

Abstract
The balance between free radicals and antioxidants is an important factor for maintaining health and slowing disease progression. The use of antioxidants, particularly natural antioxidants, has become an important strategy for dealing with this cause of widespread diseases. Natural antioxidants have been used as therapeutic tools against many diseases because they are safe, effective, and inexpensive and are among the most commonly used adjuvants in the treatment of several diseases. Camel whey protein (CWP) is considered a strong natural antioxidant because it decreases oxidative stress, enhances immune system function, and increases glutathione levels. The structure of CWP is very similar to that of other types of whey protein from different types of milk. CWP contains many components, such as lactoferrin (LF), lactalbumin, lactoglobulins, lactoperoxidase, and lysozyme, and is rich in immunoglobulins. However, in contrast to other WPs, CWP lacks β-lactoglobulin, the main cause of milk allergies in children. The components of CWP have many beneficial effects, including stimulation of both innate and adaptive immunity and anti-inflammatory, anticancer, antibacterial, and antiviral activities. Recently, it has been shown that CWP and its unique components can facilitate the treatment of impaired diabetic wound healing. However, the molecular mechanisms underlying the protective effects of CWP in human and other animal disorders are not fully understood. Therefore, the current review presents a concise summary of the scientific evidence of the beneficial effects of CWP to support its therapeutic use in disease treatment and nutritional intervention.
Keywords: Antioxidant, Camel whey protein, Free radicals, Health problems, Inflammation
Introduction
Natural antioxidants play central roles in enhancing immune system function via oxidative stress-dependent mechanisms. Oxidative stress appears to play significant roles in many human diseases. For example, we previously reported the beneficial effects of immunomodulatory roles of camel whey protein as natural antioxidants (1-7) in accelerating the healing process of diabetic wounds has been observed in experimental animal models. Therefore, many recent studies have focused on the importance of antioxidants in food, particularly the effects of milk and milk-derived peptides, which are consumed every day by humans of all ages, on overall health and immune system function (8). Lactoferrin, a bovine whey protein, is a potent non-
enzyme antioxidant. Camel whey protein (CWP) and bovine whey protein are similar, except that camel milk lacks β-lactoglobulin (β-LG) (9, 10). CWP exhibits greater antioxidant activities than bovine and other whey proteins because of increased content of antioxidant amino acids (11). Research also demonstrated that whey protein possesses antimicrobial and antioxidant activities (9). Researchers observed that animals fed an aflatoxin (AF)-contaminated diet supplemented with whey protein concentrate (WPC) exhibited decreased lipid peroxidation (LP) and significantly increased glutathione (GSH) levels in the liver and testis compared with animals fed an AF-contaminated diet alone. CWP can also restore GSH levels in the liver and testis (12). A study concluded that dietary
supplementation with WP reduces inflammation, oxidative stress, and blood pressure (13). Research showed a decrease in free radicals when diabetic rats were supplemented with CWP (3). Another study demonstrated that CWP suppresses oxidative stress by decreasing hepatic lipid peroxidation and increasing GSH levels by activating glutathione-S-transferase (14).
Numerous recent studies have examined these therapeutic actions, and mounting scientific evidence is emerging on the detailed constituents of CWP as well as the candidate therapeutic components to support the current therapeutic use of CWP (Figure 1). The following review will summarize current knowledge in these areas.
Figure 1.
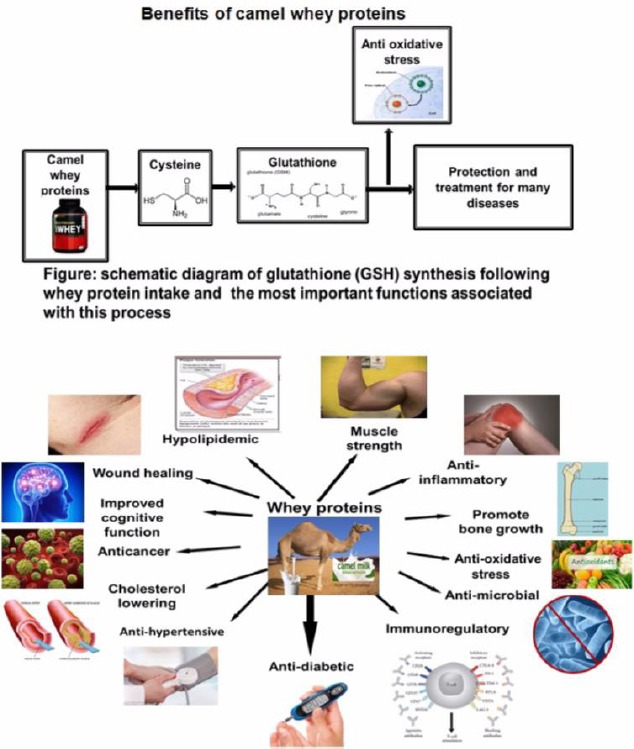
The biological benefits of camel whey protein
We summarized the biological benefits of camel whey protein for improvement of health disorders
Materials and Methods
The outline of the consensus guideline was the result of the considered opinion of the whole panel. The final review included the relevant CWP literature as searched using several electronic databases, the relative weight of the suggestions retained for the purpose of biological activities of CWP reflects the expert opinion of the panel. The consensus on these activities was expressed by the panelists using a checklist itemizing the CWP effects considered relevant after analysis of the literature. The collective judgment of the panel is expressed as a percentage of agreement among panelists. The panel decided to use a GRADE methodology for defining some treatments and biological activities of CWP.
The following electronic databases (data banks) were searched:
NCBI PubMed (1990 onwards); EMBASE (1990 onwards); UKCRN (the UK Clinical Research Network Portfolio Database); WHO ICTRP (the World Health Organization International Clinical Trials Registry Platform); ISI Web of Science; Google Scholar.
Search strategy: search covered published articles from January 1990 to August 2016. References were checked and pdf copies were provided.
Restrictions: No publication restrictions were applied. Authors were required to apply their clinical experience to compile a draft list of suitable articles for the topic, also using their experience on the immunomodulatory effect of CWP against diabetes and heat stress published from 2008 until August 2016.
Keywords used in the search
(Camel whey protein; Camel Milk; Camel milk components; Whey protein; Whey protein ingredients; Camel milk ingredients; undenatured whey protein) AND (immunomodulatory effect; Immune reaction; immune mechanism; adaptive immunity; inflammation; specific antibody; inflammatory cytokines; tumor necrosis factor alpha; chemotaxis; phagocytosis; diabetes; glucose; insulin; ROS; antioxidants; oxidative stress; wound healing; hypertension; heat stress; thermal activities; antibacterial effect; bacteriostatic; gram positive bacteria; gram-negative bacteria; viral infection; RNA viruses; DNA viruses; viral replication; carcinogenesis; colon cancer; breast cancer; hepatocytes).
Unique components of camel whey protein compared with other whey proteins
CWP represents 20-25% of total camel milk protein (15). CWP contains serum albumin, α-lactalbumin (α-LA), immunoglobulin (Ig), lacto-phorin, and peptidoglycan recognition protein (16). CWP also contains lactoperoxidase (LPO), lysozyme, and lactoferrin (LF), as well as other proteins with biological functions (17). Serum albumin is the major whey protein (WP) present in camel milk, with an average concentration of 10.8 g/l (10). WP contains essential and nonessential amino acids. CWPs are also an abundant source of glutamine and branched-chain amino acids, which are important for cell growth (18). Human and camel milk both feature high levels of α-LA and LF and a lack of β-LG (10). Camel milk is devoid of β-LG, whereas β-LG is the major WP in bovine (19), buffalo, caprine, and equine milk (20). Another study also confirmed that the main components of WPs in camel milk and colostrum are similar to those in bovine milk, except for the lack of β-LG (10). β-LG is responsible in part for allergies to bovine milk in infants. CWP has no allergenic properties and can be consumed by lactase- and/or immune-deficient populations (21). Most interestingly, the concentration of lactophorin in camel milk is approximately three-fold higher than that in bovine milk (22). LF, alone or in combination with other milk proteins, such as lysozyme and Igs, plays a vital role in innate and specific host defense mechanisms against infection by microorganisms (23-25). LF is an iron-binding glycoprotein present in various biological fluids from mammals and in neutrophils. It is involved in multiple functions, including defense against pathogens (26). Camel milk has higher concentrations of lysozyme, LF, and IgG than cow and buffalo milk. Lysozyme is also an antibacterial protein present in milk, tears, saliva, and many secretions of mammals (17) and thus plays an essential role in enhancing innate immunity. Therefore, the unique components of CWP support its therapeutic utilization in many diseases (Table 1).
Table 1.
The components of whey proteins and their biological activities The biological activities of camel whey protein components were summarized from the literature
| Components of whey protein | Biological Function | Species | References |
|---|---|---|---|
| α-lactalbumin | Enhancement of antibody response to systematic antigen stimulation and used in manufacturing of infant food | Camel, bovine, and human | (40) |
| Lactoferrin | Antimicrobial activities against microorganisms, anticancer, anti- inflammatory | Camel, bovine, and human | (90) |
| β-lactoglobulin | Source of essential and branched chain amino acid, responsible for child allergy | Bovine, buffalo, caprine, and equine | (17, 62) |
| Lysozymes | Antibacterial protein present in milk, tears, and saliva, and thus plays an important role in enhancing innate immunity | Camel and bovine | (21) |
| Immunoglobulin | Enhances immune functions | Camel, bovine, and human | (19) |
| Lactoperoxidase | Suppression of bacterial growth | Camel and bovine | (78) |
| Glycomacropeptide | Has an inhibitory effect on acid gastric secretion and modifies the concentration of blood which regulates digestive peptides | Camel and bovine | (10) |
Camel whey protein as an immunomodulatory factor that protects the immune system
Camel whey proteins are some of the very few ingredients (or dietary supplements) shown to modulate immune function using proven in vitro and in vivo models. Often the improvements have correlated with a measurable improvement in immune-meditated functions. CWP modulates different immune cell functions, such as enhancing lymphocyte activation, proliferation and chemotaxis; cytokine secretion; antibody production; phagocytic activity; and granulocyte and NK cell activity (27). WP also enhances the production of IL-1β, IL-8, IL-6, macrophage inflammatory proteins (MIP-1α, MIP-1β), and tumor necrosis factor (TNF-α) (28). CWP enhances immune cell functions during early development and plays a vital therapeutic role in some immune system disorders, including diabetes (4). CWP enhances cytoskeletal rearrangements and chemotaxis in B and T cells during diabetes, thus improving the immune response in diabetic mice (5). Another study reported that levels of GSH were increased in several GSH-deficient HIV patients following oral administration of an undenatured cysteine-rich WP isolate (29). Furthermore, WP-derived products clearly modulate immune functions in in vitro and in vivo studies (30). Whey peptides have immunomodulatory activities, such as stimulating lymphocytes and increasing phagocytosis and the secretion of IgA from Payer’s patches (31). CWP also exerts protective effects against childhood asthma (32). The anticancer (33) and immune system effects of CWP and antiapoptotic effects of CWP in diabetics have provided researchers with an opportunity to develop novel therapeutic strategies. By increasing GSH levels, CWP stimulates the proliferation of lymphocytes (34-36). Additionally, CWP increases the number of mast cells and the production of their associated cytokines and other biochemical mediators. CWP regulates the expression of TNF-α and cell death receptor (Fas) mRNAs and subsequently enhances the closure and healing of diabetic wounds (35). Individually, these fractions are established immune-enhancing constituents that modulate a range of immune functions that are linked to a range of bioactive functions such as prebiotic effects, promotion of tissue repair, maintenance of intestinal integrity, destruction of pathogens, and elimination of toxins (37). The addition of WPC to the diet is shown to significantly improve primary and secondary intestinal tract antibody responses to a variety of different vaccine antigens that are currently in medical use (38). While rodents which consumed a diet containing 20% protein from WPC, showed a significantly better immune response to influenza vaccine, diphtheria and tetanus toxoids, poliomyelitis vaccine, ovalbumin, and cholera toxins (38). Moreover, we previously showed that CWP supplementation in diabetic mice promotes tissue repair via decreasing the oxidative stress and the restoration of pro-inflammatory cytokines levels and β-defensin, which accelerate cutaneous wound healing (19).
Inflammatory cytokines stimulate many signaling cascades that lead to B cell apoptosis (39). CWP increases wound healing capacity in diabetic animals by suppressing inflammatory cytokines (14, 34). TheTable 1. The components of whey proteins and their biological activities The biological activities of camel whey protein components were summarized from the literature effects of CWP on immune functions include regulation of cytokines (1) and enhancement of leukocyte proliferation (5). The amino acid content of CWP is consistent with its immunomodulatory effects (40, 41). The bioactive components of CWP, such as LF, LPO, glycomacropeptide, serum albumin, different growth factors, and Igs, exhibit anticancer, antiviral, antibacterial, and antifungal activities (42-44). Additionally, CWP increases IL-2 and IL-8 levels but decreases IL-1α, IL-1β, IL-10, and IL- 6 levels (1).
Antidiabetic effects of camel whey protein
Diabetes mellitus (DM) is characterized by abnormally high blood glucose levels, resulting from low insulin secretion and/or increased insulin resistance. Oxidative stress is an important patho-genic factor in diabetic complications that affect cell life span. Although ROS plays crucial roles in cell signaling and in the immune response, higher levels of ROS cause oxidative stress during diabetes. CWP regulates oxidative stress and the inflamma-tory response which act as an important factor in diabetes treatment.
CWP supplementation improves the normal inflammatory process during wound healing in diabetic models by restoring oxidative stress and inflammatory cytokine levels (34). LF regulates the levels of TNF-α and IL-6, which decrease inflamma-tion and mortality (45). Whey supplementation improves wound healing by increasing GSH synthesis and improving the function of the cellular antioxidant defense system (46). CWP ameliorates impaired wound healing in diabetic patients by decreasing blood glucose levels, oxidative stress, growth factor levels, and the inflammatory response. CWP decreases wound size, and histopathological findings include increased epithelization and angio-genesis, granulation tissue formation, and extracellular matrix remodeling. CWP improves the healing of diabetic wounds by enhancing the immune response and reducing diabetic complications (3). Additionally, CWP inhibits T-helper1 (Th1)-type T cells, which play important roles in diabetic complications (35). In this context, CWP limits prolonged inflammation, improves immune system function, and enhances wound healing in diabetic mice models (2). CWP decreases blood glucose levels, levels of the free radical diphenyl-picrylhydrazyl (DPPH) and lipid peroxidation in the brain tissue of diabetic mice. Moreover, CWP reduces oxidative stress, protects central nervous system (CNS) neurons, and improves the neurobehavior of diabetic mice (47). A recent study demonstrated that the expression of MIP-1α and MIP2 processes, which play important roles in tissue repair processes, (48, 49) was increased in diabetic mice following treatment with WP, which also accelerated the healing of diabetic wounds (50). Decreased fibroblast proliferation delays collagen deposition in acute wounds of type 1 diabetic patients. WP potentially enhances tissue regeneration in diabetic mice by increasing hydroxyproline and collagen levels (51); decreases in hydroxyproline content critically impair wound healing in diabetic rats (52). Interestingly, diabetic mice supplemented with CWP exhibited a significant improvement in collagen deposition, a reduction in inflammatory stimuli, restored activation of STAT3, Akt, and NF-κB, and improvement in the closure of diabetic wounds compared with control mice. Most importantly, supplementation with CWP restored functional, long-lived wound-resident macrophages in the diabetic mice (34). Moreover, we recently provided evidence of immunomodulatory role of CWP in decreasing the tendency of the offspring of diabetic mothers to develop diabetes and related complications by restoring the expression of ATF-3 and the levels of ROS, pro-inflammatory cytokines, IL-2, and IL-7 in the offspring (53).
Antitumor effects of camel whey protein
The great anti-tumor effects of camel products are widely observed by peoples who use a mixture of camel milk and urine in the treatment of patients suffering from a variety of cancers. The current review showed studies attempting to find evidence to support these claimed anti-tumor effects of camel whey protein suggested that these anti-tumor effects could be both a direct cell cytotoxicity and cutting blood supply to tumor cells (anti-angiogenic action) or significant inhibition of the expression of the gene encoding carcinogen that could ultimately lead to the discovery of an effective anti-cancer drug. Most studies of the effects of whey protein on cancer have employed bovine whey protein. However, Habib, et al examined the impact of camel milk lactoferrin (CML) on colon cancer cells and observed that a high concentration of CML induced growth arrest in colon cancer cells (54). A study in 1990 in an animal model of colon carcinoma induced by injecting mice with 1,2-dimethylhydrazine (DMH) revealed that whey protein decreased tumor burden compared with mice fed a casein or Purina diet. Additionally, the animals fed the diet supplemented with CWP exhibited 0% mortality, whereas the animals fed casein or Purina exhibited 33% mortality (55). A study concluded that whey protein in milk had a significant role as an antitumor and anticarcinogenic agent by providing cysteine (a substrate for GSH synthesis) to enhance GSH synthesis in numerous tissues and, consequently, the detoxification of free radicals during carcinogenesis (56). The effect of the protein source (whey, casein, soybean, and red meat) on the incidence, burden, and mass index of DMH-induced intestinal tumors in male rats was examined. WPs exhibited the greatest antitumor activity against DMH-induced intestinal tumors compared with the other examined proteins. In addition, a significant increase in liver GSH levels was observed in the whey protein-fed rats (57). A study that compared the influence of whey and soy proteins on two generations of female rats indicated that WP had a two-fold greater effect on reducing tumor incidence and multiplicity compared with soy protein (58). CWP exerts its anticancer effects by increasing GSH concentrations in tissues and subsequently decreasing the tumor volume due to stimulation of the immune response (59). Researchers studied the impact of dietary supplementation with CWP on the azoxymethane-induced formation of colon tumors in male rats. The risk of colon cancer was reduced following long-term consumption of WP (60). WP in milk consists of LF, β-LG, α-LA, glycomacropeptide, and Igs, which act as biological components that enhance the immune system. WP also functions as an antioxidant, antitumor, antihypertensive, hypolipidemic, antiviral, antibacterial, and chelating agent. Most of the effects of WP are mediated by the formation of GSH from cysteine (9). Researchers reported that LF regulates the immune response and protects patients from post-surgical complications (61). Bovine whey protein is primarily composed of β-LG (approximately half of the WP content); by contrast, human milk does not contain β-LG. β-LG contains many essential amino acids and proteins, such as retinol-binding protein, which can modulate lymphatic responses by transporting small hydrophobic molecules, including retinoic acid (62). A decrease in nuclear GSH levels of 40–50% leads to DNA fragmentation and apoptosis (63). A high amount of intracellular GSH protects cells from apoptosis induced by anticancer drugs (64). Nukumi et al demonstrated that whey acidic protein (WAP) decreases the proliferation of human breast cancer cells (MCF-7 and MDA-MB-453 cells), suggesting that WAP can be used as a therapeutic factor for the treatment of breast cancer (65).
Antihypertensive effects of camel whey protein
Fermented camel milk contains angiotensin I-converting enzyme (ACE) inhibitory peptides (66). These peptides regulate blood pressure (67) and are produced by the proteolytic digestion of casein and WP (68). The C-terminal sequences of these peptides bind to the active site of ACE to inhibit ACE activity (69). Furthermore, the hypocholesterolemic effect of camel milk proteins may be due to the presence of bioactive peptides that react with and reduce cholesterol levels (70, 71). The hypoglycemic action of camel milk proteins may be due to many factors, including the presence of half-cystine, an insulin/insulin-like component present in CWP and small Igs (72, 73).
Anti-thermal effects of camel whey proteins
Heat stress causes severe economic loss in animal production. Heat-stressed animals suffer from compromised intestinal integrity, leading to endotoxemia, which is associated with reduced animal production. Pigs fed a combination of bovine whey protein and different doses of colostral whey protein (WP80, WP89, WP100) for 24 hr did not exhibit improvement of the reduced intestinal integrity resulting from severe and constant heat stress (32 °C for 24 hr continuously). Moreover, WP80 (which contained colostral WP as 20% of the diet) increased ileal permeability. Severe heat stress may have prevented positive effects of whey protein and colostral whey protein on intestinal integrity (74). However, no study has investigated the effects of CWP on mild or cyclical heat stress (which mimics natural heat stress events) in animal models, and further investigations are required.
Antibacterial properties of camel whey protein
CWP contains many components that are reported to have antibacterial effects against gram-positive and gram-negative bacteria. These components include lysozyme, LF, LPO, hydrogen peroxide, and Igs, all of which can inhibit bacterial growth (7). Additionally, lysozyme, LF, LPO, and some Igs inhibit Lactococcus lactis sup sp. cremoris, Escherichia coli, Staphylococcus aureus and rotavirus (75). LF inhibits Salmonella typhimurium growth by binding to iron, which is essential for bacterial growth, in the surroundings, thus inducing a bacteriostatic effect (75, 76). However, lysozyme inhibits the growth of some bacteria by damaging the bacterial cell wall. Additionally, camel milk lysozyme has greater activity against S. typhimurium than lysozyme from other types of milk (24). Igs, LF, and lysozyme are present at higher concentrations in CWP than in bovine milk (77, 78). α-LA is one of the most active components of CWPs and can protect newborns from infection with different pathogens (79-81). Interestingly, LPO has bacteriostatic activity against some gram-positive strains and bactericidal activity against some gram-negative strains. Moreover, camel milk contains large amounts of antibodies that protect against rotavirus (75). Inhibition of Listeria monocytogenes has been attributed to camel LPO, whereas inhibition of S. aureus may be mediated by the hydrogen peroxide present in camel milk (82). The antibacterial activity of CWPs against E. coli is significantly enhanced through hydrolysis by proteinase k (11).
Supporting this result, a study that used pepsin and pancreatin to digest camel colostral whey proteins into peptides demonstrated that the colostral whey protein still exhibited antibacterial activity against Gram-negative and gram-positive bacteria after enzymatic digestion. Additionally, different activities were enhanced by enzymatic digestion, such as antioxidant activity and ACE inhibition. Interestingly, these activities were enhanced following the release of the bioactive peptides from the hydrolysate of colostral whey protein, suggesting that the antibacterial activity of the colostrum whey peptides, such as the Lf peptides, is stronger than that of the native LF (83). These results indicate that CWPs and their hydrolysates can be used as natural antioxidants in food to reduce oxidative stress levels (11).
Antiviral activities of camel whey protein
LF displays the most potent antiviral activity of all components present in milk (84-86). LF can react with various molecules to elicit antibacterial (87), antiviral (240, iron binding (25), antioxidant, immunomodulatory, and anti-inflammatory activi-ties (25, 88), and to function as a growth factor (89). LF exhibits antiviral activity against a broad spectrum of viruses, including RNA and DNA viruses such as cytomegalovirus, hepatitis C virus (HCV), hepatitis B virus (HBV), herpes simplex virus (HSV), HIV, simian rotaviruses, and adenoviruses (90). The content of LF in CWP is higher than that in WPs from other animals. Camel lactoferrin (CLF) prevents HCV from entering Huh 7.5 cells, neutralizes intracellular HCV molecules, and suppresses HCV replication in infected Huh7.5 cells (91). CLF not only inhibits HCV (genotype 4a) entry into HepG2 cells and peripheral blood mononuclear cells (PBMCs) but also suppresses viral replication in infected cells (85). Researchers determined that the inhibitory effect of CLF on HCV (genotype 4a) was two-fold higher than that of human, sheep, and bovine LF (92). The behavior of CLF encompasses the activities of lacto-ferrin, an iron-binding protein, and an iron trans-porter protein and is thus unique among lactoferrins and transferrins present in milk from animals (93). CLF has dual functions: inhibiting lipid peroxidation and binding to and transporting iron at different pH values, thus monitoring iron content in hepatocytes (92). CLF inhibits HCV entry into HepG2 cells and PBMCs by directly blocking the viral molecules. The inhibition of HCV replication in infected HepG2 cells and PBMCs may be due to the dual nature of LF as an iron binding protein and iron transporter protein (85). Interestingly, the carboxyl region of LF resembles human CD81 (tetraspanin) and may be responsible for binding to the HCV envelope protein (94, 95). CLF has been postulated to have hepatoprotective activity because it can improve imbalances in the levels of Th1/Th2 cytokines (79, 96).
Generally, several mechanisms are suggested to mediate the antiviral activity of CLF. LF may directly interact with viral molecules such as HCV (80,84-86,97), HIV (98), and adenovirus (99). 2) LF may interact with heparin sulfate (HS), which is the viral attachment receptor on the cell surface, to prevent the attachment of the virus to the host cell, thus preventing infection. This mechanism is most accepted (100) because heparin sulfate proteoglycans (HSPGS), which are widely distributed in mammalian cells, are essential binding sites for many viruses (101-10) such as HSV (105), HCV (106), HBV (107), HIV (108), dengue virus (109), and human papilloma virus (110). 3) The virucidal venue mechanism depends on the α-helical structure and cationic nature of LF (111-114). α-LA failed to prevent entry of HCV into Huh 7.5 cells and PBMCs at concentrations of 0.5 and 1 mg/ml. Camel IgGs inhibited the entry of HCV into PBMCs and Huh 7.5 cells (84). Camel IgGs are classified into three major
subclasses (IgG1, IgG2, IgG3) (115). IgG2 and IgG3 can penetrate the active sites of some enzymes (93, 115). This mechanism may explain their ability to inhibit some enzymes of HIV-1, such as reverse transcriptase, protease, and integrase, which are all essential for the life cycle of this virus (116). Camel LPO binds and neutralizes HCV particles in HepG2 cells at concentrations of 1 and 1.5 mg/ml, with an efficiency of approximately 100%. In addition, camel LPO inhibits viral replication inside infected cells in vitro. The activity of camel LPO is more efficient than that of human and bovine proteins. Camel LPO may inhibit HCV replication by neutralizing the intracellular virus or activating the LPO-based oxidative system. This intracellular oxidative system is activated when LPO enters the cell and reacts with the cellular substrates to produce short-lived ROS. These ROS suppress the synthesis of the viral nucleic acids and proteins inside the cell. Another hypothesis suggests that LPO, as a calcium- and iron-containing glycoprotein (110, 117) may regulate iron homeostasis in the liver, thus protecting hepatocytes from HCV infection (118). Additionally, LPO reportedly inhibits both RNA and DNA viruses, such as HSV, respiratory syncytial virus, HIV, and echovirus type II (119-121).
Conclusion
Camel whey protein is a reliable source of amino acids and biologically active proteins which act as a nutritional supplement. There is growing evidence that CWP possesses therapeutic properties in different pathological conditions. The current review presents highlights about the potential beneficial effects of camel whey protein and focuses on clinical trials using camel whey protein supplementation as an immuno-modulator, antioxidant, anti-inflammatory, antidiabetic, anti-thermal, and antihypertensive agent as summariz-ed in (Table 2). In this context, this review showed that camel whey protein supplementation is shown to maintain a high concentration of cellular antioxidants and boost immune defenses that promote carcinogen detoxification. Due to the positive findings, camel whey protein supplementation is starting to be viewed as a non-pharmaceutical adjunct therapy in the treatment of cancer. Moreover, CWP provides an abundant supply of essential amino acids to organs and tissues, which stimulates tissue regenerative mechanisms and help minimize immune suppression. Furthermore, camel milk contains the angiotensin I-converting enzyme (ACE), an inhibitory peptide that regulates blood pressure and is produced by the proteolytic digestion of casein and WP. The therapeutic utilization of CWP in diabetes is through ameliorating impaired wound healing in diabetic patients by decreasing blood glucose levels, oxidative stress, growth factor levels, and the inflammatory response. Moreover, CWP reacts with various molecules to elicit antibacterial, antiviral, iron binding, antioxidant, immunomodulatory, and anti-inflammatory activities. CWP exhibits antiviral activity against a broad spectrum of viruses, including RNA and DNA viruses by neutralizing intracellular viral molecules and suppressing virus replication in cells. Finally, the current review has summarized scientific evidence for the therapeutic actions of CWP, and efforts are underway to more precisely identify the therapeutic components of CWP. Thus, the camel is proving to be a ‘gold mine’ for researchers to investigate functional therapeutic benefits of CWP.
Table 2.
The biological activities of camel whey protein The biological activities of camel whey proteins were summarized from the literature
| Biological activities of camel whey proteins | ||
|---|---|---|
| Biological activity | Functions | Ref |
| CWP enhances lymphocyte functions, chemotaxis, phagocytic activity; and granulocyte and NK cell activity | (27) | |
| CWP enhances production of IL-1β, IL-8, IL-6, macrophage inflammatory proteins (MIP-1α, MIP-1β), and tumor necrosis factor (TNF-α) | (28) | |
| CWP increases phagocytosis and the secretion of IgA from Payer’s patches | (31) | |
| CWP regulates expression of TNF-α and cell death receptor (Fas) mRNAs and subsequently enhances the closure and healing of diabetic wounds. | (35) | |
| Immunomodulatory Effects | CWP improves primary and secondary intestinal tract antibody responses to a variety of different vaccine antigens | (38) |
| CWP accelerates cutaneous wound healing via decreasing the oxidative stress and restoring pro-inflammatory cytokines levels and β-defensin. | (19) | |
| CWP increase GSH synthesis and improve the cellular antioxidant defense system | (46) | |
| Antidiabetic Effects | CWP decrease blood glucose levels, oxidative stress, growth factor levels, and the inflammatory response. | (47) |
| CWP enhances tissue regeneration in diabetic mice by increasing hydroxyproline and collagen levels | (51) | |
| Camel milk lactoferrin-induced growth arrest in colon cancer cells. | (54) | |
| Whey protein provides cysteine which enhances GSH synthesis and detoxification of free radicals during carcinogenesis | (56) | |
| Antitumor Effects | Whey acidic protein decreases the proliferation of human breast cancer cells (MCF-7 and MDA-MB-453 cells) | (65) |
| Antihypertensive Effects | Camel milk contains angiotensin I-converting enzyme (ACE) inhibitory peptides which regulate blood pressure. | (66, 67) |
| Anti-thermal Effects | CWP reduces intestinal integrity resulting from severe and constant heat stress | (74) |
| CWP components include lysozyme, LF, LPO, hydrogen peroxide, and Igs, which inhibit bacterial growth | (7). | |
| lysozyme, LF, LPO, and some Igs inhibit Lactococcus lactis sup sp. cremoris, Escherichia coli, Staphylococcus aureus, and rotavirus | (75). | |
| Antibacterial Activities | LF inhibits Salmonella typhimurium growth by binding to iron, which is essential for bacterial growth. | (75, 76) |
| LF exhibits antiviral activity against a broad spectrum of viruses such as cytomegalovirus, HCV, HBV, HSV, HIV, simian rotaviruses, and adenoviruses. | (90) | |
| Camel lactoferrin suppresses HCV replication in infected Huh7.5 cells and inhibits HCV (genotype 4a) entry into HepG2 cells and peripheral blood mononuclear cells | (91, 85) | |
| Antiviral Activities | LF prevents viral infection by interacting with heparin sulfate (HS), which represents the viral attachment receptor on the cell surface. | (100,101,105-110) |
| LPO inhibits HCV replication by neutralizing the intracellular virus or activating the LPO-based oxidative system, regulates iron homeostasis in the liver, thus protecting hepatocytes from HCV infection | (110,117,118) | |
Conflict of interest
The authors declare that they have no conflict of interest. This manuscript has not been published or submitted elsewhere. All authors have approved the final article.
References
- 1.Ebaid H, Badr G, Metwalli A. Immunoenhancing property of dietary un-denatured whey protein derived from three camel breeds in mice. Biologia. 2012;67:425–433. [Google Scholar]
- 2.Badr G, Badr BM, Mahmoud MH, Mohany M, Rabah DM, Garraud O. Treatment of diabetic mice with undenatured whey protein accelerates the wound healing process by enhancing the expression of mip-1α, mip-2, kc, cx3cl1 and tgf-βin wounded tissue. BMC Immunol. 2012;13:32. doi: 10.1186/1471-2172-13-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Badr G. Camel whey protein enhances diabetic wound healing in a streptozotocin-induced diabetic mouse model:The critical role of β-defensin-1,-2 and-3. Lipids Health Dis. 2013;12:1. doi: 10.1186/1476-511X-12-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Badr G. Supplementation with undenatured whey protein during diabetes mellitus improves the healing and closure of diabetic wounds through the rescue of functional long-lived wound macrophages. Cell Physiol Biochem. 2012;29:571–582. doi: 10.1159/000338511. [DOI] [PubMed] [Google Scholar]
- 5.Badr G, Mohany M, Metwalli A. Effects of undenatured whey protein supplementation on cxcl12-and ccl21-mediated b and t cell chemotaxis in diabetic mice. Lipids Health Dis. 2011;10:1. doi: 10.1186/1476-511X-10-203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Badr G, Ebaid H, Mohany M, Abuelsaad AS. Modulation of immune cell proliferation and chemotaxis towards cc chemokine ligand (ccl)-21 and cxc chemokine ligand (cxcl)-12 in undenatured whey protein-treated mice. J Nutr Biochem. 2012;23:1640–1646. doi: 10.1016/j.jnutbio.2011.11.006. [DOI] [PubMed] [Google Scholar]
- 7.Mohany M, Badr BM, Mahmoud MH, Garraud O, Badr G. Un-denatured whey protein expedites wound healing in diabetic mice model by enhancing the expression of beta-defensin 2, 3 and vascular endothelial growth factor (vegf) in the wounded tissue. Afr J Microbiol Res. 2012;6;:2137–2144. [Google Scholar]
- 8.Power O, Jakeman P, FitzGerald R. Antioxidative peptides:Enzymatic production in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids. 2013;44:797–820. doi: 10.1007/s00726-012-1393-9. [DOI] [PubMed] [Google Scholar]
- 9.Marshall K. Therapeutic applications of whey protein. Altern Med Rev. 2004;9:136–156. [PubMed] [Google Scholar]
- 10.El-Hatmi H, Girardet JM, Gaillard JL, Yahyaoui MH, Attia H. Characterisation of whey proteins of camel (camelus dromedarius) milk and colostrum. SmallRumin Res. 2007;70:267–271. [Google Scholar]
- 11.Salami M, Moosavi-Movahedi AA, Ehsani MR, Yousefi R, Haertleé T, Chobert JM, et al. Improvement of the antimicrobial and antioxidant activities of camel and bovine whey proteins by limite proteolysis. J AgricFoodChem. 2010;58:3297–3302. doi: 10.1021/jf9033283. [DOI] [PubMed] [Google Scholar]
- 12.Abdel-Aziem SH, Hassan AM, Abdel-Wahhab MA. Dietary supplementation with whey protein and ginseng extract counteracts oxidative stress and DNA damage in rats fed an aflatoxin-contaminated diet. Mutat Res Genet Toxicol Environ Mutagen. 2011;723:65–71. doi: 10.1016/j.mrgentox.2011.04.007. [DOI] [PubMed] [Google Scholar]
- 13.Sousa GT, Lira FS, Rosa JC, de Oliveira EP, Oyama LM, Santos RV, et al. Dietary whey protein lessens several risk factors for metabolic diseases:A review. Lipids Health Dis. 2012;11:1–9. doi: 10.1186/1476-511X-11-67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ebaid H, Ahmed OM, Mahmoud AM, Ahmed RR. Limiting prolonged inflammation during proliferation and remodeling phases of wound healing in streptozotocin-induced diabetic rats supplemented with camel undenatured whey protein. BMC Immunol. 2013;14:1–13. doi: 10.1186/1471-2172-14-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Khaskheli M, Arain M, Chaudhry S, Soomro A, Qureshi T. Physico-chemical quality of camel milk. J Agri Soc Sci. 2005;2:164–166. [Google Scholar]
- 16.Kappeler SR, Heuberger C, Farah Z, Puhan Z. Expression of the peptidoglycan recognition protein, pgrp, in the lactating mammary gland. JDairySci. 2004;87:2660–2668. doi: 10.3168/jds.S0022-0302(04)73392-5. [DOI] [PubMed] [Google Scholar]
- 17.Elagamy EI. Effect of heat treatment on camel milk proteins with respect to antimicrobial factors:A comparison with cows’ and buffalo milk proteins. Food Chem. 2000;68:227–232. [Google Scholar]
- 18.Lucas DO. Breakthrough technology produces concentrated whey protein with bioactive immunoglobulins. Clin Nutr Insight. 1999;6:1–4. [Google Scholar]
- 19.Laleye L, Jobe B, Wasesa A. Comparative study on heat stability and functionality of camel and bovine milk whey proteins. JDairySci. 2008;91:4527–4534. doi: 10.3168/jds.2008-1446. [DOI] [PubMed] [Google Scholar]
- 20.Hinz K, O’Connor PM, Huppertz T, Ross RP, Kelly AL. Comparison of the principal proteins in bovine, caprine, buffalo, equine and camel milk. JDairySci. 2012;79:185–191. doi: 10.1017/S0022029912000015. [DOI] [PubMed] [Google Scholar]
- 21.El-Agamy EI, Nawar M, Shamsia SM, Awad S, Haenlein GF. Are camel milk proteins convenient to the nutrition of cow milk allergic children?Small Rumin Res. 2009;82:1–6. [Google Scholar]
- 22.Kappeler S, Farah Z, Puhan Z. Alternative splicing of lactophorin mrna from lactating mammary gland of the camel (camelus dromedarius) J Dairy Sci. 1999;82:2084–2093. doi: 10.3168/jds.S0022-0302(99)75450-0. [DOI] [PubMed] [Google Scholar]
- 23.Cohen MS, Britigan BE, French M, Bean K. Preliminary observations on lactoferrin secretion in human vaginal mucus:Variation during the menstrual cycle, evidence of hormonal regulation, and implications for infection with neisseria gonorrhoeae. Am J Obstet Gynecol. 1987;157:1122–1125. doi: 10.1016/s0002-9378(87)80274-0. [DOI] [PubMed] [Google Scholar]
- 24.Harmsen MC, Swart PJ, de Béthune MP, Pauwels R, De Clercq E, Meijer DK. Antiviral effects of plasma and milk proteins:Lactoferrin shows potent activity against both human immunodeficiency virus and human cytomegalovirus replication in vitro. J Infect Dis. 1995;172:380–388. doi: 10.1093/infdis/172.2.380. [DOI] [PubMed] [Google Scholar]
- 25.Sanchez L, Calvo M, Brock JH. Biological role of lactoferrin. Arch Dis Child. 1992;67:657. doi: 10.1136/adc.67.5.657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Conesa C, Sánchez L, Rota C, Pérez MD, Calvo M, Farnaud S, et al. Isolation of lactoferrin from milk of different species:Calorimetric and antimicrobial studies. Comp Biochem Physiol B Biochem Mol Biol. 2008;150:131–139. doi: 10.1016/j.cbpb.2008.02.005. [DOI] [PubMed] [Google Scholar]
- 27.Gauthier SF, Pouliot Y, Saint-Sauveur D. Immunomodulatory peptides obtained by the enzymatic hydrolysis of whey proteins. Int Dairy J. 2006;16:1315–1323. [Google Scholar]
- 28.Rusu D, Drouin R, Pouliot Y, Gauthier S, Poubelle PE. A bovine whey protein extract stimulates human neutrophils to generate bioactive il-1ra through a nf-κb-and mapk-dependent mechanism. J Nutr. 2010;140:382–391. doi: 10.3945/jn.109.109645. [DOI] [PubMed] [Google Scholar]
- 29.Micke P, Beeh K, Schlaak J, Buhl R. Oral supplementation with whey proteins increases plasma glutathione levels of hiv-infected patients. Eur J Clin Invest. 2001;31:171–178. doi: 10.1046/j.1365-2362.2001.00781.x. [DOI] [PubMed] [Google Scholar]
- 30.Krissansen GW. Emerging health properties of whey proteins and their clinical implications. J Am Coll Nutr. 2007;26:713–723. doi: 10.1080/07315724.2007.10719652. [DOI] [PubMed] [Google Scholar]
- 31.Beaulieu J, Dupont C, Lemieux P. Whey proteins and peptides:Beneficial effects on immune health. 1st ed. Springer; 2006. [Google Scholar]
- 32.Loss G, Apprich S, Waser M, Kneifel W, Genuneit J, Büchele G, et al. The protective effect of farm milk consumption on childhood asthma and atopy:The gabriela study. J Allergy Clin Immunol. 2011;128:766–773e764. doi: 10.1016/j.jaci.2011.07.048. [DOI] [PubMed] [Google Scholar]
- 33.Castro G, Maria D, Bouhallab S, Sgarbieri V. In vitro impact of a whey protein isolate (wpi) and collagen hydrolysates (chs) on b16f10 melanoma cells proliferation. J Dermatol Sci. 2009;56:51–57. doi: 10.1016/j.jdermsci.2009.06.016. [DOI] [PubMed] [Google Scholar]
- 34.Ebaid H, Salem A, Sayed A, Metwalli A. Whey protein enhances normal inflammatory responses during cutaneous wound healing in diabetic rats. Lipids Health Dis. 2011;10:1. doi: 10.1186/1476-511X-10-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ebaid H. Promotion of immune and glycaemic functions in streptozotocin-induced diabetic rats treated with un-denatured camel milk whey proteins. Nutr Metab (Lond) 2014;11:31. doi: 10.1186/1743-7075-11-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ebaid H. Neutrophil depletion in the early inflammatory phase delayed cutaneous wound healing in older rats:Improvements due to the use of un-denatured camel whey protein. Diagn Pathol. 2014;9:46. doi: 10.1186/1746-1596-9-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Walzem R, Dillard C, German J. Whey components:Millennia of evolution create functionalities for mammalian nutrition:What we know and what we may be overlooking. Crit Rev Food Sci Nutr. 2002;42:353–375. doi: 10.1080/10408690290825574. [DOI] [PubMed] [Google Scholar]
- 38.Low PPL, Rutherford KJ, Gill HS, Cross ML. Effect of dietary whey protein concentrate on primary and secondary antibody responses in immunized BALB/CMice. Int Immunopharmacol. 2003;3:393–401. doi: 10.1016/S1567-5769(02)00297-7. [DOI] [PubMed] [Google Scholar]
- 39.Faloon PW, Chou DHC, Forbeck EM, Walpita D, Morgan B, Buhrlage S, et al. Identification of small molecule inhibitors that suppress cytokine-induced apoptosis in human pancreatic islet cells. Probe Reports from the NIH Molecular Libraries Program. 2011 [PubMed] [Google Scholar]
- 40.Bounous G, Batist G, Gold P. Immunoenhancing property of dietary whey protein in mice:Role of glutathione. Clin Invest Med. 1989;12:154–161. [PubMed] [Google Scholar]
- 41.Belokrylov G, Popova OY, Molchanova I, Sorochinskaya E, Anokhina V. Peptides and their constituent amino acids influence the immune response and phagocytosis in different ways. Int J Immunopharmacol. 1992;14:1285–1292. doi: 10.1016/0192-0561(92)90065-s. [DOI] [PubMed] [Google Scholar]
- 42.Kawasaki H. Comparative studies on proteodermatan sulfate of bovine gastrointestinal tract. Tohoku J Exp Med. 1993;171:255–266. doi: 10.1620/tjem.171.255. [DOI] [PubMed] [Google Scholar]
- 43.Marchetti M, Pisani S, Antonini G, Valenti P, Seganti L, Orsi N. Metal complexes of bovine lactoferrin inhibit in vitro replication of herpes simplex virus type 1 and 2. Biometals. 1998;11:89–94. doi: 10.1023/a:1009217709851. [DOI] [PubMed] [Google Scholar]
- 44.Wong CW, Watson DL. Immunomodulatory effects of dietary whey proteins in mice. J Dairy Res. 1995;62:359–368. doi: 10.1017/s0022029900031058. [DOI] [PubMed] [Google Scholar]
- 45.Machnicki M, Zimecki M, Zagulski T. Lactoferrin regulates the release of tumour necrosis factor alpha and interleukin 6 in vivo. Int J Exp Pathol. 1993;74:433. [PMC free article] [PubMed] [Google Scholar]
- 46.Ögünç V, Manukyan Cingi, Eksioglu-Demiralp Aktan Ö, Yalçın S. Dietary whey supplementation in experimental models of wound healing. Int J Vitam Nutr Res. 2008;78:70–73. doi: 10.1024/0300-9831.78.2.70. [DOI] [PubMed] [Google Scholar]
- 47.Ajarem J, Allam AA, Ebaid H, Maodaa SN, AL-Sobeai SM, Rady AM, et al. Neurochemical, structural and neurobehavioral evidence of neuronal protection by whey proteins in diabetic albino mice. Behav Brain Funct. 2015;11:1. doi: 10.1186/s12993-015-0053-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Takamiya M, Fujita S, Saigusa K, Aoki Y. Simultaneous detections of 27 cytokines during cerebral wound healing by multiplexed bead-based immunoassay for wound age estimation. J Neurotrauma. 2007;24:1833–1844. doi: 10.1089/neu.2007.0336. [DOI] [PubMed] [Google Scholar]
- 49.Lin Q, Fang D, Fang J, Ren X, Yang X, Wen F, et al. Impaired wound healing with defective expression of chemokines and recruitment of myeloid cells in tlr3-deficient mice. J Immunol. 2011;186:3710–3717. doi: 10.4049/jimmunol.1003007. [DOI] [PubMed] [Google Scholar]
- 50.Mori R, Kondo T, Ohshima T, Ishida Y, Mukaida N. Accelerated wound healing in tumor necrosis factor receptor p55-deficient mice with reduced leukocyte infiltration. FASEB J. 2002;16:963–974. doi: 10.1096/fj.01-0776com. [DOI] [PubMed] [Google Scholar]
- 51.Black E, Vibe-Petersen J, Jorgensen LN, Madsen SM, Ågren MS, Holstein PE, et al. Decrease of collagen deposition in wound repair in type 1 diabetes independent of glycemic control. Arch Surg. 2003;138:34–40. doi: 10.1001/archsurg.138.1.34. [DOI] [PubMed] [Google Scholar]
- 52.Zhang Z, Zhao M, Wang J, Ding Y, Dai X, Li Y. Oral administration of skin gelatin isolated from chum salmon (oncorhynchus keta) enhances wound healing in diabetic rats. Mar Drugs. 2011;9:696–711. doi: 10.3390/md9050696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Mahmoud MH, Badr G, El Shinnawy NA. Camel whey protein improves lymphocyte function and protects against diabetes in the offspring of diabetic mouse dams. Int J Immunopathol Pharmacol. 2016;29:632–646. doi: 10.1177/0394632016671729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Habib HM, Ibrahim WH, Schneider-Stock R, Hassan HM. Camel milk lactoferrin reduces the proliferation of colorectal cancer cells and exerts antioxidant and DNA damage inhibitory activities. Food Chem. 2013;141:148–152. doi: 10.1016/j.foodchem.2013.03.039. [DOI] [PubMed] [Google Scholar]
- 55.Papenburg R, Bounous G, Fleiszer D, Gold P. Dietary milk proteins inhibit the development of dimethylhydrazine-induced malignancy. Tumour Biol. 1990;11:129–136. doi: 10.1159/000217647. [DOI] [PubMed] [Google Scholar]
- 56.Bounous G, Batist G, Gold P. Whey proteins in cancer prevention. Cancer Lett. 1991;57:91–94. doi: 10.1016/0304-3835(91)90200-2. [DOI] [PubMed] [Google Scholar]
- 57.McIntosh GH, Regester GO, Le Leu RK, Royle PJ, Smithers GW. Dairy proteins protect against dimethylhydrazine-induced intestinal cancers in rats. J Nutr. 1995;125:809–816. doi: 10.1093/jn/125.4.809. [DOI] [PubMed] [Google Scholar]
- 58.Hakkak R, Korourian S, Shelnutt SR, Lensing S, Ronis MJ, Badger TM. Diets containing whey proteins or soy protein isolate protect against 7,12-dimethylbenz(a)anthracene-induced mammary tumors in female rats. Cancer Epidemiol Biomarkers Prev. 2000;9:113–117. [PubMed] [Google Scholar]
- 59.Bounous G. Whey protein concentrate (wpc) and glutathione modulation in cancer treatment. Anticancer Res. 2000;20:4785–4792. [PubMed] [Google Scholar]
- 60.Hakkak R, Korourian S, Ronis MJ, Johnston JM, Badger TM. Dietary whey protein protects against azoxymethane-induced colon tumors in male rats. Cancer Epidemiol Biomarkers Prev. 2001;10:555–558. [PubMed] [Google Scholar]
- 61.Zimecki M, Wlaszczyk A, Wojciechowski R, Dawiskiba J, Kruzel M. Lactoferrin regulates the immune responses in post-surgical patients. Arch Immunol Ther Exp. 2001;49:325–333. [PubMed] [Google Scholar]
- 62.Guimont C, Marchall E, Girardet JM, Linden G. Biologically active factors in bovine milk and dairy byproducts: Influence on cell culture. Crit RevFoodSci Nut. 1997;37:393–410. doi: 10.1080/10408399709527780. [DOI] [PubMed] [Google Scholar]
- 63.Morales A, Miranda M, Sanchez-Reyes A, Biete A, Fernandez-Checa JC. Oxidative damage of mitochondrial and nuclear DNA induced by ionizing radiation in human hepatoblastoma cells. Int J Radiat Oncol Biol Phys. 1998;42:191–203. doi: 10.1016/s0360-3016(98)00185-0. [DOI] [PubMed] [Google Scholar]
- 64.Friesen C, Kiess Y, Debatin KM. A critical role of glutathione in determining apoptosis sensitivity and resistance in leukemia cells. Cell Death Differ. 2004;1:S73–85. doi: 10.1038/sj.cdd.4401431. [DOI] [PubMed] [Google Scholar]
- 65.Nukumi N, Iwamori T, Kano K, Naito K, Tojo H. Reduction of tumorigenesis and invasion of human breast cancer cells by whey acidic protein (wap) Cancer Lett. 2007;252:65–74. doi: 10.1016/j.canlet.2006.12.005. [DOI] [PubMed] [Google Scholar]
- 66.Tsuda H, Miyamoto T. Angiotensin I-converting enzyme inhibitory peptides in skim milk fermented with lactobacillus helveticus 130b4 from camel milk in inner mongolia, china. J Sci Food Agric. 2008;88:2688–2692. [Google Scholar]
- 67.Smith CG, Vane JR. The discovery of captopril. FASEB J. 2003;17:788–789. doi: 10.1096/fj.03-0093life. [DOI] [PubMed] [Google Scholar]
- 68.Alhaj OAKA, Peters A. The effect of bifid bacterium lactis and trypsin on cholesterol international food and health innovation conference. Malmö, Sweden: Skåne Food Innovation Network; 2006. [Google Scholar]
- 69.López-Fandiño R, Otte J, Van Camp J. Physiological, chemical and technological aspects of milk-protein-derived peptides with antihypertensive and ace-inhibitory activity. Int Dairy J. 2006;16:1277–1293. [Google Scholar]
- 70.Li H, Papadopoulos V. Peripheral-type benzodiazepine receptor function in cholesterol transport. Identification of a putative cholesterol recognition/interaction amino acid sequence and consensus pattern 1. Endocrinol. 1998;139:4991–4997. doi: 10.1210/endo.139.12.6390. [DOI] [PubMed] [Google Scholar]
- 71.Seelig A, Seelig J. In interaction of drugs and peptides with the lipid membrane.Alfred Benzon Symposium, Munksgaard; 1996:184–191. [Google Scholar]
- 72.Agrawal R, Budania S, Sharma P, Gupta R, Kochar D, Panwar R, Sahani M. Zero prevalence of diabetes in camel milk consuming raica community of north-west rajasthan, india. DiabetesRes Clin Pract Supp. 2007;76:290–296. doi: 10.1016/j.diabres.2006.09.036. [DOI] [PubMed] [Google Scholar]
- 73.Agrawal RP, Saran S, Sharma P, Gupta RP, Kochar DK, Sahani MS. Effect of camel milk on residual β-cell function in recent onset type 1 diabetes. DiabetesRes Clin Pract. 2007;77:494–495. doi: 10.1016/j.diabres.2007.01.012. [DOI] [PubMed] [Google Scholar]
- 74.Sanz Fernandez MV, Pearce SC, Mani V, Gabler NK, Metzger L, Patience JF, Rhoads RP, et al. Effects of dairy products on intestinal integrity in heat-stressed pigs. Temperature. 2014;1:128–134. doi: 10.4161/temp.29561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.El Sayed I, Ruppanner R, Ismail A, Champagne CP, Assaf R. Antibacterial and antiviral activity of camel milk protective proteins. J Dairy Res. 1992;59:169–175. doi: 10.1017/s0022029900030417. [DOI] [PubMed] [Google Scholar]
- 76.Ochoa TJ, Cleary TG. Effect of lactoferrin on enteric pathogens. Biochimie. 2009;91:30–34. doi: 10.1016/j.biochi.2008.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Benkerroum N. Antimicrobial activity of lysozyme with special relevance to milk. Afric J Biotechnol. 2008:7. [Google Scholar]
- 78.Konuspayeva G, Faye B, Loiseau G, Levieux D. Lactoferrin and immunoglobulin contents in camel’s milk (camelus bactrianus, camelus dromedarius, and hybrids) from kazakhstan. J Dairy Sci. 2007;90:38–46. doi: 10.3168/jds.S0022-0302(07)72606-1. [DOI] [PubMed] [Google Scholar]
- 79.Hara K, Ikeda M, Saito S, Matsumoto S, Numata K, Kato N, et al. Lactoferrin inhibits hepatitis b virus infection in cultured human hepatocytes. Hepatol Res. 2002;24:228–235. doi: 10.1016/s1386-6346(02)00088-8. [DOI] [PubMed] [Google Scholar]
- 80.Redwan ERM, Tabll A. Camel lactoferrin markedly inhibits hepatitis c virus genotype 4 infection of human peripheral blood leukocytes. J Immunoassay Immunochem. 2007;28:267–277. doi: 10.1080/15321810701454839. [DOI] [PubMed] [Google Scholar]
- 81.Håkansson A, Svensson M, Mossberg AK, Sabharwal H, Linse S, Lazou I, et al. A folding variant of α-lactalbumin with bactericidal activity against streptococcus pneumoniae. Mol Microbiol. 2000;35:589–600. doi: 10.1046/j.1365-2958.2000.01728.x. [DOI] [PubMed] [Google Scholar]
- 82.Benkerroum N, Mekkaoui M, Bennani N, Hidane K. Antimicrobial activity of camel’s milk against pathogenic strains of escherichia coli and listeria monocytogenes. Int J Dairy Technol. 2004;57:39–43. [Google Scholar]
- 83.Jrad Z, El Hatmi H, Adt I, Girardet JM, Cakir-Kiefer C, Jardin J, et al. Effect of digestive enzymes on antimicrobial, radical scavenging and angiotensin i-converting enzyme inhibitory activities of camel colostrum and milk proteins. Dairy Sci Technol. 2014;94:205–224. [Google Scholar]
- 84.EL-Fakharany EM, Abedelbaky N, Haroun BM, Sánchez L, Redwan NA, Redwan EM. Anti-infectivity of camel polyclonal antibodies against hepatitis c virus in huh7. 5 hepatoma. Virol J. 2012;9:1. doi: 10.1186/1743-422X-9-201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Esmail ME, Ashraf T, El WA, Bakry MH, El-Rashdy MR. Potential activity of camel milk-amylase and lactoferrin against hepatitis c virus infectivity in hepg2 and lymphocytes. Hepat Mon. 2008;2008:101–109. [Google Scholar]
- 86.Liao Y, El-Fakkarany E, Lönnerdal B, Redwan EM. Inhibitory effects of native and recombinant full-length camel lactoferrin and its n and c lobes on hepatitis c virus infection of huh7. 5 cells. J Med Microbiol. 2012;61:375–383. doi: 10.1099/jmm.0.033894-0. [DOI] [PubMed] [Google Scholar]
- 87.Arnold RR, Russell JE, Champion WJ, Brewer M, Gauthier JJ. Bactericidal activity of human lactoferrin: Differentiation from the stasis of iron deprivation. Infect Immun. 1982;35:792–799. doi: 10.1128/iai.35.3.792-799.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Crouch S, Slater K, Fletcher J. Regulation of cytokine release from mononuclear cells by the iron-binding protein lactoferrin. Blood. 1992;80:235–240. [PubMed] [Google Scholar]
- 89.Hashizume S, Kuroda K, Murakami H. Identification of lactoferrin as an essential growth factor for human lymphocytic cell lines in serum-free medium. Biochim Biophys Acta (BBA)-Mol Cell Res. 1983;763:377–382. doi: 10.1016/0167-4889(83)90099-x. [DOI] [PubMed] [Google Scholar]
- 90.Legrand D, Pierce A, Elass E, Carpentier M, Mariller C, Mazurier J. Lactoferrin structure and functions. In Bioactive components of milk. Springer; 2008. pp. 163–194. [DOI] [PubMed] [Google Scholar]
- 91.Redwan EM, EL-Fakharany EM, Uversky VN, Linjawi MH. Screening the anti infectivity potentials of native n-and c-lobes derived from the camel lactoferrin against hepatitis c virus. BMC Complement Altern Med. 2014;14:1. doi: 10.1186/1472-6882-14-219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.EL-Fakharany EM, Sánchez L, Al-Mehdar HA, Redwan EM. Effectiveness of human, camel, bovine and sheep lactoferrin on the hepatitis c virus cellular infectivity: Comparison study. Virol J. 2013;10:1. doi: 10.1186/1743-422X-10-199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Holt LJ, Herring C, Jespers LS, Woolven BP, Tomlinson IM. Domain antibodies: Proteins for therapy. Trends Biotechnol. 2003;21:484–490. doi: 10.1016/j.tibtech.2003.08.007. [DOI] [PubMed] [Google Scholar]
- 94.Bonkovsky HL, Banner BF, Rothman AL. Iron and chronic viral hepatitis. Hepatology. 1997;25:759–768. doi: 10.1002/hep.510250345. [DOI] [PubMed] [Google Scholar]
- 95.Farinati F, Cardin R, De Maria N, Della Libera G, Marafin C, Lecis E, et al. Iron storage, lipid peroxidation and glutathione turnover in chronic anti-hcv positive hepatitis. J Hepatol. 1995;22:449–456. doi: 10.1016/0168-8278(95)80108-1. [DOI] [PubMed] [Google Scholar]
- 96.Saltanat H, Li H, Xu Y, Wang J, Liu F, Geng X. [the influences of camel milk on the immune response of chronic hepatitis b patients] Xi bao yu fen zi mian yi xue za zhi=Chinese J Cell Mol Immunol. 2009;25:431–433. [PubMed] [Google Scholar]
- 97.Ikeda M, Sugiyama K, Tanaka T, Tanaka K, Sekihara H, Shimotohno K, et al. Lactoferrin markedly inhibits hepatitis c virus infection in cultured human hepatocytes. Biochem Biophys Res Commun. 1998;245:549–553. doi: 10.1006/bbrc.1998.8481. [DOI] [PubMed] [Google Scholar]
- 98.Berkhout B, van Wamel JL, Beljaars L, Meijer DK, Visser S, Floris R. Characterization of the anti-hiv effects of native lactoferrin and other milk proteins and protein-derived peptides. Antiviral Res. 2002;55:341–355. doi: 10.1016/s0166-3542(02)00069-4. [DOI] [PubMed] [Google Scholar]
- 99.Pietrantoni A, Di Biase AM, Tinari A, Marchetti M, Valenti P, Seganti L, et al. Bovine lactoferrin inhibits adenovirus infection by interacting with viral structural polypeptides. Antimicrob Agents Chemother. 2003;47:2688–2691. doi: 10.1128/AAC.47.8.2688-2691.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Van der Strate B, Beljaars L, Molema G, Harmsen M, Meijer D. Antiviral activities of lactoferrin. Antiviral Res. 2001;52:225–239. doi: 10.1016/s0166-3542(01)00195-4. [DOI] [PubMed] [Google Scholar]
- 101.Copreni E, Castellani S, Palmieri L, Penzo M, Conese M. Involvement of glycosaminoglycans in vesicular stomatitis virus g glycoprotein pseudotyped lentiviral vector-mediated gene transfer into airway epithelial cells. J Gene Med. 2008;10:1294–1302. doi: 10.1002/jgm.1248. [DOI] [PubMed] [Google Scholar]
- 102.Leistner CM, Gruen-Bernhard S, Glebe D. Role of glycosaminoglycans for binding and infection of hepatitis b virus. Cell Microbiol. 2008;10:122–133. doi: 10.1111/j.1462-5822.2007.01023.x. [DOI] [PubMed] [Google Scholar]
- 103.Spear PG. Herpes simplex virus: Receptors and ligands for cell entry. Cell Microbiol. 2004;6:401–410. doi: 10.1111/j.1462-5822.2004.00389.x. [DOI] [PubMed] [Google Scholar]
- 104.Sapp M, Bienkowska-Haba M. Viral entry mechanisms: Human papillomavirus and a long journey from extracellular matrix to the nucleus. FEBS J. 2009;276:7206–7216. doi: 10.1111/j.1742-4658.2009.07400.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Akhtar J, Shukla D. Viral entry mechanisms: Cellular and viral mediators of herpes simplex virus entry. FEBS J. 2009;276:7228–7236. doi: 10.1111/j.1742-4658.2009.07402.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Cocquerel L, Voisset C, Dubuisson J. Hepatitis c virus entry: Potential receptors and their biological functions. J Gen Virol. 2006;87:1075–1084. doi: 10.1099/vir.0.81646-0. [DOI] [PubMed] [Google Scholar]
- 107.Schulze A, Gripon P, Urban S. Hepatitis b virus infection initiates with a large surface protein–dependent binding to heparan sulfate proteoglycans. Hepatology. 2007;46:1759–1768. doi: 10.1002/hep.21896. [DOI] [PubMed] [Google Scholar]
- 108.Belting M. Heparan sulfate proteoglycan as a plasma membrane carrier. Trends Biocheml Sci. 2003;28:145–151. doi: 10.1016/S0968-0004(03)00031-8. [DOI] [PubMed] [Google Scholar]
- 109.Burckhardt CJ, Greber UF. Virus movements on the plasma membrane support infection and transmission between cells. PLoS Pathog. 2009;5:e1000621. doi: 10.1371/journal.ppat.1000621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Selinka HC, Giroglou T, Sapp M. Analysis of the infectious entry pathway of human papillomavirus type 33 pseudovirions. Virology. 2002;299:279–287. doi: 10.1006/viro.2001.1493. [DOI] [PubMed] [Google Scholar]
- 111.Beleid R, Douglas D, Kneteman N, Kaur K. Helical peptides derived from lactoferrin bind hepatitis c virus envelope protein e2. Chem BiolDrugDes. 2008;72:436–443. doi: 10.1111/j.1747-0285.2008.00715.x. [DOI] [PubMed] [Google Scholar]
- 112.Gifford JL, Ishida H, Vogel HJ. Structural characterization of the interaction of human lactoferrin with calmodulin. PLoS One. 2012;7:e51026. doi: 10.1371/journal.pone.0051026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Lin Q, Fang D, Hou X, Le Y, Fang J, Wen F, et al. Hcv peptide (c5a), an amphipathic α -helical peptide of hepatitis virus c, is an activator of n-formyl peptide receptor in human phagocytes. J Immunol. 2011;186;:2087–2094. doi: 10.4049/jimmunol.1002340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Zhang J, Mulvenon A, Makarov E, Wagoner J, Knibbe J, Kim JO, et al. Antiviral peptide nanocomplexes as a potential therapeutic modality for hiv/hcv co-infection. Biomaterials. 2013;34:3846–3857. doi: 10.1016/j.biomaterials.2013.01.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Martin F, Volpari C, Steinkuhler C, Dimasi N, Brunetti M, Biasiol G, et al. Affinity selection of a camelized v (h) domain antibody inhibitor of hepatitis c virus ns3 protease. Protein Engin. 1997;10:607–614. doi: 10.1093/protein/10.5.607. [DOI] [PubMed] [Google Scholar]
- 116.Lauwereys M, Ghahroudi MA, Desmyter A, Kinne J, Hölzer W, De Genst E, et al. Potent enzyme inhibitors derived from dromedary heavy-chain antibodies. EMBO J. 1998;17:3512–3520. doi: 10.1093/emboj/17.13.3512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Davies MJ, Hawkins CL, Pattison DI, Rees MD. Mammalian heme peroxidases: From molecular mechanisms to health implications. AntioxidRedoxSignal. 2008;10:1199–1234. doi: 10.1089/ars.2007.1927. [DOI] [PubMed] [Google Scholar]
- 118.Redwan EM, Almehdar HA, EL-Fakharany EM, Baig AWK, Uversky VN. Potential antiviral activities of camel, bovine, and human lactoperoxidases against hepatitis c virus genotype 4. RSC Adv. 2015;5:60441–60452. [Google Scholar]
- 119.Courtois P, Van Beers D, De Foor M, Mandelbaum I, Pourtois M. Abolition of herpes simplex cytopathic effect after treatment with peroxidase generated hypothiocyanite. J BiolBuccale. 1990;18:71–74. [PubMed] [Google Scholar]
- 120.Mikola H, Waris M, Tenovuo J. Inhibition of herpes simplex virus type 1, respiratory syncytial virus and echovirus type 11 by peroxidase-generated hypothiocyanite. Antiviral Res. 1995;26:161–171. doi: 10.1016/0166-3542(94)00073-h. [DOI] [PubMed] [Google Scholar]
- 121.Yamaguchi Y, Semmel M, Stanislawski L, Strosberg A, Stanislawski M. Virucidal effects of glucose oxidase and peroxidase or their protein conjugates on human immunodeficiency virus type 1. Antimicrob Agents Chemother. 1993;37:26–31. doi: 10.1128/aac.37.1.26. [DOI] [PMC free article] [PubMed] [Google Scholar]