

Abstract
Phytoestrogens are plant‐derived dietary compounds with structural similarity to 17‐β‐oestradiol (E2), the primary female sex hormone. This structural similarity to E2 enables phytoestrogens to cause (anti)oestrogenic effects by binding to the oestrogen receptors. The aim of the present review is to present a state‐of‐the‐art overview of the potential health effects of dietary phytoestrogens. Various beneficial health effects have been ascribed to phytoestrogens, such as a lowered risk of menopausal symptoms like hot flushes and osteoporosis, lowered risks of cardiovascular disease, obesity, metabolic syndrome and type 2 diabetes, brain function disorders, breast cancer, prostate cancer, bowel cancer and other cancers. In contrast to these beneficial health claims, the (anti)oestrogenic properties of phytoestrogens have also raised concerns since they might act as endocrine disruptors, indicating a potential to cause adverse health effects. The literature overview presented in this paper illustrates that several potential health benefits of phytoestrogens have been reported but that, given the data on potential adverse health effects, the current evidence on these beneficial health effects is not so obvious that they clearly outweigh the possible health risks. Furthermore, the data currently available are not sufficient to support a more refined (semi) quantitative risk–benefit analysis. This implies that a definite conclusion on possible beneficial health effects of phytoestrogens cannot be made.
Linked Articles
This article is part of a themed section on Principles of Pharmacological Research of Nutraceuticals. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v174.11/issuetoc
Abbreviations
- ARE/EpRE
antioxidant/electrophile response element
- E2
17‐β‐oestradiol
- EFSA
European Food Safety Authority
- ERα
oestrogen receptor α
- ERβ
oestrogen receptor β
- ERs
oestrogen receptors (NRA3A)
- GPER
G protein‐coupled oestrogen receptor
- PPAR
peroxisome proliferator activated receptor (NR1C)
- TPO
thyroid peroxidase (EC number 1.11.1.8)
Tables of Links
| TARGETS | |
|---|---|
| GPCRs a | Nuclear hormone receptors c |
| GPER | ERα |
| Enzymes b | ERβ |
| AMPK | PPARγ |
| Thyroid peroxidase (TPO) | PPARa |
| LIGANDS |
|---|
| 17‐β‐oestradiol (E2) |
| Insulin |
These Tables list key protein targets and ligands in this article which are hyperlinked to corresponding entries in http://www.guidetopharmacology.org, the common portal for data from the IUPHAR/BPS Guide to PHARMACOLOGY (Southan et al., 2016) and are permanently archived in the Concise Guide to PHARMACOLOGY 2015/16 (a,b,cAlexander et al., 2015a,b,c).
Introduction
Phytoestrogens are plant‐derived dietary compounds, found in a wide variety of foods, especially in soy. They represent a diverse group of naturally occurring chemicals with structural similarity to 17‐β‐oestradiol (E2), the primary female sex hormone. Because the lack of phytoestrogens in the diet does not result in deficiency syndromes and because the phytoestrogens do not participate in any essential biological function, phytoestrogens are not considered nutrients. Their structural similarity to E2 enables them to cause (anti)oestrogenic effects by binding to the oestrogen receptors (ERs). This was already noticed in the previous century in Western Australia where sheep grazing on isoflavone‐rich red clover fields showed fertility problems (Bennetts et al., 1946; Stafford, 1997; Scherr et al., 2009). It has been hypothesized that plants use phytoestrogens as part of their natural defence to control female fertility to prevent overpopulation and overgrazing by herbivore animals (Hughes, 1988). In line with this, Setchell (Setchell, 1998) suggested that the fertility problems of zoo animals could be related to the presence of soy isoflavone phytoestrogens in the standard animal diet. Besides these adverse effects, various beneficial health effects have been ascribed to phytoestrogens, such as a lowered risk of menopausal symptoms like hot flushes and osteoporosis. As a result, phytoestrogens are present in a large number of dietary supplements and widely marketed as natural alternatives to oestrogen replacement therapy. In addition, phytoestrogen exposure has been related to lowered risks of cardiovascular disease, obesity, metabolic syndrome and type 2 diabetes, brain function disorders, breast cancer and other forms of cancer including prostate cancer, bowel cancer and other cancers (Hughes, 1988; Adlercreutz, 2002; Bhathena and Velasquez, 2002; Karahalil, 2005; Cederroth and Nef, 2009; Patisaul and Jefferson, 2010; Zhao and Mu, 2011; Jungbauer and Medjakovic, 2014).
In the last decades, soy isoflavones have received attention because of the so called ‘Japanese Phenomenon’ connected to a lower incidence of specific chronic diseases in the Japanese compared with the Western population due to a higher intake of soy foods from early life onwards (Watanabe et al., 2002; Korde et al., 2004; Korde et al., 2009). The fact that the prevalence of breast cancer in daughters of migrated Japanese Americans became similar to that of Caucasian Americans after changing their food habits is in line with this observation.
In contrast to these beneficial health claims, the (anti)oestrogenic properties of phytoestrogens have also raised concerns since they might act as endocrine disruptors, indicating a potential to cause adverse health effects. Altogether, the health benefits or risks of isoflavones and other phytoestrogens are still controversial (Wuttke et al., 2007; Andres et al., 2011; Rietjens et al., 2013), and the question of whether phytoestrogens are beneficial or harmful to human health remains unresolved.
Keeping that in mind, the aim of the present review is to present a state‐of‐the‐art overview of the potential health effects of dietary phytoestrogens. The paper presents an overview of the different phytoestrogens present in the diet and food supplements, their supposed mode(s) of action and recent evidence on their supposed beneficial effects.
To obtain an overview of the various health effects, we searched Web of Science, Scopus and PubMed for entries with the search terms ‘phytoestrogen(s)’, ‘review’, ‘meta‐analysis’ and the respective health effects in titles, abstracts and keywords.
Phytoestrogens in the diet
Figure 1 presents an overview of the major types of phytoestrogens known to be present in the diet and food supplements, also including their chemical structure compared with that of E2. The major groups of phytoestrogens present in our diet are isoflavones, prenylflavonoids, coumestans and lignans. The main isoflavones are genistein, daidzein, glycitein, formononetin and biochanin A, which are mainly found in soy, soy‐based food and legumes usually in their conjugated forms like genistin, daidzin, puerarin, glycitin, ononin and sissotrin. In countries in Asia where fermented soy products are part of the traditional diet, isoflavone intake levels may amount to about 15–50 mg isoflavones per day (Eisenbrand et al., 2007). In Western industrial countries, isoflavone intake has been reported to be less than 2 mg isoflavones per day (Eisenbrand et al., 2007), although it may be higher for menopausal women who take soy‐based preparations as an alternative to hormone replacement therapy. The dosages recommended by the manufacturers may vary with the product, and have been reported to amount to values between 20 and 80 mg isoflavones per day (Eisenbrand et al., 2007).
Figure 1.
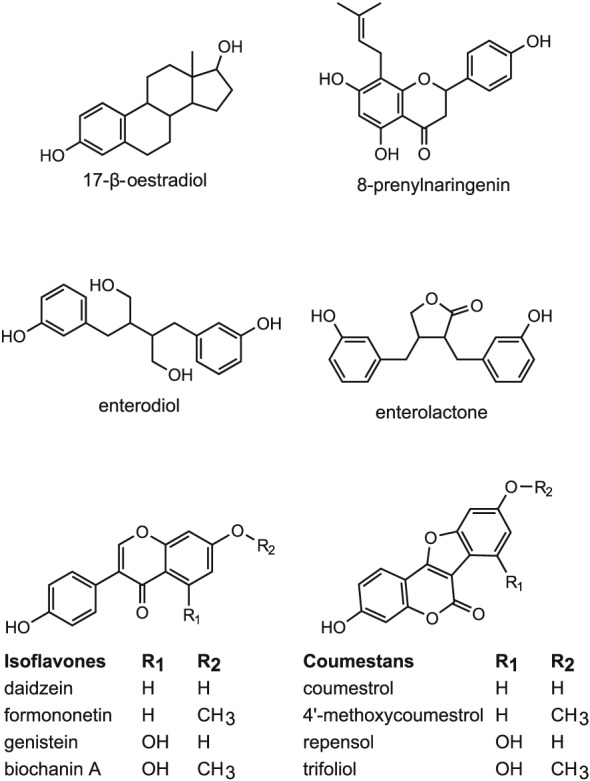
Chemical structures of E2 and the most common phytoestrogens.
Major prenylflavonoids are 6‐prenylnaringenin, 6‐geranylnaringenin, 8‐prenylnaringenin and isoxanthohumol, which can all be found in hops and beer (Stevens and Page, 2004; Dhooghe et al., 2010). Of the prenylflavonoids, 8‐prenylnaringenin (Figure 1) is the most potent phytoestrogen known. The main coumestans are coumestrol, 4′‐methoxycoumestrol, repensol and trifoliol (Figure 1). Food sources high in coumestans include split peas, pinto beans, lima beans, and especially, alfalfa and clover sprouts. Lignans that are classified as phytoestrogens are enterodiol and enterolactone (Figure 1), which are formed from lignan precursors by intestinal bacteria (Lampe, 2003). Lignan precursors include pinoresinol, lariciresinol, secoisolariciresinol, matairesinol and others (Figure 2). Lignan precursors are found in a wide variety of foods, including flaxseeds, whole grains, fruits and vegetables, sesame seeds and legumes and present a principal source of dietary phytoestrogens in the Western diet (de Kleijn et al., 2002; Valsta et al., 2003).
Figure 2.
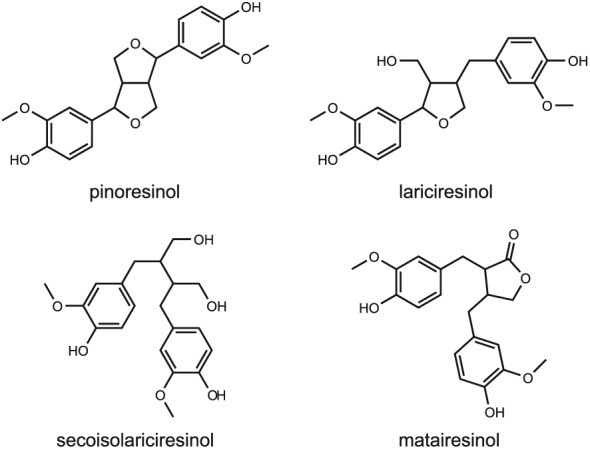
Chemical structures of some dietary lignan precursors.
Mode(s) of action: interaction with oestrogen receptors
The major mode of action by which phytoestogens may exert their possible health effects, is based on their structural similarity to E2 enabling them to cause (anti)oestrogenic effects by binding to the ER. Two main ERs, that is oestrogen receptor α (ERα) (NR3A1) and oestrogen receptor β (ERβ) (NR3A2), have been identified in rats, mice, primates and humans (Kuiper et al., 1996; Ogawa et al., 1998). These ER subtypes have different roles in gene regulation, cancer biology and therapy (Nilsson et al., 2001; Williams et al., 2008; Thomas and Gustafsson, 2011). ERα activation in breast and uterus has been shown to enhance cell proliferation, necessary for growth and tissue maintenance (Pearce and Jordan, 2004; Harris, 2007; Thomas and Gustafsson, 2011) but may also play a role in the unlimited growth of, in particular, ERα‐dependent breast tumours of which around 70% respond to anti‐oestrogen therapy with, for example, the antagonist tamoxifen (Ali and Coombes, 2000). ERβ has been shown to counteract the ERα‐mediated stimulation of cell proliferation (Bardin et al., 2004; Stossi et al., 2004; Strom et al., 2004; Sotoca et al., 2008b; Sotoca et al., 2008a; Thomas and Gustafsson, 2011). These opposite roles of ERα and ERβ in cellular responses to oestrogens have been illustrated by studies using the so‐called T47D–ERβ cell line, a breast cancer cell line with a constant level of ERα, and a tetracycline‐dependent adjustable expression level of ERβ (Strom et al., 2004). When cells of this cell line express mainly ERα, genistein and E2 both appeared to induce a concentration‐dependent increase in proliferation. When ERβ expression is induced, E2 and genistein no longer induce cell proliferation. These results support the conclusion that ERβ plays a role in counteracting ERα‐mediated cell proliferation.
Because ERα and ERβ have different roles in gene regulation, cell proliferation and related health effects, their varying ratio and relative level within tissues may influence the cellular response towards different phytoestrogens. As a result, a certain phytoestrogen may have different effects in, for example, the uterus, in which ERα is the major isoform (Pearce and Jordan, 2004), than in the prostate, in which ERβ is dominant (Enmark et al., 1997; Pearce and Jordan, 2004). These tissue‐specific effects may also result from differences in coactivators and corepressors activated upon activation of the two ERs in different tissues and/or the possible crosstalk of the ERs with other nuclear receptors (Wilson et al., 2004; Chang et al., 2008; Vanden Berghe and Haegeman, 2008; Evers et al., 2014a; Evers et al., 2014b). Furthermore, the actual mode of action of a phytoestrogen, either as an agonist or an antagonist, may also depend on the level of endogenous estrogens present (Barnes et al., 1995).
Using various in vitro models that have been developed for the detection of oestrogen activity, the relative oestrogenic potencies of a variety of phytoestrogens have been quantified. These in vitro assays include receptor binding studies (Kuiper et al., 1998; Gutendorf and Westendorf, 2001; Ikeda et al., 2002; De Angelis et al., 2005; Boue et al., 2011; Park et al., 2012; Djiogue et al., 2014; Liu et al., 2014), ERα and ERβ‐dependent reporter gene assays (Kuiper et al., 1998; Casanova et al., 1999; Dornstauder et al., 2001; Gutendorf and Westendorf, 2001; Ikeda et al., 2002; Rickard et al., 2003; De Angelis et al., 2005; Harris et al., 2005; Escande et al., 2006; ter Veld et al., 2006; Chrzan and Bradford, 2007; Sotoca et al., 2008b; Chu et al., 2009; Kwack et al., 2009; Takeuchi et al., 2009; Boue et al., 2011; Park et al., 2012; Djiogue et al., 2014; Liu et al., 2014; Tiosano et al., 2014; Islam et al., 2015) and cell proliferation assays using oestrogen‐sensitive human cell lines derived from three different female oestrogen‐sensitive tissues, including breast (MCF‐7/BOS and T47D), endometrial (ECC‐1) and ovarian (BG‐1) cells (Wang et al., 2012). Few studies also report on the oestrogenic effects of phytoestrogens in vivo in the so‐called uterotrophic assay (Ding et al., 2010; Wang et al., 2012).
For the present review, we performed a literature research on the binding affinity of E2 and phytoestrogens to ERα and ERβ (expressed as IC50 values from competitive binding assays) and the ERα‐ and ERβ‐mediated gene expression induced by E2 and phytoestrogens (expressed as EC50 values from reporter gene assays). To obtain this overview, we searched Web of Science, Scopus and PubMed for entries with the search terms ‘phytoestrogen(s)’, ‘ERα’, ‘ERβ’, ‘binding affinity’ and/or ‘reporter gene’ in titles, abstracts and keywords. In Figure 3, the reported IC50 and EC50 values for ERα are plotted against the reported IC50 and EC50 values for ERβ. In Tables S1 and S2, the IC50 values and the EC50 values, respectively, and the references to the literature are presented.
Figure 3.
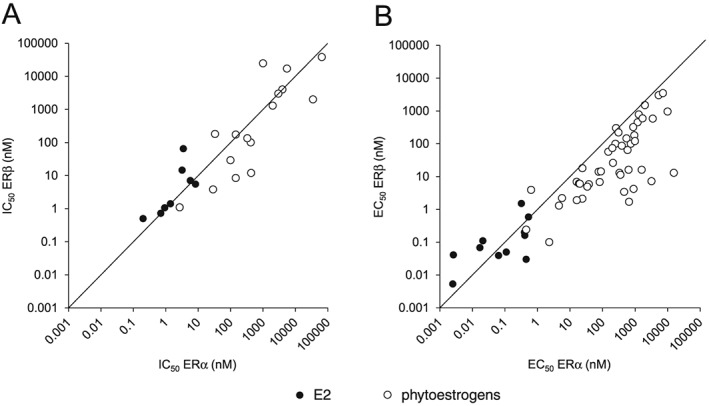
(A) Binding affinities to ERα and ERβ (expressed as IC50 values) and (B) effect concentrations in ERα and ERβ reporter gene assays (expressed as EC50 values) of E2 and phytoestrogens. In Tables S1 and S2, the IC50 and EC50 values, respectively, and the references to the literature are presented.
The overall trend emerging from this overview is that phytoestrogens are less potent oestrogens than E2 with higher IC50 and EC50 values for both receptors (Figure 3). Furthermore, the data show for most phytoestrogens that have been investigated, that the IC50 values are higher for ERα than for ERβ (Figure 3A), indicating a higher binding preference for the ERβ than for the ERα. Only for ferutinine, kievitone and for psoralidin the IC50 values are higher for ERβ than for ERα (Table S1), indicating a higher binding preference for the ERα for these phytoestrogens. A higher preference of ferutinine for ERα was also found in reporter gene studies, whereas for the majority of phytoestrogens tested in reporter gene systems, a preference for ERβ‐mediated gene expression was observed (Figure 3B and Table S2). Most research on the binding of phytoestrogens to ERα and ERβ, and the activation of ERα‐ and ERβ‐mediated gene expression has been conducted with the major soy isoflavones genistein and daidzein (Kuiper et al., 1998; Casanova et al., 1999; Dornstauder et al., 2001; Gutendorf and Westendorf, 2001; Rickard et al., 2003; Bovee et al., 2004; De Angelis et al., 2005; Harris et al., 2005; Escande et al., 2006; Chrzan and Bradford, 2007; Sotoca et al., 2008b; Chu et al., 2009; Takeuchi et al., 2009; Tiosano et al., 2014; Beekmann et al., 2015; Islam et al., 2015). Figure 4 presents an example of such a study with genistein as measured in the human osteosarcoma (U2OS) ERα or ERβ reporter cell lines, comparing the activity of genistein with that of E2 (Sotoca et al., 2008b). The data presented in Figure 4 reveal that for E2, physiological concentrations may be such that only ERα is activated since the EC50 value of E2 for activation of ERα is about 10‐fold lower than that for activation of ERβ. For the isoflavone genistein, the EC50 for ERβ activation is lower than that for ERα activation, indicating that concentrations that activate ERα will at the same time activate ERβ that will counteract the ERα‐mediated effects on cell proliferation (Strom et al., 2004; Sotoca et al., 2008a; Rietjens et al., 2013). This may result in different physiological effects induced by E2 than those induced by phytoestrogens.
Figure 4.
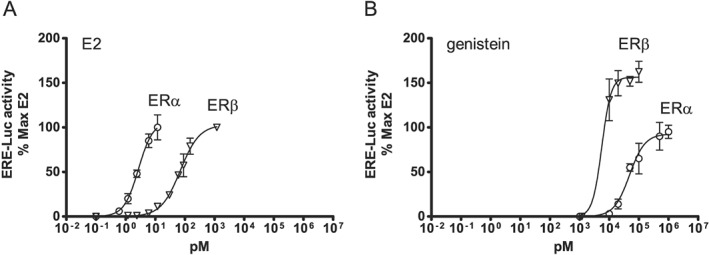
Induction of oestrogen responsive element mediated luciferase activity in the ERα‐ and ERβ‐containing U2OS reporter cell lines by (A) E2 and (B) genistein. For further details, see Rietjens et al. (2013).
It is also of interest to note the possible role of G protein‐coupled oestrogen receptors (GPERs), since many of the phytoestrogens including the soy isoflavone genistein and possibly equol activate GPERs (Prossnitz and Barton, 2011). In cell lines of thyroid, ovarian, endometrial and breast cancers, stimulation of GPERs with oestrogens including genistein, activates a signalling pathway that promotes proliferation, although inhibition of proliferation has also been reported. In particular, genistein has been reported to stimulate growth of MCF‐7 cells through a GPER‐dependent mechanism (Prossnitz and Barton, 2011).
Mode(s) of action: epigenetic effects
Another mode of action underlying the health effects of phytoestrogens may relate to epigenetic mechanisms. Various natural bioactive compounds have been shown to affect the epigenome, but for phytoestrogens, this has been mainly assessed for the soy isoflavone genistein, and to a lesser extent for the soy isoflavone daidzein and its microbial metabolite equol (Remely et al., 2015b). Especially for the reduction in cancer risk by the isoflavone genistein, a role for epigenetic changes resulting in alterations in the expression of genes that regulate cell proliferation and differentiation has been reported (Dolinoy et al., 2006; Hilakivi‐Clarke et al., 2010). Genistein consumption is also among the factors that have been associated with epigenetic modifications in obesity (Remely et al., 2015a). Among these epigenetic changes were effects on DNA methylation, histone modification and microRNA regulation (Rietjens et al., 2013). Several studies have assessed the effects of genistein on DNA methylation.
Dolinoy et al. (2006) investigated, in the so‐called agouti mouse model, the effect of genistein on DNA methylation in the offspring exposed during gestation. Genistein induced CpG hypermethylation of six CpG sites upstream of the agouti gene, resulting in a shift of the coat colour distribution towards pseudoagouti (brown), and a decrease in the incidence of the onset of obesity in the offspring at adult age (Dolinoy et al., 2006). The authors speculated that this ability of genistein to increase DNA methylation might provide an explanation for the lower incidence of certain cancers in Asians as compared with Westerners (Qin et al., 2009). This observation could provide another explanation for the different outcomes reported when studying effects of phytoestrogens on breast cancer incidence. It could be that beneficial effects result from epigenetic modifications that occur early in life perhaps even through soy intake by the mother, but that these beneficial effects may not be observed when intake of soy isoflavones starts later in life, for example when using hormone replacement therapy. Also, adolescence may reflect a sensitive period for phytoestrogens, and phytoestrogen intake during adolescence may reduce breast cancer risk later in life (Lee et al., 2009; Anderson et al., 2013; Molzberger et al., 2013).
In in vitro studies, genistein has been shown to decrease the methylation of several tumour suppressor genes (Pudenz et al., 2014), which may be mediated by the inhibition of DNA methyl transferase (DNMT) activity (Singh et al., 2013). This may allow silenced tumour suppressor genes to be re‐expressed, indicating a potential beneficial effect of genistein. However, inhibition of DNMT may also lead to a reduced methylation of proto‐oncogenes, as has been shown for coumestrol, daidzein and equol (Lyn‐Cook et al., 1995; Koo et al., 2015), which may indicate a possible adverse effect. However, whether these findings from in vitro studies are relevant for the in vivo situation remains to be established, since genistein has been shown to cause an increase in DNA methylation upon exposure in vivo, such as in the agouti mouse model as mentioned above (Dolinoy et al., 2006) and in humans (Qin et al., 2009). This human study was a double‐blind, randomized trial with 34 healthy premenopausal women, who received 40 or 140 mg isoflavones (including genistein, daidzein and glycitein) a day for the duration of one menstrual cycle, followed by characterization of the methylation status of five cancer related genes known to be methylated in breast cancer (p16, RASSF1A, RARβ2, ER and CCND2) assessed in isolated breast tissue samples (Qin et al., 2009). The results obtained revealed a treatment‐related hypermethylation of the tumour suppressor genes RARβ2 and CCND2. The implications of these findings for human health are not clear yet, although this increase in methylation of tumour suppressor genes may indicate a possible adverse epigenetic change.
Other possible modes of action
In addition to the action of phytoestrogens via the oestrogen receptors or epigenetic modes of action, some studies suggested other possible modes of action of phytoestrogens. This includes, for example, the effects of genistein resulting in activation of AMP‐activated protein kinase (EC number 2.7.11.31) in cells exposed to genistein in vitro (Hwang et al., 2005; Park et al., 2010). The exact mechanism underlying this activation has not been fully elucidated (Hsu et al., 2011) but may be related to genistein‐mediated stimulation of intracellular production of ROS (Hwang et al., 2005).
Another mode of action may relate to the action of phytoestrogens as kinase inhibitors (Dubey et al., 1999; Kang et al., 2007; Yan et al., 2010). Given that the human kinome is composed of hundreds of protein kinases of which many have been reported to be disease associated (Manning et al., 2002; Lahiry et al., 2010), one could suggest that this is a possible mode of action underlying the effects of phytoestrogens on health. It is important to note, however, that EC50 values generally reported for this inhibition of protein kinases by isoflavones seem to be around the two‐digit and lower three‐digit μM range (Dubey et al., 1999; Kang et al., 2007; Kim et al., 2009), which is substantially higher than generally reported physiological plasma concentrations of the respective phytoestrogens, which tend to be in the two‐ to three‐digit nM range, rarely reaching low μM concentrations (King and Bursill, 1998; Verkasalo et al., 2001; Setchell et al., 2003).
Phytoestrogens may also activate the PPAR (NRC1C) family including, particularly PPARα (NR1C1), and PPARγ (peroxisome proliferator activated receptor γ) (NR1C3) (Jungbauer and Medjakovic, 2014). It has been suggested that this mode of action mainly contributes to the possible effects of phytoestrogens on obesity, metabolic syndrome and type 2 diabetes, as PPARγ agonists are effective drugs for patients with type 2 diabetes. Specifically, isoflavones and their metabolites appear to be PPAR agonists with EC50 values in the two‐digit μM range (Jungbauer and Medjakovic, 2014). It remains to be established whether physiological concentrations are actually high enough to induce these PPAR‐mediated effects.
Another mode of action, suggested mainly for the beneficial health effects of phytoestrogens, is the induction of antioxidant/electrophile response element (ARE/EpRE)‐mediated gene expression by activation of nuclear factor erythroid 2‐related factor 2‐Keap 1 signalling (Jungbauer and Medjakovic, 2014). EpRE‐mediated induction of gene expression leads to increased cellular defence against the toxicity of electrophiles and ROS and related adverse health effects. Activation of this pathway may be related to protection against tumour induction and oxidative stress in cardiovascular disease.
Finally, it is of interest to consider the possible role of polymorphisms in the differential effects of phytoestrogens. So far, differences in health effects of phytoestrogens have not been related to specific gene polymorphisms. However, since ERs seem to play an important role in several of the health effects of phytoestrogens, it can be expected that phytoestrogen‐related health effects may differ between persons with differences in ER function, for example resulting from ER gene polymorphisms. Nott et al. (2008) concluded that an increasing body of evidence implicates ERα polymorphisms as one of the contributing factors for differential responses to oestrogen competitors and that polymorphisms in the ER genes may influence the individual response to hormone replacement therapy (Gennari et al., 2005). Polymorphisms of genes that play a role in the metabolism of (phyto)oestrogens may also play a role in differences in health effects of phytoestrogens in the human population (Wang et al., 2011). Also, polymorphisms related to interindividual differences in the gut microbiota and the consequences of these differences for formation of (in)active metabolites may play a role. An adequate example is the formation of equol as a metabolite of daidzein only in a part of the population. Equol has been reported to be more active as an oestrogen‐active compound than daidzein itself (Magee, 2011). Another example is the formation of enterodiol and enterolactone from ingested lignans through the gut microbiota in only a part of the population (Kuijsten et al., 2005; Hullar et al., 2015). Also, these differences can be an underlying reason for different responses to phytoestrogen exposure in the population.
Effects of phytoestrogens on menopausal symptoms
Several meta‐analyses investigated the effects of phytoestrogens or soy isoflavone extracts or supplements on menopausal symptoms. These studies reported not only a reduction in the frequency and severity of hot flashes (Howes et al., 2006; Taku et al., 2012; Chen et al., 2015) but also no conclusive evidence or only some indications for a reduction in hot flush frequency or severity (Nedrow et al., 2006; Lethaby et al., 2007; Jacobs et al., 2009; Bolanos et al., 2010; Eden, 2012; Lethaby et al., 2013), varying effects on spine bone mineral density and no effects on femoral neck, hip total and trochanter bone mineral density (Taku et al., 2010), and no protection against bone fracture (Kreijkamp‐Kaspers et al., 2004; Tempfer et al., 2007). The European Food Safety Authority (EFSA) evaluated the health claims related to the reduction of vasomotor symptoms and the maintenance of bone mineral density by soy isoflavones during menopause (EFSA NDA Panel, 2012). It was concluded that the available evidence was not sufficient to establish a relationship between the maintenance of bone mineral density and the consumption of soy isoflavones. Also, a relationship between the reduction of vasomotor symptoms associated with menopause and the consumption of soy isoflavones could not be established. EFSA evaluated 14 long‐term (>12 months) intervention studies on the effects of soy isoflavones on bone mineral density in post‐menopausal women. Of these studies, only two, reported an effect of soy isoflavones on bone mineral density and on markers of bone formation or resorption at doses of 54 mg per day (Morabito et al., 2002; Marini et al., 2007). The remaining 12 studies, testing doses of isoflavones up to 200 mg per day, showed no effects of soy isoflavones on bone mineral density or markers of bone formation or resorption [(EFSA NDA Panel, 2012) and references therein], although EFSA indicated that for some studies this negative result may have been due to the limited number of subjects included, resulting in an underpowered study.
EFSA also evaluated five human intervention studies in which soy isoflavones were administered for 6–9 months, and effects on bone mineral density were measured in peri‐/post‐menopausal women. One of these intervention studies revealed a beneficial dose–response effect on bone mineral density at the lumbar spine and femoral neck. Two of the studies reported a beneficial effect of soy isoflavones on bone mineral density at the lumbar spine, and two studies showed no effect of soy isoflavones on bone mineral density. EFSA concluded that ‘these studies provide some evidence for an effect of soy isoflavones on the attenuation of bone mineral density loss at the lumbar spine in post‐menopausal women when consumed for 6‐9 months’. Given the effects reported on parameters for bone formation and bone resorption, EFSA also concluded that these effects were possibly mediated by a decrease in bone resorption. However, taking all the evidence together, EFSA considered ‘the evidence insufficient to establish a cause and effect relationship between the consumption of soy isoflavones and maintenance of bone mineral density in post‐menopausal women’.
A later study reported by Greendale et al. (Greendale et al., 2015) investigated cross‐sectional and longitudinal relations between dietary intake of isoflavones and bone mineral density at the lumbar spine and femoral neck in Black, White, Chinese and Japanese women during the menopausal transition. In Japanese women, higher isoflavone intake appeared to be associated not only with higher peak femoral neck bone mineral density but also with greater rates of lumbar spine and femoral neck bone mineral density loss during the menopausal transition. Results for the other racial/ethnic groups did not support a relationship between dietary intake of isoflavones and either peak bone mineral density, or bone mineral density loss during the menopausal transition.
Altogether, it seems that the evidence for a beneficial effect of isoflavones and other phytoestrogens on bone mineral density in post‐ and peri‐menopausal women is limited and not convincing, and that they may even cause adverse effects, as indicated by the study of Greendale et al. (2015).
Effects of phytoestrogens on cardiovascular disease
In particular, the increase in cardiovascular risk and cardiovascular mortality upon sudden loss of ovarian function has provided a basis for the proposed role of oestrogens and phytoestrogens in the reduction in cardiovascular disease (Colditz et al., 1987; Parker et al., 2009). Reduced oestrogen levels during menopause may influence the development of obesity, fat distribution, the lipid profile in plasma, and rheological properties of plasma and platelet function (Gorodeski, 1994). These observations suggest that oestrogen deficiency may promote cardiovascular disease in women and trigger the idea that phytoestrogens may reduce the risk. The hypothesis is also supported by the low rates of cardiovascular diseases in Asian populations where the diet is particularly rich in soy, followed by the loss of this protection among the groups that have moved to Western societies (Nagata, 2000; Zhang et al., 2003). A meta‐analysis reported by Tokede et al. (2015) concluded that isoflavone supplementation had no effect on serum lipid profiles.
Oestrogens have been shown to influence atherosclerosis and the related clinical events in a differential way (Rossouw et al., 2007; Cano et al., 2010). They may act not only as protectors against atherosclerosis but also as potential disruptors of established atherosclerotic plaques, the latter being important hallmarks in the pathogenesis of the arterial forms of cardiovascular disease. The concept that phytoestrogens may act in a similar way comes from the observation that in Asian populations with high levels of their consumption, the prevalence of cardiovascular disease is lower than in populations in Western countries (Gonzalez Canete and Duran Aguero, 2014).
Cano et al. (2010) and also Gonzalez Canete and Duran Aguero (2014) reviewed the main evidence on the effects of isoflavones on the cardiovascular system at both the experimental and the clinical level. The review indicated that two clinical studies (van der Schouw et al., 2005; Kokubo et al., 2007) have examined the association of isoflavone intake with clinical cardiovascular events in a population without cardiovascular disease at baseline. One study concluded that high isoflavone intake was associated with reduced risk of cerebral and myocardial infarctions in Japanese women, with the risk reduction being especially pronounced in postmenopausal women (Kokubo et al., 2007). The second study was performed in a Western population and investigated whether a low intake of phytoestrogens would be associated with increased risks of cardiovascular disease (van der Schouw et al., 2005). The study concluded that an intake of isoflavones or lignans was not associated with decreased cardiovascular disease risk. However, when stratifying for ever versus never smokers, the risk of cardiovascular disease appeared to decrease with increased intake of lignans for ever smokers. The authors concluded that their data do not support the presence of a protective effect on cardiovascular disease risk of higher habitual intake of phytoestrogens, although for smokers a small risk reduction with higher lignan intake could not be excluded. In clinical practice, none of the phytoestrogens investigated have been proven to protect against cardiovascular disease (Tempfer et al., 2007).
Altogether, it can be concluded that it is not really clear if the administration of phytoestrogens in the early postmenopausal period may be protective for cardiovascular disease. For phytoestrogens, the current evidence appears poor compared to that available for oestrogens, and it is possible that the potential effect of oestrogens on the risk for stroke is not reproduced by isoflavones (Cano et al., 2010).
Effects of phytoestrogens on obesity, metabolic syndrome and type 2 diabetes
Dietary phytoestrogens have also been suggested to play a beneficial role in obesity, metabolic syndrome and type 2 diabetes (Bhathena and Velasquez, 2002; Cederroth and Nef, 2009; Crespillo et al., 2011; Jungbauer and Medjakovic, 2014; Struja et al., 2014). The meta‐analysis by Fang et al. (2016) reported an improvement in glucose metabolism and a significant reduction in insulin levels and insulin resistance in menopausal women, by especially genistein. Also, the meta‐analysis reported by Zhang et al. (2013) concluded that soy isoflavone supplementation could improve glucose metabolism and insulin control in non‐Asian postmenopausal women. Nutritional intervention studies in both animals and humans suggested that an intake of soy protein with isoflavones and flaxseed results in improved glucose control and insulin resistance (Bhathena and Velasquez, 2002; Cederroth and Nef, 2009; Jungbauer and Medjakovic, 2014). It is important to note that at present, it often remains unclear whether the beneficial effects are really due to the phytoestrogens or to some other dietary component in the soy protein and flax seed diet. The activation of PPAR‐mediated effects already mentioned above may provide a possible mode of action underlying these effects. However, as already concluded by Bhathena and Velasquez (2002), further investigations are needed to evaluate the long‐term effects of phytoestrogens on obesity, metabolic syndrome and type 2 diabetes.
Effects of phytoestrogens on breast cancer
The use of soy‐based preparations has been proposed for the prevention and treatment of certain types of cancer, such as for the prevention and treatment of breast cancer in women and prostate cancer in men (Eisenbrand et al., 2007). In contrast, clinical studies have reported data that suggest that isoflavones may via their oestrogenic and proliferative effects possibly raise breast cancer incidence in sensitive individuals (Petrakis et al., 1996; Hargreaves et al., 1999).
Meta‐analyses of epidemiological studies conducted in women consuming high‐soy diets concluded that there is a significant trend of decreased risk for breast cancer upon increasing intake of soy food (Trock et al., 2006; Wu et al., 2008; Dong and Qin, 2011; Chen et al., 2014). High lignan exposure has also been associated with a reduced breast cancer risk in postmenopausal women (Velentzis et al., 2009; Buck et al., 2010). Fritz et al. (2013) reported a systematic review and meta‐analysis on the potential effects of soy, red clover and isoflavone intake on breast cancer incidence and recurrence. The analysis included 40 randomized controlled trials, 11 uncontrolled trials and 80 observational studies. The authors concluded that soy consumption may be associated with reduced risk of breast cancer incidence, recurrence and mortality. Soy intake consistent with a traditional Japanese diet (2–3 servings a day containing 25–50 mg isoflavones) may also be protective against breast cancer and recurrence. Nevertheless, the authors also indicate that better evidence confirming the safety of isoflavones would be required before the use of high doses (≥100 mg) of isoflavones can be recommended for breast cancer patients. In addition, a nested case–control study and meta‐analysis of epidemiological studies revealed an inverse correlation between genistein intake and breast cancer (Taylor et al., 2009). It is important to note that this protective effect may originate from soy intake early in life (Warri et al., 2008; Hilakivi‐Clarke et al., 2010). Also of interest is the possible involvement of equol, a metabolite of daidzein produced by the human intestinal microflora, which may have similar beneficial effects on the incidence of breast cancer (Ingram et al., 1997; Goodman et al., 2009), although other studies have reported no or even adverse effects of equol (Magee, 2011). Given that only a limited percentage (30–40%) of the population has the ability to convert daidzein to equol, and taking into account that in vitro studies suggest equol to be more biologically active than its parent compound daidzein (Magee, 2011), it can be suggested that inter‐individual variation in response to daidzein may in part be related to variability in gut microflora composition resulting in inter‐individual differences in conversion of daidzein to equol. This existence of equol producers and non‐producers could provide an alternative explanation for interindividual differences in the response to phytoestrogens, since phytoestrogen metabolite formation by the gut microbiota can vary significantly between individuals with metabolites sometimes being more active than the parent phytoestrogen.
Also, the lignans enterodiol and enterolactone have been suggested to exert protective effects on breast cancer, possibly not only by oestrogen receptor‐dependent but also oestrogen receptor‐independent mechanisms (Mueller et al., 2004; Adlercreutz, 2007; Penttinen et al., 2007; Mense et al., 2008; Saarinen et al., 2010; Buck et al., 2011). There are also significant inter‐individual differences for enterodiol and enterolactone production due to different compositions of the gut microflora, which can cause a variation in the response of individuals to lignan precursors (Yoder et al., 2015). While coumestans are reported to be potent phytoestrogens in vitro, their effect on breast cancer is not known (Scarlata and Miksicek, 1995; Kuiper et al., 1998).
Recently, EFSA published a risk assessment for peri‐ and post‐menopausal women consuming food supplements containing isolated isoflavones (EFSA ANS Panel, 2015). EFSA evaluated especially the possible association between the isoflavone intake from food supplements and harmful effects observed in peri‐ and post‐menopausal women for the mammary gland, uterus and thyroid (EFSA ANS Panel, 2015). For the evaluation, EFSA selected 43 human studies and 62 animal studies. Among these studies were four epidemiological studies investigating breast cancer incidence (Rebbeck et al., 2007; Obi et al., 2009; Brasky et al., 2010; Boucher et al., 2013), eight interventional controlled studies, measuring mammographic density (Morabito et al., 2002; Atkinson et al., 2004; Marini et al., 2008; Powles et al., 2008; Verheus et al., 2008; Maskarinec et al., 2009; Colacurci et al., 2013; Delmanto et al., 2013), and two interventional controlled studies, investigating histopathological changes (Cheng et al., 2007; Khan et al., 2012). These studies did not reveal an association between isoflavone exposure and adverse effects in the mammary gland (EFSA ANS Panel, 2015). Of the 11 animal studies that investigated histopathological changes in the mammary gland upon treatment with isoflavones and the 10 studies in ovariectomized animals that investigated breast cell proliferation, the majority did not reveal an effect (EFSA ANS Panel, 2015). Only in two of the studies in ovariectomized rats, a stimulating effect on the mammary gland was observed after 90 days of dosing with genistein at 5.4 and 54 (Rimoldi et al., 2007) and 221 mg.kg bwt−1.day−1 (Wuttke et al., 2006). EFSA concluded that these findings were ‘consistent with the results from the US National Toxicology Program study conducted in non‐ovariectomized animals administered genistein at doses ranging 0.3–44 mg.kg bwt−1.day−1 (NTP, 2008), in which there was some evidence of carcinogenic activity of genistein in female rats based on an increased incidence of mammary gland adenoma or adenocarcinoma’. EFSA also indicated that the human data from observational studies did not point at an increased risk of breast cancer in menopausal women but specifically also noted that this conclusion cannot be extended to the risk of oestrogenic isoflavones‐based food supplements for postmenopausal women who have a diagnosis or history of oestrogen‐dependent cancer. Also, other endpoints considered did not reveal any effect except for some non‐malignant histopathological changes in mammary tissue after 60 months supplementation at 150 mg·day−1 of soy isoflavones which is high compared with the current estimated daily intakes.
Thus, concerns still exist that the oestrogenic activity of phytoestrogens may present a risk to patients with oestrogen‐sensitive breast cancer and to women that are at a high risk of developing breast tumours (Messina, 2008; Hilakivi‐Clarke et al., 2010), and it remains to be stablished whether exposure to isoflavones reduces or increases breast cancer risks.
Effects of phytoestrogens on other forms of cancer including prostate cancer, bowel cancer, uterine cancer and other cancers
Epidemiological studies and a nested case–control study carried out in Japan reported that isoflavone intake might be associated with a decreased risk of lung cancer (Shimazu et al., 2011). This observation also follows from a meta‐analysis indicating that the consumption of soy food is associated with lower lung cancer risk (Yang et al., 2011). Another nested case–control study performed with the Korean Multicenter cancer cohort revealed that high serum concentrations of isoflavones were associated with a decreased risk for gastric cancer (Ko et al., 2010). Furthermore, several studies report a negative correlation between prostate cancer incidence and a phytoestrogen rich diet containing lignans and/or isoflavones (Severson et al., 1989; Adlercreutz, 1995; Kurahashi et al., 2007), and a meta‐analysis revealed that the consumption of soy foods or genistein and daidzein is associated with a reduction in prostate cancer risk in men (Hwang et al., 2009; Yan and Spitznagel, 2009; He et al., 2015). Another meta‐analysis reported by Van Die et al. (2014) concluded that there is a potential role for soy and soy isoflavones in reducing risks for prostate cancer, but that a clear understanding could not be derived from the data because of limitations in sample size and study duration in the individual trials. Also equol, the gut microbiome metabolite of the soy isoflavone daidzein was suggested to have beneficial effects on the incidence of prostate cancer (Akaza et al., 2004; Ozasa et al., 2004). Epidemiological studies suggest that high dietary intake of phytoestrogens in pre‐ and post‐menopausal women is correlated with reduced thyroid cancer risk (Horn‐Ross et al., 2002; Haselkorn et al., 2003). For oestrogens in general, increased exposure has been linked to an increased risk of developing uterus cancer in women (Persson, 2000), and of prostate and testicular cancer in men (Bonkhoff et al., 1999; Maffini et al., 2006). Women who consume a diet rich in isoflavones or soy may have a lower risk of endometrial and ovarian cancer (Eden, 2012; Qu et al., 2014; Zhang et al., 2015).
The risk assessment for peri‐ and post‐menopausal women taking isoflavone‐containing food supplements performed by EFSA (EFSA ANS Panel, 2015) also evaluated the possible association between the intake of isoflavones from food supplements and harmful effects on the uterus. EFSA reported that there was no study on the association between isoflavone intake and risk of uterine cancer in the target population. As surrogate markers, data on endometrial thickness reported in 25 interventional controlled studies and on histopathology of the endometrium presented in nine interventional controlled studies were evaluated (EFSA ANS Panel, 2015). No significant adverse effects were observed in any of the human intervention studies, at dose levels up to 150 mg isoflavone·day−1 for 2.5 years. Thirteen studies in animals investigated cell proliferation in the uterus, and 22 animal studies studied uterus histopathological changes. No adverse effects were seen in any of the studies. EFSA concluded that the human and animal studies support that in post‐menopausal women adverse effects on the uterus were not noted for soy isoflavones, but that in the absence of data on the effect on uterine cancer, it was not possible to draw conclusions on effects of isoflavones on the uterus in peri‐menopausal women (Horn‐Ross et al., 2002; Haselkorn et al., 2003).
Effects of phytoestrogens on the thyroid and thyroid function
The soy isoflavones daidzein and genistein are reported to inhibit thyroid peroxidase (TPO) in vitro (EC number 1.11.1.8), an enzyme involved in the synthesis of T3 and T4 (Divi et al., 1997). Also in rats in vivo, daidzein and genistein inhibit TPO activity; however, no other adverse effects on thyroid functions could be observed (Chang and Doerge, 2000; Doerge and Sheehan, 2002). In addition, oestrogens are implied to have indirect effects on thyroid function, which has led to the concern that phytoestrogens may adversely affect thyroid function. However, human studies on the effect of soy isoflavones on thyroid function reviewed by EFSA (EFSA ANS Panel, 2015) are not conclusive. Some studies suggest that risk factors, such as iodine deficiency and subclinical hypothyroidism, might increase susceptibility of individuals to potential adverse effects of soy isoflavones on thyroid function (Doerge and Sheehan, 2002; Sathyapalan et al., 2011).
Effects of phytoestrogens on brain function
Most studies on the effect of phytoestrogens on neurological endpoints are conducted with soy isoflavones, while few studies also addressed the effects of dietary lignans. The rationale behind such studies is that oestrogens play an important role in brain health. The brain controls oestrogen release through the hypothalamus–pituitary–gonadal axis and also responds to oestrogens. In addition, oestradiol plays a key role in the neurobiology of ageing, because endocrine and neural senescence overlap in time and are mechanistically intertwined in complex feedback loops (Morrison et al., 2006). For oestrogen therapy, there are conflicting results on whether protective or detrimental effects on brain health are exerted. The reasons for the different observations appear to lie in the time at which oestrogen therapy is initiated, the neurological status of the brain at the time of oestrogen therapy initiation and the type of therapy used (Brinton, 2004). Interestingly, the loss of especially ERα seems to be associated with reduced neuroprotection by E2, and there appears to be a beneficial role for ERα in the aging brain (Schreihofer and Ma, 2013). A literature review on the neuroprotective effects of phytoestrogens found in soy reports that, while it has been demonstrated in animal research and cell culture studies that phytoestrogens from soy can exert neuroprotective effects, clinical trials and observational studies in humans have produced inconclusive findings (Soni et al., 2014). In this review, seven observational studies were reviewed; of which, three found positive effects of isoflavones on cognition (Hogervorst et al., 2008; Hogervorst et al., 2011; Greendale et al., 2012). Out of the 12 randomized control trials reviewed, six reported beneficial effects of isoflavone treatment on cognitive function (File et al., 2001; Duffy et al., 2003; Kritz‐Silverstein et al., 2003; Casini et al., 2006; Gleason et al., 2009; Thorp et al., 2009). Despite positive findings of studies investigating phytoestrogens and cognition, approximately half of the reviewed studies demonstrate negative or null effects (Soni et al., 2014). Just like for oestrogen therapy, also for phytoestrogens, there are many factors that have an effect on the outcomes, like age, gender, ethnicity, and menopausal status, as well as duration of consumption, and the cognitive test used. One important characteristic of the study population is the metabolic capacity to produce equol, which differs between Asian and non‐Asian study populations, and generally declines with age (Soni et al., 2014).
Consumption of soy or soy isoflavones has been shown to affect various endpoints related to neuronal health in rats (Zeng et al., 2004; Azcoitia et al., 2006; Huang and Zhang, 2010; Neese et al., 2010; Pan et al., 2010; Bagheri et al., 2012), and in mice (Zhao and Brinton, 2009; Bansal and Parle, 2010; Zhao et al., 2011; Yao et al., 2013), while consumption of large doses are also reported to have negative effects on the brain of rats (Choi and Lee, 2004). It is of interest to note that the protective effects of genistein on the cerebral cortex of ageing rats were suggested to have a different mode of action than that of oestradiol (Moran et al., 2013).
Different lignans have shown positive effects on cognitive performance and markers of Alzheimer's disease in mice (Um et al., 2009; Giridharan et al., 2011; Hu et al., 2012; Jeong et al., 2013; Li et al., 2014; Mao et al., 2015). In observational studies in humans, higher dietary intake of lignans is associated with better cognitive function (Franco et al., 2005; Kreijkamp‐Kaspers et al., 2007; Greendale et al., 2012; Nooyens et al., 2015), while coumestrol ingestion was unrelated to cognitive performance (Greendale et al., 2012).
Altogether, the data presented on possible beneficial effects of phytoestrogens on neurological health are inconclusive and seem to be affected by various factors. Furthermore, it is not clear, whether the observed beneficial effects are due to an oestrogen‐like mode of action, or whether other mechanisms underlie these effects.
Discussion
Many health effects including both benefits and risks have been related to exposure to phytoestrogens. Reported benefits include a lowered risk of menopausal symptoms, cardiovascular disease, breast cancer, other forms of cancer including prostate cancer, bowel cancer, uterine cancer and other cancers, and brain function disorders. On the other hand, phytoestrogens are also considered endocrine disruptors, indicating that they have the potential to cause adverse health effects such as infertility and increased risks of cancer in oestrogen‐sensitive organs. These adverse effects have been mainly suggested based on data from in vitro, animal or epidemiological studies. Clinical studies often report the absence of adverse effects. Tempfer et al. (2009) reported a meta‐analysis on possible side effects of phytoestrogens. Based on 174 randomized controlled trials, it was concluded that phytoestrogen supplements have a safe side effect profile with moderately elevated rates of gastrointestinal side effects. It is important to note, however, that these studies were generally not designed to study the safety of the phytoestrogens.
Consequently, the question of whether phytoestrogens are beneficial or harmful to human health remains of importance. The present overview reveals that the answer is rather complex and may depend on age, health status, and even the presence or absence of specific gut microflora in the population of concern. To further illustrate this complexity, Figure 5 presents a cartoon summarizing the possible health effects of phytoestrogens and the potential underlying modes of action as presented in the present paper.
Figure 5.
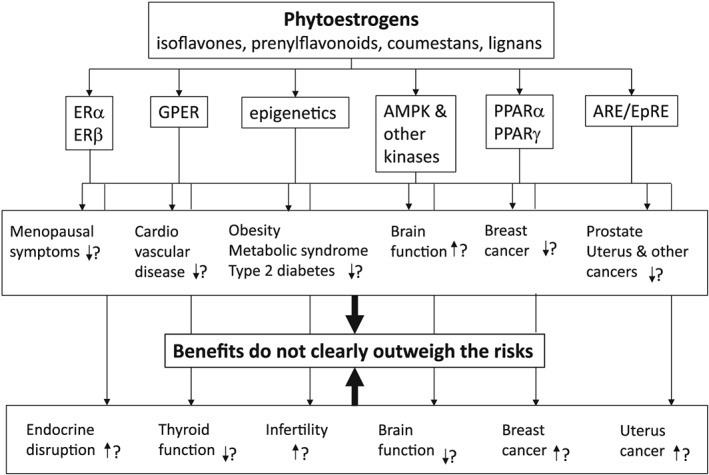
Schematic presentation summarizing the possible health effects of phytoestrogens and the potential underlying modes of action as presented in the present paper.
Given the rapid increase in global consumption of phytoestrogens and the fact that phytoestrogens are present in a wide range of dietary food supplements and widely marketed as a natural alternative to oestrogen replacement therapy, further insight into the risks and benefits of these phytoestrogens seems essential. Additional issues to be taken into account in the near future include (i) effects on children since phytoestrogens may be present in for example soy based infant foods; (ii) the role of the gut microbiota in phytoestrogen metabolism and differences in this microbial metabolism and the consequences for the health effects; (iii) the role of possible other modes of action than the oestrogenic activity in the health effects of phytoestrogens; (iv) the physiological levels of the phytoestrogens and their metabolites and how these relate to concentrations required in in vitro cellular models to actually induce the different effects; (v) the role of polymorphisms in the differential biological effects of phytoestrogens and the possible contribution of ‐omics technologies to elucidate these effects; and (vi) the fact that most phytoestrogens are weak oestrogens with anti‐oestrogenic effects when they compete with endogenous oestrogens for binding to the ERs, but pro‐oestrogenic effects in the absence of endogenous oestrogens and uncertain effects in the presence of low concentrations of endogenous oestrogens as occurs in the menopause.
The current review presents an overview of the potential health benefits of dietary phytoestrogens. It is of interest to put the observations within the guidance developed by EFSA for performing risk–benefit assessments of food. Given that in Europe food supplements are considered food this seems appropriate. EFSA recommended a stepwise approach for the risk–benefit assessment including first an assessment of whether the health risks clearly outweigh the health benefits or vice versa. The literature overview presented in this paper illustrates that at the current state‐of‐the‐art the beneficial health effects are not so obvious that they clearly outweigh the possible health risks. This implies that a further refinement of the assessment would be required, aiming at providing semi‐quantitative or quantitative estimates of risks and benefits at relevant exposures by using common metrics, and a comparison of risks and benefits using a composite metric such as disability‐adjusted life years or quality‐adjusted life years to express the outcome of the risk–benefit assessment as a single net health impact value. The currently available data set is not sufficient to support such a refined (semi) quantitative analysis. Taking all together, it can be concluded that several potential health benefits of phytoestrogens have been reported but that, given the data on potential adverse health effects, the current evidence on these beneficial health effects is not so obvious that they clearly outweigh the possible health risks. This implies that a definite conclusion on possible beneficial health effects of phytoestrogens cannot be made.
Author contributions
All authors contributed equally to writing the review.
Conflict of interest
The authors declare no conflicts of interest.
Supporting information
Table S1 binding affinity of E2 and phytoestrogens to ERα and ERβ (expressed as IC50 values from competitive binding assays).
Table S2 ERα‐ and ERβ‐ mediated gene expression induced by E2 and phytoestrogens (expressed as EC50 values from reporter gene assays).
Supporting info item
Rietjens, I. M. C. M. , Louisse, J. , and Beekmann, K. (2017) The potential health effects of dietary phytoestrogens. British Journal of Pharmacology, 174: 1263–1280. doi: 10.1111/bph.13622.
References
- Adlercreutz H (1995). Phytoestrogens: epidemiology and a possible role in cancer protection. Environ Health Perspect 103 (Suppl 7): 103–112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adlercreutz H (2002). Phyto‐oestrogens and cancer. Lancet Oncol 3: 364–373. [DOI] [PubMed] [Google Scholar]
- Adlercreutz H (2007). Lignans and human health. Crit Rev Clin Lab Sci 44: 483–525. [DOI] [PubMed] [Google Scholar]
- Akaza H, Miyanaga N, Takashima N, Naito S, Hirao Y, Tsukamoto T et al. (2004). Comparisons of percent equol producers between prostate cancer patients and controls: case‐controlled studies of isoflavones in Japanese, Korean and American residents. Jpn J Clin Oncol 34: 86–89. [DOI] [PubMed] [Google Scholar]
- Alexander SPH, Davenport AP, Kelly E, Marrion N, Peters JA, Benson HE et al. (2015a). The Concise Guide to PHARMACOLOGY 2015/16: G protein‐coupled receptors. Br J Pharmacol 172: 5744–5869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Fabbro D, Kelly E, Marrion N, Peters JA, Benson HE et al. (2015b). The Concise Guide to PHARMACOLOGY 2015/16: Enzymes. Br J Pharmacol 172: 6024–6109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander SPH, Cidlowski JA, Kelly E, Marrion N, Peters JA, Benson HE et al. (2015c). The Concise Guide to PHARMACOLOGY 2015/16: Nuclear hormone receptors. Br J Pharmacol 172: 5956–5978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali S, Coombes RC (2000). Estrogen receptor alpha in human breast cancer: occurrence and significance. J Mammary Gland Biol Neoplasia 5: 271–281. [DOI] [PubMed] [Google Scholar]
- Anderson LN, Cotterchio M, Boucher BA, Kreiger N (2013). Phytoestrogen intake from foods, during adolescence and adulthood, and risk of breast cancer by estrogen and progesterone receptor tumor subgroup among Ontario women. Int J Cancer 132: 1683–1692. [DOI] [PubMed] [Google Scholar]
- Andres S, Abraham K, Appel KE, Lampen A (2011). Risks and benefits of dietary isoflavones for cancer. Crit Rev Toxicol 41: 463–506. [DOI] [PubMed] [Google Scholar]
- Atkinson C, Warren RM, Sala E, Dowsett M, Dunning AM, Healey CS et al. (2004). Red‐clover‐derived isoflavones and mammographic breast density: a double‐blind, randomized, placebo‐controlled trial [ISRCTN42940165]. Breast Cancer Res 6: R170–R179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azcoitia I, Moreno A, Carrero P, Palacios S, Garcia‐Segura LM (2006). Neuroprotective effects of soy phytoestrogens in the rat brain. Gynecol Endocrinol 22: 63–69. [DOI] [PubMed] [Google Scholar]
- Bagheri M, Roghani M, Joghataei MT, Mohseni S (2012). Genistein inhibits aggregation of exogenous amyloid‐beta(1)(−)(4)(0) and alleviates astrogliosis in the hippocampus of rats. Brain Res 1429: 145–154. [DOI] [PubMed] [Google Scholar]
- Bansal N, Parle M (2010). Soybean supplementation helps reverse age‐ and scopolamine‐induced memory deficits in mice. J Med Food 13: 1293–1300. [DOI] [PubMed] [Google Scholar]
- Bardin A, Boulle N, Lazennec G, Vignon F, Pujol P (2004). Loss of ER beta expression as a common step in estrogen‐dependent tumor progression. Endocr Relat Cancer 11: 537–551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnes S, Peterson TG, Coward L (1995). Rationale for the use of genistein‐containing soy matrices in chemoprevention trials for breast and prostate‐cancer. J Cell Biochem : 181–187. [DOI] [PubMed] [Google Scholar]
- Beekmann K, de Haan LH, Actis‐Goretta L, Houtman R, van Bladeren PJ, Rietjens IM (2015). The effect of glucuronidation on isoflavone induced estrogen receptor (ER)alpha and ERbeta mediated coregulator interactions. J Steroid Biochem Mol Biol 154: 245–253. [DOI] [PubMed] [Google Scholar]
- Bennetts HW, Underwood EJ, Shier FL (1946). A specific breeding problem of sheep on subterranean clover pastures in Western Australia. Aust Vet J 22: 2–12. [DOI] [PubMed] [Google Scholar]
- Bhathena SJ, Velasquez MT (2002). Beneficial role of dietary phytoestrogens in obesity and diabetes. Am J Clin Nutr 76: 1191–1201. [DOI] [PubMed] [Google Scholar]
- Bolanos R, Del Castillo A, Francia J (2010). Soy isoflavones versus placebo in the treatment of climacteric vasomotor symptoms: systematic review and meta‐analysis. Menopause (New York, NY) 17: 660–666. [PubMed] [Google Scholar]
- Bonkhoff H, Fixemer T, Hunsicker I, Remberger K (1999). Estrogen receptor expression in prostate cancer and premalignant prostatic lesions. Am J Pathol 155: 641–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boucher BA, Cotterchio M, Anderson LN, Kreiger N, Kirsh VA, Thompson LU (2013). Use of isoflavone supplements is associated with reduced postmenopausal breast cancer risk. Int J Cancer 132: 1439–1450. [DOI] [PubMed] [Google Scholar]
- Boue SM, Burow ME, Wiese TE, Shih BY, Elliott S, Carter‐Wientjes CH et al. (2011). Estrogenic and antiestrogenic activities of phytoalexins from red kidney bean (Phaseolus vulgaris L.). J Agric Food Chem 59: 112–120. [DOI] [PubMed] [Google Scholar]
- Bovee TF, Helsdingen RJ, Rietjens IM, Keijer J, Hoogenboom RL (2004). Rapid yeast estrogen bioassays stably expressing human estrogen receptors alpha and beta, and green fluorescent protein: a comparison of different compounds with both receptor types. J Steroid Biochem Mol Biol 91: 99–109. [DOI] [PubMed] [Google Scholar]
- Brasky TM, Lampe JW, Potter JD, Patterson RE, White E (2010). Specialty supplements and breast cancer risk in the VITamins and Lifestyle (VITAL) Cohort. Cancer Epidemiol Biomarkers Prev 19: 1696–1708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brinton RD (2004). Impact of estrogen therapy on Alzheimer's disease: a fork in the road? CNS Drugs 18: 405–422. [DOI] [PubMed] [Google Scholar]
- Buck K, Zaineddin AK, Vrieling A, Linseisen J, Chang‐Claude J (2010). Meta‐analyses of lignans and enterolignans in relation to breast cancer risk. Am J Clin Nutr 92: 141–153. [DOI] [PubMed] [Google Scholar]
- Buck K, Zaineddin AK, Vrieling A, Heinz J, Linseisen J, Flesch‐Janys D et al. (2011). Estimated enterolignans, lignan‐rich foods, and fibre in relation to survival after postmenopausal breast cancer. Br J Cancer 105: 1151–1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cano A, Garcia‐Perez MA, Tarin JJ (2010). Isoflavones and cardiovascular disease. Maturitas 67: 219–226. [DOI] [PubMed] [Google Scholar]
- Casanova M, You L, Gaido KW, Archibeque‐Engle S, Janszen DB, Heck HA (1999). Developmental effects of dietary phytoestrogens in Sprague–Dawley rats and interactions of genistein and daidzein with rat estrogen receptors alpha and beta in vitro. Toxicol Sci 51: 236–244. [DOI] [PubMed] [Google Scholar]
- Casini ML, Marelli G, Papaleo E, Ferrari A, D'Ambrosio F, Unfer V (2006). Psychological assessment of the effects of treatment with phytoestrogens on postmenopausal women: a randomized, double‐blind, crossover, placebo‐controlled study. Fertil Steril 85: 972–978. [DOI] [PubMed] [Google Scholar]
- Cederroth CR, Nef S (2009). Soy, phytoestrogens and metabolism: A review. Mol Cell Endocrinol 304: 30–42. [DOI] [PubMed] [Google Scholar]
- Chang HC, Doerge DR (2000). Dietary genistein inactivates rat thyroid peroxidase in vivo without an apparent hypothyroid effect. Toxicol Appl Pharmacol 168: 244–252. [DOI] [PubMed] [Google Scholar]
- Chang EC, Charn TH, Park SH, Helferich WG, Komm B, Katzenellenbogen JA et al. (2008). Estrogen receptors alpha and beta as determinants of gene expression: influence of ligand, dose, and chromatin binding. Mol Endocrinol 22: 1032–1043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen M, Rao Y, Zheng Y, Wei S, Li Y, Guo T et al. (2014). Association between soy isoflavone intake and breast cancer risk for pre‐ and post‐menopausal women: a meta‐analysis of epidemiological studies. PLoS One 9: e89288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen MN, Lin CC, Liu CF (2015). Efficacy of phytoestrogens for menopausal symptoms: a meta‐analysis and systematic review. Climacteric 18: 260–269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng G, Wilczek B, Warner M, Gustafsson JA, Landgren BM (2007). Isoflavone treatment for acute menopausal symptoms. Menopause (New York, N.Y.) 14: 468–473. [DOI] [PubMed] [Google Scholar]
- Choi EJ, Lee BH (2004). Evidence for genistein mediated cytotoxicity and apoptosis in rat brain. Life Sci 75: 499–509. [DOI] [PubMed] [Google Scholar]
- Chrzan BG, Bradford PG (2007). Phytoestrogens activate estrogen receptor beta1 and estrogenic responses in human breast and bone cancer cell lines. Mol Nutr Food Res 51: 171–177. [DOI] [PubMed] [Google Scholar]
- Chu WL, Shiizaki K, Kawanishi M, Kondo M, Yagi T (2009). Validation of a new yeast‐based reporter assay consisting of human estrogen receptors alpha/beta and coactivator SRC‐1: application for detection of estrogenic activity in environmental samples. Environ Toxicol 24: 513–521. [DOI] [PubMed] [Google Scholar]
- Colacurci N, De Franciscis P, Atlante M, Mancino P, Monti M, Volpini G et al. (2013). Endometrial, breast and liver safety of soy isoflavones plus lactobacillus sporogenes in post‐menopausal women. Gynecol Endocrinol 29: 209–212. [DOI] [PubMed] [Google Scholar]
- Colditz GA, Willett WC, Stampfer MJ, Rosner B, Speizer FE, Hennekens CH (1987). Menopause and the risk of coronary heart disease in women. N Engl J Med 316: 1105–1110. [DOI] [PubMed] [Google Scholar]
- Crespillo A, Alonso M, Vida M, Pavon FJ, Serrano A, Rivera P et al. (2011). Reduction of body weight, liver steatosis and expression of stearoyl‐CoA desaturase 1 by the isoflavone daidzein in diet‐induced obesity. Br J Pharmacol 164: 1899–1915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Angelis M, Stossi F, Waibel M, Katzenellenbogen BS, Katzenellenbogen JA (2005). Isocoumarins as estrogen receptor beta selective ligands: isomers of isoflavone phytoestrogens and their metabolites. Bioorg Med Chem 13: 6529–6542. [DOI] [PubMed] [Google Scholar]
- de Kleijn MJJ, van der Schouw YT, Wilson PWF, Grobbee DE, Jacques PF (2002). Dietary intake of phytoestrogens is associated with a favorable metabolic cardiovascular risk profile in postmenopausal US women: the Framingham study. J Nutr 132: 276–282. [DOI] [PubMed] [Google Scholar]
- Delmanto A, Nahas‐Neto J, Traiman P, Uemura G, Pessoa EC, Nahas EA (2013). Effects of soy isoflavones on mammographic density and breast parenchyma in postmenopausal women: a randomized, double‐blind, placebo‐controlled clinical trial. Menopause (New York, N.Y.) 20: 1049–1054. [DOI] [PubMed] [Google Scholar]
- Dhooghe L, Naessens T, Heyerick A, De Keukeleire D, Vlietinck AJ, Pieters L et al. (2010). Quantification of xanthohumol, isoxanthohumol, 8‐prenylnaringenin, and 6‐prenylnaringenin in hop extracts and derived capsules using secondary standards. Talanta 83: 448–456. [DOI] [PubMed] [Google Scholar]
- Ding D, Xu L, Fang H, Hong H, Perkins R, Harris S et al. (2010). The EDKB: an established knowledge base for endocrine disrupting chemicals. BMC Bioinformatics 11 (Suppl 6): S5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Divi RL, Chang HC, Doerge DR (1997). Anti‐thyroid isoflavones from soybean: isolation, characterization, and mechanisms of action. Biochem Pharmacol 54: 1087–1096. [DOI] [PubMed] [Google Scholar]
- Djiogue S, Halabalaki M, Njamen D, Kretzschmar G, Lambrinidis G, Hoepping J et al. (2014). Erythroidine alkaloids: a novel class of phytoestrogens. Planta Med 80: 861–869. [DOI] [PubMed] [Google Scholar]
- Doerge DR, Sheehan DM (2002). Goitrogenic and estrogenic activity of soy isoflavones. Environ Health Perspect 110 (Suppl 3): 349–353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dolinoy DC, Weidman JR, Waterland RA, Jirtle RL (2006). Maternal genistein alters coat color and protects A(vy) mouse offspring from obesity by modifying the fetal epigenome. Environ Health Persp 114: 567–572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong JY, Qin LQ (2011). Soy isoflavones consumption and risk of breast cancer incidence or recurrence: a meta‐analysis of prospective studies. Breast Cancer Res Treat 125: 315–323. [DOI] [PubMed] [Google Scholar]
- Dornstauder E, Jisa E, Unterrieder I, Krenn L, Kubelka W, Jungbauer A (2001). Estrogenic activity of two standardized red clover extracts (Menoflavon) intended for large scale use in hormone replacement therapy. J Steroid Biochem Mol Biol 78: 67–75. [DOI] [PubMed] [Google Scholar]
- Dubey RK, Gillespie DG, Imthurn B, Rosselli M, Jackson EK, Keller PJ (1999). Phytoestrogens inhibit growth and MAP kinase activity in human aortic smooth muscle cells. Hypertension 33: 177–182. [DOI] [PubMed] [Google Scholar]
- Duffy R, Wiseman H, File SE (2003). Improved cognitive function in postmenopausal women after 12 weeks of consumption of a soya extract containing isoflavones. Pharmacol Biochem Behav 75: 721–729. [DOI] [PubMed] [Google Scholar]
- Eden JA (2012). Phytoestrogens for menopausal symptoms: a review. Maturitas 72: 157–159. [DOI] [PubMed] [Google Scholar]
- EFSA ANS Panel (2015). Scientific opinion on the risk assessment for peri‐ and post‐menopausal women taking food supplements containing isolated isoflavones. EFSA J 13: 342. [Google Scholar]
- EFSA NDA Panel (2012). Scientific opinion on the substantiation of health claims related to soy isoflavones and maintenance of bone mineral density (ID 1655) and reduction of vasomotor symptoms associated with menopause (ID 1654, 1704, 2140, 3093, 3154, 3590) (further assessment) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J 10: 2847. [Google Scholar]
- Eisenbrand G, Senate Commission on Food Safety of the German Research F (2007). Isoflavones as phytoestrogens in food supplements and dietary foods for special medical purposes. Opinion of the Senate Commission on Food Safety (SKLM) of the German Research Foundation (DFG)‐(shortened version). Mol Nutr Food Res 51: 1305–1312. [DOI] [PubMed] [Google Scholar]
- Enmark E, Pelto‐Huikko M, Grandien K, Lagercrantz S, Lagercrantz J, Fried G et al. (1997). Human estrogen receptor beta‐gene structure, chromosomal localization, and expression pattern. J Clin Endocrinol Metab 82: 4258–4265. [DOI] [PubMed] [Google Scholar]
- Escande A, Pillon A, Servant N, Cravedi JP, Larrea F, Muhn P et al. (2006). Evaluation of ligand selectivity using reporter cell lines stably expressing estrogen receptor alpha or beta. Biochem Pharmacol 71: 1459–1469. [DOI] [PubMed] [Google Scholar]
- Evers NM, van den Berg JH, Wang S, Melchers D, Houtman R, de Haan LH et al. (2014a). Cell proliferation and modulation of interaction of estrogen receptors with coregulators induced by ERalpha and ERbeta agonists. J Steroid Biochem Mol Biol 143: 376–385. [DOI] [PubMed] [Google Scholar]
- Evers NM, Wang S, van den Berg JH, Houtman R, Melchers D, de Haan LH et al. (2014b). Identification of coregulators influenced by estrogen receptor subtype specific binding of the ER antagonists 4‐hydroxytamoxifen and fulvestrant. Chem Biol Interact 220: 222–230. [DOI] [PubMed] [Google Scholar]
- Fang K, Dong H, Wang H, Gong J, Huang W, Lu F (2016). Soy isoflavones and glucose metabolism in menopausal women: A systematic review and meta‐analysis of randomized controlled trials. Mol Nutr Food res 60: 1602–1614. [DOI] [PubMed] [Google Scholar]
- File SE, Jarrett N, Fluck E, Duffy R, Casey K, Wiseman H (2001). Eating soya improves human memory. Psychopharmacology (Berl) 157: 430–436. [DOI] [PubMed] [Google Scholar]
- Franco OH, Burger H, Lebrun CEI, Peeters PHM, Lamberts SWJ, Grobbee DE et al. (2005). Higher dietary intake of lignans is associated with better cognitive performance in postmesopausal women. J Nutr 135: 1190–1195. [DOI] [PubMed] [Google Scholar]
- Fritz H, Seely D, Flower G, Skidmore B, Fernandes R, Vadeboncoeur S et al. (2013). Soy, red clover, and isoflavones and breast cancer: a systematic review. PLoS One 8: e81968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gennari L, Merlotti D, De Paola V, Calabro A, Becherini L, Martini G et al. (2005). Estrogen receptor gene polymorphisms and the genetics of osteoporosis: a HuGE review. Am J Epidemiol 161: 307–320. [DOI] [PubMed] [Google Scholar]
- Giridharan VV, Thandavarayan RA, Sato S, Ko KM, Konishi T (2011). Prevention of scopolamine‐induced memory deficits by schisandrin B, an antioxidant lignan from Schisandra chinensis in mice. Free Radic Res 45: 950–958. [DOI] [PubMed] [Google Scholar]
- Gleason CE, Carlsson CM, Barnet JH, Meade SA, Setchell KD, Atwood CS et al. (2009). A preliminary study of the safety, feasibility and cognitive efficacy of soy isoflavone supplements in older men and women. Age Ageing 38: 86–93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzalez Canete N, Duran Aguero S (2014). Soya isoflavones and evidences on cardiovascular protection. Nutr Hosp 29: 1271–1282. [DOI] [PubMed] [Google Scholar]
- Goodman MT, Shvetsov YB, Wilkens LR, Franke AA, Le Marchand L, Kakazu KK et al. (2009). Urinary phytoestrogen excretion and postmenopausal breast cancer risk: the multiethnic cohort study. Cancer Prev Res (Phila) 2: 887–894. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gorodeski GI (1994). Impact of the menopause on the epidemiology and risk factors of coronary artery heart disease in women. Exp Gerontol 29: 357–375. [DOI] [PubMed] [Google Scholar]
- Greendale GA, Huang MH, Leung K, Crawford SL, Gold EB, Wight R et al. (2012). Dietary phytoestrogen intakes and cognitive function during the menopausal transition: results from the study of women's health across the nation phytoestrogen study. Menopause (New York, N.Y.) 19: 894–903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greendale GA, Tseng CH, Han W, Huang MH, Leung K, Crawford S et al. (2015). Dietary isoflavones and bone mineral density during midlife and the menopausal transition: cross‐sectional and longitudinal results from the Study of Women's Health Across the Nation Phytoestrogen Study. Menopause (New York, N.Y.) 22: 279–288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gutendorf B, Westendorf J (2001). Comparison of an array of in vitro assays for the assessment of the estrogenic potential of natural and synthetic estrogens, phytoestrogens and xenoestrogens. Toxicology 166: 79–89. [DOI] [PubMed] [Google Scholar]
- Hargreaves DF, Potten CS, Harding C, Shaw LE, Morton MS, Roberts SA et al. (1999). Two‐week dietary soy supplementation has an estrogenic effect on normal premenopausal breast. J Clin Endocr Metab 84: 4017–4024. [DOI] [PubMed] [Google Scholar]
- Harris HA (2007). Estrogen receptor‐beta: recent lessons from in vivo studies. Mol Endocrinol 21: 1–13. [DOI] [PubMed] [Google Scholar]
- Harris DM, Besselink E, Henning SM, Go VL, Heber D (2005). Phytoestrogens induce differential estrogen receptor alpha‐ or Beta‐mediated responses in transfected breast cancer cells. Exp Biol Med (Maywood) 230: 558–568. [DOI] [PubMed] [Google Scholar]
- Haselkorn T, Stewart SL, Horn‐Ross PL (2003). Why are thyroid cancer rates so high in southeast asian women living in the United States? The bay area thyroid cancer study. Canc Epidemiol Biomarkers Prev 12: 144–150. [PubMed] [Google Scholar]
- He J, Wang S, Zhou M, Yu W, Zhang Y, He X (2015). Phytoestrogens and risk of prostate cancer: a meta‐analysis of observational studies. World J Surg Oncol 13: 231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hilakivi‐Clarke L, Andrade JE, Helferich W (2010). Is soy consumption good or bad for the breast? J Nutr 140: 2326S–2334S. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hogervorst E, Sadjimim T, Yesufu A, Kreager P, Rahardjo TB (2008). High tofu intake is associated with worse memory in elderly Indonesian men and women. Dement Geriatr Cogn Disord 26: 50–57. [DOI] [PubMed] [Google Scholar]
- Hogervorst E, Mursjid F, Priandini D, Setyawan H, Ismael RI, Bandelow S et al. (2011). Borobudur revisited: soy consumption may be associated with better recall in younger, but not in older, rural Indonesian elderly. Brain Res 1379: 206–212. [DOI] [PubMed] [Google Scholar]
- Horn‐Ross PL, Hoggatt KJ, Lee MM (2002). Phytoestrogens and thyroid cancer risk: the San Francisco Bay area thyroid cancer study. Canc Epidemiol Biomarkers Prev 11: 43–49. [PubMed] [Google Scholar]
- Howes LG, Howes JB, Knight DC (2006). Isoflavone therapy for menopausal flushes: a systematic review and meta‐analysis. Maturitas 55: 203–211. [DOI] [PubMed] [Google Scholar]
- Hsu MH, Savas U, Lasker JM, Johnson EF (2011). Genistein, resveratrol, and 5‐aminoimidazole‐4‐carboxamide‐1‐beta‐D‐ribofuranoside induce cytochrome P450 4F2 expression through an AMP‐activated protein kinase‐dependent pathway. J Pharmacol Exp Ther 337: 125–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu D, Cao Y, He R, Han N, Liu Z, Miao L et al. (2012). Schizandrin, an antioxidant lignan from Schisandra chinensis, ameliorates Abeta1‐42‐induced memory impairment in mice. Oxid Med Cell Longev 2012: 721721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang YH, Zhang QH (2010). Genistein reduced the neural apoptosis in the brain of ovariectomised rats by modulating mitochondrial oxidative stress. Br J Nutr 104: 1297–1303. [DOI] [PubMed] [Google Scholar]
- Hughes CL Jr (1988). Phytochemical mimicry of reproductive hormones and modulation of herbivore fertility by phytoestrogens. Environ Health Perspect 78: 171–174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hullar MA, Lancaster SM, Li F, Tseng E, Beer K, Atkinson C et al. (2015). Enterolignan‐producing phenotypes are associated with increased gut microbial diversity and altered composition in premenopausal women in the United States. Canc Epidemiol Biomarkers Prev 24: 546–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang JT, Park IJ, Shin JI, Lee YK, Lee SK, Baik HW et al. (2005). Genistein, EGCG, and capsaicin inhibit adipocyte differentiation process via activating AMP‐activated protein kinase. Biochem Biophys Res Commun 338: 694–699. [DOI] [PubMed] [Google Scholar]
- Hwang YW, Kim SY, Jee SH, Kim YN, Nam CM (2009). Soy food consumption and risk of prostate cancer: a meta‐analysis of observational studies. Nutr Cancer 61: 598–606. [DOI] [PubMed] [Google Scholar]
- Ikeda K, Arao Y, Otsuka H, Nomoto S, Horiguchi H, Kato S et al. (2002). Terpenoids found in the umbelliferae family act as agonists/antagonists for ER(alpha) and ERbeta: differential transcription activity between ferutinine‐liganded ER(alpha) and ERbeta. Biochem Biophys Res Commun 291: 354–360. [DOI] [PubMed] [Google Scholar]
- Ingram D, Sanders K, Kolybaba M, Lopez D (1997). Case–control study of phyto‐oestrogens and breast cancer. Lancet 350: 990–994. [DOI] [PubMed] [Google Scholar]
- Islam MA, Bekele R, Van den Berg JH, Kuswanti Y, Thapa O, Soltani S et al. (2015). Deconjugation of soy isoflavone glucuronides needed for estrogenic activity. Toxicol In Vitro 29: 706–715. [DOI] [PubMed] [Google Scholar]
- Jacobs A, Wegewitz U, Sommerfeld C, Grossklaus R, Lampen A (2009). Efficacy of isoflavones in relieving vasomotor menopausal symptoms – a systematic review. Mol Nutr Food Res 53: 1084–1097. [DOI] [PubMed] [Google Scholar]
- Jeong EJ, Lee HK, Lee KY, Jeon BJ, Kim DH, Park JH et al. (2013). The effects of lignan‐riched extract of Shisandra chinensis on amyloid‐beta‐induced cognitive impairment and neurotoxicity in the cortex and hippocampus of mouse. J Ethnopharmacol 146: 347–354. [DOI] [PubMed] [Google Scholar]
- Jungbauer A, Medjakovic S (2014). Phytoestrogens and the metabolic syndrome. J Steroid Biochem Mol Biol 139: 277–289. [DOI] [PubMed] [Google Scholar]
- Kang NJ, Lee KW, Rogozin EA, Cho YY, Heo YS, Bode AM et al. (2007). Equol, a metabolite of the soybean isoflavone daidzein, inhibits neoplastic cell transformation by targeting the MEK/ERK/p90RSK/activator protein‐1 pathway. J Biol Chem 282: 32856–32866. [DOI] [PubMed] [Google Scholar]
- Karahalil B (2005). Benefits and Risks of Phytoestrogens In: Yildiz F. (ed). Phytoestrogens In Functional Foods. CRC Press: Boca Raton, FL, USA, pp. 210–211. [Google Scholar]
- Khan SA, Chatterton RT, Michel N, Bryk M, Lee O, Ivancic D et al. (2012). Soy isoflavone supplementation for breast cancer risk reduction: a randomized phase II trial. Cancer Prev Res (Phila) 5: 309–319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim S‐H, Kim S‐H, Kim Y‐B, Jeon Y‐T, Lee S‐C, Song Y‐S (2009). Genistein inhibits cell growth by modulating various mitogen‐activated protein kinases and AKT in cervical cancer cells. Ann N Y Acad Sci 1171: 495–500. [DOI] [PubMed] [Google Scholar]
- King RA, Bursill DB (1998). Plasma and urinary kinetics of the isoflavones daidzein and genistein after a single soy meal in humans. Am J Clin Nutr 67: 867–872. [DOI] [PubMed] [Google Scholar]
- Ko KP, Park SK, Park B, Yang JJ, Cho LY, Kang C et al. (2010). Isoflavones from phytoestrogens and gastric cancer risk: a nested case–control study within the Korean Multicenter Cancer Cohort. Canc Epidemiol Biomarkers Prev 19: 1292–1300. [DOI] [PubMed] [Google Scholar]
- Kokubo Y, Iso H, Ishihara J, Okada K, Inoue M, Tsugane S et al. (2007). Association of dietary intake of soy, beans, and isoflavones with risk of cerebral and myocardial infarctions in Japanese populations: the Japan Public Health Center‐based (JPHC) study cohort I. Circulation 116: 2553–2562. [DOI] [PubMed] [Google Scholar]
- Koo J, Cabarcas‐Petroski S, Petrie JL, Diette N, White RJ, Schramm L (2015). Induction of proto‐oncogene BRF2 in breast cancer cells by the dietary soybean isoflavone daidzein. BMC Cancer 15: 905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Korde L, Fears T, Wu A, Nomura A, West D, Pike M et al. (2004). Adolescent and childhood soy intake and breast cancer risk in Asian‐American women. Breast Cancer Res Tr 88: S149–S149. [DOI] [PubMed] [Google Scholar]
- Korde LA, Wu AH, Fears T, Nomura AM, West DW, Kolonel LN et al. (2009). Childhood soy intake and breast cancer risk in Asian American women. Canc Epidemiol Biomarkers Prev 18: 1050–1059. [DOI] [PubMed] [Google Scholar]
- Kreijkamp‐Kaspers S, Kok L, Grobbee DE, de Haan EH, Aleman A, Lampe JW et al. (2004). Effect of soy protein containing isoflavones on cognitive function, bone mineral density, and plasma lipids in postmenopausal women: a randomized controlled trial. JAMA 292: 65–74. [DOI] [PubMed] [Google Scholar]
- Kreijkamp‐Kaspers S, Kok L, Grobbee DE, de Haan EH, Aleman A, van der Schouw YT (2007). Dietary phytoestrogen intake and cognitive function in older women. J Gerontol A Biol Sci Med Sci 62: 556–562. [DOI] [PubMed] [Google Scholar]
- Kritz‐Silverstein D, Von Muhlen D, Barrett‐Connor E, Bressel MA (2003). Isoflavones and cognitive function in older women: the SOy and Postmenopausal Health in Aging (SOPHIA) Study. Menopause (New York, N.Y.) 10: 196–202. [DOI] [PubMed] [Google Scholar]
- Kuijsten A, Arts ICW, Vree TB, Hollman PCH (2005). Pharmacokinetics of enterolignans in healthy men and women consuming a single dose of secoisolariciresinol diglucoside. J Nutr 135: 795–801. [DOI] [PubMed] [Google Scholar]
- Kuiper GGJM, Enmark E, PeltoHuikko M, Nilsson S, Gustafsson JA (1996). Cloning of a novel estrogen receptor expressed in rat prostate and ovary. P Natl Acad Sci USA 93: 5925–5930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuiper GG, Lemmen JG, Carlsson B, Corton JC, Safe SH, van der Saag PT et al. (1998). Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 139: 4252–4263. [DOI] [PubMed] [Google Scholar]
- Kurahashi N, Iwasaki M, Sasazuki S, Otani T, Inoue M, Tsugane S et al. (2007). Soy product and isoflavone consumption in relation to prostate cancer in Japanese men. Canc Epidemiol Biomarkers Prev 16: 538–545. [DOI] [PubMed] [Google Scholar]
- Kwack SJ, Kim KB, Kim HS, Yoon KS, Lee BM (2009). Risk assessment of soybean‐based phytoestrogens. J Toxicol Environ Health A 72: 1254–1261. [DOI] [PubMed] [Google Scholar]
- Lahiry P, Torkamani A, Schork NJ, Hegele RA (2010). Kinase mutations in human disease: interpreting genotype–phenotype relationships. Nat Rev Genet 11: 60–74. [DOI] [PubMed] [Google Scholar]
- Lampe JW (2003). Isoflavonoid and lignan phytoestrogens as dietary biomarkers. J Nutr 133: 956S–964S. [DOI] [PubMed] [Google Scholar]
- Lee SA, Shu XO, Li H, Yang G, Cai H, Wen W et al. (2009). Adolescent and adult soy food intake and breast cancer risk: results from the Shanghai Women's Health Study. Am J Clin Nutr 89: 1920–1926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lethaby AE, Brown J, Marjoribanks J, Kronenberg F, Roberts H, Eden J (2007). Phytoestrogens for vasomotor menopausal symptoms. Cochrane Database Syst Rev : Cd001395. [DOI] [PubMed] [Google Scholar]
- Lethaby A, Marjoribanks J, Kronenberg F, Roberts H, Eden J, Brown J (2013). Phytoestrogens for menopausal vasomotor symptoms. Cochrane Database Syst Rev: Cd001395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Zhao X, Xu X, Mao X, Liu Z, Li H et al. (2014). Schisantherin A recovers Abeta‐induced neurodegeneration with cognitive decline in mice. Physiol Behav 132: 10–16. [DOI] [PubMed] [Google Scholar]
- Liu X, Nam JW, Song YS, Viswanath AN, Pae AN, Kil YS et al. (2014). Psoralidin, a coumestan analogue, as a novel potent estrogen receptor signaling molecule isolated from Psoralea corylifolia . Bioorg Med Chem Lett 24: 1403–1406. [DOI] [PubMed] [Google Scholar]
- Lyn‐Cook BD, Blann E, Payne PW, Bo J, Sheehan D, Medlock K (1995). Methylation profile and amplification of proto‐oncogenes in rat pancreas induced with phytoestrogens. Proc Soc Exp Biol Med 208: 116–119. [DOI] [PubMed] [Google Scholar]
- Maffini MV, Rubin BS, Sonnenschein C, Soto AM (2006). Endocrine disruptors and reproductive health: the case of bisphenol‐A. Mol Cell Endocrinol 254‐255: 179–186. [DOI] [PubMed] [Google Scholar]
- Magee PJ (2011). Is equol production beneficial to health? Proc Nutr Soc 70: 10–18. [DOI] [PubMed] [Google Scholar]
- Manning G, Whyte DB, Martinez R, Hunter T, Sudarsanam S (2002). The protein kinase complement of the human genome. Science (New York, NY) 298: 1912–1934. [DOI] [PubMed] [Google Scholar]
- Mao X, Liao Z, Guo L, Xu X, Wu B, Xu M et al. (2015). Schisandrin C ameliorates learning and memory deficits by abeta‐induced oxidative stress and neurotoxicity in mice. Phytother Res 29: 1373–1380. [DOI] [PubMed] [Google Scholar]
- Marini H, Minutoli L, Polito F, Bitto A, Altavilla D, Atteritano M et al. (2007). Effects of the phytoestrogen genistein on bone metabolism in osteopenic postmenopausal women: a randomized trial. Ann Intern Med 146: 839–847. [DOI] [PubMed] [Google Scholar]
- Marini H, Bitto A, Altavilla D, Burnett BP, Polito F, Di Stefano V et al. (2008). Breast safety and efficacy of genistein aglycone for postmenopausal bone loss: a follow‐up study. J Clin Endocrinol Metab 93: 4787–4796. [DOI] [PubMed] [Google Scholar]
- Maskarinec G, Verheus M, Steinberg FM, Amato P, Cramer MK, Lewis RD et al. (2009). Various doses of soy isoflavones do not modify mammographic density in postmenopausal women. J Nutr 139: 981–986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mense SM, Hei TK, Ganju RK, Bhat HK (2008). Phytoestrogens and breast cancer prevention: possible mechanisms of action. Environ Health Perspect 116: 426–433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Messina M (2008). Conclusion that isoflavones exert estrogenic effects on breast tissue and may raise breast cancer risk unfounded. Mol Nutr Food Res 52: 299–300. [DOI] [PubMed] [Google Scholar]
- Molzberger AF, Soukup ST, Kulling SE, Diel P (2013). Proliferative and estrogenic sensitivity of the mammary gland are modulated by isoflavones during distinct periods of adolescence. Arch Toxicol 87: 1129–1140. [DOI] [PubMed] [Google Scholar]
- Morabito N, Crisafulli A, Vergara C, Gaudio A, Lasco A, Frisina N et al. (2002). Effects of genistein and hormone‐replacement therapy on bone loss in early postmenopausal women: a randomized double‐blind placebo‐controlled study. J Bone Miner Res 17: 1904–1912. [DOI] [PubMed] [Google Scholar]
- Moran J, Garrido P, Alonso A, Cabello E, Gonzalez C (2013). 17beta‐Estradiol and genistein acute treatments improve some cerebral cortex homeostasis aspects deteriorated by aging in female rats. Exp Gerontol 48: 414–421. [DOI] [PubMed] [Google Scholar]
- Morrison JH, Brinton RD, Schmidt PJ, Gore AC (2006). Estrogen, menopause, and the aging brain: how basic neuroscience can inform hormone therapy in women. J Neurosci 26: 10332–10348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mueller SO, Simon S, Chae K, Metzler M, Korach KS (2004). Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells. Toxicol Sci 80: 14–25. [DOI] [PubMed] [Google Scholar]
- Nagata C (2000). Ecological study of the association between soy product intake and mortality from cancer and heart disease in Japan. Int J Epidemiol 29: 832–836. [DOI] [PubMed] [Google Scholar]
- Nedrow A, Miller J, Walker M, Nygren P, Huffman LH, Nelson HD (2006). Complementary and alternative therapies for the management of menopause‐related symptoms: a systematic evidence review. Arch Intern Med 166: 1453–1465. [DOI] [PubMed] [Google Scholar]
- Neese SL, Korol DL, Katzenellenbogen JA, Schantz SL (2010). Impact of estrogen receptor alpha and beta agonists on delayed alternation in middle‐aged rats. Horm Behav 58: 878–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G et al. (2001). Mechanisms of estrogen action. Physiol Rev 81: 1535–1565. [DOI] [PubMed] [Google Scholar]
- Nooyens AC, Milder IE, van Gelder BM, Bueno‐de‐Mesquita HB, van Boxtel MP, Verschuren WM (2015). Diet and cognitive decline at middle age: the role of antioxidants. Br J Nutr 113: 1410–1417. [DOI] [PubMed] [Google Scholar]
- Nott SL, Huang Y, Fluharty BR, Sokolov AM, Huang M, Cox C et al. (2008). Do estrogen receptor beta polymorphisms play a role in the pharmacogenetics of estrogen signaling? Curr Pharmacogenomics Person Med 6: 239–259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NTP (2008). Toxicology and carcinogenesis studies of genistein (Cas No. 446‐72‐0) in Sprague–Dawley rats (feed study). Natl Toxicol Program Tech Rep Ser : 1–240. [PubMed] [Google Scholar]
- Obi N, Chang‐Claude J, Berger J, Braendle W, Slanger T, Schmidt M et al. (2009). The use of herbal preparations to alleviate climacteric disorders and risk of postmenopausal breast cancer in a German case–control study. Canc Epidemiol Biomarkers Prev 18: 2207–2213. [DOI] [PubMed] [Google Scholar]
- Ogawa S, Inoue S, Watanabe T, Orimo A, Hosoi T, Ouchi Y et al. (1998). Molecular cloning and characterization of human estrogen receptor beta cx: a potential inhibitor of estrogen action in human. Nucleic Acids Res 26: 3505–3512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ozasa K, Nakao M, Watanabe Y, Hayashi K, Miki T, Mikami K et al. (2004). Serum phytoestrogens and prostate cancer risk in a nested case–control study among Japanese men. Cancer Sci 95: 65–71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pan M, Li Z, Yeung V, Xu RJ (2010). Dietary supplementation of soy germ phytoestrogens or estradiol improves spatial memory performance and increases gene expression of BDNF, TrkB receptor and synaptic factors in ovariectomized rats. Nutr Metab (Lond) 7: 75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park CE, Yun H, Lee EB, Min BI, Bae H, Choe W et al. (2010). The antioxidant effects of genistein are associated with AMP‐activated protein kinase activation and PTEN induction in prostate cancer cells. J Med Food 13: 815–820. [DOI] [PubMed] [Google Scholar]
- Park J, Kim d H, Ahn HN, Song YS, Lee YJ, Ryu JH (2012). Activation of estrogen receptor by bavachin from Psoralea corylifolia . Biomol Ther (Seoul) 20: 183–188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker WH, Broder MS, Chang E, Feskanich D, Farquhar C, Liu ZM et al. (2009). Ovarian conservation at the time of hysterectomy and long‐term health outcomes in the nurses' health study. Obstet Gynecol 113: 1027–1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patisaul HB, Jefferson W (2010). The pros and cons of phytoestrogens. Front Neuroendocrinol 31: 400–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearce ST, Jordan VC (2004). The biological role of estrogen receptors alpha and beta in cancer. Crit Rev Oncol Hematol 50: 3–22. [DOI] [PubMed] [Google Scholar]
- Penttinen P, Jaehrling J, Damdimopoulos AE, Inzunza J, Lemmen JG, van der Saag P et al. (2007). Diet‐derived polyphenol metabolite enterolactone is a tissue‐specific estrogen receptor activator. Endocrinology 148: 4875–4886. [DOI] [PubMed] [Google Scholar]
- Persson I (2000). Estrogens in the causation of breast, endometrial and ovarian cancers – evidence and hypotheses from epidemiological findings. J Steroid Biochem Mol Biol 74: 357–364. [DOI] [PubMed] [Google Scholar]
- Petrakis NL, Barnes S, King EB, Lowenstein J, Wiencke J, Lee MM et al. (1996). Stimulatory influence of soy protein isolate on breast secretion in pre‐ and postmenopausal women. Canc Epidemiol Biomarkers Prev 5: 785–794. [PubMed] [Google Scholar]
- Powles TJ, Howell A, Evans DG, McCloskey EV, Ashley S, Greenhalgh R et al. (2008). Red clover isoflavones are safe and well tolerated in women with a family history of breast cancer. Menopause Int 14: 6–12. [DOI] [PubMed] [Google Scholar]
- Prossnitz ER, Barton M (2011). The G‐protein‐coupled estrogen receptor GPER in health and disease. Nat Rev Endocrinol 7: 715–726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pudenz M, Roth K, Gerhauser C (2014). Impact of soy isoflavones on the epigenome in cancer prevention. Nutrients 6: 4218–4272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin W, Zhu W, Shi H, Hewett JE, Ruhlen RL, MacDonald RS et al. (2009). Soy isoflavones have an antiestrogenic effect and alter mammary promoter hypermethylation in healthy premenopausal women. Nutr Cancer 61: 238–244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qu XL, Fang Y, Zhang M, Zhang YZ (2014). Phytoestrogen intake and risk of ovarian cancer: a meta‐ analysis of 10 observational studies. Asian Pac J Cancer Prev: APJCP 15: 9085–9091. [DOI] [PubMed] [Google Scholar]
- Rebbeck TR, Troxel AB, Norman S, Bunin GR, DeMichele A, Baumgarten M et al. (2007). A retrospective case–control study of the use of hormone‐related supplements and association with breast cancer. Int J Cancer 120: 1523–1528. [DOI] [PubMed] [Google Scholar]
- Remely M, de la Garza AL, Magnet U, Aumueller E, Haslberger AG (2015a). Obesity: epigenetic regulation ‐ recent observations. Biomol Concepts 6: 163–175. [DOI] [PubMed] [Google Scholar]
- Remely M, Lovrecic L, de la Garza AL, Migliore L, Peterlin B, Milagro FI et al. (2015b). Therapeutic perspectives of epigenetically active nutrients. Br J Pharmacol 172: 2756–2768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rickard DJ, Monroe DG, Ruesink TJ, Khosla S, Riggs BL, Spelsberg TC (2003). Phytoestrogen genistein acts as an estrogen agonist on human osteoblastic cells through estrogen receptors alpha and beta. J Cell Biochem 89: 633–646. [DOI] [PubMed] [Google Scholar]
- Rietjens IM, Sotoca AM, Vervoort J, Louisse J (2013). Mechanisms underlying the dualistic mode of action of major soy isoflavones in relation to cell proliferation and cancer risks. Mol Nutr Food Res 57: 100–113. [DOI] [PubMed] [Google Scholar]
- Rimoldi G, Christoffel J, Seidlova‐Wuttke D, Jarry H, Wuttke W (2007). Effects of chronic genistein treatment in mammary gland, uterus, and vagina. Environ Health Perspect 115 (Suppl 1): 62–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rossouw JE, Prentice RL, Manson JE, Wu L, Barad D, Barnabei VM et al. (2007). Postmenopausal hormone therapy and risk of cardiovascular disease by age and years since menopause. J Womens Health (Larchmt) 16: 927–928. [DOI] [PubMed] [Google Scholar]
- Saarinen NM, Abrahamsson A, Dabrosin C (2010). Estrogen‐induced angiogenic factors derived from stromal and cancer cells are differently regulated by enterolactone and genistein in human breast cancer in vivo. Int J Cancer 127: 737–745. [DOI] [PubMed] [Google Scholar]
- Sathyapalan T, Manuchehri AM, Thatcher NJ, Rigby AS, Chapman T, Kilpatrick ES et al. (2011). The effect of soy phytoestrogen supplementation on thyroid status and cardiovascular risk markers in patients with subclinical hypothyroidism: a randomized, double‐blind, crossover study. J Clin Endocrinol Metab 96: 1442–1449. [DOI] [PubMed] [Google Scholar]
- Scarlata S, Miksicek R (1995). Binding properties of coumestrol to expressed human estrogen receptor. Mol Cell Endocrinol 115: 65–72. [DOI] [PubMed] [Google Scholar]
- Scherr FF, Sarmah AK, Di HJ, Cameron KC (2009). Degradation and metabolite formation of 17beta‐estradiol‐3‐sulphate in New Zealand pasture soils. Environ Int 35: 291–297. [DOI] [PubMed] [Google Scholar]
- Schreihofer DA, Ma Y (2013). Estrogen receptors and ischemic neuroprotection: who, what, where, and when? Brain Res 1514: 107–122. [DOI] [PubMed] [Google Scholar]
- Setchell KD (1998). Phytoestrogens: the biochemistry, physiology, and implications for human health of soy isoflavones. Am J Clin Nutr 68: 1333S–1346S. [DOI] [PubMed] [Google Scholar]
- Setchell KD, Faughnan MS, Avades T, Zimmer‐Nechemias L, Brown NM, Wolfe BE et al. (2003). Comparing the pharmacokinetics of daidzein and genistein with the use of 13C‐labeled tracers in premenopausal women. Am J Clin Nutr 77: 411–419. [DOI] [PubMed] [Google Scholar]
- Severson RK, Nomura AMY, Grove JS, Stemmermann GN (1989). A prospective‐study of demographics, diet, and prostate‐cancer among men of Japanese ancestry in Hawaii. Cancer Res 49: 1857–1860. [PubMed] [Google Scholar]
- Shimazu T, Inoue M, Sasazuki S, Iwasaki M, Sawada N, Yamaji T et al. (2011). Plasma isoflavones and the risk of lung cancer in women: a nested case–control study in Japan. Canc Epidemiol Biomarkers Prev 20: 419–427. [DOI] [PubMed] [Google Scholar]
- Singh V, Sharma P, Capalash N (2013). DNA methyltransferase‐1 inhibitors as epigenetic therapy for cancer. Curr Cancer Drug Targets 13: 379–399. [DOI] [PubMed] [Google Scholar]
- Soni M, Rahardjo TB, Soekardi R, Sulistyowati Y, Lestariningsih Y‐UA et al. (2014). Phytoestrogens and cognitive function: a review. Maturitas 77: 209–220. [DOI] [PubMed] [Google Scholar]
- Sotoca AM, van den Berg H, Vervoort J, van der Saag P, Strom A, Gustafsson JA et al. (2008a). Influence of cellular ERalpha/ERbeta ratio on the ERalpha‐agonist induced proliferation of human T47D breast cancer cells. Toxicol Sci 105: 303–311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sotoca AM, Ratman D, van der Saag P, Strom A, Gustafsson JA, Vervoort J et al. (2008b). Phytoestrogen‐mediated inhibition of proliferation of the human T47D breast cancer cells depends on the ERalpha/ERbeta ratio. J Steroid Biochem Mol Biol 112: 171–178. [DOI] [PubMed] [Google Scholar]
- Southan C, Sharman JL, Benson HE, Faccenda E, Pawson AJ, Alexander SP et al. (2016). The IUPHAR/BPS Guide to PHARMACOLOGY in 2016: towards curated quantitative interactions between 1300 protein targets and 6000 ligands. Nucl. Acids Res. 44: D1054–D1068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stafford HA (1997). Roles of flavonoids in symbiotic and defense functions in legume roots. Bot Rev 63: 27–39. [Google Scholar]
- Stevens JF, Page JE (2004). Xanthohumol and related prenylflavonoids from hops and beer: to your good health! Phytochemistry 65: 1317–1330. [DOI] [PubMed] [Google Scholar]
- Stossi F, Barnett DH, Frasor J, Komm B, Lyttle CR, Katzenellenbogen BS (2004). Transcriptional profiling of estrogen‐regulated gene expression via estrogen receptor (ER) alpha or ERbeta in human osteosarcoma cells: distinct and common target genes for these receptors. Endocrinology 145: 3473–3486. [DOI] [PubMed] [Google Scholar]
- Strom A, Hartman J, Foster JS, Kietz S, Wimalasena J, Gustafsson JA (2004). Estrogen receptor beta inhibits 17beta‐estradiol‐stimulated proliferation of the breast cancer cell line T47D. Proc Natl Acad Sci U S A 101: 1566–1571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Struja T, Richard A, Linseisen J, Eichholzer M, Rohrmann S (2014). The association between urinary phytoestrogen excretion and components of the metabolic syndrome in NHANES. Eur J Nutr 53: 1371–1381. [DOI] [PubMed] [Google Scholar]
- Takeuchi S, Takahashi T, Sawada Y, Iida M, Matsuda T, Kojima H (2009). Comparative study on the nuclear hormone receptor activity of various phytochemicals and their metabolites by reporter gene assays using Chinese hamster ovary cells. Biol Pharm Bull 32: 195–202. [DOI] [PubMed] [Google Scholar]
- Taku K, Melby MK, Takebayashi J, Mizuno S, Ishimi Y, Omori T et al. (2010). Effect of soy isoflavone extract supplements on bone mineral density in menopausal women: meta‐analysis of randomized controlled trials. Asia Pac J Clin Nutr 19: 33–42. [PubMed] [Google Scholar]
- Taku K, Melby MK, Kronenberg F, Kurzer MS, Messina M (2012). Extracted or synthesized soybean isoflavones reduce menopausal hot flash frequency and severity: systematic review and meta‐analysis of randomized controlled trials. Menopause (New York, N.Y.) 19: 776–790. [DOI] [PubMed] [Google Scholar]
- Taylor CK, Levy RM, Elliott JC, Burnett BP (2009). The effect of genistein aglycone on cancer and cancer risk: a review of in vitro, preclinical, and clinical studies. Nutr Rev 67: 398–415. [DOI] [PubMed] [Google Scholar]
- Tempfer CB, Bentz EK, Leodolter S, Tscherne G, Reuss F, Cross HS et al. (2007). Phytoestrogens in clinical practice: a review of the literature. Fertil Steril 87: 1243–1249. [DOI] [PubMed] [Google Scholar]
- Tempfer CB, Froese G, Heinze G, Bentz EK, Hefler LA, Huber JC (2009). Side effects of phytoestrogens: a meta‐analysis of randomized trials. Am J Med 122: 939–946.e939. [DOI] [PubMed] [Google Scholar]
- ter Veld MG, Schouten B, Louisse J, van Es DS, van der Saag PT, Rietjens IM et al. (2006). Estrogenic potency of food‐packaging‐associated plasticizers and antioxidants as detected in ERalpha and ERbeta reporter gene cell lines. J Agric Food Chem 54: 4407–4416. [DOI] [PubMed] [Google Scholar]
- Thomas C, Gustafsson JA (2011). The different roles of ER subtypes in cancer biology and therapy. Nat Rev Cancer 11: 597–608. [DOI] [PubMed] [Google Scholar]
- Thorp AA, Sinn N, Buckley JD, Coates AM, Howe PR (2009). Soya isoflavone supplementation enhances spatial working memory in men. Br J Nutr 102: 1348–1354. [DOI] [PubMed] [Google Scholar]
- Tiosano D, Paris F, Grimaldi M, Georgescu V, Servant N, Hochberg Z et al. (2014). Evidence of ERalpha and ERbeta selectivity and partial estrogen agonism in traditional Chinese medicine. Reprod Biol Endocrinol 12: 97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tokede OA, Onabanjo TA, Yansane A, Gaziano JM, Djousse L (2015). Soya products and serum lipids: a meta‐analysis of randomised controlled trials. Br J Nutr 114: 831–843. [DOI] [PubMed] [Google Scholar]
- Trock BJ, Hilakivi‐Clarke L, Clarke R (2006). Meta‐analysis of soy intake and breast cancer risk. J Natl Cancer Inst 98: 459–471. [DOI] [PubMed] [Google Scholar]
- Um MY, Ahn JY, Kim S, Kim MK, Ha TY (2009). Sesaminol glucosides protect beta‐amyloid peptide‐induced cognitive deficits in mice. Biol Pharm Bull 32: 1516–1520. [DOI] [PubMed] [Google Scholar]
- Valsta LM, Kilkkinen A, Mazur W, Nurmi T, Lampi AM, Ovaskainen ML et al. (2003). Phyto‐oestrogen database of foods and average intake in Finland. Br J Nutr 89 (Suppl 1): S31–S38. [DOI] [PubMed] [Google Scholar]
- van der Schouw YT, Kreijkamp‐Kaspers S, Peeters PH, Keinan‐Boker L, Rimm EB, Grobbee DE (2005). Prospective study on usual dietary phytoestrogen intake and cardiovascular disease risk in Western women. Circulation 111: 465–471. [DOI] [PubMed] [Google Scholar]
- van Die MD, Bone KM, Williams SG, Pirotta MV (2014). Soy and soy isoflavones in prostate cancer: a systematic review and meta‐analysis of randomized controlled trials. BJU Int 113: E119–E130. [DOI] [PubMed] [Google Scholar]
- Vanden Berghe W, Haegeman G (2008). Beneficial effects of soy isoflavones focus on epigenetic and anti‐inflammatory effects. Agro Food Ind Hi Tech 19: 57–60. [Google Scholar]
- Velentzis LS, Cantwell MM, Cardwell C, Keshtgar MR, Leathem AJ, Woodside JV (2009). Lignans and breast cancer risk in pre‐ and post‐menopausal women: meta‐analyses of observational studies. Br J Cancer 100: 1492–1498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Verheus M, van Gils CH, Kreijkamp‐Kaspers S, Kok L, Peeters PH, Grobbee DE et al. (2008). Soy protein containing isoflavones and mammographic density in a randomized controlled trial in postmenopausal women. Canc Epidemiol Biomarkers Prev 17: 2632–2638. [DOI] [PubMed] [Google Scholar]
- Verkasalo PK, Appleby PN, Allen NE, Davey G, Adlercreutz H, Key TJ (2001). Soya intake and plasma concentrations of daidzein and genistein: validity of dietary assessment among eighty British women (Oxford arm of the European Prospective Investigation into Cancer and Nutrition). Br J Nutr 86: 415–421. [DOI] [PubMed] [Google Scholar]
- Wang Q, Li H, Tao P, Wang YP, Yuan P, Yang CX et al. (2011). Soy isoflavones, CYP1A1, CYP1B1, and COMT polymorphisms, and breast cancer: a case–control study in southwestern China. DNA Cell Biol 30: 585–595. [DOI] [PubMed] [Google Scholar]
- Wang S, Aarts JM, Evers NM, Peijnenburg AA, Rietjens IM, Bovee TF (2012). Proliferation assays for estrogenicity testing with high predictive value for the in vivo uterotrophic effect. J Steroid Biochem Mol Biol 128: 98–106. [DOI] [PubMed] [Google Scholar]
- Warri A, Saarinen NM, Makela S, Hilakivi‐Clarke L (2008). The role of early life genistein exposures in modifying breast cancer risk. Br J Cancer 98: 1485–1493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe S, Uesugi S, Kikuchi Y (2002). Isoflavones for prevention of cancer, cardiovascular diseases, gynecological problems and possible immune potentiation. Biomed Pharmacother 56: 302–312. [DOI] [PubMed] [Google Scholar]
- Williams C, Edvardsson K, Lewandowski SA, Strom A, Gustafsson JA (2008). A genome‐wide study of the repressive effects of estrogen receptor beta on estrogen receptor alpha signaling in breast cancer cells. Oncogene 27: 1019–1032. [DOI] [PubMed] [Google Scholar]
- Wilson VS, Bobseine K, Gray LE Jr (2004). Development and characterization of a cell line that stably expresses an estrogen‐responsive luciferase reporter for the detection of estrogen receptor agonist and antagonists. Toxicol Sci 81: 69–77. [DOI] [PubMed] [Google Scholar]
- Wu AH, Yu MC, Tseng CC, Pike MC (2008). Epidemiology of soy exposures and breast cancer risk. Br J Cancer 98: 9–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wuttke W, Rimoldi G, Christoffel J, Seidlova‐Wuttke D (2006). Plant extracts for the treatment of menopausal women: Safe? Maturitas 55: S92–S100. [Google Scholar]
- Wuttke W, Jarry H, Seidlova‐Wuttke D (2007). Isoflavones‐‐safe food additives or dangerous drugs? Ageing Res Rev 6: 150–188. [DOI] [PubMed] [Google Scholar]
- Yan L, Spitznagel EL (2009). Soy consumption and prostate cancer risk in men: a revisit of a meta‐analysis. Am J Clin Nutr 89: 1155–1163. [DOI] [PubMed] [Google Scholar]
- Yan GR, Xiao CL, He GW, Yin XF, Chen NP, Cao Y et al. (2010). Global phosphoproteomic effects of natural tyrosine kinase inhibitor, genistein, on signaling pathways. Proteomics 10: 976–986. [DOI] [PubMed] [Google Scholar]
- Yang WS, Va P, Wong MY, Zhang HL, Xiang YB (2011). Soy intake is associated with lower lung cancer risk: results from a meta‐analysis of epidemiologic studies. Am J Clin Nutr 94: 1575–1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao J, Zhao L, Mao Z, Chen S, Wong KC, To J et al. (2013). Potentiation of brain mitochondrial function by S‐equol and R/S‐equol estrogen receptor beta‐selective phytoSERM treatments. Brain Res 1514: 128–141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoder SC, Lancaster SM, Hullar MAJ, Lampe JW (2015). Chapter 7 – Gut microbial metabolism of plant lignans: influence on human health A2 – Rio, Kieran TuohyDaniele Del In: Diet‐Microbe Interactions in the Gut. Academic Press: San Diego, pp. 103–117. [Google Scholar]
- Zeng H, Chen Q, Zhao B (2004). Genistein ameliorates beta‐amyloid peptide (25‐35)‐induced hippocampal neuronal apoptosis. Free Radic Biol Med 36: 180–188. [DOI] [PubMed] [Google Scholar]
- Zhang XL, Shu XO, Gao YT, Yang G, Li Q, Li HL et al. (2003). Soy food consumption is associated with lower risk of coronary heart disease in Chinese women. J Nutr 133: 2874–2878. [DOI] [PubMed] [Google Scholar]
- Zhang YB, Chen WH, Guo JJ, Fu ZH, Yi C, Zhang M et al. (2013). Soy isoflavone supplementation could reduce body weight and improve glucose metabolism in non‐Asian postmenopausal women – a meta‐analysis. Nutrition (Burbank, Los Angeles County, Calif) 29: 8–14. [DOI] [PubMed] [Google Scholar]
- Zhang GQ, Chen JL, Liu Q, Zhang Y, Zeng H, Zhao Y (2015). Soy intake is associated with lower endometrial cancer risk: a systematic review and meta‐analysis of observational studies. Medicine 94: e2281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao L, Brinton RD (2009). In search of estrogen alternatives for the brain In: Hormones, Cognition and Dementia: State of The Art and Emergent Therapeutic Strategies, Cambridge University Press: Cambridge: pp. 93–100. [Google Scholar]
- Zhao E, Mu Q (2011). Phytoestrogen biological actions on Mammalian reproductive system and cancer growth. Sci Pharm 79: 1–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao L, Mao Z, Schneider LS, Brinton RD (2011). Estrogen receptor beta‐selective phytoestrogenic formulation prevents physical and neurological changes in a preclinical model of human menopause. Menopause (New York, N.Y.) 18: 1131–1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1 binding affinity of E2 and phytoestrogens to ERα and ERβ (expressed as IC50 values from competitive binding assays).
Table S2 ERα‐ and ERβ‐ mediated gene expression induced by E2 and phytoestrogens (expressed as EC50 values from reporter gene assays).
Supporting info item