

Abstract
Overt similarities exist between the effects of systemic cannabinoid CB1 inverse agonists and dopamine (DA) antagonists on appetitive behavior. The present set of studies was undertaken to apply a fine-grained analysis of food-reinforced operant lever pressing in rats in order to compare the pattern of effects produced by administration of the CB1 inverse agonist AM 251 and those induced by the DA D1 antagonist SKF 83566, and the D2 antagonist raclopride. Three groups of rats were trained on a fixed-ratio 5 (FR5) schedule and administered these compounds over a range of doses expected to suppress responding. All three drugs produced a dose-related suppression of total lever pressing. In addition to main effects of dose, regression analyses were performed to determine which of several response timing- and rate-related variables correlated most strongly with overall responding in each group. It was found that total session time spent pausing from responding was significantly better at predicting responding in the AM 251 group, while both DA antagonists produced significantly stronger regression coefficients (vs. AM 251) from fast responding measures. These results suggest that, while several similarities exist, CB1, D1, and D2 antagonists are not identical in their pattern of suppression of food-maintained lever pressing.
Keywords: cannabinoid, motivation, operant conditioning, fixed ratio, rimonabant appetite, microanalysis
Drugs from numerous pharmacological classes are known to suppress food-motivated behavior. For example, cannabinoid CB1 receptor antagonists and inverse agonists and dopamine (DA) D1 and D2 receptor antagonists have been shown to reduce food intake and food-reinforced operant responding when administered to rats systemically (Arnone et al., 1997; Barbano et al., 2009; Colombo et al., 1998; McLaughlin et al., 2003, 2005a, 2006; Pério et al., 2001; Rusk and Cooper, 1994; Salamone et al., 2002; Trevitt et al., 1998). Fixed-ratio (FR) operant schedules are often employed to assess the effects of food-reinforced operant behavior. FR schedules typically generate high response rates (Carriero et al., 1998; Killeen, 1994; Niv et al., 2007; Salamone et al., 2002) and are sensitive to the effects of several drug classes. Recently, CB1 inverse agonists such as rimonabant (SR 141716A; Rinaldi-Carmona et al., 1994), AM 251 (Gatley et al., 1996), and AM 1387 (McLaughlin et al., 2006) have been shown to reduce food-maintained operant responding, as defined by both reductions in numbers of responses on FR schedules (Freedland et al., 2000, 2003; McLaughlin et al., 2003, 2006; Sink et al., 2008) and also by lower break points in progressive ratio designs (Ward and Dykstra, 2005). As CB1 inverse agonists suppress lever pressing for food over a dose range similar to that which inhibits food intake (McLaughlin et al., 2003, 2006), it has been suggested that this class of compounds reduces operant responding by influencing some aspect of appetitive behavior toward the food stimulus. This is supported by a detailed analysis of lick rate, an unlearned motor response to food stimuli, in which 1.0 and 3.0 mg/kg SR 141716A decreased number of licks while preserving number of feeding bouts, which suggested that decreased motivation for food may be one of the effects of CB1 inverse agonism (Higgs et al., 2003). Similarly, both D1 and D2 DA receptor antagonists have been found to reduce number of operant responses (Fowler and Liao, 1998; Rick et al., 2006; Varvel et al., 2002) and to lower break points in progressive ratio schedules (Mobini et al., 2000). It has been suggested that CB1 inverse agonists may exert their effects on food motivated behavior via actions on dopaminergic systems (Tanda et al., 1997; Melis et al., 2007), which is consistent with reports of dense CB1 expression throughout striatum (Herkenham et al., 1990). However, several differences have been established between the actions of AM 251 and dopaminergic manipulations on food intake and food-maintained operant responding. First, while systemic DA antagonism and DA depletion of either whole forebrain or ventrolateral striatum inhibited food intake by reducing feeding rate and impairing food-handling behavior (Salamone et al., 1990, 1993a), systemic anorectic doses of the CB1 inverse agonist AM 251 did not produce deficits in food-handling or feeding efficiency (McLaughlin et al., 2005a). More recently, it was shown that compensatory increases in chow intake were not observed alongside reductions in FR5 lever-pressing induced by AM 251 (Sink et al., 2008), as has been shown for both D1 and D2 antagonist administration (Farrar et al., 2007; Salamone et al., 1991, 1996, 2002; Sink et al., 2008), and local antagonism of nucleus accumbens DA receptors (Nowend et al., 2001).
Such differentiation between the effects of cannabinergic and dopaminergic manipulations suggests that a fine-grained analysis of operant responding may yield response patterns that are characteristic of CB1 inverse agonists, D1 antagonists, and D2 antagonists, and can be used to further dissociate the effects of these drugs on food-maintained responding. In FR schedules, rate-decreasing manipulations can be analyzed for effects on the distribution of times (IRTs). Analysis of the distribution of IRTs has become a useful tool for assessing the effects of drugs or brain manipulations on operant behavior (Salamone et al., 1993b, 1995; Carriero et al., 1998; McLaughlin et al., 2005b; Mingote et al., 2005, 2008). Fast IRTs (i.e., smaller IRT values), which characterize bursts of responding, can be contrasted with pauses during the response session (i.e., disengaging the lever for more than a few seconds), and analysis of these components has been used to differentiate effects of response-suppressing manipulations. For instance, on a continuous reinforcement (CRF) schedule, 6-OHDA depletions of nucleus accumbens reduced responding by decreasing the percentage of responses with IRTs less than 1.5 s, and those between 3.0 and 4.5 s, while increasing pauses in responding, defined as IRTs > 12.0 s. In contrast, while reductions in responding produced by extinction were also characterized by increases in pauses, extinction tended to increase, rather than decrease, the proportion of fast responses (Salamone et al., 1995). This indicates that these manipulations do not produce the same effects on food-reinforced behavior. DA depletions in nucleus accumbens can also be distinguished from the effects of satiation via prefeeding, which reduced responding not by altering the IRT distribution, but rather by increasing total session time spent in pauses (Salamone et al. 1999).
Furthermore, regression analyses have been employed to assess the relation between different measures related to the IRT distribution and the overall number of responses. For instance, it was found that the reduction in FR30 responding seen with increasing doses of the opioid agonist alfentanil correlated with both time spent in pauses and time spent in response runs, while that seen with cocaine and ketamine were more strongly associated with changes in time spent in response runs (Skjoldager et al., 1991). Linear regression can therefore be used to evaluate the extent to which overall responding is associated with, and possibly mediated by, timing- and rate-related variables, in addition to simply identifying those variables that are affected at response-suppressing doses. In the present study, separate groups of rats trained on a food-reinforced FR5 schedule were given systemic dose regimens of the CB1 inverse agonist AM 251, the D1 antagonist SKF 83566, and the D2 antagonist raclopride. The FR5 schedule was chosen as the ratio parameter because it reliably produces both dense reinforcement (approx. 13 – 18 g of pellets per session) and high responses rates, allowing for extensive data to be gathered. It was predicted that, while all three drugs would inhibit food-maintained responding, differences between them would be evident in terms of IRT-related measures. As DA depletion of the nucleus accumbens reduced responses with faster IRT values (Salamone et al., 1993b, 1995), it is likely that the reduction in responding predicted in the SKF 83566 and raclopride groups would be more related to slowing of faster responses. In contrast, AM 251 was predicted to reduce responding by decreasing motivation to respond; therefore, the reduction in responding predicted for AM 251 may be more related to more frequent or longer pauses (during which time the animal may have disengaged the lever), rather than slowing fast responses. A portion of the data set (i.e., total number of session responses) generated by the group administered AM 251 was reported previously (McLaughlin et al., 2003), although in that publication, no data from the microanalysis was reported.
Methods
Subjects
A total of 24 adult male Sprague-Dawley rats (Harlan, Indianapolis, IN) weighing approximately 280-330 g on arrival were used. Rats were housed in pairs, and all studies were conducted in the light portion of the 12h light-dark cycle (lights on 0800-2000h). Approximately a week after arrival, rats were food-restricted by removing access to chow in the home cage. This required them to earn all of their daily food allotment during daily experimental sessions, except on weekends, when they were allowed to feed ad libitum. Rats were weighed daily and maintained at or above 85% of free-feeding body weight. Animals with weights under 85% of free-feeding weight were fed lab chow supplementally until their 85% weight was reattained. As animals were allowed to gain weight during the study, no animals required supplemental feeding in this manner once injection sessions had begun. Water was freely available in the home cage throughout the experiments. Animal protocols were approved by the University of Connecticut's Institutional Animal Care and Use Committee, and the methods were in accord with the Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, 1996).
Drugs
AM 251 (supplied by the Makriyannis lab) was dissolved in DMSO and Tween-80 (both Fisher, St. Louis, MO), and then the solution was brought to volume with artificial cerebrospinal fluid (aCSF), in a final ratio of 1:2:7. This solution also was used as the vehicle control treatment. SKF 83566 and raclopride (both Sigma, St. Louis, MO) were dissolved in 0.3% tartaric acid, which served as the vehicle control. See Procedures section for specific doses of each compound. Doses for each were selected based on pilot studies (data not shown). All injections were via the i.p. route, and in a volume of 1.0 ml/kg.
Procedures
All experiments were carried out in operant chambers (Med Associates, St. Albans, VT) controlled by IBM-compatible computers running custom software written in QBasic. These programs controlled reinforcement contingencies and also recorded time and number of lever responses (see Measures section). Following food restriction, subjects were exposed to approximately one gram of operant pellets in the home cage to reduce neophobia. Operant sessions were carried out 5 days per week, and all sessions were 30 min in length. Subjects were first given one magazine training session in the operant chambers in which pellets were delivered every 30 s, and each lever response was reinforced with one pellet. Beginning the following day, subjects were placed on a CRF schedule in which each response was reinforced. This schedule continued until subjects were reliably making greater than 300 responses in a single session. At this point, they were moved to an FR5 schedule, in which every fifth response was reinforced with a single pellet. Injection sessions commenced when performance stabilized; that is, when no subject had three consecutive days of increasing responses. Once baseline performance was attained, overall daily responding was monitored but allowed to vary freely.
Experiments
For all experiments, injections were administered prior to one operant session per week, and data (not shown) from subsequent sessions were analyzed to ensure that there were no carryover effects. All doses were administered in a counterbalanced, within-subjects design. In Experiment 1, subjects (n=10) were administered AM 251, a cannabinoid CB1 inverse agonist, in doses of 0.5, 1.0, 2.0, 4.0, and 8.0 mg/kg in addition to vehicle control. In Experiment 2, the DA D1 receptor antagonist SKF 83566 was administered in doses of 0.01875, 0.0375, 0.075, 0.15, 0.3, and 0.6 mg/kg, plus vehicle, to (n=7) rats. In Experiment 3, subjects (n=7) were administered vehicle, and doses of 0.05, 0.1, 0.2, and 0.4 mg/kg of the DA D2 antagonist raclopride. AM 251 and SKF 83566 were administered 30 min prior to testing, and raclopride was injected 20 min prior to testing.
Measures
As is typical in this type of design, the overall number of responses within the session was recorded. Total pause time (TPT; in s) was defined as the sum of the length of all pauses in each session. Average pause length was calculated as TPT divided by the raw number of pauses. In addition, the session IRT distribution was divided into several variables related to the timing and rate of lever pressing, as well as pauses in responding. As described previously (McLaughlin et al., 2005b), IRTs were placed into 250 ms time bins. 20 IRT bins represented all IRTs which were 5 s in length or shorter (20 bins × 250 ms per bin = 5 s), and one additional bin (Bin 21) represented pauses, defined as IRTs >5 s in length. During baseline (non-injected) sessions, the majority of IRTs were found to be within 500 ms, which corresponded to the first two IRT bins. Because of this, where significant dose X bin interactions were found, three bins were selected a priori for analysis of simple main effects of dose. In addition to the pause bin (Bin 21), Bin 1 (IRTs <= 250 ms) and Bin 2 (250 ms < IRTs <= 500 ms) were expressed as a percentage of total session IRTs and analyzed as well. Other bins (i.e., Bins 3-20) were not analyzed separately because responding within these bins is typically less than 4% per bin across dose conditions.
Statistical Analyses
Mean responses, TPT, and average pause length were analyzed for dose effects using repeated measures ANOVA with dose as a within-subjects factor. Changes in the overall IRT distribution were analyzed by entering the 21 bins as a second within-subjects factor in a dose X IRT bin ANOVA. As IRT bins are expressed as a percentage of all responding (and therefore sum to 100% at each dose), main effects of dose were not predicted for this measure; however, an interaction was interpreted as evidence that the drug altered the overall distribution of responses. Where significant dose X bin interactions were found, simple main effects of dose were analyzed via repeated-measures ANOVA for Bin 1, Bin 2, and Bin 21 (the pause bin). Separate analyses were performed for each experiment. For the dose analyses of the variables above, individual dose effects were analyzed using non-orthogonal planned comparisons (Keppel and Wickens, 2004) in which data from each dose were compared to those from its own vehicle. Regression analyses were further performed with all data points analyzed regardless of dose. TPT, Bin 1, Bin 2, and percentage (i.e., Bin 21) and length of pauses were each analyzed with a separate equation with overall responding as the dependent variable. The regression slopes were taken as an indication of the strength of the relationship between each IRT measure, and responding (the predicted variable). Regression slopes can be compared for significant differences using ANOVA, rather than “eyeballing” the different slope values across groups (Raudenbush et al., 1997). This ANOVA was performed on a multiple regression equation for which four new variables were created for each analysis. First, two dummy variables were created that were coded by group: in each, one drug group was arbitrarily selected (AM 251 for the first variable and SKF 83566 for the second) and assigned a value of 1, and a value of 0 was assigned to both of the other groups. Then, two variables were found from the product of the IV and the dummy-coded variable. Thus, each of these two variables contained values identical to the IV for AM 251 and SKF 83566, respectively, and values of 0 for all other cells. In the multiple regression analysis, the IV and both dummy variables were entered simultaneously. The last two variables described, containing the products of the IV and each dummy variable, were then entered. The incremental F ratio was found; this analysis determines whether more variance is explained by the equation when a new set of predictors is added. By analyzing whether a set of variables weighted by drug group contribute more variability than the IRT variable alone, we tested the null hypothesis that the regression slopes were the same across drug group (Raudenbush et al., 1997). Where significant overall effects were found, post-hoc analyses were conducted by employing a similar procedure using pairwise comparisons of drugs. Sessions with fewer than 10 responses were excluded from regression analysis.
Results
Dose Analyses
Experiment 1 (AM 251) – As seen in Table 1, the CB1 inverse agonist AM 251 suppressed overall responding (F(5,45) = 17.44, p < .001). Total pause time (TPT; i.e., total time not engaging the lever for more than 5 s) was dose-dependently increased (F(5,45) = 11.66, p < .001), but average length of pauses was not altered by AM 251 (F(5,45) = 1.42, ns). AM 251 altered overall response rate, as revealed by a significant dose X IRT bin interaction (F(105, 945) = 4.63, p < .001). Bin 1 (i.e., all IRTs <= 250 ms; F(5,45) = 6.05, p < .001) decreased significantly, but Bin 2 did not (F(5,45) = 1.76, ns). Bin 21, containing all IRTs that were pauses (i.e., > 5 s) increased significantly F(5,45) = 18.82, p < .001). Planned comparisons showed that the 2.0, 4.0, and 8.0 mg/kg doses reduced overall responding as well as Bin 1 responding, while increasing TPT and Bin 21, relative to vehicle.
Table 1.
Summary of dose effects on operant responding for AM 251-treated subjects. See text for specific analyses. Values represent means (±S.E.M.).
| Dose AM 251 (mg/kg) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Measure | Vehicle | 0.5 | 1.0 | 2.0 | 4.0 | 8.0 | Regression Equation (DV = Response) | ||
| Overall Responses | 1508.2 (117.2) |
1353.7 (93.1) |
1250.6 (131.7) |
931.7 (100.7)** |
712.8 (73.5)** |
454.1 (68.7)** |
Slope | Y-intercept | R2 |
| Total Pause Time (s) | 505.1 (92.0) |
649.1 (91.3) |
710.2 (98.6) |
877.0 (76.1)** |
1062.2 (74.5)** |
1248.8 (83.4)** |
− 1.26 †† | 2093.3 | .929 |
| Average Pause Length (s) | 23.92 (6.46) |
25.1 (5.70) |
24.1 (4.05) |
26.81 (3.19) |
29.2 (4.44) |
36.9 (5.31) |
−19.7 †† | 1579.6 | .413 |
| Bin 1 (% all IRTs) | 31.18 (3.97) |
30.93 (3.66) |
29.48 (4.14) |
23.63 (3.94)* |
22.2 (2.66)** |
19.6 (3.69)** |
16.6 †† | 601.3 | .175 |
| Bin 2 (% all IRTs) | 29.71 (3.17) |
29.69 (2.58) |
30.88 (3.36) |
28.6 (3.29) |
30.0 (2.62) |
25.68 (3.02) |
9.27 | 766.2 | .032 |
| Pauses (% all IRTs) | 1.99 (.28) |
2.43 (0.24) |
3.39 (0.79) |
4.46 (0.64)* |
6.56 (0.81)** |
9.72 (1.06)** |
−114.8 †† | 1582.4 | .670 |
p < .05,
p < .01 versus vehicle via planned comparisons. Values in the regression equation predicting responding from each measure are also presented.
Slope of regression equation is significantly different from zero, p < .01.
Experiment 2 (SKF 83566) – Overall responding was significantly decreased (F(6,36) = 24.01, p < .001). TPT also increased (F(6,36) = 18.71, p < .001), as did the average length of pauses (F(6,36) = 10.22, p < .001). The IRT bin distribution was altered, as evidenced by a dose X IRT bin interaction (F(126,756) = 6.32, p < .001). Proportion of responses in IRT Bin 1 (F(6,36) = 12.08, p < .001) and Bin 2 decreased (F(6,36) = 14.17, p < .001), while those in Bin 21 increased (F(6,36) = 7.98, p < .001). The 0.075, 0.15, 0.3, and 0.6 mg/kg doses were significantly lower than vehicle performance according to planned comparisons. Furthermore, responses were fewer, while TPT was greater, at the 0.075, 0.15, 0.3, and 0.6 mg/kg doses relative to vehicle. Only the 0.15, 0.3, and 0.6 mg/kg doses altered the proportion of responses in Bin 1. Lastly, percentage and length of pauses, as well as proportion of responses in Bin 2 were altered by the 0.3 and 0.6 mg/kg doses relative to vehicle. These effects are seen in Table 2.
Table 2.
Values represent means (±S.E.M.) for each dose of the D1 antagonist SKF 83566 on measures of operant responding. See text for specific analyses.
| Dose SKF 83566 (mg/kg) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Measure | Vehicle | 0.01875 | 0.0375 | 0.075 | 0.15 | 0.3 | 0.6 | Regression Equation (DV = Response) | ||
| Overall Responses | 1359.6 (125.8) |
1459.1 (134.1) |
1283.4 (50.5) |
961.4 (172.7)* |
656.7 (209.5)** |
218.9 (173.2)** |
7.29 (3.00)** |
Slope | Y-intercept | R2 |
| Total Pause Time (s) | 463.0 (88.3) |
377.5 (70.8) |
556.4 (77.2) |
862.9 (142.67)* |
1025.3 (215.5)** |
1536.0 (197.6)** |
1784.7 (8.49)** |
−1.04 †† | 1828.3 | .909 |
| Average Pause Length (s) | 10.96 (2.28) |
8.18 (1.40) |
14.03 (4.36) |
29.76 (15.16) |
142.6 (123.4) |
422.7 (236.1)* |
1189.4 (232.4)** |
−13.9 †† | 1387.7 | .466 |
| Bin 1 (% all IRTs) | 24.36 (2.15) |
23.19 (2.62) |
29.04 (4.79) |
23.71 (6.00) |
11.24 (1.58)** |
4.99 (1.87)** |
2.24 (1.61)** |
31.7 †† | 363.7 | .433 |
| Bin 2 (% all IRTs) | 33.43 (2.15) |
35.06 (1.79) |
34.33 (3.16) |
30.25 (3.53) |
27.5 (3.56) |
15.4 (4.34)** |
4.47 (3.22)** |
30.6 †† | 86.2 | .308 |
| Pauses (% all IRTs) | 4.11 (0.81) |
4.05 (1.04) |
4.51 (1.00) |
7.42 (1.74) |
12.0 (3.07) |
37.0 (11.6)** |
57.45 (13.9)** |
−86.8 †† | 1683.7 | .796 |
p < .05,
p < .01 versus vehicle via planned comparisons. Constants of the linear regression equation predicting responding are also presented for each measure.
Slope of regression equation is significantly different from zero, p < .01. Slope and Y-intercept for Pauses excludes sessions with >15%; those for Pause Length exclude values >60s.
Experiment 3 (Raclopride) – As in Experiments 1 and 2, responding was suppressed (F(4,24) = 11.57, p < .001). As seen in Table 3, TPT (F(4,20) = 10.09, p < .001), and average length of pauses (F(4,24) = 7.04, p < .001) were dose-dependently increased. The IRT bin distribution was altered (dose X bin interaction: F(84,504) = 6.64, p < .001). Raclopride also reduced the proportion of IRTs in Bin 1 (F(4,24) = 11.49, p < .001) and Bin 2 (F(4,24) = 18.98, p < .001). Proportion of IRTs in Bin 21 was increased (F(4,24) = 10.70, p < .001). Planned comparisons revealed that responding (as well as Bin 1) was significantly decreased at the 0.2 and 0.4 mg/kg doses. TPT, average length and percentage of pauses (i.e., Bin 21) and Bin 2 were significantly different from vehicle only at the 0.4 mg/kg dose.
Table 3.
Effects of the D2 antagonist raclopride on measures of operant responding, as well as constants of the regression equation with responding entered as the DV. See text for specific analyses. Values represent means (±S.E.M.).
| Dose Raclopride | ||||||||
|---|---|---|---|---|---|---|---|---|
| Measure | Vehicle | 0.05 | 0.1 | 0.2 | 0.4 | Regression Equation (DV = Responses) | ||
| Overall Responses | 1140.1 (63.9) |
1258.0 (88.5) |
1035.0 (144.9) |
724.4 (176.5)* |
172.9 (156.4)** |
Slope | Y- Intercept | R2 |
| Total Pause Time (s) | 637.3 (82.8) |
485.9 (83.9) |
685.7 (144.7) |
936.1 (177.1) |
1579.0 (184.7)** |
−.95 †† | 1687.3 | .881 |
| Average Pause Length (s) | 16.9 (4.31) |
12.2 (3.88) |
16.4 (4.75) |
28.3 (9.30) |
883.9 (326.5)** |
−18.3 †† | 1363.1 | .506 |
| Bin 1 (% all IRTs) | 27.2 (2.75) |
24.9 (2.80) |
20.9 (1.94) |
17.6 (3.77)* |
3.27 (3.02)** |
34.8 †† | 239.7 | .573 |
| Bin 2 (% all IRTs) | 33.6 (2.29) |
34.75 (1.85) |
34.0 (1.39) |
25.5 (4.27) |
5.42 (3.62)** |
31.7 †† | 25.5 | .510 |
| Pauses (% all IRTs) | 4.18 (0.44) |
4.35 (0.71) |
7.03 (2.41) |
13.5 (4.24) |
51.0 (13.3)** |
−51.6 †† | 1400.6 | .524 |
p < .05,
p < .01 versus vehicle via planned comparisons.
Slope of regression equation is significantly different from zero, p < .01. Slope and Y-intercept for Pauses excludes sessions with >15%; those for Pause Length exclude values >60s.
Regression analyses: comparisons of all groups
For all drug experiments, slopes of all equations predicting responding were significant (Tables 1-3; p's < .05), with the exception of Bin 2, which was significant for raclopride and SKF 83566, but not AM 251. To account for nonlinear relationships, sessions with more than 15 percent pauses and average pause lengths greater than 60 s (both values approximated the maximum obtained in AM 251-treated rats) were removed from their respective analyses. In all three experiments, it was found that substantial decreases in responding (i.e., sessions with fewer than 100 responses were often related to very long pause lengths and high percentages of pauses, producing data that violated assumptions of normality. Such sessions were found only in the SKF 83566 and raclopride studies; as has been found previously (McLaughlin et al., 2003; 2006), CB1 inverse agonists rarely suppress responding to such levels. While the potential to investigate nonlinear relationships was lost, this omission was necessary for a comparison using linear regression across groups.
Analysis of variance of regression coefficients revealed significant differences between the coefficients of the relationships between overall responding and TPT (Fig. 1; F(2,128) = 7.561, p < .001), Bin 21 (Fig 2., F(2,112) = 10.17, p < .001), and fast responses, including Bin 1 (Fig. 3; F(2,128) = 3.342, p < .05), and Bin 2 (Fig. 4; F(2,128) = 3.542, p < .05), across drug conditions. Post hoc analyses revealed that the regression coefficient for TPT was significantly greater in absolute value for the AM 251 group than for the SKF 83566 or raclopride groups (p's < .001; Fig. 1). As can be seen in Figure 2, the slope of predicted responding as a function of pause number (i.e., Bin 21) was significantly lower for raclopride than for either of the two other drugs, which were not different from one another. In contrast, for both Bin 1 (Fig. 3) and Bin 2 (Fig. 4), the AM 251 group produced a significantly lower regression coefficient than either of the other two groups, which did not differ from each other.
Figure 1.
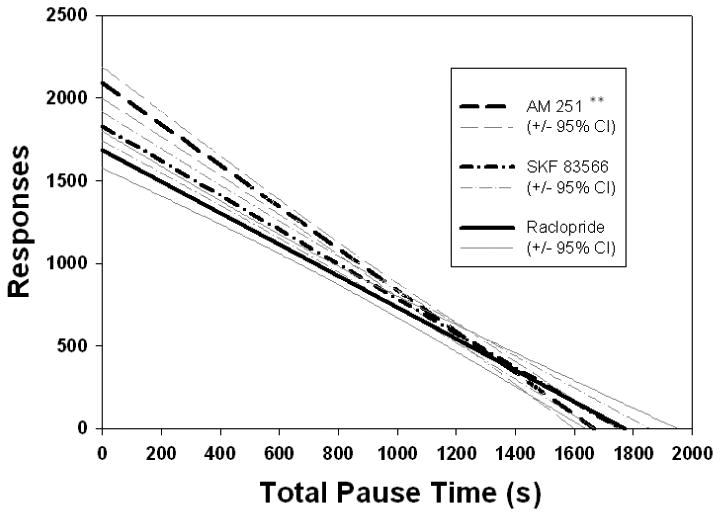
Linear regression plots between total pause time (TPT) and overall responding. The slope produced by AM 251 was significantly more negative than those produced by either DA antagonist, which did not differ from one another. **p < .01 difference between AM 251 and both SKF 83566 and raclopride. Regression equations: AM 251: Y = 2093.3 – 1.26 X. SKF 83566: Y = 1828.3 – 1.04 X. Raclopride: Y = 1687.3 – 0.952 X.
Figure 2.
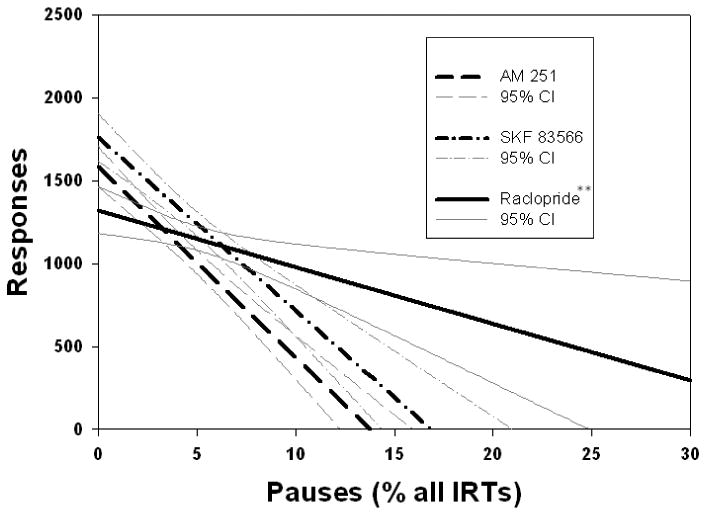
Least-squares regression lines predicting responding from the percent of IRTs which were pauses (IRTs > 5s). AM 251 and SKF 83566 produced a significantly stronger slope than raclopride. **p < .01 difference between raclopride and both SKF 83566 and AM 251. Regression equations: AM 251: Y = 1582.4 - 114.8 X. SKF 83566: Y = 1683.7 – 86.8 X. Raclopride: Y = 1400.6 – 51.6 X.
Figure 3.
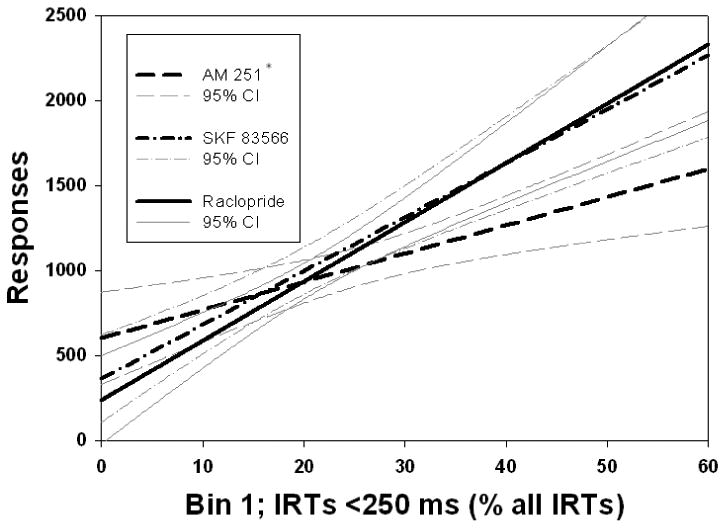
Linear regression predicting overall responding from Bin 1, the first, shortest IRT bin as a percent of all IRTs. AM 251 produced a significantly lower slope than either DA antagonist. *p < .05 difference between AM 251 and both SKF 83566 and raclopride. Regression equations: AM 251: Y = 601.3 + 16.61 X. SKF 83566: Y = 363.7 + 31.69 X. Raclopride: Y = 239.7 + 34.82 X.
Figure 4.
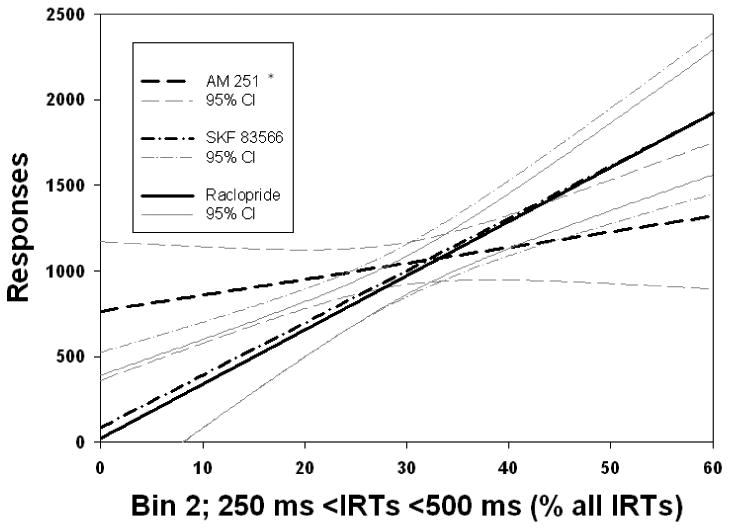
Lines of best fit predicting overall responding from the second-shortest IRT bin, expressed as a percent of all IRTs. As with the relationship between Bin 1 and responding, both DA antagonists were better at producing a relationship between Bin 2 and responding than was AM 251. *p < .05 difference between AM 251 and both SKF 83566 and raclopride. Regression equations: AM 251: Y = 766.2 + 9.27 X. SKF 83566: Y = 86.17 + 30.57 X. Raclopride: Y = 25.5 + 31.7 X.
Discussion
The cannabinoid CB1 inverse agonist AM 251, the DA D1 antagonist SKF 83566, and the D2 antagonist raclopride all dose-dependently suppressed lever pressing for food reinforcement. There were some general similarities between the effects of these drugs on operant responding. All three compounds increased total pause time (TPT), increased pauses (IRTs > 5 s) relative to other IRTs, and also decreased the relative number of IRTs in Bin 1 (fast responses, or IRTs <= 250 ms). Overall, responding was typically decreased at doses that altered these other parameters of lever pressing. These findings corroborate previous reports on the effects of AM 251 (McLaughlin et al., 2003; Sink et al., 2008) and DA antagonist administration (Fowler and Liao, 1998; Salamone et al., 2002; Trevitt et al., 1998; Varvel et al., 2002) on operant responding and response slowing. Possibly, there are certain core features of lever pressing behavior that tend to be affected in similar ways across multiple pharmacological manipulations. Furthermore, similarities could be expected based upon the fact that CB1, D1, and D2 receptors are expressed in many of the same central systems. Dense CB1 radiolabeling is found in striatal areas, including nucleus accumbens and neostriatum, as well as in substantia nigra pars reticulata and ventral pallidum (Herkenham et al., 1990), and in striatum is restricted to GABAergic medium spiny neurons (Herkenham et al., 1991) that also express DA receptors (Herkenham et al., 1991; Hermann et al., 2002). Likewise, spiny neurons in nucleus accumbens are also known to contain cannabinoid and DA receptors (Sesack & Pickel, 1990; Pickel et al., 2006). Moreover, systemic SR 141716A was recently found to increase DA outflow in prefrontal cortex and nucleus accumbens at doses that also inhibited food intake (Need et al., 2006; Melis et al., 2007). Taken together, as CB1 and DA receptors are densely expressed in areas believed to be crucial for sensorimotor integration and regulation of food intake, it is reasonable to expect that dopaminergic and cannabinergic manipulations could share many overt similarities with regard to effects upon food-reinforced behavior. It should also be noted that a wide dose range was chosen that may permit the influence of motivational and motoric processes, and that the dose range of AM 251 is similar to that used previously by this group and others (McLaughlin et al., 2003, 2005a; Xi et al., 2006; Sink et al., 2008; Merroun et al., 2009).
As all three compounds substantially reduced responding at high doses, it is not surprising that measures related to timing and rate of responding appear to have been affected similarly in each experiment. Nevertheless, it should be emphasized that there also were some differences between the effects of AM 251 and the DA antagonists. For example, both DA antagonists (but not AM 251) decreased responding in Bin 2 (250 ms < IRTs <= 500 ms), and also decreased the average length of pauses. Furthermore, linear regression analysis was used to predict responding from each timing and rate measure, and the regression coefficients were then compared among the different compounds (Raudenbush et al., 1997). It was found that the regression coefficients for total pause time were significantly stronger for the AM 251 group than for the SKF 83566 group and the raclopride group, even though both DA antagonists produced more pauses. Raclopride was also distinguished from the two other compounds; in raclopride-treated rats, there was a relation between pause number and overall responding that was significantly weaker than that seen with AM 251 or SKF 83566. On the other hand, ANOVAs of regression coefficients also indicated that, unlike AM 251, both SKF 83566 and raclopride produced a pattern of overall response suppression more associated with slowing of Bins 1 and 2, together representing IRTs < 500 ms (Figs. 3 and 4). These results may suggest that AM 251 mediates responding more by decreasing time engaged with the lever, while the effects of SKF 83566 and especially raclopride were more strongly related to a relative reduction in fast responding. Disengaging the lever may indicate increasing satiety, as has been found by increasing satiety via prefeeding (Salamone et al. 1999; Lowe et al., 1974; Powell, 1969), although the CB1 agonist AM 411 has also been found to increase total pause time (McLaughlin et al., 2005a). Although AM 251 significantly reduced the number of fast responses, regression analysis revealed that this effect was less strongly associated with overall responding in the AM 251-treated animals than in those treated with SKF 83566 or raclopride. Of the three compounds tested, the pattern of responding produced by AM 251 may therefore be most similar to that of prefeeding, that is, reducing food-maintained responding by allowing access to food prior to the session. Prefeeding has been found to increase pausing without altering fast responses (Salamone et al. 1999; Lowe et al., 1974; Powell, 1969). However, there also may be substantial differences between the effects of CB1 receptor blockade and prefeeding. AM 251 and SR 141716A, at doses at or below those that reduce food intake, have been shown to produce nonmotivational effects that potentially interfere with food intake, such as nausea (McLaughlin et al., 2005a) and biting and scratching (Hodge et al., 2008; Tallett et al., 2007, 2008). Similar to the suppression of fast responding in the present study, these effects, while relevant to the therapeutic potential of CB1 blockade as an antiobesity treatment, are not likely the main cause of the anorectic effect (Chambers et al., 2006; Hodge et al., 2008; Sink et al., 2008).
The results reported in the present studies support previous findings suggesting that the effects of CB1 inverse agonists can be distinguished from those of dopaminergic manipulations. Recently, the effects of AM 251, as well as the CB1 neutral antagonist AM 4113, were compared with those of the D1 antagonist SCH 39166 (ecopipam) and the D2 antagonist eticlopride on a concurrent lever pressing/chow feeding task (Sink et al., 2008). With this task, rats are given the choice between lever pressing on a FR5 schedule for a preferred food (high carbohydrate operant pellets) versus simply approaching and feeding upon a less preferred food (laboratory chow) that is concurrently available in the chamber. Consistent with other studies (Nowend et al., 2001; Salamone et al., 1991, 2002), at low to moderate doses the DA D1 and D2 family antagonists suppressed lever pressing for the preferred food, but along with the drug-induced reductions in lever pressing, these rats showed a concomitant increase in consumption of the concurrently available chow (Sink et al., 2008). In contrast, interference with CB1 transmission by injection of AM 251 (at doses that significantly suppressed responding in the present study) or AM 4113 suppressed FR5 responding, but without a concurrent increase in chow consumption (Sink et al., 2008). These effects of CB1 antagonism or inverse agonism, which differed substantially from those produced by DA antagonists, resembled effects reported previously for the serotonergic appetite suppressant fenfluramine (Salamone et al., 2002) and prefeeding (Salamone et al., 1991). The present results further indicate that the pattern of the operant response suppression itself can also be used to dissociate the effects of CB1 inverse agonism, and D1 as well as D2 blockade. It could be argued that differences may be due to differences in pharmacological action between AM 251, an inverse agonist, and both dopaminergic compounds, which are neutral antagonists. However, SKF 83566 and raclopride were distinguishable from one another, while the effects of AM 251 on food-motivated behavior are quite similar to those of CB1 antagonists such as AM 4113 (Hodge et al., 2008; Sink et al., 2008). In fact, it is possible that dopaminergic manipulations (particularly D2 antagonists) produce effects that are more similar to those of CB1 agonists, rather than CB1 inverse agonists or antagonists. Low doses of apomorphine activate inhibitory DA autoreceptors (thus reducing DA release), which produced similar effects to the CB1 agonist delta-9-THC on a microanalysis of operant responding in a task in which rats were required to respond with high (>50 g) force to obtain reinforcement (McLaughlin et al., 2000). Moreover, a detailed study of the tetrad of tasks used to screen CB1 agonists (Martin et al., 1991) revealed that typical antipsychotics produced effects that were more difficult to distinguish from CB1 agonists than several other tested drug classes (Wiley and Martin, 2003).
As differences appear to exist between the effects of CB1 and DA antagonists on food reinforced behavior, future studies should investigate the brain structures involved by direct local application of relevant compounds. Given the present results, as well as the pattern of expression of brain cannabinoid and DA receptors, it is possible that distinct sites of action for each class of compounds exist, as well as sites of convergence.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Arnone M, Maruani J, Chaperon F, Thiébot M-H, Poncelet M, Soubrié P, et al. Selective inhibition of sucrose and ethanol intake by SR 141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology (Berl) 1997;132(1):104–6. doi: 10.1007/s002130050326. [DOI] [PubMed] [Google Scholar]
- Barbano MF, Le Saux M, Cador M. Involvement of dopamine and opioids in the motivation to eat: Influence of palatability, homeostatic state, and behavioral paradigms. Psychopharmacology (Berl) 2009;203(3):475–87. doi: 10.1007/s00213-008-1390-6. [DOI] [PubMed] [Google Scholar]
- Carriero D, Aberman J, Lin SY, Hill A, Makriyannis A, Salamone JD. A detailed characterization of the effects of four cannabinoid agonists on operant lever pressing. Psychopharmacology (Berl) 1998;137(2):147–56. doi: 10.1007/s002130050604. [DOI] [PubMed] [Google Scholar]
- Chambers AP, Koopmans HS, Pittman QJ, Sharkey KA. AM 251 produces sustained reductions in food intake and body weight that are resistant to tolerance and conditioned taste aversion. Br J Pharmacol. 2006;147(1):109–16. doi: 10.1038/sj.bjp.0706439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colombo G, Agabio R, Diaz G, Lobina C, Reali R, Gessa GL. Appetite suppression and weight loss after the cannabinoid antagonist SR 141716. Life Sci. 1998;63(8):PL113–7. doi: 10.1016/s0024-3205(98)00322-1. [DOI] [PubMed] [Google Scholar]
- Farrar AM, Pereira M, Velasco F, Hockemeyer J, Muller CE, Salamone JD. Adenosine A(2A) receptor antagonism reverses the effects of dopamine receptor antagonism on instrumental output and effort-related choice in the rat: Implications for studies of psychomotor slowing. Psychopharmacology (Berl) 2007;191(3):579–86. doi: 10.1007/s00213-006-0554-5. [DOI] [PubMed] [Google Scholar]
- Freedland CS, Poston JS, Porrino LJ. Effects of SR141716A, a central cannabinoid receptor antagonist, on food-maintained responding. Pharmacol Biochem Behav. 2000;67(2):265–70. doi: 10.1016/s0091-3057(00)00359-2. [DOI] [PubMed] [Google Scholar]
- Freedland CS, Whitlow CT, Smith HR, Porrino LJ. Functional consequences of the acute administration of the cannabinoid receptor antagonist, SR141716A, in cannabinoid-naïve and – tolerant animals: A quantitative 2-[14C]deoxyglucose study. Brain Res. 2003;962(1-2):169–79. doi: 10.1016/s0006-8993(02)03999-9. [DOI] [PubMed] [Google Scholar]
- Fowler SC, Liao JR. Haloperidol, raclopride, and eticlopride induce microcatalepsy during operant performance in rats, but clozapine and SCH 23390 do not. Psychopharmacology (Berl) 1998;140(1):81–90. doi: 10.1007/s002130050742. [DOI] [PubMed] [Google Scholar]
- Gatley SJ, Gifford AN, Volkow ND, Lan R, Makriyannis A. 123I-labeled AM 251: A radioiodinated ligand which binds in vivo to mouse brain cannabinoid CB1 receptors. Eur J Pharmacol. 1996;307(3):331–8. doi: 10.1016/0014-2999(96)00279-8. [DOI] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa BR, et al. Cannabinoid receptor localization in brain. Proc Nat Acad Sci USA. 1990;87(5):1932–6. doi: 10.1073/pnas.87.5.1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, de Costa BR, Richfield EK. Neuronal localization of cannabinoid receptors in the basal ganglia of the rat. Brain Res. 1991;547(2):267–74. doi: 10.1016/0006-8993(91)90970-7. [DOI] [PubMed] [Google Scholar]
- Hermann H, Marsicano G, Lutz B. Coexpression of the cannabinoid receptor type 1 with dopamine and serotonin receptors in distinct neuronal subpopulations of the adult mouse forebrain. Neuroscience. 2002;109(3):451–60. doi: 10.1016/s0306-4522(01)00509-7. [DOI] [PubMed] [Google Scholar]
- Higgs S, Williams CM, Kirkham TC. Cannabinoid influences on palatability: Microstructural analysis of sucrose drinking after delta(9)-tetrahydrocannabinol, anandamide, 2-arachidonoyl glycerol and SR141716. Psychopharmacology (Berl) 2003;165(4):370–7. doi: 10.1007/s00213-002-1263-3. [DOI] [PubMed] [Google Scholar]
- Hodge J, Bow JP, Plyler KS, Vemuri VK, Wisniecki A, Salamone JD, et al. The cannabinoid CB1 receptor inverse agonist and antagonist AM 4113 produce similar effects on the behavioral satiety sequence in rats. Behav Brain Res. 2008;193(2):298–305. doi: 10.1016/j.bbr.2008.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Institute of Laboratory Animal Resources Guide for the Care and Use of Laboratory Animals. Washington DC: National Academy Press; 1996. [Google Scholar]
- Keppel G, Wickens TD. Design and Analysis: A researcher's Handbook. 4. Englewood Cliffs, NJ: Prentice-Hall; 2004. [Google Scholar]
- Killeen PR. Mathematical principles of reinforcement. Behav Brain Sci. 1994;17(1):105–72. [Google Scholar]
- Lowe CF, Davey GC, Harzem P. Effects of reinforcement magnitude on interval and ratio schedules. J Exp Anal Behav. 1974;22(3):553–60. doi: 10.1901/jeab.1974.22-553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin BR, Compton DR, Thomas BF, Prescott WR, Little PJ, Razdan RK, et al. Behavioral, biochemical, and molecular modeling evaluations of cannabinoid analogs. Pharmacol Biochem Behav. 1991;40(3):471–8. doi: 10.1016/0091-3057(91)90349-7. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Delevan CE, Carnicom S, Robinson JK, Brener J. Fine motor control in rats is disrupted by delta-9-tetrahydrocannabinol. Pharmacol Biochem Behav. 2000;66(4):803–9. doi: 10.1016/s0091-3057(00)00281-1. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Winston K, Swezey L, Wisniecki A, Aberman J, Tardif DJ, et al. The cannabinoid CB1 antagonists SR 141716A and AM 251 suppress food intake and food-reinforced behavior in a variety of tasks in rats. Behav Pharmacol. 2003;14(8):583–8. doi: 10.1097/00008877-200312000-00002. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Winston KM, Limebeer CL, Parker LA, Makriyannis A, Salamone JD. The cannabinoid CB1 antagonist AM 251 produces food avoidance and behaviors associated with nausea but does not impair feeding efficiency in rats. Psychopharmacology (Berl) 2005a;180(2):286–93. doi: 10.1007/s00213-005-2171-0. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Lu D, Winston KM, Thakur G, Swezey LA, Makriyannis A, et al. Behavioral effects of the novel cannabinoid full agonist AM 411. Pharmacol Biochem Behav. 2005b;81(1):78–88. doi: 10.1016/j.pbb.2005.02.005. [DOI] [PubMed] [Google Scholar]
- McLaughlin PJ, Qian L, Wood JT, Wisniecki A, Winston KM, Swezey LA, et al. Suppression of food intake and food-reinforced behavior produced by the novel CB1 receptor antagonist/inverse agonist AM 1387. Pharmacol Biochem Behav. 2006;83(3):396–402. doi: 10.1016/j.pbb.2006.02.022. [DOI] [PubMed] [Google Scholar]
- Melis T, Succu S, Sanna F, Boi A, Argiolas A, Melis MR. The cannabinoid antagonist SR 141716A (rimonabant) reduces the increase of extra-cellular dopamine release in the rat nucleus accumbens induced by a novel high palatable food. Neurosci Lett. 2007;419(3):231–5. doi: 10.1016/j.neulet.2007.04.012. [DOI] [PubMed] [Google Scholar]
- Mingote S, Weber SM, Ishiwari K, Correa M, Salamone JD. Ratio and time requirements on operant schedules: Effort-related effects of nucleus accumbens dopamine depletions. Eur J Neurosci. 2005;21(6):1749–57. doi: 10.1111/j.1460-9568.2005.03972.x. [DOI] [PubMed] [Google Scholar]
- Mingote S, Font L, Farrar AM, Vontell R, Worden LT, Stopper CM, et al. Nucleus accumbens adenosine A2A receptors regulate exertion of effort by acting on the ventral striatopallidal pathway. J Neurosci. 2008;28(36):9037–46. doi: 10.1523/JNEUROSCI.1525-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mobini S, Chiang TJ, Ho MY, Bradshaw CM, Szabade E. Comparison of the effects of clozapine, haloperidol, chlorpromazine, and d-amphetamine on performance on a time-constrained progressive ratio schedule and on locomotor behaviour in the rat. Psychopharmacology (Berl) 2000;152(1):47–54. doi: 10.1007/s002130000486. [DOI] [PubMed] [Google Scholar]
- Need AB, Davis RJ, Alexander-Chacko JT, Eastwood B, Chernet E, Phebus LA, et al. The relationship of in vivo central CB1 receptor occupancy to changes in cortical monoamine release and feeding elicited by CB1 receptor antagonists in rats. Psychopharmacology (Berl) 2006;184(1):26–35. doi: 10.1007/s00213-005-0234-x. [DOI] [PubMed] [Google Scholar]
- Niv Y, Daw ND, Joel D, Dayan P. Tonic dopamine: Opportunity costs and the control of response vigor. Psychopharmacology (Berl) 2007;191(3):507–20. doi: 10.1007/s00213-006-0502-4. [DOI] [PubMed] [Google Scholar]
- Nowend KL, Arizzi M, Carlson BB, Salamone JD. D1 or D2 antagonism in nucleus accumbens core or dorsomedial shell suppresses lever pressing for food but leads to compensatory increases in chow consumption. Pharmacol Biochem Behav. 2001;69(3-4):373–82. doi: 10.1016/s0091-3057(01)00524-x. [DOI] [PubMed] [Google Scholar]
- Pério A, Barnouin MC, Poncelet M, Soubrié P. Activity of SR141716 on post-reinforcement pauses in operant responding for sucrose reward in rats. Behav Pharmacol. 2001;12(8):641–5. doi: 10.1097/00008877-200112000-00009. [DOI] [PubMed] [Google Scholar]
- Powell RW. The effect of reinforcement magnitude upon responding under fixed-ratio schedules. J Exp Anal Behav. 1969;12(4):605–8. doi: 10.1901/jeab.1969.12-605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raudenbush SW, Gleser L, Hedges L, Johnson E, Petkova E. Comparing regression coefficients between models: Concepts and illustrative examples. National Institute of Statistical Sciences. 1997 Technical Report No. 65. [Google Scholar]
- Rick JH, Horvitz JC, Balsam PD. Dopamine receptor blockade and extinction differentially affect behavioral variability. Behav Neurosci. 2006;120(2):488–92. doi: 10.1037/0735-7044.120.2.488. [DOI] [PubMed] [Google Scholar]
- Rinaldi-Carmona M, Barth F, Héaulme M, Shire D, Calandra B, Congy C, et al. SR141716A, a potent and selective antagonist of the brain cannabinoid receptor. FEBS Letters. 1994;350(2-3):240–4. doi: 10.1016/0014-5793(94)00773-x. [DOI] [PubMed] [Google Scholar]
- Rusk IN, Cooper SJ. Parametric studies of selective D1 or D2 antagonists: Effects on appetitive and feeding behaviour. Behav Pharmacol. 1994;5(6):615–22. doi: 10.1097/00008877-199410000-00007. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Zigmond MJ, Stricker EM. Characterization of the impaired feeding behavior in rats given haloperidol or dopamine-depleting brain lesions. Neuroscience. 1990;39(1):17–24. doi: 10.1016/0306-4522(90)90218-s. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Steinpreis RE, McCullough LD, Smith P, Grebel D, Mahan K. Haloperidol and nucleus accumbens dopamine depletion suppress lever pressing for food but increase free food consumption in a novel food-choice procedure. Psychopharmacology (Berl) 1991;104(4):515–21. doi: 10.1007/BF02245659. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Mahan K, Rogers S. Ventrolateral striatal dopamine depletions impair feeding and food handling in rats. Pharmacol Biochem Behav. 1993a;44(3):605–10. doi: 10.1016/0091-3057(93)90174-r. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Kurth PA, McCullough LD, Sokolowski JD, Cousins MS. The role of brain dopamine in response initiation: Effects of haloperidol and regionally specific dopamine depletions on the local rate of instrumental responding. Brain Res. 1993b;628(1-2):218–26. doi: 10.1016/0006-8993(93)90958-p. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Kurth P, McCullough LD, Sokolowski JD. The effects of nucleus accumbens dopamine depletions on continuously reinforced operant responding: Contrasts with the effects of extinction. Pharmacol Biochem Behav. 1995;50(3):437–43. doi: 10.1016/0091-3057(94)00294-s. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Cousins MS, Maio C, Champion M, Turski T, Kovach J. Different behavioral effects of haloperidol, clozapine and thioridazine in an instrumental lever pressing/feeding procedure. Psychopharmacology (Berl) 1996;125(2):105–12. doi: 10.1007/BF02249408. [DOI] [PubMed] [Google Scholar]
- Salamone JD, Aberman JE, Sokolowski JD, Cousins MS. Nucleus accumbens dopamine and rate of responding: Neurochemical and behavioral studies. Psychobiology. 1999;27(2):236–47. [Google Scholar]
- Salamone JD, Arizzi MN, Sandoval MD, Cervone KM, Aberman JE. Dopamine antagonists alter response allocation but do not suppress appetite for food in rats: Contrast between the effects of SKF 83566, raclopride and fenfluramine on a concurrent choice task. Psychopharmacology (Berl) 2002;160(4):371–80. doi: 10.1007/s00213-001-0994-x. [DOI] [PubMed] [Google Scholar]
- Sink KS, Vemuri VK, Olszewska T, Makriyannis A, Salamone JD. Cannabinoid CB1 antagonists and dopamine antagonists produce different effects on a task involving response allocation and effort-related choice in food-seeking behavior. Psychopharmacology (Berl) 2008;196(4):565–74. doi: 10.1007/s00213-007-0988-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skjoldager P, Winger G, Woods JH. Analysis of fixed-ratio behavior maintained by drug reinforcers. J Exp Anal Behav. 1991;56(2):331–43. doi: 10.1901/jeab.1991.56-331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tallett AJ, Blundell JE, Rodgers RJ. Acute anorectic response to cannabinoid CB1 receptor antagonist/inverse agonist AM 251 in rats: Indirect behavioural mediation. Behav Pharmacol. 2007;18(7):591–600. doi: 10.1097/FBP.0b013e3282eff0a9. [DOI] [PubMed] [Google Scholar]
- Tallett AJ, Blundell JE, Rodgers RJ. Endogenous opioids and cannabinoids: System interactions in the regulation of appetite, grooming and scratching. Physiol Behav. 2008;94(3):422–31. doi: 10.1016/j.physbeh.2008.02.009. [DOI] [PubMed] [Google Scholar]
- Tanda G, Pontieri FE, Di Chiara G. Cannabinoid and heroin activation of mesolimbic dopamine transmission by a common mu1 opioid receptor mechanism. Science. 1997;276(5321):2048–50. doi: 10.1126/science.276.5321.2048. [DOI] [PubMed] [Google Scholar]
- Trevitt J, Atherton A, Aberman J, Salamone JD. Effects of subchronic administration of clozapine, thioridazine and haloperidol on tests related to extrapyramidal motor function in the rat. Psychopharmacology (Berl) 1998;137(1):61–6. doi: 10.1007/s002130050593. [DOI] [PubMed] [Google Scholar]
- Varvel SA, Vann RE, Wise LE, Philibin SD, Porter JH. Effects of antipsychotic drugs on operant responding after acute and repeated administration. Psychopharmacology (Berl) 2002;160(2):182–91. doi: 10.1007/s00213-001-0969-y. [DOI] [PubMed] [Google Scholar]
- Ward SJ, Dykstra LA. The role of CB1 receptors in sweet vs. fat reinforcement: Effect of CB1 receptor deletion, CB1 receptor antagonism (SR141716A) and CB1 receptor agonism (CP-55940) Behav Pharmacol. 2005;16(5-6):381–8. doi: 10.1097/00008877-200509000-00010. [DOI] [PubMed] [Google Scholar]
- Wiley JL, Martin BR. Cannabinoid pharmacological properties common to other centrally acting drugs. Eur J Pharmacol. 2003;471(3):185–93. doi: 10.1016/s0014-2999(03)01856-9. [DOI] [PubMed] [Google Scholar]