

Abstract
Regulation of biological processes occurs through complex, synergistic mechanisms. In this study, we discovered the synergistic orchestration of multiple mechanisms regulating the normal and diseased state (age related macular degeneration, AMD) in the retina. We uncovered gene networks with overlapping feedback loops that are modulated by nuclear hormone receptors (NHR), miRNAs, and epigenetic factors. We utilized a comprehensive filtering and pathway analysis strategy comparing miRNA and microarray data between three mouse models and human donor eyes (normal and AMD). The mouse models lack key NHRS (Nr2e3, RORA) or epigenetic (Ezh2) factors. Fifty-four total miRNAs were differentially expressed, potentially targeting over 150 genes in 18 major representative networks including angiogenesis, metabolism, and immunity. We identified sixty-eight genes and 5 miRNAS directly regulated by NR2E3 and/or RORA. After a comprehensive analysis, we discovered multimodal regulation by miRNA, NHRs, and epigenetic factors of three miRNAs (miR-466, miR1187, and miR-710) and two genes (Ell2 and Entpd1) that are also associated with AMD. These studies provide insight into the complex, dynamic modulation of gene networks as well as their impact on human disease, and provide novel data for the development of innovative and more effective therapeutics.
Introduction
Regulating gene expression is a fundamental mechanism used by cells to orchestrate the complex development of all tissues. This multi-tiered event is modulated by many processes including modification of DNA, regulation of pre- and post-transcripts, and protein modifications1–3. Gene regulation at the DNA level occurs through several mechanisms, including chromatin modification directed by DNA methylation, and noncoding RNA (ncRNA) or DNA-binding proteins4, 5. Transcription factors are the main contributors in regulating networks at the transcription level. Additionally, cells regulate how much mRNA is translated into proteins by modulating capping, splicing, addition of a Poly (A) tail, the sequence-specific nuclear export rates, and by sequestering the RNA transcript2, 6–8. The translation of mRNA is also controlled by a number of mechanisms at initiation or by mRNA silencing. In the initiation process, recruitment of small ribosomal subunits is modulated by mRNA secondary structure, antisense RNA binding, or protein binding while the translational repression and gene silencing is modulated by microRNAs (miRNAs)9–11. In this study, we discover the synergistic manner in which three modulators (miRNA, nuclear hormone receptors (NHRs), and epigenetic factors) influence the retina in a normal and diseased state.
miRNAs have recently emerged as an important class of post-transcriptional regulatory factors and play a crucial role in regulating gene expression in the retina12–14. miRNA coding sequence typically resides in intergenic regions or on the anti-sense strand of genes15, 16. miRNAs are self-sufficient, retain promoters and regulatory elements, and have the capacity to regulate their own expression. Recent studies revealed that there are more than 400 miRNAs expressed in the retina, and miRNA gene regulation has been shown to affect retinal development, function, and disease14, 17, 18. Previous studies have demonstrated significant differences between the expression profiles of miRNAs in the embryonic and adult retina18. These profiles suggest specific roles for miRNAs in the developing and mature retina. Different groups and clusters of miRNAs have been identified and are usually co-expressed under similar conditions19. Recent studies also demonstrate that variations in gene expression involving transcription factors or miRNAs are implicated in numerous retinal diseases14. miRNAs play an important role in the development and maintenance of photoreceptors cells. Therefore, lack or misexpression of miRNAs are associated with diseases such as retinitis pigmentosa, age-related macular degeneration (AMD), light-induced retinal degeneration, and Stargardt disease20–22. Complete loss of the miR-24a, for example, impacts the rate of apoptosis in retinal precursors, leading to a decrease in the size of the eye23. Global loss of retinal miRNAs by loss of Dicer expression has severe and detrimental effects on retinal development and physiology. Several loss of function studies reveal miRNAs are crucial regulators of differentiation and promote the survival of retinal neuronal progenitors14, 24, 25. In addition, altered miRNA expression has been demonstrated in rodent models of Retinitis Pigmentosa (RP), Induced Uveitis, Retinoblastoma, and Diabetic Retinopathy26–28. Consistent with these studies, retinas have a different expression miRNA profile than normal retinas29. Furthermore, it is increasingly evident that the two classes of gene modulators, transcription factors and miRNAs, work synergistically within given gene networks30–32.
Nuclear hormone receptors (NHRs) are part of a large superfamily of ligand-dependent steroid hormone receptors that often regulate transcription in a ligand-dependent manner and can function as activators or repressors of gene expression. NHRs regulate many biological processes to maintain cellular homeostasis and are significant regulators of retinal development, maintenance, and disease33–37. miRNAs are regulated by nuclear receptors via three main mechanisms: binding to the promoter region, indirect regulation by interacting with nuclear receptor target genes, and regulation of miRNA biogenesis38. Nuclear receptor class 2, subfamily e, member 3 (Nr2e3) and retinoic acid receptor-related orphan receptor alpha (Rora) are key regulators of retinal development and function33, 39–45. Mutations in NR2E3 are associated with several retinal diseases including enhanced S-cone syndrome, Goldman-Favre Syndrome, Clumped Pigmentary Retinal Degeneration, and Autosomal Recessive and Dominant Retinitis Pigmentosa46. Mice lacking Nr2e3, Nr2e3 rd7/rd7, rd7, exhibit a slow progressive retinal degeneration, characterized clinically by pan-retinal spotting, and histologically by whorls and rosettes due to excess production of blue opsin expressing cone cells followed by progressive degeneration of both rod and cone photoreceptor cells41, 47, 48.
Our prior studies show Nr2e3 regulates Rora 42 and both are implicated in AMD pathogenesis49–51. RORA is ubiquitously expressed and plays a role in lipid metabolism, bone morphogenesis, and photoreceptor development50. Studies of the knockout mouse model for Rora (Staggerer) show it is highly expressed in cerebellar Purkinje cells, and plays a crucial role in the CNS51. Staggerer mice are a model for human ataxia characterized by reduced number of Purkinje cells52, 53. In the retina, Rora is expressed in the ganglion cell layer and the inner nuclear layer of the adult retina54, 55, and regulates the expression of the cone opsin genes, Opn1sw and Opn1mw 45. More recently, our colleagues discovered Nr2e3 is upregulated in the conditional knockout of epigenetic factor Enhancer Of Zeste 2 Polycomb Repressive Complex 2 Subunit, Ezh2, suggesting a link between Nr2e3 and epigenetics54.
In addition to miRNA and NHR regulation, epigenetic events such as DNA methylation and post-translational modifications of the histones regulate gene expression. The marker histone-3, on lysine-27 (H3K27me3), trimethylates histones is a strong epigenetic regulator that serves to silence genes and is regulated by EZH256. EZH2 is a subunit of the Polycomb Repressor Complex2 (PRC2) and is involved in the regulation of a key development transcription factor family of genes including homeobox (HOX), SRY box (SOX), paired box (PAX) and wingless type (WNT). EZH2 functions in the PCR2 histone methyltransferase complex involved in the silencing of HOX genes during development57. In the PCR2 complex, EZH2 acts as a catalytic agent that methylates lysine 9 and 27 in histone H356, 58. EZH2 as well as other genes from the PCR2 complex induce the overexpression of E2F, a cell cycle regulator, in tumor cell lines59. EZH2 also interacts with retinoic acid (RA) to induce neural stem cell differentiation59–61. Interestingly, microarray data of Ezh2 −/− retinas detected upregulation of Nr2e3 consistent with a phenotype of suppression of neural proliferation and differentiation62. Ezh2 −/− mice exhibit retinal degeneration characterized by functional loss of photoreceptors, detected by loss of scotopic and photopic ERG responses, and histologically by retinal thinning at one month of age.
Macular degeneration is a leading cause of blindness yet there are no known genetic causes, and therapies are not effective on a significant number of patients. It is clear from many studies that gene expression and related disease occur via a dynamic and complex synergistic integration of multiple mechanisms of regulation. In this study, we examine the synergistic modulation of NHRs Nr2e3 and Rora, miRNAs, and the epigenetic factor Ezh2. As both NR2E3 and RORA have been implicated in AMD, we evaluated our miRNA potential gene targets as well as differentially expressed genes from each mouse model against gene expression data from a large cohort of AMD patients’ donor eyes from unrelated subjects. Lastly, we determined which of these differentially expressed genes and miRNAs are directly regulated by NR2E3 and/or RORA. This is the first comprehensive study to evaluate multiple mechanisms of gene regulation and determine their combined impact on biological pathways and human disease.
Results
The retina has progressive, dynamic miRNA expression
In this study, we employed a comprehensive analysis and filtering strategy to identify key miRNA and their target genes that are differentially expressed in rd7, regulated by Rora, Nr2e3, and Ezh2, and implicated in AMD (Fig. 1). A miRNA microarray analysis was conducted on retinas from Nr2e3 rd7/rd7 and control C57BL6/J mice at embryonic (E) 18.5 day and postnatal (P) days 6, 14, and 30 to capture the dynamic changes that occur in a developing retina to its maturity. Differential miRNA expression was observed across multiple time points, (Supplemental Figure S1). Interestingly, this heat map shows clear differences in the expression of miRNAs between the embryonic and adult points and postnatal day 6 (P6) illustrates a transition between development and maturity. In the initial analysis and filtering, redundant probes, erroneous control spots, and miRNAs with no known gene targets were removed from analysis and miRNAs differentially expressed in more than one time point were classified as “shared”. Using these criteria, 54 miRNAs were differentially expressed between Nr2e3 rd7/rd7 and C57BL6/J mice with a minimum 2-fold change and a p value of p < 0.001. Of the differentially expressed miRNAs, 37 miRNAs were unique to E18, 4 miRNAs were unique to P6, 3 miRNAs were unique to P14, and 10 miRNAs were shared between embryonic time point and one or multiple postnatal time points (Fig. 2A,B). Interestingly, while most of the miRNA that were differentially expressed at multiple time points were either consistently increased or decreased, four miRNAS showed altered pattern of expression at each time point they were differentially expressed (Fig. 2C). Previous studies of miRNA profiles expressed in the embryonic and adult retina were cross-referenced revealing common and unique miRNAs to our study17, 18. Additionally, prior studies from various models of retinal degeneration identified over 300 differentially expressed miRNAs63–90, a total of 16 common miRNAs were identified (miR-1187, miR-125b-5p, miR-331-3p, miR466d-3p, miR-467f, miR-542-3p, miR-574-5p, miR654-3p, miR669h-3p, miR-882, miR-342-3p, miR-466a-5p, miR-466d-5p, miR-706, miR-345-3p, miR532-5p). This study reveals 38 unique miRNAs that may be particular to the Nr2e3 regulated pathway. All 54 differentially expressed miRNAs were subsequently evaluated against the mirBase to determine all potential target genes for each miRNA (Table 1). Each miRNA can have up to 700–1500 potential target genes. Potential miRNA target genes were cross-referenced to previously published microarray expression data (GSE24512) of genes misexpressed in Nr2e3 rd7/rd7 retinas at E18 and P30.
Figure 1.
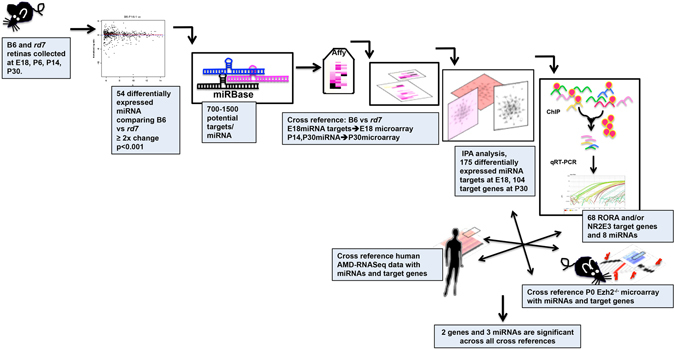
Comprehensive filtering strategy to identify multimodal regulation of miRNAs and their target genes. Step 1. 10 retinas per time point (E18, P6, P14, P30) were collected from B6 and rd7. Step 2. miRNA microarray was performed on samples from step 1. 54 differentially miRNAS were identified with 2-fold change, P < 0.001. Step 3. Subsequent filtering analysis of miRNA expression data was performed using BRB Array Tools along with Stanford Statistical Analysis of Microarrays (SAM) and evaluated using miRBase to identify their potential target genes. Step 4. miRNA targets were then cross-referenced with E18 microarray and P30 rd7 microarray data. Steps 5, 6. Genes that were found to be statistically significant were analyzed using Ingenuity Pathway analysis to algorithmically generated gene networks in which the default cutoffs identify those genes that were significantly differentially regulated in the network. Step 7. Genes that were identified through the networks were screened for the nuclear receptor-binding site of Nr2e3 and RORA and further analyzed by chromatin immunoprecipitation. A total of 68 genes were direct targets of RORA and/or Nr2e3 as well as 8 miRNAs (out of the 54 differentially expressed miRNAs identified). The results obtained were then cross-referenced to AMD-RNAseq data and microarray data from the epigenetic factor Ezh2. This filtering strategy allowed us to identify 2 genes (Ell2 and Entpd1) as well as 3 miRNAs (mir466, miR1187 and miR710) that are regulated by epigenetic factors and nuclear hormone receptors and are associated with AMD pathogenesis.
Figure 2.
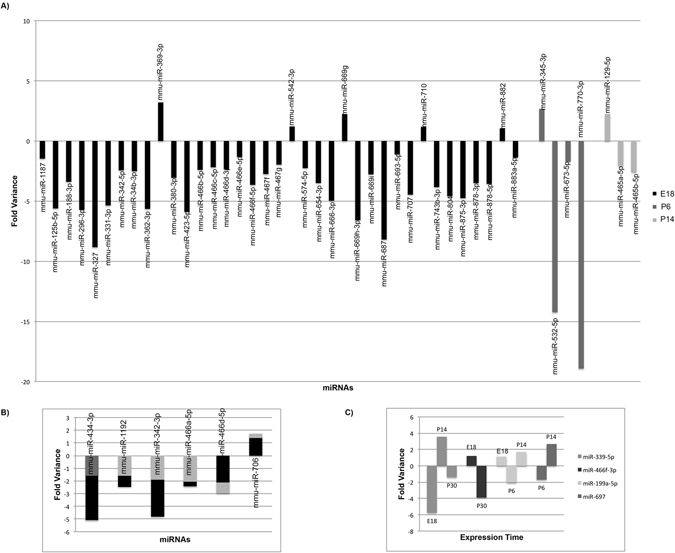
miRNAs differentially expressed between B6 (wildtype) and rd7 (mutant) retinas. (A) miRNAs that are differentially expressed at E18, P6, or P14 (B) miRNAs differentially expressed between the embryonic time point and one adult time point, and (C) miRNAs differentially expressed in more that one time point and have an alternate pattern of expression. Minimum 2 fold change and P < 0.001.
Table 1.
miRNAs differentially expressed in rd7 vs B6 and their potential target genes.
| Expression TP | miRNA | Target Genes |
|---|---|---|
| E18 | miR-1187 | Abcc9, Acpp, B3gnt9, Bnc2, Casp12, Cpm, Crhbp, Dpt, Egfl6, Entpd1, Fndc1, Fras1, Has2, Lcp1, Mylk4, Nox4, Otx1, Papss2, Peg3, Pgm5, Rgs5, Stard8, Stat6, Tek, Tfcp2l1, Vamp5, Wnt4 |
| miR-125b-5p | Acer2, Atp1b4, Bmf, C77080, Ehd4, Enpep, Ets1, Eva1a, Hapln1, Jade2, Mgll, Msrb3, Rhov, Serpinb5, Snx31, Ucp2 | |
| miR-188-3p | Alcam, Arhgef3, Atp1b4, Cd248, Cxcl12, Egln3, Ell2, Flrt2, Gbp7, Grhl2, Ifi202b, Itgb3, Itpripl2, Msrb3, Muc4, Myot, Scara5, Slc16a12, Smoc2 | |
| miR-296-3p | Epb4.1l1 | |
| miR-327 | Abca9, Arhgap36, Col18a1, Crim1, Cspg4, Cxcl12, Entpd1, Fbn2, Iigp1, Kcnq5, Lamp2, Ppfibp2, Ptprb, Ryr1, Thbs1, Tmem88 | |
| miR-331-3p | Mgll | |
| miR-342-5p | Ppargc1a | |
| miR-34b-3p | Fras1 | |
| miR-362-3p | Il13ra1, Mef2c, Ppp1r3b, Prdm16, Sspn, Unc45b | |
| miR-369-3p | Afap1l2, Aspn, Mylk4, Sema3d, She | |
| miR-380-3p | Dst | |
| miR-423-5p | Bmf, Ccnd2, Dlk1, Mgll, Myrf, Slc7a8 | |
| miR-466b-5p | Acer2, Fam46a, Ppargc1a, Tbx22 | |
| miR-466c-5p | Acer2, Fam46a, Ppargc1a, Tbx22 | |
| miR-466d-3p | Arap2, B3gnt9, Dmrta2, Dsg2, Dst, Ets1, Fam46a, Fgfr2, Gdf10, Jag1, Matn2, Piezo2, Sgms1, Stard8, Tbx22, Tgfbr3 | |
| miR-466e-5p | Abcc9, Acpp, B3gnt9, Bnc2, Casp12, Crhbp, Dpt, Egfl6, Entpd1, Fndc1, Fras1, Has2, Lcp1, Mylk4, Nox4, Otx1, Papss2, Peg3, Pgm5, Rgs5, Stard8, Stat6, Tek, Tfcp2l1, Vamp5, Wnt4 | |
| miR-466f-5p | Fzd4 | |
| miR-467f | Dmrta2, Fam46a, Fam84a, Fndc1, Hmcn1, Itpripl2, Jag1, Mef2c, Msrb3, Stard8, Tbx22, Tek, Thbs2, Trp63 | |
| miR-467g | Arap2, B3gnt9, Dmrta2, Dsg2, Dst, Ets1, Fam46a, Fgfr2, Fgl2, Gdf10, Jag1, Matn2, Piezo2, Sgms1, Stard8, Tbx22, Tgfbr3 | |
| miR-542-3p | Cgnl1, Fgl2, Jade2, Lpp, Ppp1r3b, Slc7a8, Sspn, Tgfbr3 | |
| miR-574-5p | Acpp, Fndc1, Mylk4, Ntrk2, Rgs5, Vcan | |
| miR-654-3p | Cd55, Lamc1, Ntrk2, Ppp1r3b, Thbs2 | |
| miR-666-3p | Bmp7, C77080, Ets1, Fam174b, Fzd4, Ptgfrn, Rab27a | |
| miR-669g | Ntrk2 | |
| miR-669h-3p | Dsc2, Fndc1, Foxc1, Frk, Tbx22 | |
| miR-669i | Cd36, Fgl2, Fndc1, Gatm, Tbx22 | |
| miR-687 | Crim1, Dsg2, Mfsd7c, Msrb3 | |
| miR-693-5p | Egfl6, Foxc1, Il13ra1, Mef2c, Papss2 | |
| miR-707 | Ehf | |
| miR-710 | Dsg2, Jag1, Lpar1, Mef2c | |
| miR-743b-3p | Acer2, Afap1l2, Bnc2, Cast, Ell2, Elovl7, Flrt2, Fndc3b, Lpp, Lrig1, Npr3, Ntrk2, Ptgfrn, Scara5, Sema3c, Tgfbr2 | |
| miR-804 | B3gnt9, Dpt, Foxc1, Osr1 | |
| miR-875-3p | Bnc2, Cd44, Ehf, Fam46a, Fgl2, Frk, Il13ra1, Lamc1, Mef2c, Otx1, Ptgfrn, Rbms1, Sema3c, Sgms2, Sspn, Tfcp2l1 | |
| miR-878-3p | Cgnl1, Elk3, Mef2c, Sema3c, Sema3d | |
| miR-878-5p | Ptprb, Trdn | |
| miR-882 | Atp1b4, Bmf, Dsc2, Fosl2, Mgll, Ntrk2, Rgs5, Sgms1, Tfcp2l1 | |
| miR-883a-5p | Bnc2, Eef2k, Fam174b, Lyz1 | |
| E18, P6 | miR-434-3p | Sulf1 |
| E18, P14 | miR-1192 | Bmp7, Boc, Ccbe1, Ccnd2, Cd44, Cd55, Col25a1, Dennd2c, Dmp1, Ehf, Ell2, Elovl7, Ets1, Fam101b, Fbn2, Flt1, Fosl2b, Foxc1, Fzd6, Hapln1, Itpripl2, Kdelc2, Lmo7, Lpp, Mef2c, Mylk4, Nrp2, Ntrk2, Pdlim7, Piezo2, Prickle1, Sema3c, Sgms1Sgms2, Slc7a8, Tbx18, Tgfbr3, Thbs2, Trp63, Unc45 |
| miR-342-3p | Ntrk2 | |
| miR-466a-5p | Abcc9, Acpp, B3gnt9, Bnc2, Casp12, Crhbp, Dpt, Egfl6, Entpd1, Fndc1, Fras1, Has2, Lcp1, Mylk4, Nox4, Otx1, Papss2, Peg3, Pgm5, Rgs5, Stard8, Stat6, Tek, Tfcp2l1, Vamp5, Wnt4 | |
| miR-466d-5p | Abcc9, Acpp, B3gnt9, Bmp7, Bnc2, Ccbe1, Cpm, Eef2k, Fam114a1, Fam174b, Fam84a, Has2, Lpp, Msrb3, Nrk, Osr1, Otx1, Papss2, Peg3, Pgm5, Rgs5, Slc7a8, Sspn, Stard8, Stat6, Sulf1, Vamp5, Wnt4 | |
| miR-706 | Atp1b4, Dpt, Has2, Il13ra1, Kdelc2, Mef2c, Mylk4, Ostf1, Sema3d, Sgms2 | |
| E18, P30 | miR-466f-3p | Adora2b, Col25a1, Fam174b, Fam46a, Fam84a, Fbn2, Fndc1, Hmcn1, Itpripl2, Jag1, Lpar1, Macc1, Matn2, Mef2c, Msrb3, Mylk4, Ntrk2, Otx1, Pdgfc, Sspn, Stard8, Tfcp2l1, Thbs2, Trp63 |
| E18, P6, P14 | miR-199a-5p | Afap1l2, Cd36, Cgnl1, Ctgf, Ets1, Fzd4, Fzd6, Hapln1, Hmcn1, Itga8, Lamc1, Lrrc173, Matn2, Mertk, Myrf, Ppargc1a, Scara, Sulf1, Zc3hav1 |
| E18, P14, P30 | miR-339-5p | Kdelc2, Lamc1, Sema3c, Sspn, Tfcp2l1 |
| P6 | miR-345-3p | Cpm, Eef2k |
| miR-532-5p | Alcam, Ets1, Gbp7, Itpripl2, Lpp, Ppp1r3b | |
| miR-770-3p | Hapln1, Ucp2 | |
| miR-673-5p | Abca1, Fzd4, Kdelc2, Nrk, Tfcp2l1 | |
| P6, P14 | miR-697 | Lamp2, Flt1, Rbsm1 |
| P14 | miR-129-5p | Flt1, Stat6 |
| miR-465a-5p | Abcc9, Acpp, B3gnt9, Bnc2, Casp12, Crhbp, Dpt, Egfl6, Entpd1, Fndc1, Fras1, Has2, Lcp1, Mylk4, Nox4, Otx1, Papss2, Peg3, Pgm5, Rgs5, Stard8, Stat6, Tek, Tfcp2l1, Vamp5, Wnt4 | |
| miR-465b-5p | Acer2, Fam46a, Ppargc1a, Tbx22 |
Genes identified through microarray data of differentially expressed genes in rd7 vs B6. Minimum 2 fold change and p < 0.05.
Identification of microRNA target genes and their gene networks
Once significantly differentiated miRNA were identified, the analysis focused on miRNAs differentially expressed at embryonic day E18.5 and postnatal day P30.5 for further analysis. These time points were chosen to represent miRNAs important in development when Nr2e3 is first expressed, and those miRNAs important to maintain normal function in the mature retina. Potential miRNA target genes at each of the two time points were then cross-referenced to microarray expression data91. To identify differentially expressed miRNA potential target genes showing biological significance, miRNA target genes were cross-referenced with microarray data mentioned above (GSE24512) and those exhibiting greater than 2-fold change of gene expression in Nr2e3 rd7/rd7 when compared to C57BL6/J mice were further analyzed. Duplicate genes (targeted by more than one miRNA within a time point) were counted once in the total number of unique miRNA target genes. This filtering strategy identified over 150 unique genes (Supplemental Table S1) which were further analyzed by Ingenuity Pathway Analysis (IPA). IPA analysis revealed eighteen unique gene networks including those for metabolism, development, immune response, cancer, and cellular function (Table 2). These putative miRNA targets are associated with over 50 diseases including diabetes, mellitus, cell proliferation of cancer, neuro-inflammation, apoptosis of neuroblastomas, and heart disease (Table 3). Figure 3 summarizes the combined networks and their common targets and pathways.
Table 2.
Ingenuity pathway analysis (IPA) of miRNA, RORA and NR2E3 target genes.
| E18 Network Function | Genes |
|---|---|
| Cardiovascular | ANTXR2, ARHGEF3, ASPN, B3GNT9, BMP7, CAST, DSC2, DSG2, EGLN3, EHF, ELL2, ETSS1, FAM46A, FGFR2, FOXC1, GATM, GRHL2, HAS2, JAG1, LAMC1, LAMP2, LRIG1, MGLL, MUC4, NTRK2, OSR1, PPARGC1A, PRDM16, PTPRB, RSPO1, RYR1, SEMA3C, SEMA3D, SGMS1, SLC7A8, TGFBR2, TGFBR3, THBS1, THBS2, TRDN, WNT4 |
| Immune Response/Injury | CD44, COL18A1, DPT, EGFL6, ELK3, ENTPD1, FGL2, FOSL2, FRAS1, KCNQ5, LPP, LYZ, MYRF, NPR3, OTX1, RAB27A, RGS5, VCAN |
| Cellular Function | ALCAM, ARAP2, BACH2, BMF, BNC2, CCND2, CD36, CRHBP, CSPG4, DLK1, DST, ELOVL7, FLRT2, IFI202B, IL13RA1, ITGB3, LCP1, MYLK4, PPFIBP2, PPP1R3B, PRDM1, SCARA5, SGMS2, SHE, STAT6, TBX22, TEK, TFCP2L1, UNC45B |
| Development | ACER2, CGNL1, CRIM1, DMRTA2, EPB41L1, FBN2, FNDC3B, FOXC1, FZD4, GBP7, JADE2, JAG1, LPAR1, MATN2, MEF2C, MSRB3, MYOT, PAPSS2, PEG3, PGM5, RBSM1, SLC16A12, SMOC2, SSPN, STARD8, TRP63, VAMP5 |
| Molecular Transport | ABCA9, CD248, FAM174B, FAM84A, FNDC1, ITPRIPL2, LRFN2, PIEZO2, PRTG |
| Metabolism | AFAP1L2, ARHGAP36, CXCL12, EEF2K, FRK, HMCN1, NOX4, PTGFRN |
| Vision | CGNL1, CRIM1, DMRTA2, EPB41L1, FBN2, FNDC3B, FZD4, KIAA1522, MATN2, MSRB3, PAPSS2, PGM5, RBSM1, SLC16A12, STARD8, VAMP5 |
| PN Network Function | Genes |
| Cardiovascular | ARG1, BACH2, BMP7, BOC, CAV3, CD44, COL25A1, DLK1, ELK3, ETS1, FLT1, FOXC1, HAPLN1, HAS2, JAG1, LAMC1, LCP1, LPP, NOX4, NRP2, NTRK2, OSTF1, RSPO1, SEMA3C, SEMA3D, SLC7A8, SULF1, TEK, TGFBR3, THBS2, VCAN, WNT4 |
| Structure | ABCA1, ABCC9, B3GNT9, CASP12, CCBE1, CCND2, CD55, CD93, EHF, ELL2, FAM46A, IL13RA1, OSR1, PPARGC1A, PRDM1, STAT6, UCP2 |
| Hematological Disease | FAM101B, FAM114A1, FAM174B, FAM84A, FZD6, KDELC2, MSRB3, PAPSS2, PDLIM7, PIEZO2, PRICKLE1, SGMS2, TMEM164, VAMP5 |
| Cellular Function | ACPP, DPT, EEF2K, EGFL6, EHD4, ENTPD1, FOSL2, FRAS1, LMO7, LYZ1, MYRF, OTX1, RGS5, SDPR, TNNT2 |
| Immune Response/Injury | ASPN, BNC2, CPM, DENND2C, FBN2, FIBIN, FNDC1, ITPRIPL2, NEBL, PPP1R3B, STARD8, TBX18 |
| Metabolism | CRHBP, DEK, DMRTA2, FBXL7, GATM, NRK, RBSM1, SGMS1, SMPX, TFCP2L1, DST, ELOVL7, MYLK4, PGM5, TBX22, UNC45B, SLC7A10 |
| Development | ACER2, DSG2, EGFR, FZD4, LMNA, MEF2C, PEG3, SSPN, TRP63 |
Genes regulated by NR2E3 are in bold; genes regulated by RORA are in italics, and genes regulated by NR2E3 and RORA are bold and italics.
Table 3.
Ingenuity pathway analysis (IPA) of miRNA differentially expressed between rd7 and B6.
| miRNA | Disease Annotation | |
|---|---|---|
| Cancer | miR-185-5p, miR-199a-5p | Early invasive cervical squamous cell carcinoma |
| miR-542-3p | Cell proliferation of bone cancer cell lines | |
| miR-542-3p | Cell proliferation of ovarian cancer cell lines | |
| miR-542-3p | Cell proliferation of brain cancer cell lines | |
| miR-125b-5p | Colony formation of bladder cancer cell lines | |
| miR-125b-5p, miR-185-5p, miR-199a-5p miR-331-3p, miR-542-3p | Proliferation of tumor cell lines | |
| miR-125b-5p | Advanced Dukes’ stage colorectal cancer | |
| miR-185-5p, miR-199a-5p | Proliferation of cervical cancer cell lines | |
| miR-125b-5p, miR-185-5p, miR-199a-5p | Squamous-cell carcinoma | |
| miR-125b-5p | Apoptosis of neuroblastoma cells | |
| miR-125b-5p | Metastatic HER2 negative hormone receptor | |
| miR-125b-5p, miR-339-5p, miR-542-3p | Hematologic cancer | |
| miR-125b-5p | Apoptosis of mammary tumor cells | |
| miR-125b-5p, miR-199a-5p, miR-329-3p miR-331-3p, miR-423-5p, miR-495-3p, miR-542-3p, miR-574-5p, miR-654-3p | Colorectal cancer | |
| miR-185-5p | Arrest in G1 phase of colon cancer cell lines | |
| miR-185-5p, miR-199a-5p | Renal clear cell adenocarcinoma | |
| miR-339-5p | Precursor B-cell acute lymphoblastic leukemia | |
| miR-125b-5p, miR-331-3p | Proliferation of prostate cancer cell lines | |
| miR-125b-5p | Differentiation of embryonic stem cell lines | |
| miR-542-3p | Cell proliferation of adenocarcinoma cell lines | |
| miR-125b-5p, miR-339-5p | Lymphoproliferative malignancy | |
| miR-199a-5p | Response of endothelial cell lines | |
| miR-125b-5p | Migration of microvascular endothelial cells | |
| Diabetes | miR-125b-5p, miR-327, miR-434-3p | Non-insulin-dependent diabetes mellitus |
| miR-125b-5p, miR-185-5p, miR-327 miR-329-3p, miR-434-3p | Diabetes mellitus | |
| Cardiovascular | miR-199a-5p, miR-423-5p | Failure of heart |
| miR-199a-5p | Interstitial fibrosis of heart | |
| miR-125b-5p*, miR-199a-5p | Fibrosis of heart | |
| miR-199a-5p | Vasoconstriction of aorta | |
| miR-199a-5p | Enlargement of heart | |
| Inflammation | miR-125b-5p, miR-185-5p, miR-188-3p miR-199a-5p, miR-423-5p | Inflammation of body region |
| miR-188-3p | Inflammation of hippocampus | |
| miR-125b-5p, miR-185-5p, miR-199a-5p miR-423-5p | Inflammation of body cavity | |
| miR-125b-5p, miR-185-5p, miR-188-3p miR-199a-5p, miR-423-5p | Inflammation of organ | |
| miR-188-3p | Neuroinflammation | |
| miR-125b-5p | Differentiation of granulocyte progenitors | |
| miR-125b-5p | Transendothelial migration of monocytes | |
| miR-125b-5p | Activation GM-CSF dependent marrow macrophage | |
| miR-125b-5p | M1 polarization of M2 macrophages | |
| Disease | miR-125b-5p, miR-423-5p | Rapidly progressive idiopathic pulmonary fibrosis |
| miR-125b-5p | Myelodysplastic syndrome with 5q- syndrome | |
| miR-125b-5p | Formation of tubules | |
| miR-199a-5p | Chronic hepatitis B | |
| miR-125b-5p, miR-199a-5p, miR-423-5p | Fibrosis | |
| miR-199a-5p, miR-339-5p, miR-495-3p | Nonobstructive azoospermia | |
| miR-329-3p | Dent’s disease | |
| miR-125b-5p | Systemic sclerosis | |
| miR-125b-5p, miR-185-5p | Chromosomal aberration | |
| miR-125b-5p | Androgenic alopecia | |
| miR-185-5p | DiGeorge syndrome |
Five major groups with a P value p < 0.05 were identified with the majority of the miRNAs associated with cancer.
Figure 3.
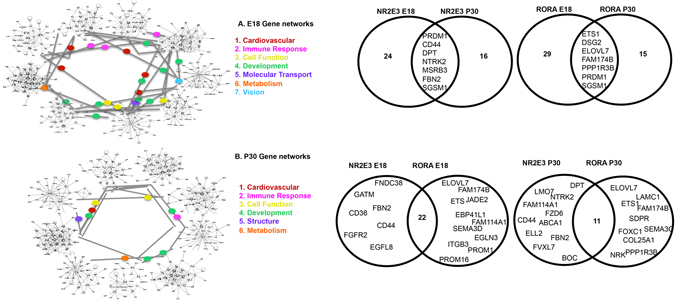
Interacting NR2E3 and RORA Associated Gene Networks. (A) IPA analysis of E18 targets identified 9 gene networks with 7 biological classifications. (B) IPA analysis of P30 targets identified 9 gene networks with 6 biological classifications. Venn Diagrams show unique and overlapping gene targets of NR2E3 and RORA at E18 and P30. Comparisons of RORA E18/P30 or NR2E3 E18/P30 show less overlap than RORA/NR2E3 at E18 or RORA/NR2E3 at P30.
Nr2e3 and Rora co-regulate AMD pathways with miRNAS
Previous work from our lab and others has demonstrated that the nuclear hormone receptors Nr2e3 and Rora are key regulators of retinal development and function41–45. To determine which miRNA potential target genes and which miRNAs are directly regulated by NR2E3 or RORA, we scanned approximately 100 kilobases (kb) upstream and into intron 1 of each target gene for putative response element (RE) binding sites. We identified 87 genes and 10 miRNAs that had putative Nr2e3 and/or RORA RE binding sites. Chromatin immunoprecipitation (ChIP) revealed 68 direct target genes of NR2E3 or RORA in the developing (E18) and/or mature retina (P30) (Figs 4–6) as well as 7 miRNAs (Fig. 7). At E18.5 there were 36 target genes identified for RORA (Fig. 4) and 32 for NR2E3 (Fig. 5) with 23 of these genes common targets for both RORA and NR2E3. At P30, we identified 34 RORA targets and 27 NR2E3 target genes, 12 of which were common targets of both (Fig. 6). In addition, all potential target genes were cross-referenced against an RNA-Seq expression analysis of human macular RPE/choroid and macular retina from an AMD cohort of donor eyes. This cross-reference revealed 145 common genes. In particular, three genes (vascular endothelial growth factor receptor VEGFR1 (FLT1), protogenin (PRTG), and the synaptic adhesion molecule (LRFN2)) were significantly differentially expressed between normal RPE and Neo AMD RPE. PRTG, LRFN2, and the leucine zipper transcription factor, BACH2 were significantly differentially expressed between neovascular AMD RPE and intermediate (AREDS3) RPE.
Figure 4.
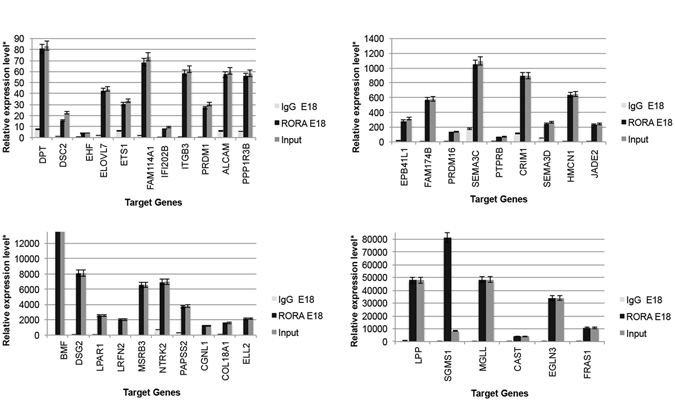
Chromatin IP (chIP) quantitative real time PCR (chIP-qRT-PCR) of gene targets for RORA at E18. Input (positive control), RORA-chIP, and immunoglobulin G (IgG) negative control. *Relative expression is calculated as molecules/1000 molecules β-actin. Standard errors are indicated for each gene. All genes had a minimum significance of p < 0.05.
Figure 6.

Chromatin IP (chIP) quantitative real time PCR (chIP-qRT-PCR) of gene targets for RORA and NR2E3 at P30. Input (positive control), RORA-chIP, NR2E3-chIP, and immunoglobulin G (IgG) negative control. *Relative expression is calculated as molecules/1000 molecules β-actin. Standard errors are indicated for each gene. All genes had a minimum significance of p < 0.05.
Figure 7.
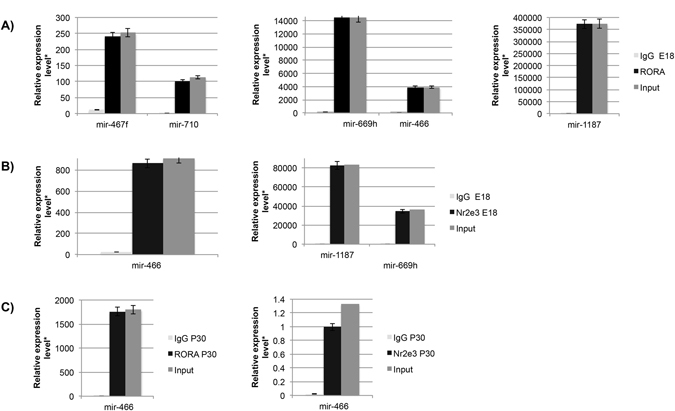
Chromatin IP (chIP) quantitative real time PCR (chIP-qRT-PCR) of miRNA targets for RORA and NR2E3 at E18 and P30. (A) RORA targeted miRNAs identified at E18 (B) NR2E3 targeted miRNAs identified at E18, (C) RORA and NR2E3 targeted miRNAs identified at E18. Input (positive control), RORA-chIP, NR2E3-chIP, and immunoglobulin G (IgG) negative control. *Relative expression is calculated as molecules/1000 molecules β-actin. Standard errors are indicated for each gene. All genes had a minimum significance of p < 0.05.
Figure 5.

Chromatin IP (chIP) quantitative real time PCR (chIP-qRT-PCR) of gene targets for NR2E3 at E18. Input (positive control), NR2E3-chIP, and immunoglobulin G (IgG) negative control. *Relative expression is calculated as molecules/1000 molecules β-actin. Standard errors are indicated for each gene. All genes had a minimum significance of p < 0.05.
Multimodal regulation of AMD gene networks by NRs, miRNAs, and Epigenetic factors
In addition to NHRs and miRNAs, a key component of gene regulation is epigenetics, and Nr2e3 has been implicated in the Ezh2 network59. In order to determine the epigenetic influence on miRNA and NHR target genes and gene networks potentially regulated by miRNAs and NHRs and associated with AMD. we compared the miRNA target genes against microarray data from P0 Ezh2 −/− retinas as E18 and P0 time points have similar epigenetic effects (personal communication Dr. Chen). Although our studies were performed at E18, mouse gestation is 18–20 days and our prior unpublished work concurred that evaluating gene expression at E18 is quite similar to P0. In this cross-reference, we identified 7 genes and their 13 putative miRNAs as being significantly differentially expressed in Ezh2 −/− compared to normal and also potential target genes of the miRNA identified in this study (Table 4). Furthermore, two of these genes (Ell2 and Entpd1) are commonly modulated by the NHRs Nr2e3 and Rora and are also differentially expressed in the RPE versus retina of AMD cohort.
Table 4.
miRNAs and their potential target genes that are differentially expressed in rd7 vs B6 and Ezh2 vs B6.
| Gene | miRNA | Expression TP |
|---|---|---|
| Pgm5 | mmu-miR-466a-5p | E18, P6, P14, P30 |
| mmu-miR-466d-5p | E18, P6, P14, P30 | |
| mmu-miR-466e-5p | E18, P6, P14, P30 | |
| mmu-miR-1187 | E18, P6, P14 | |
| Ell2 | mmu-miR-1192 | E18, P6, P14, P30 |
| mmu-miR-188-3p | E18 | |
| mmu-miR-743b-3p | E18 | |
| Aspn | mmu-miR-369-3p | E18, P6, P14 |
| Jag1 | mmu-miR-466f-3p | E18, P6, P14, P30 |
| mmu-miR-467f | E18, P6, P14, P30 | |
| mmu-miR-467g | E18, P6, P14, P30 | |
| mmu-miR-466d-3p | E18, P6, P14 | |
| mmu-miR-710 | E18, P6, P14 | |
| Slc16a12 | mmu-miR-188-3p | E18 |
| Crhbp | mmu-miR-466a-5p | E18, P6, P14, P30 |
| mmu-miR-466e-5p | E18, P6, P14, P30 | |
| mmu-miR-1187 | E18, P6, P14 | |
| Entpd1 | mmu-miR-466a-5p | E18, P6, P14, P30 |
| mmu-miR-466e-5p | E18, P6, P14, P30 | |
| mmu-miR-327 | E18 | |
| mmu-miR-1187 | E18, P6, P14 |
Genes and miRNAs regulated by NR2E3 are in bold; genes and miRNAs regulated by RORA are in italics, and genes and miRNAs regulated by NR2E3 and RORA are bold and italics. Minimum 2 fold change and p < 0.05.
Discussion
This study is the first to evaluate multimodal regulation of gene expression by evaluating the combined impact of miRNAs, NHRs, and epigenetic factors. Recently, miRNAs have emerged as an important class of regulatory molecules that affect gene expression and in turn normal human development, function, and disease15, 16. miRNAs are endogenous non-coding RNA that regulate gene expression. Approximately 21,264 miRNAs have been identified to date and are classified in families based on sequence or the pre-miRNA structure41, 48–92. Over 400 miRNAs have been identified in the retina and function in the similar, parallel biological pathways in different tissues. This study reveals the synergistic impact of miRNAs, NHRs, and epigenetic factors on common biological networks in the normal and diseased state of the retina.
Several studies show that miRNA expression is subject to precise regulation that varies dramatically by cell type, developmental stage, environmental conditions, and disease pathology14. Currently, over 400 hundred miRNAs have been identified as key modulators of gene expression in the retina14, 17, 93. Here, we utilize a comprehensive strategy to identify and characterize multimodal nodes of gene regulation important in human retinal development and disease. We identified miRNAs that, along with the NHRs Nr2e3 and Rora and the epigenetic factor Ezh2, play a synergistic role in modulating key biological networks affected in the complex human disease AMD. Eighteen overlapping gene networks were identified at E18 and P30 (Fig. 3A,B). These networks contained genes that regulate metabolism, immune response, development, and other cellular functions. We identified novel genes that are co-regulated by miRNAs, the nuclear hormone receptors Nr2e3 and/or Rora. When comparing gene targets, NR2E3 and RORA have more targets in common at each time point (E18, 22 genes or P30, 11 genes) than for the same NHR at different time points (Fig. 3A,B). Furthermore, our study adds novel components to the list of Nr2e3 and Rora targets identified thus far42, 94. We identified 4 miRNAs that are differentially expressed in the retina of affected Nr2e3 rd7/rd7 mice. These findings support the hypothesis that genetic variations strongly influence phenotypic outcome by perturbing gene regulation. We discovered 25 unique miRNAs with novel function in the retina. miR-466 and miR-1187, in particular, emerge as significant and regulators, sharing common target genes with Nr2e3, Rora, Ezh2 that are also differentially expressed in AMD.
This study revealed several miRNAs and genes functioning in the developmental gene network. miR-466 was discovered in a recent study to identify miRNAs that target the Prospero homeobox 1 (Prox1) gene, a known regulator of retinal neurogenesis. More recent studies show Prox1 influences inflammation-induced lymph-angiogenesis by regulating the expression of VEGF-C receptor. miR-466 in turn impacts lymph-angiogenesis by targeting Prox1 3′UTR sequence and suppressing the expression of the gene and therefore reducing lymphangiogenesis95. miR-1187 has no known function in the retina. Current studies show miR-1187 is down-regulated in a mouse model for acute liver failure and plays a role in hepatocyte apoptosis96. miR-1187 potential targets include Carboxypeptidase M (Cpm) a protein coding gene involved in monocyte and macrophage differentiation97, Ectonucleoside triphosphate diphosphohydrolase 1 (Entpd1) which functions in the hydrolyzes of phosphates into ATP, ADP, and AMP98, and Fraser extracellular matrix complex subunit 1(Fras1) that encodes for an extracellular matrix protein that regulates the epidermal-basement membrane adhesion as well as organogenesis in development99. Alcam codes for a transmembrane glycoprotein that belongs to the immunoglobulin family. Alcam has been implicated in the control of numerous developmental and pathological processes such as activation of T cells, melanoma cells, activation of monocytes, and has lately been used as a cancer stem cell marker100, 101. Alcam mutant retinas exhibit evaginated or invaginated regions with photoreceptor rosettes that resembled the retinal folds observed in some human retinopathies and also observed in the Nr2e3 rd7/rd7 retina100.
We identified seven putative miRNA target genes whose expression are also regulated by nuclear hormone receptors Nr2e3, Rora, and Ezh2: Phosphoglucomutase 5(Pgm5), Elongation factor for RNA polymerase II 2(Ell2), Asporin (Aspn), Jagged 1 (Jag1), and solute carrier family 16 member 12(Slc16a12), Corticotropin releasing hormone binding protein(Crhbp), Ectonucleoside Triphosphate Diphosphohydrolase (Entpd1). Jag1 is the ubiquitously expressed, well known ligand of Notch and is part of a critical signaling pathway for the development of most tissues102–104. Our results add Nr2e3 and Ezh2 to the recently identified regulatory interplay between Jag1 and 17beta-estradiol that is important in cancer and other tissue specific disorders including retinal diseases105. Ell2, codes for an RNA polymerase elongation factor and helps regulate the activity of the elongation complex. Ell2 increases the rate of elongation during promoter dependent and independent transcription106. Pgm5 is a phosphotransferase involved in the conversion of glucose107. In contrast the Aspn gene is involved in collagen mineralization and helps regulate chondrogenesis108. Slc16a12 codes for a transmembrane transporter and plays a role in monocarboxylic acid transport. Mutations in Slc16a12 are associated with juvenile cataracts109. Lastly, Crhbp is a corticotropin hormone and plays a role in development of the optic cup110. Our study connects several known, important genes and biological processes with novel miRNAs in normal and diseased retina.
Age-related macular degeneration is a leading cause of blindness in the world. In this study we discovered three key known AMD genes, Fms related tyrosine kinase 1(Flt1), the receptor for vascular endothelial growth factor (VEGF) (putative miR1192 target), ATP binding cassette subfamily A member 1 (Abca1) (putative miR673-5p target), and Activated leukocyte cell adhesion molecule(Alcam) (putative miR188-3p target) are regulated by Nr2e3 and Rora and their potential corresponding miRNAS that are differentially expressed in Nr2e3 rd7/rd7 retinas. Flt1, a receptor tyrosine kinase, functions in the VEGF pathway, is predominantly expressed in endothelial cells, and is the target for transducing signal in VEGF treatment111. Moreover, our group previously determined Flt1 is a target of Nr2e3, and demonstrated genetic variants in Flt1 in four ethnically diverse patient populations associated with AMD112. Abca1 is an ATP transporter that is expressed in the retina as well as in the RPE113. Alleles of the Abca1 gene are associated with a reduction in the levels of HDL contributing to the formation of drusen accumulation in AMD114. Variants in Abca1 have also been shown to be associated with risk of AMD115. In addition, the miRNAs mir466 and mir1186 have 5 potential target genes that are differentially expressed in AMD patients compared to unaffected individuals: lymphocyte cytosolic protein1 (Lcp1), NADPH oxidase 4 (Nox 4) regulator of G-protein signaling (Rgs5), Dystonin (Dst), and Cpm.
Recent studies show an important role for the Nr2e3 in cancer116–118. Interestingly, in this study we identified several miRNAs that are differentially expressed in rd7 and play a significant role in cancer. The rd7 model does not develop cancer, however it is well documented that rd7 mice have a defect in cell proliferation41, 119. Consistent with reports that miRNAs are predominantly downregulated in cancer tumors120, our studies show that 85% of the miRNAs that are differentially expressed in rd7 are downregulated. Many of these miRNAs are also subject to epigenetic regulation and are located within CpG islands and regulated by methylation121. Interestingly, our IPA analysis also revealed the miRNA miR125b-5p is associated with diabetes and cancer (Table 3). A study in breast cancer lines demonstrated a direct link between the methylation status of the cancer miRNA promoters and the development of cancer122. These findings further substantiate an important role for nuclear receptors such as Nr2e3 in broader-impact diseases such as cancer. Our future studies will further explore the role of Nr2e3 in cancer and regulation of metastasis via miRNAs and NHRs.
While numerous studies show the significant singular impact that NHRs, miRNAs, and epigenetics have on mammalian development and human disease, in vivo, the process is dynamic and complex involving multiple factors simultaneously converging for singular biological events. This study is the first to demonstrate the synergistic and combined impact of multiple factors on gene expression in the developing and mature retina and its impact on human disease. While these studies were conducted in the retina, the genes and microRNAs identified here are not unique to the retina or even the CNS. The findings of this study will provide a better understanding of normal development, genomic signatures modulating complex human retinal disease, and valuable tools for better prognostics and development of improved therapies. Future studies will explore developing combination therapies to combat human retinal diseases such as AMD.
Materials and Methods
Ethics Statement
All animals were bred and maintained under standard conditions at The University of Nebraska Medical Center or Schepens Eye Research Institute research vivarium in accordance with protocol numbers 04086, approved by the Animal Care and Use Committee at the University of Nebraska Medical Center, and 3980417 and 4060617 at Schepens Eye Research Institute. Mice were housed in microisolator cages and provided food and water ad libitum. The University of Nebraska Medical Center is in compliance with the NIH policy on the use of animals in research (Animal Welfare Act P.L. 89–544, as amended by P.L. 91–579 and P.L. 94–279) as well as the Guide for the Care and Use of Laboratory Animals, NIH Publication No. 86–23. All experiments were performed in accordance with relevant guidelines and regulations.
Availability of Data and Materials
The miRNA dataset supporting the conclusion of this article has been made MIAME compliant and is publicly available through the Gene Expression Omnibus (GEO) database under the series record GSE91029, and the prior microarray used to cross reference miRNA targets are available under the series record GSE24512. AMD data used as cross reference in this study is in preparation for publication.
Microarray Analysis
MicroRNA microarrays were performed using mirVana 2-color microRNA array chips hybridized to 100 nanogram (ng) of total miRNA from 10 pool retinas for each time point from Nr2e3 rd7/rd7 and C57BL6/J (B6) control retinas extracted at E18, P6, P14, or P30. Samples were hybridized to mirVana miRNA chips followed by normalization using Lowess normalization and generation of scatter plots using GenePix to determine the strong data points. Chips were normalized using both whole chip normalization and control spot normalization. Heat map of raw data is found in Supplemental Figure 1. Differential miRNA expression was determined utilizing a merged data set of both normalization techniques and the following criteria: (1) >20% spots showing >2-fold change; (2) P < 0.001 for log-ratio variation; (3) exclusion of array spots with >50% missing values. Additional analysis of differentially expressed miRNAs was performed using BRB Array Tools for Excel, as well as the Stanford Statistical Analysis of Microarrays (SAM) plug-in for Excel. miRNA expression data were input into SAM using a false discovery rate (FDR) of <0.05. Heat map for the microarray analysis with a hierarchical cluster analysis has been previously published91. Heat map depicting differential expression of miRNA as observed by miRNA array. Values represent median intensity values for the respective label (cy5/cy3). Color-coding of heat map is relative to percentile higher expression values represented in orange and lower expression values in dark indigo (Supplemental Figure 1). All differentially expressed miRNAs were subsequently evaluated against the mirBase to determine all potential target genes for each miRNA. Each miRNA can have up to 700–1500 potential target genes.
A cross-comparison was made between the miRNA array data (differentially expressed miRNAs) with microarray data (gene expression >2 fold change). RNA was isolated at E18.5, P6, P14, and P30 from Nr2e3 rd7/rd7 retinas as previously described42. Eyes were enucleated from 7 different animals (at each time point) and placed in phosphate buffered saline (PBS) on ice. Retinas were dissected using a stereomicroscope (Zeiss SV11) and RNA was isolated by TRIzol® extraction. RNA was hybridized to Mouse 420A 2.0 (Affymetrix, Santa Clara, CA) chips by the UNMC Microarray Core Facility according to manufacturer specifications (Affymetrix, Santa Clara, CA). Data quality was assessed using the affyPLM package for the R programming language. Consistency of expression levels was confirmed by validation across multiple redundant probe sets. Differential expression analysis was performed using the Linear Models for Microarray Analysis portion of Bioconductor. Genes found to be differentially expressed for each pair wise comparison using an FDR-adjusted p-value of 0.001 and at least a 2-fold change were combined and used to perform clustering analysis. A self-organizing map (SOM) clustering algorithm was applied to genes showing significant expression differences as judged by mean log2 intensity per strain. The gap statistic was used to estimate the optimal number of clusters.
Quantitative RT-PCR
Each gene and miRNA identified through microarray studies were validated through quantitative real-time PCR (qRT-PCR). Real time PCR was performed as previously described42 to confirm differential expression observed in the microarray data. RNA was isolated using the samples from the microarray analysis as well as additional individual samples from mice for each strain (total n = 7 per sample type). Two µg of total RNA was reverse transcribed using Retroscript (Ambion). The cDNA samples were diluted 1:100 and real time PCR was performed in triplicates for each primer using Sybr Green PCR master mix (Thermo fisher). Reactions were quantified using an ABI Step One Plus Real Time PCR and analyzed with the corresponding software. Relative expression levels were determined by normalizing cycle thresholds values to the amount of β-actin expressed (1000/2^Ct gene − Ct β-actin) Statistical significance of differential expression was assessed using a T-test and p < 0.05.
Pathway Generation and Analysis
miRNA and microarray target genes were analyzed using Ingenuity Pathway Analysis (IPA, Ingenuity Systems, www.ingenuity.com). Gene identifiers and statistically significant expression values were uploaded into Ingenuity. Default cutoffs were set to identify genes whose expression was significantly differentially regulated and overlaid onto a global molecular network developed from information contained in the Ingenuity Pathways Knowledge Base. Networks were algorithmically generated based on their connectivity. Genes or gene products in the networks are represented as nodes, and the biological relationship between two nodes is represented as an edge (line). All edges are supported by at least 1 reference from the literature, from a textbook, or from canonical information stored in the Ingenuity Pathways Knowledge Base. Nodes are displayed using various shapes that represent the functional class of the gene product.
Chromatin Immunoprecipitation
Putative miRNA target genes were analyzed for nuclear receptor binding sites using: 1) classic binding sites of Nr2e3 (AAGTCA(n1-4)AAGTCA) or Rora ([AGT][TA][AT][TA]C[AT]AGGTCA); 2) general nuclear hormone receptor response elements as determined algorithmically by NUBIscan123. Binding sites were selected at a maximum of 100 kilobases (kb) upstream of each gene’s start site to intron 1. Chromatin immunoprecipitation was performed as described previously42. Briefly, retinas were dissected from P30 C57Bl/6J mice. A total of 8–10 retinas were used per Chip reaction and placed into 400 uL of a PI solution (phosphate buffered saline (PBS) and protease inhibitor cocktail from Roche). Chilled tissue was dissociated and homogenized and cross-linked in 37% formaldehyde for a period of 60 minutes at room temperature in a rotating platform. 1 M glycine was added (final concentration 0.125 mM) and later rinsed with the phosphatase inhibitor (PI) solution in PBS (Thermo, cat#78430). Sonication was performed using a Qsonica Ultrasonic Processor amplitude 50%, 10 pulses x 30 with 1-second pauses between pulses. Immunoprecipitation was performed overnight using 1 μg of NR2E3 antibody at 4 °C on a rotating platform. The NR2E3 antibody used in this experiment had been previously generated in the laboratory43 and validated42. For a negative control goat IgG antibody was used and the input (control) was not incubated with antibody. Immunoprecipitated samples were resuspended with Protein G dynabeads (Invitrogen, cat#10003D) and a series of washes using wash buffer #1 (Tris, EDTA 0.5 M, Triton, NaCl 5 M) and wash buffer #2 (Tris, EDTA 0.5 M, NP-40, LiCl) were performed at room temperature. Samples were reversed cross-inked with NaCl (200 mM final concentration). Quantitative RT-PCR was performed using 1 ul of 1:100 dilution (input) and 1:10 dilution (samples and immunoglobulin G (IgG) control) using conditions described previously36. All sample data was normalized to IgG control.
Comparison to Human RNA-Seq Data
Resulting mouse data was compared to differentially expressed genes found in the Utah human donor eye AMD repository and RNA-Seq dataset (personal communication DeAngelis lab) from macular neural retina and macular RPE/choroid. Cohort is comprised of Caucasians over the age of 60 including representatives of both intermediate and neovascular subtypes with age matched controls.
Electronic supplementary material
Acknowledgements
This work was supported by the following funding agencies: NIH-NCRR P20-RRO18788-03 (NBH), NIH-NEI RO1EY017653 (NBH), NIH-NEI RO1EY017653-01A2S1 (NBH), an unrestricted grant from Research to Prevent Blindness, Inc., New York, NY to Department of Ophthalmology, Harvard Medical School (NBH), Hope for Vision (NBH), Massachusetts Lions Eye Research Fund (NBH), American Macular Degeneration Foundation (NBH), Webster Foundation (NBH), Edward N. & Della L. Thome Memorial Foundation (NBH, MMD), NIH/NEI EY014800 (MMD), an unrestricted grant from Research to Prevent Blindness, Inc., New York, NY to the Department of Ophthalmology & Visual Sciences, University of Utah (MMD), The Skaggs Foundation for Research (MMD), The Carl Marshall Reeves & Mildred Almen Reeves Foundation, Inc., (MMD), and the Macular Degeneration Foundation, Inc. (MMD), NIH/NEI R41 EY025913 (DFC), and NIH/NEI R01EY025259 (DFC). We wish to thank Gloria Chen for assistance with R and generating the heat map.
Author Contributions
Conceived and design the study: A.S.J., D.F.C., M.M.D., N.B.H. Conducted the experiments: A.M.O., A.S.J., B.L., A.H., M.A.M., L.C. Analyzed the data: A.M.O., A.S.J., L.C., D.F.C., M.M.D., N.B.H. Drafted the manuscript: A.M.O., N.B.H. All authors reviewed the manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Electronic supplementary material
Supplementary information accompanies this paper at doi:10.1038/s41598-017-00788-3
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Waterland RA, Jirtle RL. Transposable elements: targets for early nutritional effects on epigenetic gene regulation. Mol. Cell. Biol. 2003;23:5293–300. doi: 10.1128/MCB.23.15.5293-5300.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Filipowicz W, Bhattacharyya S, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat. Rev. Genet. 2008;9:102–114. doi: 10.1038/nrg2290. [DOI] [PubMed] [Google Scholar]
- 3.Vasudevan S, Tong Y, Steitz JA. Switching from Repression to Activation: MicroRNAs Can Up-Regulate Translation. Science (80-.). 2007;318:1931–1934. doi: 10.1126/science.1149460. [DOI] [PubMed] [Google Scholar]
- 4.Morgan DJ, Deangelis MM. Differential Gene Expression in Age-Related Macular Degeneration. Cold Spring Harb. Perspect. Med. 2014;5:a017210. doi: 10.1101/cshperspect.a017210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Olivares AM, Moreno-Ramos OA, Haider NB. Role of Nuclear Receptors in Central Nervous System Development and Associated Diseases. Journal of experimental neuroscience. 2015;9:93–121. doi: 10.4137/JEN.S25480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hu W, Coller J. What comes first: translational repression or mRNA degradation? The deepening mystery of microRNA function. Cell Res. 2012;22:1322–1324. doi: 10.1038/cr.2012.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Nogueira T, Springer M. Post-transcriptional control by global regulators of gene expression in bacteria. Curr Opin Microbiol. 2000;3:154–8. doi: 10.1016/S1369-5274(00)00068-0. [DOI] [PubMed] [Google Scholar]
- 8.Chen M, Manley J. Mechanisms of alternative splicing regulation: Insights from molecular and genomics approaches. Nat. Rev. Mol. Cell Biol. 2009;10:741–54. doi: 10.1038/nrm2777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kumar M, Carmichael G. Antisense RNA: Function and Fate of Duplex RNA in Cells of Higher Eukaryotes. Microbiol. Mol. Biol. Rev. 1998;62:1415–1434. doi: 10.1128/mmbr.62.4.1415-1434.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Aroni N. Target selectivity in mRNA silencing. Gene Ther. 2006;13:509–516. doi: 10.1038/sj.gt.3302726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fabian M, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem. 2010;79:351–79. doi: 10.1146/annurev-biochem-060308-103103. [DOI] [PubMed] [Google Scholar]
- 12.Xu S, Witmer P, Lumayag S, Kovacs B, Valle D. MicroRNA (miRNA) transcriptome of mouse retina and identification of a sensory organ-specific miRNA cluster. J Biol Chem. 2007;282:25053–25066. doi: 10.1074/jbc.M700501200. [DOI] [PubMed] [Google Scholar]
- 13.Jin Z, Hirokawa G, Gui L. Targeted deletion of miR-182, an abundant retinal microRNA. Mol Vis. 2009;15:523–533. [PMC free article] [PubMed] [Google Scholar]
- 14.Sundermeier TR, Palczewski K. The physiological impact of microRNA gene regulation in the retina. Cell Mol Life Sci. 2012;69:2739–2750. doi: 10.1007/s00018-012-0976-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bartel DP. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell. 2004;116:281–297. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 16.Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- 17.Soundara Pandi SP, Chen M, Guduric-Fuchs J, Xu H, Simpson DA. Extremely Complex Populations of Small RNAs in the Mouse Retina and RPE/ChoroidmicroRNAs in the Retina and RPE/Choroid. Invest. Ophthalmol. Vis. Sci. 2013;54:8140–8151. doi: 10.1167/iovs.13-12631. [DOI] [PubMed] [Google Scholar]
- 18.Hackler L, Wan J, Swaroop A, Qian J, Zack DJ. MicroRNA Profile of the Developing Mouse Retina. Invest. Ophthalmol. Vis. Sci. 2010;51:1823–1831. doi: 10.1167/iovs.09-4657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Baskerville S, Bartel DP. Microarray profiling of microRNAs reveals frequent coexpression with neighboring miRNAs and host genes. RNA. 2005;11:241–7. doi: 10.1261/rna.7240905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Organisciak DT, Vaughan DK. Retinal Light Damage: Mechanisms and Protection. Prog Retin Eye Res. 2010;29:113–134. doi: 10.1016/j.preteyeres.2009.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sun H, Nathans J. The Challenge of macular degeneration. Sci Am. 2001;285:68–75. doi: 10.1038/scientificamerican1001-68. [DOI] [PubMed] [Google Scholar]
- 22.Sanuki R, et al. miR-124a is required for hippocampal axogenesis and retinal cone survival through Lhx2 suppression. Nat Neurosci. 2011;14:1125–34. doi: 10.1038/nn.2897. [DOI] [PubMed] [Google Scholar]
- 23.Walker J, Harland R. microRNA-24a is required to repress apoptosis in the developing neural retina. Genes Dev. 2009;23:1046–1051. doi: 10.1101/gad.1777709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.La Torre A, Georgi S, Reh Ta. Conserved microRNA pathway regulates developmental timing of retinal neurogenesis. Proc. Natl. Acad. Sci. USA. 2013;110:E2362–70. doi: 10.1073/pnas.1301837110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Georgi SA, Reh TA. Dicer is required for the maintenance of notch signaling and gliogenic competence during mouse retinal development. Dev. Neurobiol. 2011;71:1153–1169. doi: 10.1002/dneu.20899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kovacs B, Lumayag S, Cowan C, Xu S. MicroRNAs in early diabetic retinopathy in streptozotocin-induced diabetic rat. Investig. Ophthalmol. Vis. Sci. 2011;52:4402–9. doi: 10.1167/iovs.10-6879. [DOI] [PubMed] [Google Scholar]
- 27.Ishida W, et al. Dynamic changes of microRNAs in the eye during the development of experimental autoimmune uveoretinitis. Invest. Ophthalmol. Vis. Sci. 2011;52:611–7. doi: 10.1167/iovs.10-6115. [DOI] [PubMed] [Google Scholar]
- 28.McArthur K, Feng B, Wu Y, Chen S, Chakrabarti S. MicroRNA-200b regulates vascular endothelial growth factor-mediated alterations in diabetic retinopathy. Diabetes. 2011;60:1314–1323. doi: 10.2337/db10-1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Loscher CJ, et al. Altered retinal microRNA expression profile in a mouse model of retinitis pigmentosa. Genome Biol. 2007;8:R248. doi: 10.1186/gb-2007-8-11-r248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Yu X, Lin J, Zack DJ, Mendell JT, Qian J. Analysis of regulatory network topology reveals functionally distinct classes of microRNAs. Nucleic Acids Res. 2008;36:6494–6503. doi: 10.1093/nar/gkn712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tsang J, Zhu J, van Oudenaarden A. MicroRNA-mediated feedback and feedforward loops are Recurrent Network Motifs in Mammals. Mol Cell. 2007;26:753–767. doi: 10.1016/j.molcel.2007.05.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Shalgi R, Lieber D, Oren M, Pilpel Y. Global and local architecture of the mammalian microRNA-transcription factor regulatory network. PLoS Comput. Biol. 2007;3:1291–1304. doi: 10.1371/journal.pcbi.0030131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jetten AM. Retinoid-related orphan receptors (RORs): critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl. Recept. Signal. 2009;7:1–32. doi: 10.1621/nrs.07003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Roberts M, Srinivas M, Forrest D, Morreale de Escobar G, Reh T. Making the gradient: thyroid hormone regulates cone opsin expression in the developing mouse retina. Proc. Natl. Acad. Sci. USA. 2006;103:6218–6223. doi: 10.1073/pnas.0509981103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chow L, Levine EM, Reh TA. The nuclear receptor transcription factor, retinoid-related orphan receptor β, regulates retinal progenitor proliferation. Mech. Dev. 1998;77:149–164. doi: 10.1016/S0925-4773(98)00135-X. [DOI] [PubMed] [Google Scholar]
- 36.Mollema NJ, et al. Nuclear Receptor Rev-erb Alpha (Nr1d1) Functions in Concert with Nr2e3 to Regulate Transcriptional Networks in the Retina. PLoS One. 2011;6:e17494. doi: 10.1371/journal.pone.0017494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Forrest D, Swaroop A. Minireview: the role of nuclear receptors in photoreceptor differentiation and disease. Mol Endocrinol. 2012;26:905–915. doi: 10.1210/me.2012-1010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Yang Z, Wang L. Regulation of microRNA expression and function by nuclear receptor signaling. Cell Biosci. 2011;1:31. doi: 10.1186/2045-3701-1-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Nothnick WB. Steroidal regulation of uterine miRNAs is associated with modulation of the miRNA biogenesis components Exportin-5 and Dicer1. Endocrine. 2010;37:265–273. doi: 10.1007/s12020-009-9293-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mollema N, Haider N. Focus on Molecules: Nuclear hormone receptor Nr2e3: Impact on retinal development and disease. Exp. Eye Res. 2010;91:116–117. doi: 10.1016/j.exer.2010.04.013. [DOI] [PubMed] [Google Scholar]
- 41.Haider N, Naggert J, Nishina P. Excess cone cell proliferation due to lack of a functional NR2E3 causes retinal dysplasia and degeneration in rd7/rd7 mice. Hum. Mol. Genet. 2001;10:1619–1626. doi: 10.1093/hmg/10.16.1619. [DOI] [PubMed] [Google Scholar]
- 42.Haider N, et al. Nr2e3-directed transcriptional regulation of genes involved in photoreceptor development and cell-type specific phototransduction. Exp. Eye Res. 2009;89:365–372. doi: 10.1016/j.exer.2009.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Haider N, et al. The transcription factor Nr2e3 functions in retinal progenitors to suppress cone cell generation. Vis. Neurosci. 2006;23:917–929. doi: 10.1017/S095252380623027X. [DOI] [PubMed] [Google Scholar]
- 44.Corbo JC, Cepko CL. A hybrid photoreceptor expressing both rod and cone genes in a mouse model of enhanced S-cone syndrome. PLoS Genet. 2005;1:0140–0153. doi: 10.1371/journal.pgen.0010011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Fujieda H, Bremner R, Mears AJ, Sasaki H. Retinoic acid receptor-related orphan receptor alpha regulates a subset of cone genes during mouse retinal development. J. Neurochem. 2009;108:91–101. doi: 10.1111/j.1471-4159.2008.05739.x. [DOI] [PubMed] [Google Scholar]
- 46.Schorderet DF, Escher P. NR2E3 mutations in enhanced S-cone sensitivity syndrome (ESCS), Goldmann-Favre syndrome (GFS), clumped pigmentary retinal degeneration (CPRD), and retinitis pigmentosa (RP) Hum. Mutat. 2009;30:1475–1485. doi: 10.1002/humu.21096. [DOI] [PubMed] [Google Scholar]
- 47.Akhmedov NB, et al. A deletion in a photoreceptor-specific nuclear receptor mRNA causes retinal degeneration in the rd7 mouse. Proc. Natl. Acad. Sci. USA. 2000;97:5551–5556. doi: 10.1073/pnas.97.10.5551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yanagi Y, Takezawa S, Kato S. Distinct Functions of Photoreceptor Cell–Specific Nuclear Receptor, Thyroid Hormone Receptor β2 and CRX in Cone Photoreceptor Development. Invest. Ophthalmol. Vis. Sci. 2002;43:3489–3494. [PubMed] [Google Scholar]
- 49.Jun G, et al. Influence of ROBO1 and RORA on risk of age-related macular degeneration reveals genetically distinct phenotypes in disease pathophysiology. PLoS One. 2011;6:e25775. doi: 10.1371/journal.pone.0025775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Silveira AC, et al. Convergence of linkage, gene expression and association data demonstrates the influence of the RAR-related orphan receptor alpha (RORA) gene on neovascular AMD: A systems biology based approach. Vision Res. 2010;50:698–715. doi: 10.1016/j.visres.2009.09.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Schaumberg DA, et al. A prospective study of common variants in the RAR-related orphan receptor alpha (RORalpha) gene and risk of neovascular age-related macular degeneration. Arch Ophthalmol. 2010;128:1462–1471. doi: 10.1001/archophthalmol.2010.261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Nakagawa S, Watanabe M, Inoue Y. Prominent expression of nuclear hormone receptor RORa in Purkinjecells from early development. Neurosci. Res. 1997;28:177–184. doi: 10.1016/S0168-0102(97)00042-4. [DOI] [PubMed] [Google Scholar]
- 53.Dussault I, Fawcett D, Matthyssen A, Bader J, Giguere V. Orphan nuclear receptor ROR alpha-deficient mice display the cerebellar defects of staggerer. Mech Dev. 1998;70:147–53. doi: 10.1016/S0925-4773(97)00187-1. [DOI] [PubMed] [Google Scholar]
- 54.Ino H. Immunohistochemical characterization of the orphan nuclear receptor ROR alpha in the mouse nervous system. J Histochem Cytochem. 2004;52:311–23. doi: 10.1177/002215540405200302. [DOI] [PubMed] [Google Scholar]
- 55.Steinmayr M, et al. Staggerer Phenotype in Retinoid-Related Orphan Receptor Alpha-Deficient Mice. Proc. Natl. Acad. Sci. USA. 1998;95:3960–5. doi: 10.1073/pnas.95.7.3960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Cao R, et al. Role of histone H3 lysine 27 methylation in Polycomb-group silencing. Science (80-.). 2002;298:1039–43. doi: 10.1126/science.1076997. [DOI] [PubMed] [Google Scholar]
- 57.Bracken AP, et al. EZH2 is downstream of the pRB-E2F pathway, essential for proliferation and amplified in cancer. EMBO J. 2003;22:5323–5335. doi: 10.1093/emboj/cdg542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kuzmichev A, Nishioka K, Erdjument-Bromage H, Tempst P, Reinberg D. Histone methyltransferase activity associated with a human multiprotein complex containing the Enhancer of Zeste protein. Genes Dev. 2002;16:2893–2905. doi: 10.1101/gad.1035902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wang C, et al. EZH2 mediates epigenetic silencing of neuroblastoma suppressor genes CASZ1, CLU, RUNX3, and NGFR. Cancer Res. 2012;72:315–324. doi: 10.1158/0008-5472.CAN-11-0961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Surface L, Thornton S, Boyer L. Polycomb group proteins set the stage for early lineage commitment. Cell Stem Cell. 2010;7:288–98. doi: 10.1016/j.stem.2010.08.004. [DOI] [PubMed] [Google Scholar]
- 61.Sauvageau M, Sauvageau G. Polycomb group proteins: multi-faceted regulators of somatic stem cells and cancer. Cell Stem Cell. 2010;7:299–313. doi: 10.1016/j.stem.2010.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yan N, et al. Postnatal onset of retinal degeneration by loss of embryonic Ezh2 repression of Six1. Sci. Rep. 2016;6:33887. doi: 10.1038/srep33887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang, J. et al. MicroRNA regulation in an animal model of acute ocular hypertension. Acta Ophthalmol. 1–12, 10.1111/aos.13227 (2016). [DOI] [PMC free article] [PubMed]
- 64.Palfi A, et al. microRNA regulatory circuits in a mouse model of inherited retinal degeneration. Sci. Rep. 2016;6:31431. doi: 10.1038/srep31431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Tian B, et al. miR-17-3p Exacerbates Oxidative Damage in Human Retinal Pigment Epithelial Cells. PLoS One. 2016;11:eD160887. doi: 10.1371/journal.pone.0160887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jiang C, et al. MicroRNA-184 promotes differentiation of the retinal pigment epithelium by targeting the AKT2/mTOR signaling pathway. Oncotarget. 2016;7:52340–52353. doi: 10.18632/oncotarget.10566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Chung SH, Gillies M, Yam M, Wang Y, Shen W. Differential expression of microRNAs in retinal vasculopathy caused by selective Müller cell disruption. Sci. Rep. 2016;6:28993. doi: 10.1038/srep28993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Choi SW, et al. miR-410 Inhibition Induces RPE Differentiation of Amniotic Epithelial Stem Cells via Overexpression of OTX2 and RPE65. Stem Cell Rev. Reports. 2015;11:376–386. doi: 10.1007/s12015-014-9568-2. [DOI] [PubMed] [Google Scholar]
- 69.Zhou Q, et al. Strand and Cell Type-specific Function of microRNA-126 in Angiogenesis. Mol. Ther. 2016;24:1823–1835. doi: 10.1038/mt.2016.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Bhattacharjee S, Zhao Y, Dua P, Rogaev EI, Lukiw WJ. MicroRNA-34a-mediated down-regulation of the microglial-enriched triggering receptor and phagocytosis-sensor TREM2 in age-related macular degeneration. PLoS One. 2016;11:1–21. doi: 10.1371/journal.pone.0150211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Garcia TY, Gutierrez M, Reynolds J, Lamba DA. Modeling the dynamic AMD-associated chronic oxidative stress changes in human ESC and ipsc-derived RPE cells. Investig. Ophthalmol. Vis. Sci. 2015;56:7480–7488. doi: 10.1167/iovs.15-17251. [DOI] [PubMed] [Google Scholar]
- 72.Hill JM, Pogue AI, Lukiw WJ. Pathogenic microRNAs common to brain and retinal degeneration; recent observations in Alzheimer's disease and age-related macular degeneration. Front. Neurol. 2015;6:3–7. doi: 10.3389/fneur.2015.00232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Szemraj M, et al. Serum MicroRNAs as Potential Biomarkers of AMD. Med. Sci. Monit. 2015;21:2734–2742. doi: 10.12659/MSM.893697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Xu S. microRNAs and inherited retinal dystrophies. Proc. Natl. Acad. Sci. USA. 2015;112:8805–8806. doi: 10.1073/pnas.1511019112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Ohana R, et al. MicroRNAs are essential for differentiation of the retinal pigmented epithelium and maturation of adjacent photoreceptors. Development. 2015;142:2487 LP–2498. doi: 10.1242/dev.121533. [DOI] [PubMed] [Google Scholar]
- 76.Conte I, et al. MiR-204 is responsible for inherited retinal dystrophy associated with ocular coloboma. Proc. Natl. Acad. Sci. USA. 2015;112:E3236–45. doi: 10.1073/pnas.1401464112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Chung SH, et al. Profiling of Micro RNAs Involved in Retinal Degeneration Caused by Selective M??ller Cell Ablation. PLoS One. 2015;10:1–18. doi: 10.1371/journal.pone.0118949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Saxena K, Rutar MV, Provis JM, Natoli RC. Identification of miRNAs in a Model of Retinal DegenerationsmiRNAs in a Model of Retinal Degeneration. Invest. Ophthalmol. Vis. Sci. 2015;56:1820–1829. doi: 10.1167/iovs.14-15449. [DOI] [PubMed] [Google Scholar]
- 79.Tang Y, et al. Mertk Deficiency Alters Expression of microRNAs in the Retinal Pigment Epithelium Cells. Metab Brain Dis. 2015;30:943–50. doi: 10.1007/s11011-015-9653-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Murad N, et al. MiR-184 regulates ezrin, LAMP-1 expression, affects phagocytosis in human retinal pigment epithelium and is downregulated in age-related macular degeneration. FEBS J. 2014;281:5251–5264. doi: 10.1111/febs.13066. [DOI] [PubMed] [Google Scholar]
- 81.Grassmann F, et al. A Circulating MicroRNA Profile Is Associated with Late- Stage Neovascular Age-Related Macular Degeneration. PLoS One. 2014;9:e107461. doi: 10.1371/journal.pone.0107461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Genini S, Guziewicz KE, Beltran WA, Aguirre GD. Altered miRNA expression in canine retinas during normal development and in models of retinal degeneration. BMC Genomics. 2014;15:172. doi: 10.1186/1471-2164-15-172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kutty RK, et al. Differential regulation of microRNA-146a and microRNA-146b-5p in human retinal pigment epithelial cells by interleukin-1β, tumor necrosis factor-α, and interferon-γ. Mol. Vis. 2013;19:737–50. [PMC free article] [PubMed] [Google Scholar]
- 84.Lumayag S, et al. Inactivation of the microRNA-183/96/182 cluster results in syndromic retinal degeneration. Proc. Natl. Acad. Sci. USA. 2013;110:E507–16. doi: 10.1073/pnas.1212655110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Haque R, et al. MicroRNA-30b-Mediated Regulation of Catalase Expression in Human ARPE-19 Cells. PLoS One. 2012;7:e42542. doi: 10.1371/journal.pone.0042542. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lukiw WJ, Surjyadipta B, Dua P, Alexandrov PN. Common micro RNAs (miRNAs) target complement factor H (CFH) regulation in Alzheimer's disease (AD) and in agerelated macular degeneration (AMD) Int. J. Biochem. Mol. Biol. 2012;3:105–116. [PMC free article] [PubMed] [Google Scholar]
- 87.Li Y, et al. A microrna, mir133b, suppresses melanopsin expression mediated by failure dopaminergic amacrine cells in rcs rats. Cell. Signal. 2012;24:685–698. doi: 10.1016/j.cellsig.2011.10.017. [DOI] [PubMed] [Google Scholar]
- 88.Lin H, et al. Effect of miR-23 on oxidant-induced injury in human retinal pigment epithelial cells. Invest Ophthalmol Vis Sci. 2011;52:6308–6314. doi: 10.1167/iovs.10-6632. [DOI] [PubMed] [Google Scholar]
- 89.Zhou Q, et al. Regulation of angiogenesis and choroidal neovascularization by members of microRNA-23~27~24 clusters. Proc. Natl. Acad. Sci. USA. 2011;108:8287–8292. doi: 10.1073/pnas.1105254108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Arora A, et al. Prediction of microRNAs affecting mRNA expression during retinal development. BMC Dev. Biol. 2010;10:1. doi: 10.1186/1471-213X-10-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Jelcick AS, et al. Genetic variations strongly influence phenotypic outcome in the mouse retina. PLoS One. 2011;6:e21858. doi: 10.1371/journal.pone.0021858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ding J, Zhou S, Guan J. miRFam: an effective automatic miRNA classification method based on n-grams and a multiclass SVM. BMC Bioinformatics. 2011;12:216. doi: 10.1186/1471-2105-12-216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Krol J, et al. Characterizing Light-Regulated Retinal MicroRNAs Reveals Rapid Turnover as a Common Property of Neuronal MicroRNAs. Cell. 2010;141:618–631. doi: 10.1016/j.cell.2010.03.039. [DOI] [PubMed] [Google Scholar]
- 94.Peng G, Ahmad O, Ahmad F, Liu J, Chen S. The photoreceptor-specific nuclear receptor Nr2e3 interacts with Crx and exerts opposing effects on the transcription of rod versus cone genes. Human molecular genetics. Hum Mol Genet. 2005;14:747–764. doi: 10.1093/hmg/ddi070. [DOI] [PubMed] [Google Scholar]
- 95.Seo M, Choi J-S, Rho CR, Joo C-K, Lee SK. MicroRNA miR-466 inhibits Lymphangiogenesis by targeting prospero-related homeobox 1 in the alkali burn corneal injury model. J. Biomed. Sci. 2015;22:3. doi: 10.1186/s12929-014-0104-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Yu DS, et al. The regulatory role of microRNA-1187 in TNF-??-mediated hepatocyte apoptosis in acute liver failure. Int. J. Mol. Med. 2012;29:663–668. doi: 10.3892/ijmm.2012.888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kas K, Schoenmakers E, Van de Ven W. Physical map location of the human carboxypeptidase M gene (CPM) distal to D12S375 and proximal to D12S8 at chromosome 12q15. Genomics. 1995;30:403–405. [PubMed] [Google Scholar]
- 98.Sevigny J, Levesque FP, Grondin G, Beaudoin AR. Purification of the blood vessel ATP diphosphohydrolase, identification and localisation by immunological techniques. Biochim. Biophys. Acta - Gen. Subj. 1997;1334:73–88. doi: 10.1016/S0304-4165(96)00079-7. [DOI] [PubMed] [Google Scholar]
- 99.McGregor L, et al. Fraser syndrome and mouse blebbed phenotype caused by mutations in FRAS1/Fras1 encoding a putative extracellular matrix protein. Nat Genet. 2003;34:203–208. doi: 10.1038/ng1142. [DOI] [PubMed] [Google Scholar]
- 100.Bowen Ma, et al. Cloning, mapping, and characterization of activated leukocyte-cell adhesion molecule (ALCAM), a CD6 ligand. J. Exp. Med. 1995;181:2213–2220. doi: 10.1084/jem.181.6.2213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Levin TG, et al. Characterization of the intestinal cancer stem cell marker, CD166/ALCAM, in the human and mouse gastrointestinal tract. Gastroenterology. 2010;139:2072–2082. doi: 10.1053/j.gastro.2010.08.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Weiner J, et al. Axon fasciculation defects and retinal dysplasias in mice lacking the immunoglobulin superfamily adhesion molecule BEN/ALCAM/SC1. Mol Cell Neurosci. 2004;27:59–69. doi: 10.1016/j.mcn.2004.06.005. [DOI] [PubMed] [Google Scholar]
- 103.Li L, et al. The human homolog of rat Jagged1 expressed by marrow stroma inhibits differentiation of 32D cells through interaction with Notch1. Immunity. 1998;8:43–55. doi: 10.1016/S1074-7613(00)80457-4. [DOI] [PubMed] [Google Scholar]
- 104.Bao ZZ, Cepko CL. The expression and function of Notch pathway genes in the developing rat eye. J. Neurosci. 1997;17:1425–1434. doi: 10.1523/JNEUROSCI.17-04-01425.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Selcuklu SD, Donoghue MTA, Kerin MJ, Spillane C. Regulatory interplay between miR-21, JAG1 and 17beta-estradiol (E2) in breast cancer cells. Biochem. Biophys. Res. Commun. 2012;423:234–239. doi: 10.1016/j.bbrc.2012.05.074. [DOI] [PubMed] [Google Scholar]
- 106.Shilatifard a, et al. ELL2, a new member of an ELL family of RNA polymerase II elongation factors. Proc. Natl. Acad. Sci. USA. 1997;94:3639–3643. doi: 10.1073/pnas.94.8.3639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Human, M. R. C. & Genetics, B. A Novel Human Phosphoglucomutase (PGM5) Maps to the Centromeric Region of Chromosome 9. 353, 350–353 (1995). [DOI] [PubMed]
- 108.Henry S, et al. Expression Pattern and Gene Characterization of Asporin. A newly discovered member of the leucine-rich repeat protein family. J. Biol. Chem. 2001;276:12212–12221. doi: 10.1074/jbc.M011290200. [DOI] [PubMed] [Google Scholar]
- 109.Kloeckener-Gruissem B, et al. Mutation of Solute Carrier SLC16A12 Associates with a Syndrome Combining Juvenile Cataract with Microcornea and Renal Glucosuria. Am. J. Hum. Genet. 2008;82:772–779. doi: 10.1016/j.ajhg.2007.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Trimarchi JM, Cho SH, Cepko CL. Identification of genes expressed preferentially in the developing peripheral margin of the optic cup. Dev. Dyn. 2009;238:2327–2339. doi: 10.1002/dvdy.21973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Waltenberger J, Claessonwelsh L, Siegbahn A, Shibuya M, Heldin C. Different Signal-Transduction Properties of Kdr and Flt1, 2 Receptors for Vascular Endothelial Growth-Factor. J. Biol. Chem. 1994;269:26988–26995. [PubMed] [Google Scholar]
- 112.Owen LA, et al. FLT1 genetic variation predisposes to neovascular AMD in ethnically diverse populations and alters systemic FLT1 expression. Investig. Ophthalmol. Vis. Sci. 2014;55:3543–3554. doi: 10.1167/iovs.14-14047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Duncan K, et al. Expression of reverse cholesterol transport proteins ATP-binding cassette A1 (ABCA1) and scavenger receptor BI (SR-BI) in the retina and retinal pigment epithelium. Br J Ophthalmol. 2009;93:1116–1120. doi: 10.1136/bjo.2008.144006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Yu Y, et al. Association of variants in the LIPC and ABCA1 genes with intermediate and large drusen and advanced age-related macular degeneration. Investig. Ophthalmol. Vis. Sci. 2011;52:4663–4670. doi: 10.1167/iovs.10-7070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Fritsche LG, et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat. Genet. 2015;48:134–43. doi: 10.1038/ng.3448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Park Y-YY, et al. Reconstruction of nuclear receptor network reveals that NR2E3 is a novel upstream regulator of ESR1 in breast cancer. EMBO Mol. Med. 2012;4:52–67. doi: 10.1002/emmm.201100187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zhao Z, et al. Systematic Analyses of the Cytotoxic Effects of Compound 11a, a Putative Synthetic Agonist of Photoreceptor-Specific Nuclear Receptor (PNR), in Cancer Cell Lines. PLoS One. 2013;8:e75198. doi: 10.1371/journal.pone.0075198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Zhao Z, Wang L, Xu W. IL-13R[alpha]2 mediates PNR-induced migration and metastasis in ER[alpha]-negative breast cancer. Oncogene. 2015;34:1596–1607. doi: 10.1038/onc.2014.53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Cheng H, Khan NW, Roger JE, Swaroop A. Excess cones in the retinal degeneration rd7 mouse, caused by the loss of function of orphan nuclear receptor Nr2e3, originate from early-born photoreceptor precursors. Hum. Mol. Genet. 2011;20:4102–4115. doi: 10.1093/hmg/ddr334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Lu J, et al. MicroRNA expression profiles classify human cancers. Nature. 2005;435:834–838. doi: 10.1038/nature03702. [DOI] [PubMed] [Google Scholar]
- 121.Weber B, Stresemann C, Brueckner B, Lyko F. Methylation of human MicroRNA genes in normal and neoplastic cells. Cell Cycle. 2007;6:1001–1005. doi: 10.4161/cc.6.9.4209. [DOI] [PubMed] [Google Scholar]
- 122.Starlard-Davenport A, Kutanzi K, Tryndyak V, Word B, Lyn-Cook B. Restoration of the methylation status of hypermethylated gene promoters by microRNA-29b in human breast cancer: A novel epigenetic therapeutic approach. J Carcinog. 2013;12:15. doi: 10.4103/1477-3163.115720. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Podvinec M, Kaufmann MR, Handschin C, Meyer UA. NUBIScan, an in Silico Approach for Prediction of Nuclear Receptor Response Elements. Mol. Endocrinol. 2002;16:1269–1279. doi: 10.1210/mend.16.6.0851. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
The miRNA dataset supporting the conclusion of this article has been made MIAME compliant and is publicly available through the Gene Expression Omnibus (GEO) database under the series record GSE91029, and the prior microarray used to cross reference miRNA targets are available under the series record GSE24512. AMD data used as cross reference in this study is in preparation for publication.