687. [DOI] [PubMed] [Google Scholar]
- 145.El-Sheikh M. Parental drinking problems and children’s adjustment: vagal regulation and emotional reactivity as pathways and moderators of risk. J Abnorm Psychol. 2001;110:499–515. doi: 10.1037/0021-843X.110.4.499. [DOI] [PubMed] [Google Scholar]
- 146.Kok BE, Fredrickson BL. Upward spirals of the heart: autonomic flexibility, as indexed by vagal tone, reciprocally and prospectively predicts positive emotions and social connectedness. Biol Psychol. 2010;85:432–6. doi: 10.1016/j.biopsycho.2010.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Ennis S, Ward D, Murray A. Nonlinear association between CGG repeat number and age of menopause in FMR1 premutation carriers. Eur J Hum Genet. 2006;14:253–5. doi: 10.1038/sj.ejhg.5201510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Tassone F, Iwahashi C, Hagerman PJ. FMR1 RNA within the intranuclear inclusions of fragile X-associated tremor/ataxia syndrome (FXTAS) RNA Biol. 2004;1:103–5. doi: 10.4161/rna.1.2.1035. [DOI] [PubMed] [Google Scholar]
- 149.Koldewyn K, Hessl D, Adams J, Tassone F, Hagerman PJ, Hagerman RJ, Rivera S. Reduced hippocampal activation during recall is associated with elevated FMR1 mRNA and psychiatric symptoms in men with the fragile X premutation. Brain Imaging Behav. 2008;2:105–16. doi: 10.1007/s11682-008-9020-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Tassone F, Greco CM, Hunsaker MR, Seritan AL, Berman RF, Gane LW, Jacquemont S, Basuta K, Jin LW, Hagerman PJ. Neuropathological, clinical and molecular pathology in female fragile X premutation carriers with and without FXTAS. Genes Brain Behav. 2012;11:577–85. doi: 10.1111/j.1601-183X.2012.00779.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Seritan AL, Nguyen DV, Farias ST, Hinton L, Grigsby J, Bourgeois JA, Hagerman RJ. Dementia in fragile X‐associated tremor/ataxia syndrome (FXTAS): comparison with Alzheimer’s disease. Am J Med Genet B Neuropsychiatr Genet. 2008;147:1138–44. doi: 10.1002/ajmg.b.30732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Adams J, Adams P, Nguyen D, Brunberg J, Tassone F, Zhang W, Koldewyn K, Rivera S, Grigsby J, Zhang L. Volumetric brain changes in females with fragile X-associated tremor/ataxia syndrome (FXTAS) Neurology. 2007;69:851–9. doi: 10.1212/01.wnl.0000269781.10417.7b. [DOI] [PubMed] [Google Scholar]
- 153.Kraan CM, Hocking DR, Bradshaw JL, Fielding J, Cohen J, Georgiou-Karistianis N, Cornish KM. Neurobehavioural evidence for the involvement of the FMR1 gene in female carriers of fragile X syndrome. Neurosci Biobehav Rev. 2013;37:522–47. doi: 10.1016/j.neubiorev.2013.01.010. [DOI] [PubMed] [Google Scholar]
- 154.Roberts JE, Boccia ML, Bailey DB, Hatton DD, Skinner M. Cardiovascular indices of physiological arousal in boys with fragile X syndrome. Dev Psychobiol. 2001;39:107–23. doi: 10.1002/dev.1035. [DOI] [PubMed] [Google Scholar]
- 155.Hall SS, Lightbody AA, Huffman LC, Lazzeroni LC, Reiss AL. Physiological correlates of social avoidance behavior in children and adolsecents with fragile X syndrome. J Am Acad Child Adolesc Psychiatry. 2009;48:320–9. doi: 10.1097/CHI.0b013e318195bd15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Grossman P, van Beek J, Wientjes C. A comparison of three quantification methods for the estimation of respiratory sinus arrhythmia. Psychophysiology. 1990;27:702–14. doi: 10.1111/j.1469-8986.1990.tb03198.x. [DOI] [PubMed] [Google Scholar]
- 157.Billman GE. Heart rate variability—a historical perspective. Front Physiol. 2011;2:1–13. doi: 10.3389/fphys.2011.00086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Lewis P, Abbeduto L, Murphy M, Richmond E, Giles N, Bruno L, Schroeder S, Anderson J, Orsmond G. Psychological well-being of mothers of youth with fragile X syndrome: syndrome specificity and within-syndrome variability. J Intellect Disabil Res. 2006;50:894–904. doi: 10.1111/j.1365-2788.2006.00907.x. [DOI] [PubMed] [Google Scholar]
- 159.McLaughlin KA, Rith-Najarian L, Dirks MA, Sheridan MA. Low vagal tone magnifies the association between psychosocial stress exposure and internalizing psychopathology in adolescents. J Clin Child Adolesc Psychol. 2015;44:314–28. doi: 10.1080/15374416.2013.843464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Root AE, Hastings PD, Rubin KH. The parenting behaviors of shy–anxious mothers: The moderating role of vagal tone. J Child Fam Stud. 2015;4:1–9.
- 161.Robinson AR, Roberts JE, McQuillin SD, Brady N, Warren S. Physiological correlates of maternal responsivity in mothers of preschoolers with fragile X syndrome. Am J Intellect Dev Disabil. 2016;121:111–20. doi: 10.1352/1944-7558-121.2.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Weber CS, Thayer JF, Rudat M, Wirtz PH, Zimmermann-Viehoff F, Thomas A, Perschel FH, Arck PC, Deter HC. Low vagal tone is associated with impaired post stress recovery of cardiovascular, endocrine, and immune markers. Eur J Appl Physiol. 2010;109:201–11. doi: 10.1007/s00421-009-1341-x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.