

Abstract
Background:
Decreased high-energy phosphate level is involved in endothelial cell injury and dysfunction. Reduced telomerase activity in endothelial cells in parallel with reduced energy levels might be due to altered direction of alternative splicing machine as a complication of depleted energy during the process of atherosclerosis.
Materials and Methods:
Isolated human umbilical vein endothelial cells (HUVECs) were treated for 24 hours by oligomycine (OM) and 2-deoxy glucose (2-DG). After 24 hours, the effect of energy depletion on telomerase splicing pattern was evaluated using RT-PCR. Indeed, in both treated and untargeted cells, nitric oxide (NO) and von Willebrand factor (vWF) were measured.
Results:
ATP was depleted in treated cells by 43.9% compared with control group. We observed a slight decrease in NO levels (P = 0.09) and vWF (P = 0.395) in the setting of 49.36% ATP depletion. In both groups, no telomerase gene expression was seen. Telomerase and housekeeping gene expression were found in positive control group (colon cancer tissue) and sample tissue.
Conclusions:
The absence of telomerase gene expression in HUVECs might be due to the mortality of these cells or the low level of telomerase gene expression in these cells under normal circumstances.
Keywords: ATP depletion, endothelial cell dysfunction, telomerase, telomere
Introduction
Telomerase is a reverse transcriptase enzyme, which is necessary for proper function of telomeres. It adds TTAGGG repeats to the ends of chromosomes. In this way, it protects terminal ends of chromosomes.[1,2,3,4] Telomerase activity is controlled through various mechanisms, in which alternative splicing is one of them.[5] Alternative splicing is a process in which various proteins are produced from single gene. Among eight alternatively spliced variants of telomerase gene, just full length variant is truly functional.[6] Decreased telomerase activity in endothelial cells is seen in the process of atherosclerosis.[7] This decreased telomerase activity is shown experimentally in regions susceptible to atherosclerosis.[8] The cause of this decrease in telomere length is not well known, however, the theory of telomere shortening is vastly considered during the process of atherogenesis.[9]
Endothelial cell dysfunction is one of the earliest events in the process of atherosclerosis.[10] Indeed, activated endothelial cells pour out UTP, ADP and ATP from themselves. ATP depletion and decrease in energy levels are in parallel with endothelial cell dysfunction.[11,12,13] Reduction in adenosine nucleotide level plays an important role in the process of endothelial cell dysfunction.[13,14] Diminished respiratory rate and ATP synthesis are due to the presence of different stimuli as free fatty acidosis, hypoxia, acidosis, etc.[15,16] Decreased basic level of nitric oxide (NO) is associated with decreased level of ATP/ADP ratio, which is a marker of impaired recovery of cellular phosphorylation potential.[13,14,15,16,17,18] Factors with the ability to maintain endothelial cell function, as 17-beta-hydroxy estrogen, increases efficiency of energy production in endothelial cells.[19] Alternative splicing needs correct regulation, in which its mechanisms are not yet fully understood. Alternative splicing is an energy dependent process and in the setting of appropriate energy levels, incorrect splicing will occur.[20,21]
Decreased telomerase activity occurs in the process of endothelial cell dysfunction and atherosclerosis. Indeed, decreased ATP/ADP ratio is in parallel with endothelial cell dysfunction and atherosclerosis. Thus, it seems that this telomere dysfunction might be due to decreased energy levels, which could be attributed to impaired alternative splicing. Thus, in this study, we aimed to investigate the role of impaired energy balance on telomerase alternative splicing during the process of endothelial cell dysfunction and atherosclerosis.
Materials and Methods
Isolation of HVECs
Isolated placenta and cord were inserted in cold environment (4°C). Umbilical cord was inserted in a paper towel to get rid from blood and collection medium. A transverse incision was made at terminal end of placenta to determine vein and artery. Then, vein was cannulated after ligation of both terminal ends of vein. Internal lumen of cannula was filled with phosphate-buffer saline (PBS) (in both ends). After dilation of lumen, vein was transferred into incubator (37°C for 10 min). Then, umbilical cord was exited from incubator. Cells were isolated by manual massage and suction. Venus lumen was re-filled with prewarmed PBS. Internal lumen was filled by collagenase solution and cells were centrifugated for 5 min at 1000 rpm. Supernatant was transferred into tissue culture flask T25 with 4 ml of serum including culture medium without growth factor included. The next day, tissue culture medium was removed. Cells were washed using prewarmed serum-free culture medium. Then, 4 ml of tissue culture was added into cells. Cells were feeded with fresh serum plus culture medium. Cells were recultured after removing culture medium from confluent cells. Then, 1 ml of Trypsin-EDTA was added into flask. Flask was transferred into incubator for 5 min. After this stage, equal amounts of culture medium plus growth factor was added to cells. Cells were transferred into incubator and were feeded by changing culture medium every other day. In this study, cells were passaged for three times. Cultured cells were divided into test and control groups. Test cells were treated by oligomycine (OM) and 2-DG for 6 hours.[1] Simultaneous administration of these agents leads to 50% reduction in ATP level without induction of apoptosis.[1]
Endothelial cell characterization
Endothelial cells were characterized using observation of cobble-stone morphology under microscopic evaluation [Figure 1]. Endothelial cells were identified using flow cytometry by anti-CD34 (PECAM) monoclonal antibody.
Figure 1.
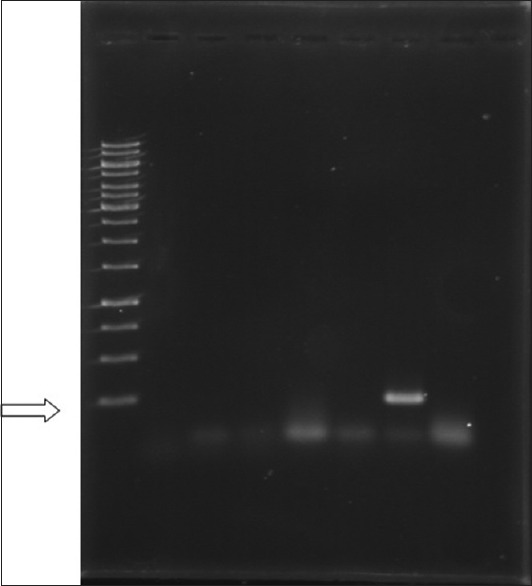
Photo indicates the presence of telomerase (4120 bp), 100 bp DNA ladder
Nucleotide measurement
ATP/ADP ratio was measured as a marker of endothelial cell dysfunction. Changes in ATP/ADP ratio were measured using measured by bioluminator detection. This kit works based on luciferase enzyme. About 10000 cells were used. Nearly 103-104 (10 μL) of cells were transferred into tissue culture palates after removing culture medium. Next, 100 μL of nucleus releasing buffer was added into cells and cells were incubated in room temperature for 5 min. A total of 1 μL of ATP monitoring enzyme was added into cell lysate and samples were read out in luminator.
Nitric oxide measurement
NO was measured in conditioned medium indirectly using quantification of nitrites by total NO and nitrate/nitrite assay kit per protocol. NO level of both treated and nontreated cells were measured.
Von Willebrand factor measurement
Extracellular von Willebrand factor (vWF) was measured in HUVCs embedded in 96-well plates pre- and post-ATP depletion through Asserachrom vWF kit per protocol (Diagnostica Stago, France).
Expression of telomerase alternative splicing variants after ATP depletion
About 6 hours after induction of ATP depletion, cellular RNA was extracted using RNA extraction kit, per protocol. The following primer pairs were used:[22]
hTERT980S/hTERT1303A, hTERT2164S/hTERT 2620A, hTERT1241S/hTERT1835A, hTERT1783S/hTERT2247A
PCR conditions were as follows:
94°C for 60 seconds
33 cycles of (96°C for 20 seconds, 68°C for 50 seconds, 72°C for 50 seconds)
PCR products were electrophorezed on polyacrylamide gel.
As a positive control, we used a tissue with certain telomerase activity (colon cancer tissue). Beta-actin gene expression was used as a house-keeping gene to verify the accuracy of RNA extraction. Beta-actin primer pairs were as follows:
Forward primer 1 GCACTCTTCCAGCCTTCCTTC 21
Template 862.882
Reverse primer 1 CAGCACTGTGTTGGCGTACA 20
Template 981. 962
Statistical analysis was performed using SPSS version 17.00 by K2 test.
Results
ATP depletion
ATP was reduced in treated cells by 43.9%. Luminecance change in ATP-depleted cells was 5082.5000 ± 2147.48329 (mean ± SD). In these cells, ATP level was nonsignificantly reduced, which was due to the applied method to avoid cell apoptosis. In control cells, ATP level showed nonsignificant change. In cells of control group, ATP was 22711.5000 ± 7845.34974 (mean ± SD).
ATP depletion and NO release
We observed a slight decrease (16.32%) in NO release between induced and uninduced endothelial cells (7.23 ± 1.13 vs. 8.64 ± 0.85 μm/L, P = 0.09, n = 3). Indeed, our data demonstrated a marginal decrease (15.73%) in secreted vWF between treated and untreated cells (0.0876 ± 0.0087 vs. 0.1056 ± 0.023885 μm/L, P = 0.395, n = 3) [Table 1].[23]
Table 1.
Effect of ATP depletion on NO level and vWF secretion of human umbilical vein endothelial cells

Telomerase gene expression
In both groups, no expression of telomerase gene expression was found. Since no telomerase gene expression was found in human umbilical vein endothelial cells (HUVECs), we investigated telomerase gene expression in colon cancer tissue. In this tissue sample, the presence of teloermase gene expression (380 bp) and full length telomerase gene (475 bp) were seen [Figures 2 and 3]. Other variants were not found in this sample. Expression of beta-actin as a house keeping gene confirmed the accuracy of RNA expression method in our survey [Figure 4].
Figure 2.
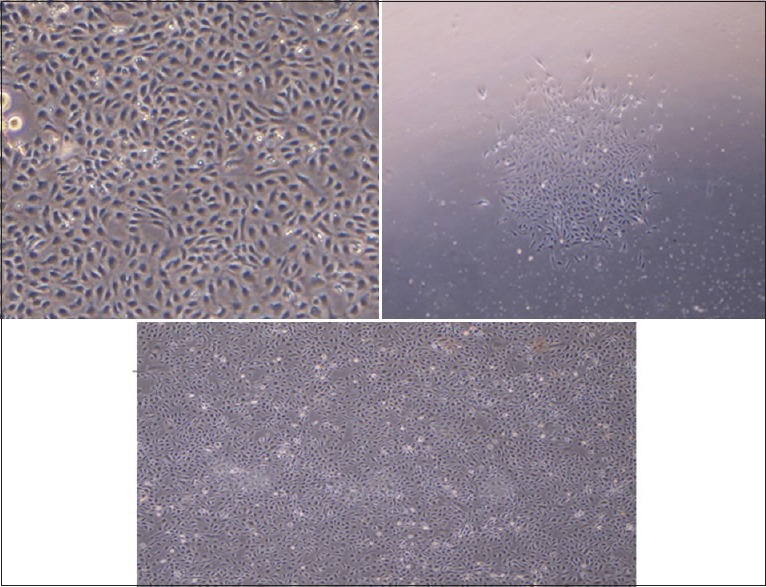
Cobble-stone morphology of HUVCs
Figure 3.
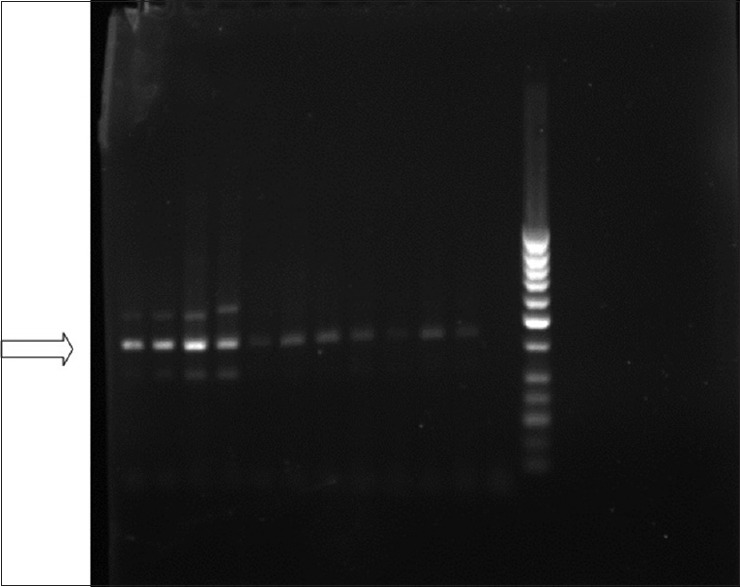
The photo indicates the presence of telomerase (323 bp) using hTERT980S and hTERT1303A Primer pairs (50 bp DNA ladder)
Figure 4.
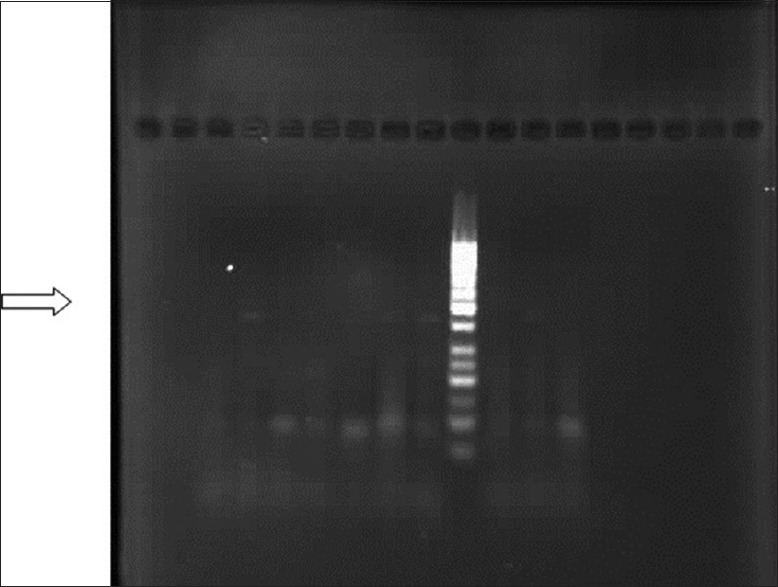
The photo indicates the presence of telomerase (457 bp) using hTERT2164S and hTERT2620A primer pairs (50 bp DNA ladder)
Discussion
Our data implies induced ATP depletion and consequently induced decreased energy level in treated cells. In this setting, NO and vWF factors were measured in which full discussion in values are explained by Behjati et al.[23] In the setting of both ATP depletion and normal energy balance, we observed no telomerase gene expression in HUVECs; thus we used a tissue with certain telomerase gene expression. Full length telomerase gene was found in this tissue. Other variants were negative in this tissue. Expression of beta-actin gene, confirms the accuracy of RNA extraction. These data demonstrate that telomerase gene expression is very low or zero in isolated HUVECs, which is undetectable by RT-PCR. These cells are mortal. In previous studies, immortalized HUVECs using transfected TERT or other vectors were used.[24,25] In this way, endothelial cell lines were preferred. But since we aimed to evaluate the effect of impaired energy balance on telomerase activity, we were not allowed to use immortal cells. It may be possible that these cells possess very low amounts of telomerase activity, undetectable by RT-PCR. These cells may also have no telomerase activity.
In order to evaluate telomerase gene expression in HUVECs, various methods have been used. Quantitative real time RT-PCR, PCR-based TRAP analysis, linear range of TRAP assay are most often used methods.[26,27,28,29,30] Some investigators have also used RT-PCR for assessment of telomerase gene expression in HUVECs in instances like assessment of telomerase activity by RT-PCR after induction of TERT over-expression.[31,32] Indeed, RT-PCR was used to assess telomerase gene expression in HUVECs after administration of agents, which increase telomerase gene expression as 17beta-estradiol. Since both morphology and flow cytometry analysis, confirm the through isolation procedure of endothelial cells, it seems interesting that why telomerase gene expression was evaluated by semi-quantitative RT-PCR by some researchers but with similar applied method, it was undetectable in our survey. So, is semi-quantitative RT-PCR a suitable method for evaluation of telomerase gene expression in HUVECs?
Conclusions
The absence of telomerase gene expression in HUVECs might be due to the mortality of these cells or the low level of telomerase gene expression in these cells under normal circumstances.
Financial support and sponsorship
Nil.
Conflicts of interest
There are no conflicts of interest.
References
- 1.Kilian A, Bowtell DD, Abud HE, Hime GR, Venter DJ, Keese PK, et al. Isolation of a candidate human telomerase catalytic subunit gene, which reveals complex splicing patterns in different cell types. Hum Mol Genet. 1997;6:2011–9. doi: 10.1093/hmg/6.12.2011. [DOI] [PubMed] [Google Scholar]
- 2.Derradji H, Bekaert S, Van Oostveldt P, Baatout S. Comparison of different protocols for telomere length estimation by combination of quantitative fluorescence in situ hybridization (Q-FISH) and flow cytometry in human cancer cell lines. Anticancer Res. 2005;25:1039–50. [PubMed] [Google Scholar]
- 3.Pandita TK, Roti Roti JL. Role of telomerase in radiocurability (review) Oncol Rep. 2003;10:263–70. [PubMed] [Google Scholar]
- 4.Lantuéjoul S, Salon C, Soria JC, Brambilla E. Telomerase expression in lung preneoplasia and neoplasia. Int J Cancer. 2007;120:1835–41. doi: 10.1002/ijc.22473. [DOI] [PubMed] [Google Scholar]
- 5.Liu WJ, Zhang YW, Zhang ZX, Ding J. Alternative splicing of human telomerase reverse transcriptase may not be involved in telomerase regulation during all-trans-retinoic acid-induced HL-60 cell differentiation. J Pharmacol Sci. 2004;96:106–14. doi: 10.1254/jphs.fp0030600. [DOI] [PubMed] [Google Scholar]
- 6.Ulaner GA, Hu JF, Vu TH, Giudice LC, Hoffman AR. Tissue-specific alternate splicing of human telomerase reverse transcriptase (hTERT) influences telomere lengths during human development. Int J Cancer. 2001;91:644–9. [PubMed] [Google Scholar]
- 7.Pu DR, Liu L. HDL slowing down endothelial progenitor cells senescence: A novel anti-atherogenic property of HDL. Med Hypotheses. 2008;70:338–42. doi: 10.1016/j.mehy.2007.05.025. [DOI] [PubMed] [Google Scholar]
- 8.Minamino T, Miyauchi H, Yoshida T, Ishida Y, Yoshida H, Komuro I. Endothelial cell senescence in human atherosclerosis: Role of telomere in endothelial dysfunction. Circulation. 2002;105:1541–4. doi: 10.1161/01.cir.0000013836.85741.17. [DOI] [PubMed] [Google Scholar]
- 9.Minamino T, Komuro I. Role of telomere in endothelial dysfunction in atherosclerosis. Curr Opin Lipidol. 2002;13:537–43. doi: 10.1097/00041433-200210000-00010. [DOI] [PubMed] [Google Scholar]
- 10.Jian W, Arora JS, Oe T, Shuvaev VV, Blair IA. Induction of endothelial cell apoptosis by lipid hydroperoxide-derived bifunctional electrophiles. Free Radic Biol Med. 2005;39:1162–76. doi: 10.1016/j.freeradbiomed.2005.06.008. [DOI] [PubMed] [Google Scholar]
- 11.Beldi G, Enjyoji K, Wu Y, Miller L, Banz Y, Sun X, et al. The role of purinergic signaling in the liver and in transplantation: Effects of extracellular nucleotides on hepatic graft vascular injury, rejection and metabolism. Front Biosci. 2008;13:2588–603. doi: 10.2741/2868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mathews MT, Berk BC. PARP-1 inhibition prevents oxidative and nitrosative stress-induced endothelial cell death via transactivation of the VEGF receptor 2. Arterioscler Thromb Vasc Biol. 2008;28:711–7. doi: 10.1161/ATVBAHA.107.156406. [DOI] [PubMed] [Google Scholar]
- 13.Carlucci F, Marinello E, Rosi F, Floccari F, Gerunda G, Neri D, et al. Nitric oxide generation is associated with an unbalance of protein tyrosine phosphatases during liver transplantation. Biomed Pharmacother. 2007;61:216–21. doi: 10.1016/j.biopha.2007.03.005. [DOI] [PubMed] [Google Scholar]
- 14.Hurst RD, Azam S, Hurst A, Clark JB. Nitric-oxide-induced inhibition of glyceraldehyde-3-phosphate dehydrogenase may mediate reduced endothelial cell monolayer integrity in an in vitro model blood-brain barrier. Brain Res. 2001;894:181–8. doi: 10.1016/s0006-8993(01)01992-8. [DOI] [PubMed] [Google Scholar]
- 15.Hermans MP. Diabetes and the endothelium. Acta Clin Belg. 2007;62:97–101. doi: 10.1179/acb.2007.017. [DOI] [PubMed] [Google Scholar]
- 16.Agulló L, Garcia-Dorado D, Escalona N, Inserte J, Ruiz-Meana M, Barrabés JA, et al. Hypoxia and acidosis impair cGMP synthesis in microvascular coronary endothelial cells. Am J Physiol Heart Circ Physiol. 2002;283:H917–25. doi: 10.1152/ajpheart.01067.2001. [DOI] [PubMed] [Google Scholar]
- 17.de la Torre JC, Stefano GB. Evidence that Alzheimer's disease is a microvascular disorder: The role of constitutive nitric oxide. Brain Res Brain Res Rev. 2000;34:119–36. doi: 10.1016/s0165-0173(00)00043-6. [DOI] [PubMed] [Google Scholar]
- 18.Ciapaite J, van Bezu J, van Eikenhorst G, Bakker SJ, Teerlink T, Diamant M, et al. Palmitate and oleate have distinct effects on the inflammatory phenotype of human endothelial cells. Biochim Biophys Acta. 2007;1771:147–54. doi: 10.1016/j.bbalip.2006.12.005. [DOI] [PubMed] [Google Scholar]
- 19.Razmara A, Sunday L, Stirone C, Wang XB, Krause DN, Duckles SP, et al. Mitochondrial effects of estrogen are mediated by estrogen receptor alpha in brain endothelial cells. J Pharmacol Exp Ther. 2008;325:782–90. doi: 10.1124/jpet.107.134072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Krainer AR, Maniatis T, Ruskin B, Green MR. Normal and mutant human beta-globin pre-mRNAs are faithfully and efficiently spliced in vitro. Cell. 1984;36:993–1005. doi: 10.1016/0092-8674(84)90049-7. [DOI] [PubMed] [Google Scholar]
- 21.Hartl DL, Elizabeth W. Genetics: Analysis of Genes and Genomes. Sudbury, Massachusetts: Jones and Bartlett Publishers [My paper]; 2005. Jan 1, 2005. [Google Scholar]
- 22.Ulaner GA, Hu JF, Vu TH, Giudice LC, Hoffman AR. Tissue-specific alternate splicing of human telomerase reverse transcriptase (hTERT) influences telomere lengths during human development. Int J Cancer. 2001;91:644–9. [PubMed] [Google Scholar]
- 23.Behjati M, Hashemi M, Kazemi M, Salehi M. Nitric oxide level and von Willebrand factor (vWF) secretion are not candidate markers of endothelial cell dysfunction in adenosine triphosphate (ATP) depleted endothelial cells. Afr J Biotechnol. 2011;10:8871–6. [Google Scholar]
- 24.Noguchi H, Kobayashi N, Westerman KA, Sakaguchi M, Okitsu T, Totsugawa T, et al. Controlled expansion of human endothelial cell populations by Cre-loxP -based reversible immortalization. Hum Gene Ther. 2002;13:321–34. doi: 10.1089/10430340252769833. [DOI] [PubMed] [Google Scholar]
- 25.Tentori L, Vergati M, Muzi A, Levati L, Ruffini F, Forini O, et al. Generation of an immortalized human endothelial cell line as a model of neovascular proliferating endothelial cells to assess chemosensitivity to anticancer drugs. Int J Oncol. 2005;27:525–35. [PubMed] [Google Scholar]
- 26.Falchetti ML, Mongiardi MP, Fiorenzo P, Petrucci G, Pierconti F, D’Agnano I, et al. Inhibition of telomerase in the endothelial cells disrupts tumor angiogenesis in glioblastoma xenografts. Int J Cancer. 2008;122:1236–42. doi: 10.1002/ijc.23193. [DOI] [PubMed] [Google Scholar]
- 27.Chang H, Rha SY, Jeung HC, Park KH, Kim TS, Kim YB, et al. Telomerase- and angiogenesis -related gene responses to irradiation in human umbilical vein endothelial cells. Int J Mol Med. 2013;31:1202–8. doi: 10.3892/ijmm.2013.1300. [DOI] [PubMed] [Google Scholar]
- 28.Ling S, Zhou L, Li H, Dai A, Liu JP, Komesaroff PA, et al. Effects of 17 beta-estradiol on growth and apoptosis in human vascular endothelial cells: Influence of mechanical strain and tumor necrosis factor-alpha. Steroids. 2006;71:799–808. doi: 10.1016/j.steroids.2006.05.010. [DOI] [PubMed] [Google Scholar]
- 29.Vasa M, Breitschopf K, Zeiher AM, Dimmeler S. Nitric oxide activates telomerase and delays endothelial cell senescence. Circ Res. 2000;87:540–2. doi: 10.1161/01.res.87.7.540. [DOI] [PubMed] [Google Scholar]
- 30.Mortuza R, Chen S, Feng B, Sen S, Chakrabarti S. High glucose induced alteration of SIRTs in endothelial cells causes rapid aging in a p300 and FOXO regulated pathway. PLoS One. 2013;8:e54514. doi: 10.1371/journal.pone.0054514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Takano H, Murasawa S, Asahara T. Functional and gene expression analysis of hTERT over expressed endothelial cells. Biologics. 2008;2:547–54. doi: 10.2147/btt.s2479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Ingram DA, Mead LE, Moore DB, Woodard W, Fenoglio A, Yoder MC. Vessel wall-derived endothelial cells rapidly proliferate because they contain a complete hierarchy of endothelial progenitor cells. Blood. 2005;105:2783–6. doi: 10.1182/blood-2004-08-3057. [DOI] [PubMed] [Google Scholar]