

Abstract
The ability to identify the largest amount of prey available is fundamental for optimizing foraging behaviour in several species. To date, this cognitive skill has been observed in all vertebrate groups except reptiles. In this study we investigated the spontaneous ability of ruin lizards to select the larger amount of food items. In Experiment 1, lizards proved able to select the larger food item when presented with two alternatives differing in size (0.25, 0.50, 0.67 and 0.75 ratio). In Experiment 2 lizards presented with two groups of food items (1 versus 4, 2 versus 4, 2 versus 3 and 3 versus 4 items) were unable to select the larger group in any contrast. The lack of discrimination in the presence of multiple items represents an exception in numerical cognition studies, raising the question as to whether reptiles' quantitative abilities are different from those of other vertebrate groups.
Keywords: lizard, reptile cognition, numerical abilities, continuous quantities
1. Introduction
The ability to estimate and compare quantities is of prime importance for several aspects of the relationship between an animal and its natural environment. Such competence is useful to assess the relative number of opponents in potentially aggressive interactions (e.g. hyenas, Crocuta crocuta, [1]) and provides an advantage in foraging decisions as several mammals, birds, amphibians and fish maximize their energy intake by selecting the larger group of food items available (e.g. chimpanzees, Pan troglodytes, [2]; pigeons, Columba livia, [3]; salamanders, Plethodon cinereus, [4]; and guppies, Poecilia reticulata, [5]). Quantitative abilities are essential for both prey and predator survival. On one hand, the risk of an individual of being spotted by a predator diminishes as the quantity of conspecifics in the group increases (a phenomenon called ‘dilution effect’: [6]). On the other hand, predators adjust their predation strategy depending on the number of prey available [7]. Interestingly, the performance of mammals, birds, amphibians and fish on relative quantity judgements varies as a function of ratio in line with Weber's law which states that the just-noticeable difference between two stimuli is proportional to the magnitude of the stimuli themselves. These similarities suggest that animals most likely share a similar numerical system [8].
To date, existing studies in this field have investigated all vertebrate groups except reptiles. However, there is no reason to believe that the selective pressures in favour of the ability to estimate different quantities should not have acted on this vertebrate group. To fill this gap, we investigated quantitative abilities in ruin lizards (Podarcis sicula) using the most common procedure adopted in this field, a free choice test in the presence of food items differing in quantity [2–5,9]. In detail, in Experiment 1 we tested whether lizards attempt to maximize the amount of food intake. Subjects could choose between two pieces of food items differing in size. Four different ratios comparing items' sizes were used: 0.25, 0.50, 0.67 and 0.75. Experiment 2 was designed to assess quantitative abilities of lizards in the presence of two groups of food items differing in number. The same ratios as in Experiment 1 were presented: 1 versus 4 (ratio 0.25), 2 versus 4 (0.50), 2 versus 3 (0.67) and 3 versus 4 (0.75).
2. Material and methods
(a). Subjects
Subjects were 27 adult ruin lizards, Podarcis sicula (12 subjects in Experiment 1 and 15 in Experiment 2), collected from the area of Ferrara and maintained at the Department of Life Sciences and Biotechnology, University of Ferrara. However, a total of nine lizards (three in Experiment 1 and six in Experiment 2) were not admitted to the testing phase. As a consequence, the total sample consisted of nine subjects in Experiment 1 (eight males, one female), and nine subjects in Experiment 2 (seven males, two females).
(b). Apparatus and stimuli
Subjects were tested in a Y-shaped enclosure divided into a tunnel that served as holding area and an experimental compartment where stimuli were presented during testing (figure 1). Lizards could view the stimuli from the top of a ramp inserted into the tunnel before entering the experimental compartment (additional details can be found in the electronic supplementary material, S1).
Figure 1.
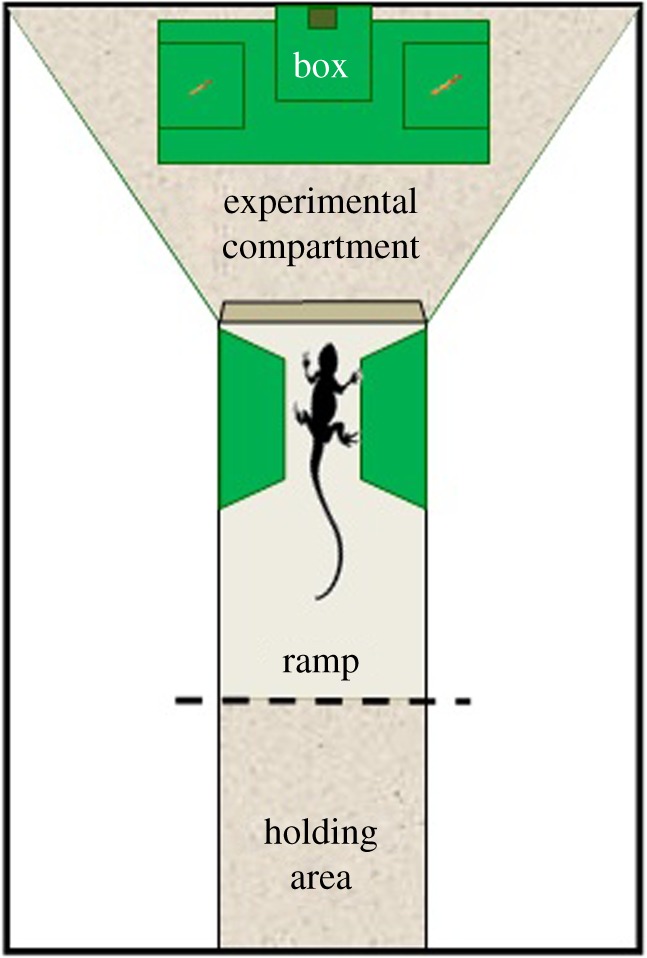
Schematic representation of the experimental apparatus used in both experiments (top view). Example of a trial of Experiment 1 (i.e. the lizards were presented with a choice between two food items of different size). (Online version in colour.)
Stimuli consisted of Musca domestica larvae placed on two supports in the centre of the experimental compartment; they were equidistant with respect to the subject's path of approach. The size, number and arrangements of the larvae on each support varied according to the schedule of each experiment. After the subject made its choice, a green box was placed over the unchosen stimulus to prevent its consumption.
To reduce the possibility of subjects using olfactory cues, the experimental compartment was saturated with the smell of six non-visible larvae (for details see the electronic supplementary material, S1).
(c). Experimental procedure
We performed two experiments with the same apparatus and basic procedure (see the electronic supplementary material, S1).
After a 3-day acclimation phase to familiarize the lizards with the experimental apparatus and procedure, the subjects entered the test phase. The lizards underwent a series of trials in which two stimuli differing either in size or in number of food items were presented. Only one food set could be selected per trial. Once the subject had chosen one stimulus (by approaching one support), it was allowed to eat the content while the other was gently covered with the green box (see the electronic supplementary material, video S2). Stimuli were presented in a pseudo-random sequence with the restriction that the larger/smaller stimulus was never presented more than twice in a row on the same side. The left–right position of the stimuli was counterbalanced over trials.
(d). Experiment 1
In this Experiment, we investigated whether lizards are able to discriminate between food items differing in size. Subjects were observed in their spontaneous preference between pairs (1 versus 1) of differently sized larvae (range from 5 to 25 mm2), one on each support, with four size ratios within each pair (0.25, 0.50, 0.67 and 0.75). The lizards underwent a total of 64 trials (16 for each discrimination) over 11 days in which the size discriminations were intermingled across trials.
(e). Experiment 2
We investigated lizards’ choices between two sets of equally sized food items (5 mm2) differing in number. Four numerical discriminations were presented: 1 versus 4, 2 versus 4, 2 versus 3 and 3 versus 4 (the same ratios used in Experiment 1: 0.25, 0.50, 0.67 and 0.75, respectively). The lizards underwent a total of 64 trials (16 for each discrimination) over 16 days in which the numerical discriminations were intermingled across trials.
For statistical analyses we computed the proportion of choices for the set with the larger food item (Experiment 1) or the larger number of food items (Experiment 2).
3. Results
A mixed-model ANOVA on Ratio (0.25/0.50/0.67/0.75) as within-subjects factor and Experiment (Experiment 1/Experiment 2) as between-subjects factor showed a main effect of Experiment (F1,16 = 55.012, p < 0.001, partial eta squared 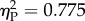 ) and Ratio (F3,48 = 9.735, p < 0.001,
) and Ratio (F3,48 = 9.735, p < 0.001, 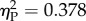 ). The interaction was marginally non-significant (F3,48 = 2.44, p = 0.076,
). The interaction was marginally non-significant (F3,48 = 2.44, p = 0.076, 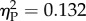 ). Trend analysis indicated a significant decrease of performance as a function of Ratio (p = 0.035). We also found a significant Experiment × Ratio trend interaction (p = 0.032). Separate analyses for the two experiments showed that this was due to the fact that discrimination significantly decreased with Ratio in Experiment 1 (repeated measures ANOVA: F3,24 = 12.674, p < 0.001,
). Trend analysis indicated a significant decrease of performance as a function of Ratio (p = 0.035). We also found a significant Experiment × Ratio trend interaction (p = 0.032). Separate analyses for the two experiments showed that this was due to the fact that discrimination significantly decreased with Ratio in Experiment 1 (repeated measures ANOVA: F3,24 = 12.674, p < 0.001, 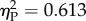 ; linear trend: p < 0.001) but not in Experiment 2 (F3,24 = 2.139, p = 0.122,
; linear trend: p < 0.001) but not in Experiment 2 (F3,24 = 2.139, p = 0.122, 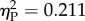 ; figure 2). Bonferroni post hoc tests in Experiment 1 revealed that 0.25 ratio differed from 0.67 (p = 0.017) and 0.75 (p = 0.002) and 0.50 differed from 0.67 (p = 0.045) and 0.75 (p = 0.023). Lizards preferred the larger quantity in any ratio in Experiment 1. No significant preference for any ratio in Experiment 2 was found (table 1).
; figure 2). Bonferroni post hoc tests in Experiment 1 revealed that 0.25 ratio differed from 0.67 (p = 0.017) and 0.75 (p = 0.002) and 0.50 differed from 0.67 (p = 0.045) and 0.75 (p = 0.023). Lizards preferred the larger quantity in any ratio in Experiment 1. No significant preference for any ratio in Experiment 2 was found (table 1).
Figure 2.
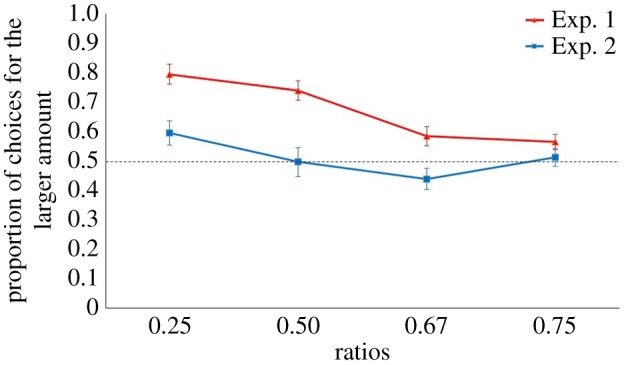
Results. Proportion of choices for the larger amount is plotted against the ratio of quantities presented in Experiments 1 and 2. A significant choice for the larger amount was found in all contrasts of Experiment 1, while no choice in any numerical ratio was found in Experiment 2. The dotted line represents chance level (0.50). Bars refer to standard errors. (Online version in colour.)
Table 1.
Performance of lizards as a function of the ratio in Experiments 1 and 2.
| experiment | ratio | one-sample t-test | Cohen's d |
|---|---|---|---|
| Experiment 1 | 0.25 | t8 = 7.476, p < 0.001 | 2.492 |
| 0.50 | t8 = 6.459, p < 0.001 | 2.153 | |
| 0.67 | t8 = 2.501, p = 0.037 | 0.837 | |
| 0.75 | t8 = 2.488, p = 0.038 | 0.829 | |
| Experiment 2 | 0.25 (1 versus 4) | t8 = 2.141, p = 0.065 | 0.714 |
| 0.50 (2 versus 4) | t8 = −0.093, p = 0.928 | 0.031 | |
| 0.67 (2 versus 3) | t8 = −1.685, p = 0.130 | 0.561 | |
| 0.75 (3 versus 4) | t8 = 0.378, p = 0.715 | 0.126 |
4. Discussion
This study provides the first evidence of quantitative abilities in reptiles, a group of vertebrates not previously investigated in this field.
Experiment 1 showed that lizards prefer to reach the larger amount of food, being able to select the largest items in all contrasts presented, probably in order to optimize food intake. In this test, numerical information was made irrelevant (1 versus 1); hence the ability to select the larger amount was entirely based on the comparison of items’ size. However, their performance was increasingly more precise as the ratio between the smaller and the larger quantity decreased, in line with Weber's law. The capacity to discriminate these ratios and ratio-dependence of the performance strictly resemble those observed in other vertebrates tested with similar size-discrimination tasks (e.g. chimpanzees [2]; salamanders [4]; guppies [5]).
On the contrary, in Experiment 2 lizards seem to be unable to optimize food intake when multiple items are available, showing no preference for either group in the presence of food presented in discrete quantities. As far as we are aware, these data represent a true exception in numerical cognition studies. For instance, when presented with food items differing in number, fish are able to discriminate up to a 0.50 ratio [5], dogs up to 0.67 [9], pigeons up to a 0.86 ratio [3], and apes up to a 0.90 ratio [10].
Two hypotheses could be advanced. First, it is possible that lizards’ motivation is different in the two tests. In nature, while an individual is reaching one prey in the patch, other conspecifics could try to capture other ones. Hence, we could hypothesize an advantage for the individual that detects and consumes the largest prey first. As suggested in other species [2,5], natural selection might have favoured decision mechanisms that prioritize the search for larger prey instead of the search for patches containing more overall prey.
As an alternative, the lack of preference observed in Experiment 2 may reflect a true limit in lizards’ ability to process quantitative information. If so, this study would raise an interesting issue in the evolution of numerical abilities. As different species show similar performance in numerical tasks, the existence of a core number system shared by all vertebrates and inherited by a common ancestor is widely accepted [8,11]. Hence, a similar performance between reptiles and closely related vertebrate groups would be expected. We did not observe this pattern, as fish species show higher numerical abilities than lizards. One potential explanation for this evolutionary paradox could be related to the potential genetic advantage enjoyed by teleost fish during their early evolution, a whole-genome duplication that took place within the ray-finned fishes but after their divergence from the lineage leading to land vertebrates. Recently, Schartl et al. [12] found higher duplicate retention rate for putative cognition-related genes in fish. The possibility exists that this early evolutionary event may have actually promoted the appearance in fish of complex cognitive skills (including numerical abilities) that are similar to mammals' and not observed in closely related vertebrate groups, a hypothesis that needs to be tested in the future.
Supplementary Material
Supplementary Material
Ethics
All husbandry and experimental procedures complied with the European Legislation for the Protection of Animals Used for Scientific Purposes (Directive 2010/63/EU). The experimental protocol was previously authorized by the University of Ferrara Institutional Animal Care and Use Committee and by the Italian Ministry of Health (authorization number 235/2015-PR).
Data accessibility
The dataset is available from the electronic supplementary material, S3.
Authors' contributions
M.E.M.P., A.B., C.B. and A.F. developed the study concept; M.E.M.P., C.A., A.B., C.B. and A.F. contributed to the study design; I.F. and F.G. collected the data; M.E.M.P., I.F. and F.G. analysed the data; M.E.M.P., C.A. and A.B. drafted the manuscript. All authors contributed to the revisions of this manuscript, agreed to be held accountable for this work and approved the final version of the manuscript for publication.
Competing interests
We declare we have no competing interests.
Funding
Funding was provided to M.E.M.P. and C.A. from MIUR (FIRB 2013, N. RBFR13KHFS) and to C.B. and A.F. from the University of Ferrara (FAR 2013 and 2014).
References
- 1.Benson-Amram S, Heinen VK, Dryer SL, Holekamp KE. 2011. Numerical assessment and individual call discrimination by wild spotted hyaenas, Crocuta crocuta. Anim. Behav. 82, 743–752. ( 10.1016/j.anbehav.2011.07.004) [DOI] [Google Scholar]
- 2.Beran MJ, Evans TA, Harris EH. 2008. Perception of food amounts by chimpanzees based on the number, size, contour length and visibility of items. Anim. Behav. 75, 1793–1802. ( 10.1016/j.anbehav.2007.10.035) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Emmerton J, Delius JD. 1993. Beyond sensation: visual cognition in pigeons. In Vision, brain, and behavior in birds (eds Zeigler HP, Bischof HJ), pp. 377–390. Cambridge, MA: MIT Press. [Google Scholar]
- 4.Uller C, Jaeger R, Guidry G, Martin C. 2003. Salamanders (Plethodon cinereus) go for more: rudiments of number in an amphibian. Anim. Cogn. 6, 105–112. ( 10.1007/s10071-003-0167-x) [DOI] [PubMed] [Google Scholar]
- 5.Lucon-Xiccato T, Miletto Petrazzini ME, Agrillo C, Bisazza A. 2015. Guppies discriminate between two quantities of food items but prioritize item size over total amount. Anim. Behav. 107, 183–191. ( 10.1016/j.anbehav.2015.06.019) [DOI] [Google Scholar]
- 6.Foster WA, Treherne JE. 1981. Evidence for the dilution effect in the selfish herd from fish predation on a marine insect. Nature 293, 466–467. ( 10.1038/293466a0) [DOI] [Google Scholar]
- 7.Botham MS, Kerfoot CJ, Louca V, Krause J. 2005. Predator choice in the field; grouping guppies, Poecilia reticulata, receive more attacks. Behav. Ecol. Sociobiol. 59, 181–184. ( 10.1007/s00265-005-0018-7) [DOI] [Google Scholar]
- 8.Feigenson L, Dehaene S, Spelke ES. 2004. Core systems of number. Trends Cogn. Sci. 8, 307–314. ( 10.1016/j.tics.2004.05.002) [DOI] [PubMed] [Google Scholar]
- 9.Miletto Petrazzini ME, Wynne CD. 2016. What counts for dogs (Canis lupus familiaris) in a quantity discrimination task? Behav. Proc. 122, 90–97. ( 10.1016/j.beproc.2015.11.013) [DOI] [PubMed] [Google Scholar]
- 10.Hanus D, Call J. 2007. Discrete quantity judgments in the great apes (Pan paniscus, Pan troglodytes, Gorilla gorilla, Pongo pygmaeus). J. Comp. Psychol. 121, 241–249. ( 10.1037/0735-7036.121.3.241) [DOI] [PubMed] [Google Scholar]
- 11.Beran MJ. 2008. The evolutionary and developmental foundations of mathematics. PLoS Biol. 6, e19 ( 10.1371/journal.pbio.0060019) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schartl M, et al. 2013. The genome of the platyfish, Xiphophorus maculatus, provides insights into evolutionary adaptation and several complex traits. Nat. Genet. 45, 567–572. ( 10.1038/ng.2604) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The dataset is available from the electronic supplementary material, S3.