

ABSTRACT
Tumor-associated macrophages (TAM) are well known as a key player in the tumor microenvironment, which support cancer progression. More recently, a lineage of monocytes characterized by the expression of the TIE-2/Tek angiopoietin receptor identified a subset of circulating and tumor-associated monocytes endowed with proangiogenic activity. TIE-2 expressing monocytes (TEM) were found both in humans and mice. Here, we review the phenotypes and functions of TEM reported so far in human cancer and their potential use as markers of cancer progression and metastasis. Finally, we discuss the therapeutic approaches currently used or proposed to target TEM.
KEYWORDS: Human cancer, monocytes, TEM, TIE-2-expressing monocytes, tumor microenvironment
Introduction
Tumor-associated monocytes or macrophages (TAM) are highly heterogeneous and plastic cells displaying pro-tumoral activities, largely shaped by tumor microenvironmental cues. TAM are well known to promote tumor vascularization, 1-3 metastasis,4,5 and immune suppression in pre-clinical models6,7 and to correlate with poor patient outcome.8 Recent trials have efficiently targeted TAM in different tumor settings, thus demonstrating the feasibility of this approach.9,10 A unique subset of monocytes, which express the Tek tyrosine kinase receptor TIE-2 (TEM: TIE-2 expressing monocytes) comprises a functionally distinct myeloid lineage of paracrine inducers of angiogenesis and tumor growth, identified in 2005 by De Palma and collaborators.11 In experimental mouse models, TEM recruited to tumors displayed a profoundly M2-skewed phenotype12,13 and accounted for most of the angiogenic activity of bone marrow-derived cells since the genetic ablation of TIE-2 on TEM completely prevented tumor angiogenesis.11 TIE-2 binds to Angiopoietins 1–4 (ANG 1–4) and is critically involved in vascular embryogenesis and adult angiogenesis.14 Although TIE-2 has been long regarded as an endothelial cell-specific receptor, TEM, endothelial progenitor cells and pericyte precursors of mesenchymal origin express also TIE-2.11 In vitro, TEM migrated toward angiopoietin-2 released by activated endothelial cells (EC) and angiogenic vessels, suggesting a homing mechanism for TEM to tumors.15 In 2007, Venneri et al. detected TEM in human tumors, including those of the kidney, colon, pancreas and lung as well as soft tissue carcinomas, while they were excluded from surrounding healthy tissues.15 We made similar observation in human breast cancer (BC) and reported that TEM are also highly pro-angiogenic16 and pro-lymphangiogenic cells.17 Further, TEM display an immune suppressive activity evidenced by their ability to secrete IL-10 and VEGF in large amounts and to dampen in vitro tumor-specific T-cell proliferation mediated by tumor dendritic cells (DC).18 In line with the previous studies,19,20 we observed in sections of BC that TEM density inversely correlated with the expression of HLA-DR on DC, thus limiting their ability to present antigens. In addition, TEM, but not DC, massively form multiple tight conjugates with Tregs in BC, and induce in vitro a conversion of CD4+ T cells into CD25+CD127lowFoxP3+ suppressive CD4+ T cells. This conversion was largely due to the overexpression of CD86 on TEM. Last, the observation in mice of tumor microenvironment of metastases (TMEM) and in BC patients of some parallel structures, where the presence of monocytes together with other cells seem to be involved in the metastatic process, might set the bases for a contribution of TEM toward human tumor cell dissemination.4,21,22
In 2007, Venneri et al. reported also that TEM accounted for 2–7% of blood mononuclear cells in healthy donors and were distinct from rare circulating endothelial cells and progenitors. Further, like their murine counterparts, circulating TEM displayed a marked tumor proangiogenic activity when co-injected with human A87 gliomas in immuno-compromised mice.15 It appears clear that TEM start to emerge as central player in the tumor microenvironment and tumor progression and dissemination. We review here the pro-tumoral functions of TEM in human cancers and the corresponding clinical strategies targeting them.
Origins of TEM
In 2012, Wong et al. conducted a whole genome analysis of monocytes circulating in healthy donors, which they categorized in three groups based on their expression of CD14 and CD16: non-classical (CD14+/CD16++), intermediate (CD14++/CD16+) and classical (CD14++/CD16−) monocytes.23 A recent genomic analysis confirmed the expression of Tie-2, Endoglin and VEGF-R2 in the CD14++/CD16+ population, and showed that it has the highest capacity to induce CD4+ T-cell activation.18,24 Circulating TEM are enriched but not restricted to the intermediate subset, which expresses higher levels of surface molecules involved in antigen-presenting cell–T cell interactions: particularly MHC class II, CD40 and CD74, CD40 and CD54.23
Although monocytes are commonly described as TAM precursors, it is not yet clear which of the two main circulating monocytes populations, “Ly6C+ inflammatory” or “Ly6C− resident” monocytes, is the main source of TAM in mice.25,26 Movahedi and colleagues labeled Ly6Chi and Ly6Clo monocytes with latex beads and injected them into tumor-bearing mice. The authors found labeled-TAM only in mice transplanted with labeled Ly6Chi monocytes and postulated that inflammatory but not resident monocytes contain TAM precursors.26 In contrast, in tumor-bearing mice, depletion of tissue resident macrophages with CSFR-1-specific antibodies (CSFR-1, colony stimulating factor receptor 1) was balanced by an increase in inflammatory monocytes and associated with depletion of TAM from the tumor mass but not around the invasion front.25 A comparative gene expression profiling of murine tumor TEM with that of TAM, myeloid-derived suppressor cells, inflammatory and resident monocytes, and tumor-derived EC revealed that TEM and TAM profiles were highly related and clearly distinct from that of EC. Circulating TEM express a gene signature more closely related to resident monocytes and TIE-2+ embryonic/fetal macrophages. Relative to TAM, TEM signature show enhanced pro-angiogenic/tissue remodeling activity and lower pro-inflammatory activity. Hence, resident monocytes and TEMs on one hand, and inflammatory monocytes and TAM appear to be committed to distinct functions in the tumor microenvironment. The authors postulate that TIE-2+ embryonic/fetal macrophages, resident blood monocytes, and tumor TEMs represent distinct developmental stages of a TEM lineage committed to execute physiologic proangiogenic and tissue-remodeling programs, which can be coopted by tumors.12 However, additional studies are needed to understand the relationships between human TEM and TAM.
Other authors also hypothesize a common origin with endothelial precursor cells (EPC): hematopoietic and circulating EPC originate from the bone marrow (BM) and have the capacity to differentiate into EC in vivo and to participate to the formation of new blood and lymphatic vessels.27 The contribution of EPC to the vasculature of mouse tumors was first underlined by Lyden and colleagues, who reported that the transplantation of wild type BM stem cells into angiogenic defective mice restores tumor growth and angiogenesis.28 Blood and lymphatic human EPC derive from CD34+ hematopoietic progenitors29,30 and share endothelial specific markers with mature ECs, such as TIE-1, TIE-2, VEGFR-2, VEGFR-3 or vascular endothelial (VE)-cadherin.31 In several human cancers, EPC have been detected at higher frequency in the peripheral blood of patients relative to healthy individuals30,32,33 and associated with poor patient survival, cancer dissemination and progression.34 Further, EPC are home to sites of neovascularization and have been found inserted, in very low numbers, into the tumor endothelium in several human cancers.35-37 However, to date, evidence for the contribution of EPC to human tumor neovascularization remains scarce: Capillary forming CD133+ structures were detected in NSCLC,33 while circulating EPC in multiple myeloma displayed a capillary-like network formation in matrigel.38 Recently, we reported that CD34+CD133+ EPC possess the ability to differentiate in vitro into myeloid lymphatic endothelial cells (LEC) expressing VEGF-A, -C and -D that consistently display hemangiogenic and lymphangiogenic activities.39 Finally and most importantly, myeloid LEC differentiated in vitro in the presence of patient plasma recapitulated both the phenotype and the dual angiogenic functions of TEM infiltrating breast tumors.17 Hence, the lineage relationships of EPC with angiogenic monocytes or TEM, as well as the extent of their contribution to neovascularization of human cancers remain to be clarified, but a possible link between the two lineages might not be completely excluded.
TEM involvement in different human solid tumors
Even though the first evidence of a pro-tumoral effect of TEM in mice11 and the first report of TEM in peripheral blood of cancer patient dates back to 2007,40,15 only in the recent years the complexity of the interactions between TEM and the tumor microenvironment has been examined by several groups into different human cancer settings, among which BC is the most studied. While TEM have been reported in many cancer types and frequently associated to a worse prognosis, they remain less well characterized in other cancer types. Here, we review and summarize the state of the art in this field (Table 1).
Table 1.
Summary of TEM detection and importance in tumor development in different cancer types
| Cancer type | Tumor TEM/circulating TEM | TEM role and association with tumor features | Ref. |
|---|---|---|---|
| BC | TEM: > 95% of TAM | Tumor TEM immune suppressive and pro-angiogenic activity are both controlled by TIE-2 and VEGFR pathways. TEM limit DC's Ag presentation and promote CD4+ T-cell conversion into Tregs | 16,17,18,43 |
| BC (metastatic) | Circulating TEM | Elevated levels of circulating TEM in metastatic BC | 41 |
| RCC | TEM: around 62% of TAM | Tumor TEM frequency correlates with tumor grade, patient stage, metastases and microvessel density | 45 |
| HCC | TEM: most of TAM | Tumor TEM correlate with circulating TEM and are envisioned as a diagnostic marker for HCC. TEM frequency correlates with microvessel density | 46,47 |
| Hilar cholangiocarcinoma | TEM infiltration correlates with prolonged survival | 48 | |
| CRC | TEM: minority of TAM | Low frequency of circulating TEM. No correlation with tumor, microvessel density, stage, pathological and clinical end points | 49,50 |
| Glioblastoma | TEM are mostly at the tumor periphery | TEM are associated with an invasive glioma phenotype and are a proposed biomarker of resistance to anti-angiogenic treatment | 52,53 |
| GEP-NE | Circulating TEM | 54 | |
| AML | Circulating TEM | TEM are associated with increased proliferative activity of blasts and pro-angiogenic features | 56 |
| CLL | Circulating TEM | High frequency of TEM correlates with ANG-2 secreted by CLL cells | 58 |
| Primary myelofibrosis | CD14brightCD16lowTie-2+ | TEM frequency is higher than in healthy donors | 59 |
Breast cancer
Relative to healthy women, metastatic BC patients have elevated frequencies of TEM in their peripheral blood that remained unchanged by chemotherapy (paclitaxel) or anti-angiogenic (bevacizumab) therapy.41 We reported that in the breast tumor microenvironment, most (> 95%) of monocytes (CD11b+, CD14+ CD45+ cells) express high levels of TIE-2, but also of VEGFR-1, which correlated with TIE-2 expression, and we confirmed that they are endowed with high pro-angiogenic and pro-tumoral activities.17 These TEM functions and phenotypes were shaped by the tumor microenvironment as they decreased dramatically from tumor to adjacent non-neoplastic tissue and to peripheral blood.17,16
It is well known that BC is a deeply immunosuppressed environment, characterized by an impaired APC presentation capacity, due to lack of CD80 and CD86 expression,19 and by an increased infiltration of Tregs.20 We recently showed that TEM have immunosuppressive functions, which are mediated by TIE-2 and VEGFR.18 By combining Boolean modeling and experimental approaches,42 we have predicted in silico combinations of treatments transitioning the highly pro-angiogenic phenotype of BC TEM to the weak pro-angiogenic phenotype of their peripheral blood counterparts and vice versa. In silico predicted treatments were validated experimentally using patient TEM. We have uncovered that complex cross-talks between TIE-2, VEGFR1, TNF-α and TGF-β pathways control TEM angiogenic activity.16 Importantly, we show that both immune suppressive effects and pro-angiogenic activity mediated by TEM are controlled by the synergistic action of TIE-2 and VEGFR kinase activity and could be abolished by combined use of specific kinase inhibitors of these receptors.18,16 Forget et al. show that macrophage colony-stimulating factor (CSF1) enhanced TEM pro-angiogenic activity by inducing TIE-2 expression and lowered the threshold of Ang-2 signaling needed to induce TEM migration toward breast tumor cells in vitro.43 Consistent with the observations of Forget et al., exposure of TEM to PlGF/TIE-2 kinase inhibitor/TGF-β combined treatment impaired TEM pro-angiogenic activity and ability to migrate toward tumors cells.16 Hence, the tumor microenvironment attracts TEM in tumor areas of intense neo-vascularization17 associated with high levels of Ang-2 expression.44 Ang-2 synergizes with PlGF and TNF-α to trigger TEM pro-angiogenic activity, thus tilting the balance toward a strongly immunosuppressive environment. Finally, the unraveled synergies between TIE-2 and VEGFR-1 pathways and their ligands Ang-2 and PlGF, which underlie TEM pro-angiogenic and suppressive activities, have a strong predictive value for BC patient relapse-free survival.16
Other solid tumor types
Renal cell carcinoma (RCC)
Like in BC, TEM infiltrating RCC represent a large fraction (62.12 ± 17.4%) of CD45+CD14+ monocytes and TIE-2 expression is largely induced in the TME relative to adjacent non-tumor tissue. In RCC, the frequency of tumor-infiltrated TEM correlates with tumor grade, patient stage, lymph node and distant metastases. Further, the overall tumor microvessel density correlates with TEM frequency.45
Hepatocellular carcinoma (HCC)
Similarly to RCC, in HCC tissue, most of CD14+ cells are TEM. Further, the frequency of TEM was significantly higher in HCC patients relative to healthy individuals and associated to cancer and not to hepatitis C virus infection.46,47 In addition, the frequency of TEM in peripheral blood correlates with that in the tumor and with microvessel density and changes with the therapeutic response or recurrence, thus establishing TEM as a diagnostic marker for HCC and suggesting a role for TEM in HCC-induced angiogenesis.46,47 In contrast to TEM infiltrating BC,17 HCC TEM were found enriched in perivascular areas of HCC tissues. Last, and opposite to these observations, one study described that infiltration of TEM in hilar cholangiocarcinoma (a cancer arising from the proximal biliary tree) defines a subgroup of patients with beneficial tumor characteristics and prolonged survival.48
Colorectal cancer (CRC)
In marked contrast to HCC, BC and RCC, TEM represent a minority of CD14+ cells in sections of CRC tissues.49 TEM frequency in the peripheral blood of CRC patients and healthy individuals did not differ significantly49,50 but the levels of TIE-2 expression at the surface of CD14+ cells were significantly higher in patients relative to healthy individuals.49 More recently, the low levels of circulating TEM in CRC were confirmed, even after the addition of anti-VEGF to standard chemotherapy.51 Finally, in CRC, TEM frequencies and levels of TIE-2 expression did not correlate with tumor microvessel density, tumor stage, or pathological and clinical end points in CRC patients.49
Glioblastoma
In 2014, the group of Gomez-Manzano was the first to describe the presence of TEM in surgical samples of human malignant gliomas after bevacizumab treatment and in preclinical xenograft models of glioma.52 TEM are localized at the tumor periphery in sections of human brain tissues and observed at the leading edge of the tumor following anti-VEGF therapy. The authors demonstrated that tumor TEM are associated with the development of an invasive glioma phenotype. Two years later, Gabrusiewicz et al. proposed TEM as a biomarker of tumor resistance to classical anti-angiogenic therapies in glioblastoma52,53 but, however, did not examine the presence of TEM in patient peripheral blood.
Circulating TEM in the peripheral blood of patients with solid tumors
Another open question resides on the presence of circulating TEM, which in some cases can be correlated with an advanced or metastatic status of tumors.
Peripheral blood TEM have been detected and reported in gastroenteropancreatic-neuroendocrine (GEP-NE) cancer patients, although they have not yet been characterized. In GEP-NE tumors, besides an increased level of soluble TIE-2 in the serum of metastatic patients, circulating TEM show an enhanced expression of TIE-2 and can migrate in response to Ang-2.54
Further, circulating leukocyte populations not strictly assigned to monocytes were reported as expressing TIE-2. The first one represents up to 30% of circulating cells in ovarian cancer patients and consists of a population of vascular leukocytes cells (VLC) characterized by the expression of CD14, CD45, CD11c, CD11b, VE-cadherin, CD31 and CD146 (hence their names, due to the expression of myeloid and vascular markers).55 VLC display a pro-angiogenic phenotype, secrete TGF-β, VEGF, IL-8, pro-angiogenic molecules and contribute to tumor angiogenesis in mouse models. In human ovarian cancer, most of VLCs are TEM, but only 50% of them express VE-cadherin. Interestingly, both VLC and TEM express CD52 (a glycosylphosphatidylinositol-anchored protein showing abundant surface expression in lymphoid neoplasia), which represents a promising target for adoptive immunotherapy of this neoplasm.55
TEM in hematological tumors
In a few hematological tumors, the presence of TEM is just merely described, but their importance is increasing. Noteworthy, a remarkable percentage of acute myeloid leukemia (AML) patients have circulating TEM.56 TIE-2 expression in AML is associated with increased proliferative activity of leukemic blasts, potentially through an Ang-1 autocrine loop. Last, these cells also express VEGFRs, show potential pro-angiogenic features, 56 and exhibit high FMS-related tyrosine kinase 3, a possible therapeutic intervention point of the corresponding tyrosine kinase inhibitors.57 Further, in chronic lymphocytic leukemia (CLL) patients, TEM counts were found high and correlated with plasma ANG-2 secreted by CLL cells.58 Finally, in primary myelofibrosis, a myeloproliferative neoplasm, the frequency of CD14bright CD16low TIE-2+ cells—but not of CD14lowCD16bright TIE-2+ cells—was significantly higher than in healthy individuals or patients with polycythemia vera or thrombocytemia.59
TEM in metastasis
Multiple evidence have suggested that TAM are critically involved into the dissemination of several human solid tumors by promoting tumor lymphangiogenesis and lymphatic remodeling, which are associated with lymph node metastasis and poor patient outcome.60-67 The role of TEM in metastasis is largely unknown but multiple recent observations have depicted a possible relationship between TEM and circulating tumor cells or TEM and tumor cell intravasation through the tumor endothelium.
Cancer associated macrophage-like cells (CAMCL) expressing variable levels of TIE-2, CD14 and CD45 were specifically detected in the peripheral blood of breast, pancreatic and prostate cancer patients, but not found in healthy individuals.68 Although the relationship between CAMLC and TEM remain yet largely unexplored, CAMLC may encompass a population of TEM. CAMLC in peripheral blood were found bound to circulating tumor cells (CTC), suggesting that CAMLC may contribute to tumor cells dissemination by binding to CTC while in circulation or by interacting with tumor cells at the tumor site and migrating together into circulation.68 Moreover, the involvement of TEM in tumor cell dissemination might be strengthened by the observation in mice of TMEM. TMEM are sites of tumor cell intravasation, consisting of direct contact between a macrophage, an endothelial cell and a tumor cell. Moreover, TMEM density correlates with metastasis in different mouse models.4,21,22 Preliminary data have indicated that human macrophages in TMEM may express TIE-2.4 and further work is required to establish whether mouse and human TMEM have parallel functions and share similar mechanisms of tumor cell intravasation. Along these lines, we show that a fraction of TEM were found inserted into tumor lymphatics, but not with lymphatics of adjacent non-neoplastic tissue, where TEM show drastically reduced expression of TIE-2 and LEC markers.17 Interestingly, we observed that all BC patients with metastasis to the lymph node (LN) show TEM inserted into their tumor lymphatics, whereas this was only the case for 57% of the patients without LN metastasis. 17 Thus, given the lymphangiogenic activity of TEM and the correlation between tumor lymphatics with sentinel and non-sentinel LN metastasis in BC,69,70 it is reasonable to suggest that TEM insertion into lymphatics may contribute to the spreading of tumor cells to the proximal LN. The underlying mechanisms of TEM pro-metastatic activity and insertion into tumor LV are currently under investigation in our group.
Therapeutic opportunities
Strategies targeting TEM
Preclinical and clinical strategies to target TAM have been recently reviewed71,72 and fall into four main groups: (i) killing-induced depletion of TAM either by drugs, immunotoxin-conjugated antibodies or by immune cells, (ii) prevention of recruitment of macrophages to tumor through the blockade of chemokine gradients such as SDF1 (stromal cell-derived factor 1) or CCL2, (iii) reprogramming pro-tumoral M2-like macrophages toward an antitumor (M1-like) phenotype, and (iv) reprogramming of the immunosuppressive TME to enhance tumor-specific T-cell response, e.g., through the use of CD40 agonistic antibodies or checkpoint blockade inhibitors.71,72 Most of the clinical strategies currently in progress are focused on CSFR-1 inhibition and are mainly phase I or II clinical trials. The rationale for using CSFR-1 inhibitors is largely based on the depletion or/and reprogramming of macrophages depending on the TME.71,72
TEM pro-angiogenic activity and recruitment to tumor is critically controlled by TIE-2-ANG axis, which has been the target of therapeutic strategies in clinical trials summarized below and displayed in red in Fig. 1. TIE-2-ANG axis is involved in both physiologic and tumor angiogenesis, but since the physiologic vasculature is quiescent, disruption of TIE-2-ANG axis is remarkable for the lack of side effects typically associated with inhibitors of angiogenesis (hemorrhage, arterial thromboembolic events or proteinuria). However, antiangiogenic therapy induces intratumoral hypoxia, which directly modifies TAM and other stromal cells, which in turn become active promoters of the metastatic dissemination of tumor cells. A promising molecule is AMG 386, a peptibody that prevents the binding of ANG to TIE-2: Its efficacy has been demonstrated in many tumors alone or in combination with chemotherapy, and in particular in ovarian cancer (Fig. 1, intervention 1).71,73 Similarly, CVX-060 is a fusion protein composed of two ANG-2-binding peptides. Aside from the TIE-2-ANG axis, also the VEGF-VEGFR one (Fig. 1, intervention 2) alone or in combination is a critical target for current therapeutic approaches: CVX-241 is a bispecific Ab-binding VEGF an Ang-2, whose efficacy was evaluated in a phase-II clinical trial either alone or in combination with Axitinib (a kinase inhibitor of VEGFR-1, VEGFR-2, VEGFR-3, PDGFRα/β and c-Kit), which discontinued due to lack of tolerability in patients with previously treated metastatic renal cell carcinoma.74 More recently, Nesvacumab (REGN910), an ANG-2 function blocking Ab (which does not bind to ANG-1), was evaluated in patients with different solid tumors (HCC, BC, adrenocortical carcinoma, CRC, RCC, NSCLC and pancreatic cancer) and showed acceptable safety and evidence of objective treatment effects in one adrenocortical carcinoma patient and a few hepatocellular carcinoma patients (Fig. 1, intervention 1).74 Nesvacumab is now entering a phase-II clinical trial and a phase-Ib study in combination with Aflibercept (a fusion protein composed of VEGFR binding portion fused to a Fc fragment) in patients with advanced solid malignancies. CEP-11981 potently and specifically inhibits VEGFR-1 and VEGFR-2 and its efficiency was evaluated in patients with advanced, relapsed or refractory solid tumors in a phase-I study. Although no patient experienced complete or partial response, 44% of the patients had stable disease at ≥ 6 weeks, which occurred more frequently in the higher dose cohorts.75
Figure 1.
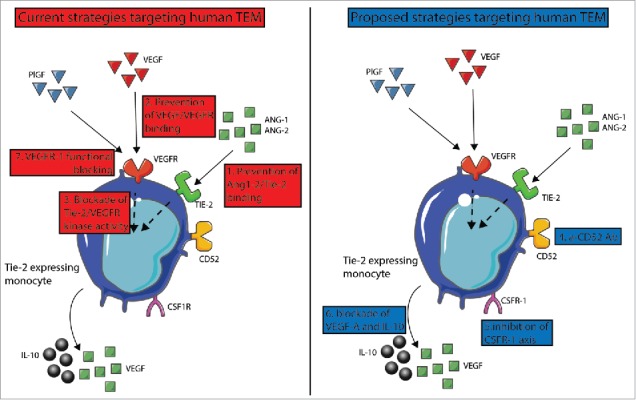
Therapeutic approaches targeting human TEM. On the left in red, strategies reported in clinical trials, while proposed strategies are shown on the right in blue. Strategies aim at preventing the interaction of (1) TIE-2/ANG2 and/or (2) VEGF/VEGFR. (3) In BC, combined blockade of TIE-2 and VEGFR kinase activity induce reprogramming of TEM toward an antitumoral functional phenotype, whose effects are summarized in Figure 2. Other approaches are based on (4) CD52-mediated TEM killing, (5) co-inhibition of CSFR-1 and VEGFR axes, (6) combined IL-10 and VEGF-A blockades. (7) VEGFR-1 function blocking antibodies impair the differentiation of CD34+ precursor cells into angiogenic TEM.
Re-programming of pro-tumoral TAM toward monocytes able to promote antitumor-specific responses has emerged recently as a powerful therapeutic strategy,71 which may apply also to TEM (Fig. 1, intervention 3). We show that TIE-2 and VEGFR-1 kinase activity synergistically control TEM angiogenic16 and immune suppressive25 activities: The combined blockade of these receptors with specific kinase inhibitors reprogrammed tumor TEM into monocytes sharing feature of myeloid-derived DC, which in turn can present tumor antigens and enhance tumor-specific T-cell responses. This treatment also dampened tumor TEM angiogenic16 and lymphangiogenic16 activities, by decreasing TEM paracrine secretion of VEGFs.16 Most importantly, this combined kinase inhibitor treatment abolished TEM immune suppressive activity by impairing the aptitude of TEM to massively convert T cells into Tregs and to secrete the immunosuppressive cytokines VEGF-A and IL-10.16 Finally, this treatment prevented also tumor TEM to interfere with DC maturation.16 All these effects are summarized in Fig. 2.
Figure 2.

Effects of combined blockade of TIE-2 and VEGFR kinase activities on BC TEM. This treatment induces reprogramming of BC TEM toward an antitumoral functional phenotype enhancing tumor-specific T cell responses, dampens TEM pro-angiogenic and lymphangiogenic activities by decreasing their paracrine secretion of VEGFs, impairs TEM-mediated conversion of T cells into Tregs, impedes TEM paracrine secretion of the immunosuppressive cytokines IL-10 and VEGF-A and prevents TEM to interfere with DC maturation.
Besides the therapeutic strategies targeting TIE-2-ANG axis, we propose below a rationale for additional approaches targeting TEM (displayed in blue in Fig. 1). In ovarian cancer, most of TEM co-express the leukocyte marker CD52, which is the target of Alemtuzumab, a monoclonal antibody approved by the FDA for the treatment of chronic lymphoid leukemia (CLL).55 Alemtuzumab depletes circulating T and B cells through antibody-dependent cellular cytolysis and complement-mediated lysis, and thus might be useful to deplete TEM in ovarian tumors (Fig. 1, intervention 4). The inhibition of CSFR-1 in clinical trials resulted in effective TAM depletion and objective clinical responses in > 50% of the patients. 71 Recently, Forget et al. observed that CSF-1 increased TIE-2 surface expression on human TEM,43 while in PyMT tumor bearing mice TIE-2 upregulation leads to an increased secretion of VEGF-A and to TEM expansion. 76 Hence, we postulate that co-inhibition of CSFR-1 and VEGFR axes may synergistically impair human TEM recruitment and pro-tumoral activities (Fig. 1, intervention 5). Along these lines, Priceman et al. demonstrated that the combined inhibition of CSF-1R and VEGF-R2 signaling pathways in Lewis lung carcinoma model prevented the recruitment of TAM and the tumor from evading anti-angiogenic therapies.77 Finally, targeting TEM secretion of IL-10 and VEGF by use of specific function antibodies (Fig. 1, intervention 6) might be synergic when combined with the interventions described above.
Strategies reprograming TEM precursors
Importantly, human and mouse circulating TEM are already committed to an angiogenic function11,15, and thus re-programming or altering mobilization or progenitor cells might result in improved disease outcome. We report that plasma from BC patients induced differentiation of CD34+ cord blood progenitors into hemangiogenic and lymphangiogenic CD11b+ myeloid cells, which express TIE-2, whereas plasma from healthy women did not have this effect.15 Consistently, we observed that circulating CD11b+ cells from BC patients, but not from healthy women, displayed a similar dual angiogenic activity.15 Further, the commitment in vitro of human CD34+ hematopoietic progenitors into CD11b+ proangiogenic cells was shown to involve PlGF and to be impaired by VEGFR-1 function blocking antibodies (Fig. 1, intervention 7). 78 In line with these results, silenced PlGF expression in the murine mammary carcinoma-derived cell line 4T1 impaired in vivo the commitment of myeloid precursors into CD11b+ angiogenic cells and dampened tumor growth and vascularization.78 Preliminary results obtained recently in our group suggest that in human BC, the signals and pathways that control the differentiation of TEM precursors into lymphangiogenic TEM and those that support tumor TEM lymphangiogenic activity differ. Hence, these results raise the critical question of whether combining therapeutic strategies targeting both TEM precursor differentiation and tumor TEM pro-tumoral activities would improve patient relapse-free survival and limit resistance to TEM-targeted therapies. To address these issues, pre-clinical models of TEM truly reflecting the functions and phenotypes of patient TEM are required. In human BC, we report that more than 95% of tumor TEM co-expressed TIE-2 and VEGFR-1 in a co-regulated manner17 and synergistically control TEM proangiogenic,16 immune suppressive,18 and lymphangiogenic17 activities. By contrast, we observed that in 4T1, FARN168, and MMTV-PyMT, currently held as the closest model to human BC model, a much lower fraction of TEM co-express TIE-2 and VEGFR-1 (28%, 45% and 5% of monocytes, respectively) and this co-expression was not co-regulated. This observation questions the validity of these pre-clinical models to the identification of treatments that can translate into effective therapeutic strategies in BC patients.
Concluding remarks
TEM have now been detected in multiple solid and hematological human tumors and represent the main population of TAM in BC, RCC and HCC. In most of cancers, TEM have been reported to display an inherent vascular growth-promoting activity, which is largely shaped by the TME or to be associated with tumor vascularization. The tumor signals and the corresponding TEM pathways that control TIE-2 expression, TEM frequency and TEM angiogenic activity remain still poorly understood. Nevertheless, ANG-2, PlGF and SDF-1 have emerged as tumor factors critically controlling TEM pro-angiogenic activity, and in parallel TIE-2 and VEGFR signaling axes arose as an attractive therapeutic target. Most importantly, besides their pro-angiogenic activity, TEM infiltrating BC endow a lymphangiogenic, immune suppressive and pro-metastatic activity. TEM also represent a circulating reservoir of cells committed to a pro-angiogenic function, which in some cancers, but not others, function as a diagnostic marker. Thus, TEM represent attractive therapeutic and diagnostic targets, but further studies are needed to elucidate their pro-tumoral functions and interactions with the TME.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
This work was supported by grants from the Swiss National Foundation (M.-A. D. project 310030-120473) and the Medic foundation (M.-A. D.).
References
- 1.Du R, Lu KV, Petritsch C, Liu P, Ganss R, Passegue E, Song H, Vandenberg S, Johnson RS, Werb Z et al.. HIF1alpha induces the recruitment of bone marrow-derived vascular modulatory cells to regulate tumor angiogenesis and invasion. Cancer Cell 2008; 13:206-20; PMID:18328425; http://dx.doi.org/ 10.1016/j.ccr.2008.01.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lin EY, Li JF, Gnatovskiy L, Deng Y, Zhu L, Grzesik DA, Qian H, Xue XN, Pollard JW. Macrophages regulate the angiogenic switch in a mouse model of breast cancer. Cancer Res 2006; 66:11238-46; PMID:17114237; http://dx.doi.org/ 10.1158/0008-5472.CAN-06-1278 [DOI] [PubMed] [Google Scholar]
- 3.Lin P, Polverini P, Dewhirst M, Shan S, Rao PS, Peters K. Inhibition of tumor angiogenesis using a soluble receptor establishes a role for Tie2 in pathologic vascular growth. J Clin Invest 1997; 100:2072-8; PMID:9329972; http://dx.doi.org/ 10.1172/JCI119740 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Harney AS, Arwert EN, Entenberg D, Wang Y, Guo P, Qian BZ, Oktay MH, Pollard JW, Jones JG, Condeelis JS. Real-time imaging reveals local, transient vascular permeability, and tumor cell intravasation stimulated by TIE2hi macrophage-derived VEGFA. Cancer Discov 2015; 5:932-43; PMID:26269515; http://dx.doi.org/ 10.1158/2159-8290.CD-15-0012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Smith HA, Kang Y. The metastasis-promoting roles of tumor-associated immune cells. J Mol Med 2013; 91:411-29; PMID:23515621; http://dx.doi.org/ 10.1007/s00109-013-1021-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Costa NL, Valadares MC, Souza PP, Mendonca EF, Oliveira JC, Silva TA, Batista AC. Tumor-associated macrophages and the profile of inflammatory cytokines in oral squamous cell carcinoma. Oral Oncol 2013; 49:216-23; PMID:23089461; http://dx.doi.org/ 10.1016/j.oraloncology.2012.09.012 [DOI] [PubMed] [Google Scholar]
- 7.Han Q, Shi H, Liu F. CD163(+) M2-type tumor-associated macrophage support the suppression of tumor-infiltrating T cells in osteosarcoma. Int Immunopharmacol 2016; 34:101-6; PMID:26938675; http://dx.doi.org/ 10.1016/j.intimp.2016.01.023 [DOI] [PubMed] [Google Scholar]
- 8.Wang R, Zhang J, Chen S, Lu M, Luo X, Yao S, Liu S, Qin Y, Chen H. Tumor-associated macrophages provide a suitable microenvironment for non-small lung cancer invasion and progression. Lung Cancer 2011; 74:188-96; PMID:21601305; http://dx.doi.org/ 10.1016/j.lungcan.2011.04.009 [DOI] [PubMed] [Google Scholar]
- 9.Ries CH, Cannarile MA, Hoves S, Benz J, Wartha K, Runza V, Rey-Giraud F, Pradel LP, Feuerhake F, Klaman I et al.. Targeting tumor-associated macrophages with anti-CSF-1R antibody reveals a strategy for cancer therapy. Cancer Cell 2014; 25:846-59; PMID:24898549; http://dx.doi.org/ 10.1016/j.ccr.2014.05.016 [DOI] [PubMed] [Google Scholar]
- 10.Tap WD, Wainberg ZA, Anthony SP, Ibrahim PN, Zhang C, Healey JH, Chmielowski B, Staddon AP, Cohn AL, Shapiro GI et al.. Structure-Guided Blockade of CSF1R Kinase in Tenosynovial Giant-Cell Tumor. N Eng J Med 2015; 373:428-37; PMID:26222558; http://dx.doi.org/ 10.1056/NEJMoa1411366 [DOI] [PubMed] [Google Scholar]
- 11.De Palma M, Venneri MA, Galli R, Sergi L, Politi LS, Sampaolesi M, Naldini L. Tie2 identifies a hematopoietic lineage of proangiogenic monocytes required for tumor vessel formation and a mesenchymal population of pericyte progenitors. Cancer Cell 2005; 8:211-26; PMID:16169466; http://dx.doi.org/ 10.1016/j.ccr.2005.08.002 [DOI] [PubMed] [Google Scholar]
- 12.Pucci F, Venneri MA, Biziato D, Nonis A, Moi D, Sica A, Di Serio C, Naldini L, De Palma M. A distinguishing gene signature shared by tumor-infiltrating Tie2-expressing monocytes, blood “resident” monocytes, and embryonic macrophages suggests common functions and developmental relationships. Blood 2009; 114:901-14; PMID:19383967; http://dx.doi.org/ 10.1182/blood-2009-01-200931 [DOI] [PubMed] [Google Scholar]
- 13.Coffelt SB, Tal AO, Scholz A, De Palma M, Patel S, Urbich C, Biswas SK, Murdoch C, Plate KH, Reiss Y et al.. Angiopoietin-2 regulates gene expression in TIE2-expressing monocytes and augments their inherent proangiogenic functions. Cancer Res 2010; 70:5270-80; PMID:20530679; http://dx.doi.org/ 10.1158/0008-5472.CAN-10-0012 [DOI] [PubMed] [Google Scholar]
- 14.Asahara T, Chen D, Takahashi T, Fujikawa K, Kearney M, Magner M, Yancopoulos GD, Isner JM. Tie2 receptor ligands, angiopoietin-1 and angiopoietin-2, modulate VEGF-induced postnatal neovascularization. Circ Res 1998; 83:233-40; PMID:9710115; http://dx.doi.org/ 10.1161/01.RES.83.3.233 [DOI] [PubMed] [Google Scholar]
- 15.Venneri MA, De Palma M, Ponzoni M, Pucci F, Scielzo C, Zonari E, Mazzieri R, Doglioni C, Naldini L. Identification of proangiogenic TIE2-expressing monocytes (TEMs) in human peripheral blood and cancer. Blood 2007; 109:5276-85; PMID:17327411; http://dx.doi.org/ 10.1182/blood-2006-10-053504 [DOI] [PubMed] [Google Scholar]
- 16.Guex N, Crespo I, Bron S, Ifticene-Treboux A, Faes-Van't Hull E, Kharoubi S, Liechti R, Werffeli P, Ibberson M, Majo F et al.. Angiogenic activity of breast cancer patients' monocytes reverted by combined use of systems modeling and experimental approaches. PLoS Comput Biol 2015; 11:e1004050; PMID:25768678; http://dx.doi.org/ 10.1371/journal.pcbi.1004050 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bron S, Henry L, Faes-Van't Hull E, Turrini R, Vanhecke D, Guex N, Ifticene-Treboux A, Marina Iancu E, Semilietof A, Rufer N et al.. TIE-2-expressing monocytes are lymphangiogenic and associate specifically with lymphatics of human breast cancer. Oncoimmunology 2016; 5:e1073882; PMID:27057438; http://dx.doi.org/ 10.1080/2162402X.2015.1073882 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ibberson M, Bron S, Guex N, Faes-van't Hull E, Ifticene-Treboux A, Henry L, Lehr HA, Delaloye JF, Coukos G, Xenarios I et al.. TIE-2 and VEGFR kinase activities drive immunosuppressive function of TIE-2-expressing monocytes in human breast tumors. Clin Cancer Res 2013; 19:3439-49; PMID:23649001; http://dx.doi.org/ 10.1158/1078-0432.CCR-12-3181 [DOI] [PubMed] [Google Scholar]
- 19.Vesosky B, Hurwitz AA. Modulation of costimulation to enhance tumor immunity. Cancer Immunol Immunother 2003; 52:663-9; PMID:12920481; http://dx.doi.org/ 10.1007/s00262-003-0424-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Facciabene A, Motz GT, Coukos G. T-regulatory cells: key players in tumor immune escape and angiogenesis. Cancer Res 2012; 72:2162-71; PMID:22549946; http://dx.doi.org/ 10.1158/0008-5472.CAN-11-3687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Robinson BD, Sica GL, Liu YF, Rohan TE, Gertler FB, Condeelis JS, Jones JG. Tumor microenvironment of metastasis in human breast carcinoma: a potential prognostic marker linked to hematogenous dissemination. Clin Cancer Res 2009; 15:2433-41; PMID:19318480; http://dx.doi.org/ 10.1158/1078-0432.CCR-08-2179 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rohan TE, Xue X, Lin HM, D'Alfonso TM, Ginter PS, Oktay MH et al.. Tumor microenvironment of metastasis and risk of distant metastasis of breast cancer. J Natl Cancer Inst 2014; 106; PMID:24895374; http://dx.doi.org/ 10.1093/jnci/dju136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wong KL, Yeap WH, Tai JJ, Ong SM, Dang TM, Wong SC. The three human monocyte subsets: implications for health and disease. Immunol Res 2012; 53:41-57; PMID:22430559; http://dx.doi.org/ 10.1007/s12026-012-8297-3 [DOI] [PubMed] [Google Scholar]
- 24.Zawada AM, Rogacev KS, Rotter B, Winter P, Marell RR, Fliser D, Heine GH. SuperSAGE evidence for CD14++CD16+ monocytes as a third monocyte subset. Blood 2011; 118:e50-61; PMID:21803849; http://dx.doi.org/ 10.1182/blood-2011-01-326827 [DOI] [PubMed] [Google Scholar]
- 25.MacDonald KP, Palmer JS, Cronau S, Seppanen E, Olver S, Raffelt NC, Kuns R, Pettit AR, Clouston A, Wainwright B et al.. An antibody against the colony-stimulating factor 1 receptor depletes the resident subset of monocytes and tissue- and tumor-associated macrophages but does not inhibit inflammation. Blood 2010; 116:3955-63; PMID:20682855; http://dx.doi.org/ 10.1182/blood-2010-02-266296 [DOI] [PubMed] [Google Scholar]
- 26.Movahedi K, Laoui D, Gysemans C, Baeten M, Stange G, Van den Bossche J, Mack M, Pipeleers D, In't Veld P, De Baetselier P et al.. Different tumor microenvironments contain functionally distinct subsets of macrophages derived from Ly6C(high) monocytes. Cancer Res 2010; 70:5728-39; PMID:20570887; http://dx.doi.org/ 10.1158/0008-5472.CAN-09-4672 [DOI] [PubMed] [Google Scholar]
- 27.Asahara T, Murohara T, Sullivan A, Silver M, van der Zee R, Li T, Witzenbichler B, Schatteman G, Isner JM. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997; 275:964-7; PMID:9020076; http://dx.doi.org/ 10.1126/science.275.5302.964 [DOI] [PubMed] [Google Scholar]
- 28.Lyden D, Hattori K, Dias S, Costa C, Blaikie P, Butros L, Chadburn A, Heissig B, Marks W, Witte L et al.. Impaired recruitment of bone-marrow-derived endothelial and hematopoietic precursor cells blocks tumor angiogenesis and growth. Nat Med 2001; 7:1194-201; PMID:11689883; http://dx.doi.org/ 10.1038/nm1101-1194 [DOI] [PubMed] [Google Scholar]
- 29.Salven P, Mustjoki S, Alitalo R, Alitalo K, Rafii S. VEGFR-3 and CD133 identify a population of CD34+ lymphatic/vascular endothelial precursor cells. Blood 2003; 101:168-72; PMID:12393704; http://dx.doi.org/ 10.1182/blood-2002-03-0755 [DOI] [PubMed] [Google Scholar]
- 30.Dome B, Hendrix MJ, Paku S, Tovari J, Timar J. Alternative vascularization mechanisms in cancer: pathology and therapeutic implications. Am J Pathol 2007; 170:1-15; PMID:17200177; http://dx.doi.org/ 10.2353/ajpath.2007.060302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Resch T, Pircher A, Kahler CM, Pratschke J, Hilbe W. Endothelial progenitor cells: current issues on characterization and challenging clinical applications. Stem Cell Rev 2012; 8:926-39; PMID:22095429; http://dx.doi.org/ 10.1007/s12015-011-9332-9 [DOI] [PubMed] [Google Scholar]
- 32.Sussman LK, Upalakalin JN, Roberts MJ, Kocher O, Benjamin LE. Blood markers for vasculogenesis increase with tumor progression in patients with breast carcinoma. Cancer Biol Ther 2003; 2:255-6; http://dx.doi.org/ 10.4161/cbt.2.3.363 [DOI] [PubMed] [Google Scholar]
- 33.Hilbe W, Dirnhofer S, Oberwasserlechner F, Schmid T, Gunsilius E, Hilbe G, Wöll E, Kähler CM. CD133 positive endothelial progenitor cells contribute to the tumour vasculature in non-small cell lung cancer. J Clin Pathol 2004; 57:965-9; PMID:15333659; http://dx.doi.org/ 10.1136/jcp.2004.016444 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pirro M, Cagini L, Mannarino MR, Andolfi M, Potenza R, Paciullo F, Bianconi V, Frangione MR, Bagaglia F, Puma F et al.. Reduced survival in patients with early-stage non-small-cell lung cancer is associated with high pleural endothelial progenitor cell levels. Eur J Cardiothorac Surg 2016; 50:1053-9; PMID:27301385; http://dx.doi.org/ 10.1093/ejcts/ezw197 [DOI] [PubMed] [Google Scholar]
- 35.Arbab AS, Janic B, Knight RA, Anderson SA, Pawelczyk E, Rad AM, Read EJ, Pandit SD, Frank JA. Detection of migration of locally implanted AC133+ stem cells by cellular magnetic resonance imaging with histological findings. FASEB J 2008; 22:3234-46; PMID:18556461; http://dx.doi.org/ 10.1096/fj.07-105676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Peters BA, Diaz LA, Polyak K, Meszler L, Romans K, Guinan EC, Antin JH, Myerson D, Hamilton SR, Vogelstein B et al.. Contribution of bone marrow-derived endothelial cells to human tumor vasculature. Nat Med 2005; 11:261-2; PMID:15723071; http://dx.doi.org/ 10.1038/nm1200 [DOI] [PubMed] [Google Scholar]
- 37.Dome B, Timar J, Dobos J, Meszaros L, Raso E, Paku S, Kenessey I, Ostoros G, Magyar M, Ladanyi A et al.. Identification and clinical significance of circulating endothelial progenitor cells in human non-small cell lung cancer. Cancer Res 2006; 66:7341-7; PMID:16849585; http://dx.doi.org/ 10.1158/0008-5472.CAN-05-4654 [DOI] [PubMed] [Google Scholar]
- 38.Zhang H, Vakil V, Braunstein M, Smith EL, Maroney J, Chen L, Dai K, Berenson JR, Hussain MM, Klueppelberg U et al.. Circulating endothelial progenitor cells in multiple myeloma: implications and significance. Blood 2005; 105:3286-94; PMID:15618473; http://dx.doi.org/ 10.1182/blood-2004-06-2101 [DOI] [PubMed] [Google Scholar]
- 39.Van't Hull EF, Bron S, Henry L, Ifticene-Treboux A, Turrini R, Coukos G, Delaloye JF, Doucey MA. Bone marrow-derived cells are implicated as a source of lymphatic endothelial progenitors in human breast cancer. Oncoimmunology 2014; 3:e29080; PMID:25101222; http://dx.doi.org/ 10.4161/onci.29080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.De Palma M, Murdoch C, Venneri MA, Naldini L, Lewis CE. Tie2-expressing monocytes: regulation of tumor angiogenesis and therapeutic implications. Trends Immunol 2007; 28:519-24; PMID:17981504; http://dx.doi.org/ 10.1016/j.it.2007.09.004 [DOI] [PubMed] [Google Scholar]
- 41.Cattin S, Fellay B, Pradervand S, Trojan A, Ruhstaller T, Ruegg C, Fürstenberger G. Bevacizumab specifically decreases elevated levels of circulating KIT+CD11b+ cells and IL-10 in metastatic breast cancer patients. Oncotarget 2016; 7:11137-50; PMID:26840567 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Doucey MA, Xenarios I. Toward a rational design of combination therapy in cancer. Oncoimmunology 2015; 4:e1046674; PMID:26451320; http://dx.doi.org/ 10.1080/2162402X.2015.1046674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Forget MA, Voorhees JL, Cole SL, Dakhlallah D, Patterson IL, Gross AC, Moldovan L, Mo X, Evans R, Marsh CB et al.. Macrophage colony-stimulating factor augments Tie2-expressing monocyte differentiation, angiogenic function, and recruitment in a mouse model of breast cancer. PLoS One 2014; 9:e98623; PMID:24892425; http://dx.doi.org/ 10.1371/journal.pone.0098623 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 44.Tsutsui S, Inoue H, Yasuda K, Suzuki K, Takeuchi H, Nishizaki T, Higashi H, Era S, Mori M. Angiopoietin 2 expression in invasive ductal carcinoma of the breast: its relationship to the VEGF expression and microvessel density. Breast Cancer Res Treat 2006; 98:261-6; PMID:16538528; http://dx.doi.org/ 10.1007/s10549-005-9157-9 [DOI] [PubMed] [Google Scholar]
- 45.Ji J, Zhang G, Sun B, Yuan H, Huang Y, Zhang J, Wei X, Zhang X, Hou J. The frequency of tumor-infiltrating Tie-2-expressing monocytes in renal cell carcinoma: its relationship to angiogenesis and progression. Urology 2013; 82:974. e9-13; PMID:23769120; http://dx.doi.org/ 10.1016/j.urology.2013.05.026 [DOI] [PubMed] [Google Scholar]
- 46.De Palma M, Coukos G, Semela D. TIE2-expressing monocytes: a novel cellular biomarker for hepatocellular carcinoma? Hepatology 2013; 57:1294-6; PMID:22911438; http://dx.doi.org/ 10.1002/hep.26025 [DOI] [PubMed] [Google Scholar]
- 47.Matsubara T, Kanto T, Kuroda S, Yoshio S, Higashitani K, Kakita N, Miyazaki M, Sakakibara M, Hiramatsu N, Kasahara A et al.. TIE2-expressing monocytes as a diagnostic marker for hepatocellular carcinoma correlates with angiogenesis. Hepatology 2013; 57:1416-25; PMID:22815256; http://dx.doi.org/ 10.1002/hep.25965 [DOI] [PubMed] [Google Scholar]
- 48.Atanasov G, Hau HM, Dietel C, Benzing C, Krenzien F, Brandl A, Englisch JP, Wiltberger G, Schierle K, Robson SC et al.. Prognostic significance of TIE2-expressing monocytes in hilar cholangiocarcinoma. J Surg Oncol 2016; 114:91-8; PMID:27111031; http://dx.doi.org/ 10.1002/jso.24249 [DOI] [PubMed] [Google Scholar]
- 49.Goede V, Coutelle O, Shimabukuro-Vornhagen A, Holtick U, Neuneier J, Koslowsky TC, Weihrauch MR, von Bergwelt-Baildon M, Hacker UT. Analysis of Tie2-expressing monocytes (TEM) in patients with colorectal cancer. Cancer Invest 2012; 30:225-30; PMID:22171993; http://dx.doi.org/ 10.3109/07357907.2011.636114 [DOI] [PubMed] [Google Scholar]
- 50.Schauer D, Starlinger P, Reiter C, Jahn N, Zajc P, Buchberger E, Bachleitner-Hofmann T, Bergmann M, Stift A, Gruenberger T et al.. Intermediate monocytes but not TIE2-expressing monocytes are a sensitive diagnostic indicator for colorectal cancer. PLoS One 2012; 7:e44450; PMID:22973451; http://dx.doi.org/ 10.1371/journal.pone.0044450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Schauer D, Starlinger P, Alidzanovic L, Zajc P, Maier T, Feldman A, Padickakudy R, Buchberger E, Elleder V, Spittler A et al.. Chemotherapy of colorectal liver metastases induces a rapid rise in intermediate blood monocytes which predicts treatment response. Oncoimmunology 2016; 5:e1160185; PMID:27471631; http://dx.doi.org/ 10.1080/2162402X.2016.1160185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Gabrusiewicz K, Liu D, Cortes-Santiago N, Hossain MB, Conrad CA, Aldape KD, Fuller GN, Marini FC, Alonso MM, Idoate MA et al.. Anti-vascular endothelial growth factor therapy-induced glioma invasion is associated with accumulation of Tie2-expressing monocytes. Oncotarget 2014; 5:2208-20; PMID:24809734; http://dx.doi.org/ 10.18632/oncotarget.1893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Cortes-Santiago N, Hossain MB, Gabrusiewicz K, Fan X, Gumin J, Marini FC, Alonso MM, Lang F, Yung WK, Fueyo J et al.. Soluble Tie2 overrides the heightened invasion induced by anti-angiogenesis therapies in gliomas. Oncotarget 2016; 7:16146-57; PMID:26910374 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Figueroa-Vega N, Diaz A, Adrados M, Alvarez-Escola C, Paniagua A, Aragones J, Martín-Pérez E, Leskela S, Moreno-Otero R, González-Amaro R et al.. The association of the angiopoietin/Tie-2 system with the development of metastasis and leukocyte migration in neuroendocrine tumors. Endocr Relat Cancer 2010; 17:897-908; PMID:20696814; http://dx.doi.org/ 10.1677/ERC-10-0020 [DOI] [PubMed] [Google Scholar]
- 55.Pulaski HL, Spahlinger G, Silva IA, McLean K, Kueck AS, Reynolds RK, Coukos G, Conejo-Garcia JR, Buckanovich RJ. Identifying alemtuzumab as an anti-myeloid cell antiangiogenic therapy for the treatment of ovarian cancer. J Transl Med 2009; 7:49; PMID:19545375; http://dx.doi.org/ 10.1186/1479-5876-7-49 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Riccioni R, Calzolari A, Biffoni M, Senese M, Riti V, Petrucci E, Pasquini L, Cedrone M, Lo-Coco F, Diverio D et al.. Podocalyxin is expressed in normal and leukemic monocytes. Blood Cells Mol Dis 2006; 37:218-25; PMID:17059890; http://dx.doi.org/ 10.1016/j.bcmd.2006.09.001 [DOI] [PubMed] [Google Scholar]
- 57.Riccioni R, Diverio D, Mariani G, Buffolino S, Riti V, Saulle E, Petrucci E, Cedrone M, Lo-Coco F, Foà R et al.. Expression of Tie-2 and other receptors for endothelial growth factors in acute myeloid leukemias is associated with monocytic features of leukemic blasts. Stem Cells 2007; 25:1862-71; PMID:17446561; http://dx.doi.org/ 10.1634/stemcells.2006-0700 [DOI] [PubMed] [Google Scholar]
- 58.Maffei R, Bulgarelli J, Fiorcari S, Bertoncelli L, Martinelli S, Guarnotta C, Castelli I, Deaglio S, Debbia G, De Biasi S et al.. The monocytic population in chronic lymphocytic leukemia shows altered composition and deregulation of genes involved in phagocytosis and inflammation. Haematologica 2013; 98:1115-23; PMID:23349302; http://dx.doi.org/ 10.3324/haematol.2012.073080 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Campanelli R, Rosti V, Fois G, Bonetti E, Barosi G, Massa M. CD14(bright)CD16(low) intermediate monocytes expressing Tie2 are increased in the peripheral blood of patients with primary myelofibrosis. Exp Hematol 2014; 42:244-6; PMID:24333662; http://dx.doi.org/ 10.1016/j.exphem.2013.12.002 [DOI] [PubMed] [Google Scholar]
- 60.Ran S, Montgomery KE. Macrophage-mediated lymphangiogenesis: the emerging role of macrophages as lymphatic endothelial progenitors. Cancers 2012; 4:618-57; PMID:22946011; http://dx.doi.org/ 10.3390/cancers4030618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Alitalo A, Detmar M. Interaction of tumor cells and lymphatic vessels in cancer progression. Oncogene 2012; 31:4499-508; PMID:22179834; http://dx.doi.org/ 10.1038/onc.2011.602 [DOI] [PubMed] [Google Scholar]
- 62.Ran S, Volk L, Hall K, Flister MJ. Lymphangiogenesis and lymphatic metastasis in breast cancer. Pathophysiology 2010; 17:229-51; http://dx.doi.org/ 10.1016/j.pathophys.2009.11.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Schoppmann SF, Birner P, Stockl J, Kalt R, Ullrich R, Caucig C, Kriehuber E, Nagy K, Alitalo K, Kerjaschki D. Tumor-associated macrophages express lymphatic endothelial growth factors and are related to peritumoral lymphangiogenesis. Am J Pathol 2002; 161:947-56; PMID:12213723; http://dx.doi.org/ 10.1016/S0002-9440(10)64255-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Storr SJ, Safuan S, Mitra A, Elliott F, Walker C, Vasko MJ, Ho B, Cook M, Mohammed RA, Patel PM et al.. Objective assessment of blood and lymphatic vessel invasion and association with macrophage infiltration in cutaneous melanoma. Mod Pathol 2012; 25:493-504; PMID:22080065; http://dx.doi.org/ 10.1038/modpathol.2011.182 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ding P, Wang W, Wang J, Yang Z, Xue L. Expression of tumor-associated macrophage in progression of human glioma. Cell Biochem Biophys 2014; 70:1625-31; PMID:25182001; http://dx.doi.org/ 10.1007/s12013-014-0105-3 [DOI] [PubMed] [Google Scholar]
- 66.Zhang W, Wang L, Zhou D, Cui Q, Zhao D, Wu Y. Expression of tumor-associated macrophages and vascular endothelial growth factor correlates with poor prognosis of peripheral T-cell lymphoma, not otherwise specified. Leuk Lymphoma 2011; 52:46-52; PMID:21077742; http://dx.doi.org/ 10.3109/10428194.2010.529204 [DOI] [PubMed] [Google Scholar]
- 67.Kurahara H, Shinchi H, Mataki Y, Maemura K, Noma H, Kubo F, Sakoda M, Ueno S, Natsugoe S, Takao S. Significance of M2-polarized tumor-associated macrophage in pancreatic cancer. J Surg Res 2011; 167:e211-9; PMID:19765725; http://dx.doi.org/ 10.1016/j.jss.2009.05.026 [DOI] [PubMed] [Google Scholar]
- 68.Adams DL, Martin SS, Alpaugh RK, Charpentier M, Tsai S, Bergan RC, Ogden IM, Catalona W, Chumsri S, Tang CM et al.. Circulating giant macrophages as a potential biomarker of solid tumors. Proc Natl Acad Sci USA 2014; 111:3514-9; PMID:24550495; http://dx.doi.org/ 10.1073/pnas.1320198111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kerjaschki D, Bago-Horvath Z, Rudas M, Sexl V, Schneckenleithner C, Wolbank S, Bartel G, Krieger S, Kalt R, Hantusch B et al.. Lipoxygenase mediates invasion of intrametastatic lymphatic vessels and propagates lymph node metastasis of human mammary carcinoma xenografts in mouse. J Clin Invest 2011; 121:2000-12; PMID:21540548; http://dx.doi.org/ 10.1172/JCI44751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Van den Eynden GG, Vandenberghe MK, van Dam PJ, Colpaert CG, van Dam P, Dirix LY, Vermeulen PB, Van Marck EA. Increased sentinel lymph node lymphangiogenesis is associated with nonsentinel axillary lymph node involvement in breast cancer patients with a positive sentinel node. Clin Cancer Res 2007; 13:5391-7; PMID:17875768; http://dx.doi.org/ 10.1158/1078-0432.CCR-07-1230 [DOI] [PubMed] [Google Scholar]
- 71.Quail DF, Joyce JA. Molecular pathways: deciphering mechanisms of resistance to macrophage-targeted therapies. Clin Cancer Res 2016; 23(4):876-84; PMID:27895033; http://dx.doi.org/ 10.1158/1078-0432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Tang X, Mo C, Wang Y, Wei D, Xiao H. Anti-tumour strategies aiming to target tumour-associated macrophages. Immunology 2013; 138:93-104; PMID:23113570; http://dx.doi.org/ 10.1111/imm.12023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Monk BJ, Poveda A, Vergote I, Raspagliesi F, Fujiwara K, Bae DS, Oaknin A, Ray-Coquard I, Provencher DM, Karlan BY et al.. Final results of a phase 3 study of trebananib plus weekly paclitaxel in recurrent ovarian cancer (TRINOVA-1): long-term survival, impact of ascites, and progression-free survival-2. Gynecol Oncol 2016; 143:27-34; PMID:27546885; http://dx.doi.org/ 10.1016/j.ygyno.2016.07.112 [DOI] [PubMed] [Google Scholar]
- 74.Pfizer PF-04856884 (CVX-060) in Combination with Axitinib in Patients with Previously Treated Metastatic Renal Cell Carcinoma. Bethesda, MD, USA: National Library of Medicine 2000–2016; Available from: https://clinicaltrialsgov/ct2/show/NCT01441414 2012 [Google Scholar]
- 75.Pili R, Carducci M, Brown P, Hurwitz H. An open-label study to determine the maximum tolerated dose of the multitargeted tyrosine kinase inhibitor CEP-11981 in patients with advanced cancer. Invest New Drugs 2014; 32:1258-68; PMID:25152243; http://dx.doi.org/ 10.1007/s10637-014-0147-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.De Palma M, Naldini L. Angiopoietin-2 TIEs up macrophages in tumor angiogenesis. Clin Cancer Res 2011; 17:5226-32; PMID:21576085; http://dx.doi.org/ 10.1158/1078-0432.CCR-10-0171 [DOI] [PubMed] [Google Scholar]
- 77.Priceman SJ, Sung JL, Shaposhnik Z, Burton JB, Torres-Collado AX, Moughon DL, Johnson M, Lusis AJ, Cohen DA, Iruela-Arispe ML et al.. Targeting distinct tumor-infiltrating myeloid cells by inhibiting CSF-1 receptor: combating tumor evasion of antiangiogenic therapy. Blood 2010; 115:1461-71; PMID:20008303; http://dx.doi.org/ 10.1182/blood-2009-08-237412 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Laurent J, Hull EF, Touvrey C, Kuonen F, Lan Q, Lorusso G, Doucey MA, Ciarloni L, Imaizumi N, Alghisi GC et al.. Proangiogenic factor PlGF programs CD11b(+) myelomonocytes in breast cancer during differentiation of their hematopoietic progenitors. Cancer Res 2011; 71:3781-91; PMID:21507936; http://dx.doi.org/ 10.1158/0008-5472.CAN-10-3684 [DOI] [PubMed] [Google Scholar]