

Abstract
Achievement in school is crucial for students to be able to pursue successful careers and lead happy lives in the future. Although many psychological attributes have been found to be associated with academic performance, the neural substrates of academic performance remain largely unknown. Here, we investigated the relationship between brain structure and academic performance in a large sample of high school students via structural magnetic resonance imaging (S-MRI) using voxel-based morphometry (VBM) approach. The whole-brain regression analyses showed that higher academic performance was related to greater regional gray matter density (rGMD) of the left dorsolateral prefrontal cortex (DLPFC), which is considered a neural center at the intersection of cognitive and non-cognitive functions. Furthermore, mediation analyses suggested that general intelligence partially mediated the impact of the left DLPFC density on academic performance. These results persisted even after adjusting for the effect of family socioeconomic status (SES). In short, our findings reveal a potential neuroanatomical marker for academic performance and highlight the role of general intelligence in explaining the relationship between brain structure and academic performance.
Introduction
Academic performance at the end of high school plays a crucial role in students’ future academics and career development. For instance, in China, the score of the Chinese National College Entrance Examination (CNCEE) (also known as Gaokao) taken at the end of high school is the sole criterion for admission to Chinese universities. Success in this examination offers not only a key opportunity for students to acquire subsequent academic and vocational achievement but also represents a critical promising opportunity for poverty-stricken families to change their fortunes1, 2. Therefore, exploring the factors related to academic performance in adolescents at the end of high school might be critical for possible reforms in education and curriculum.
Evidence from numerous studies has showed that a myriad of psychosocial factors contribute to academic performance3, which is usually measured by standardized tests (e.g., the Achievement College Test and the Stanford Achievement Test) or Grade Point Average (GPA)4, 5. Among these factors, general intelligence is the most stable and powerful predictor of academic performance6, 7. The mean correlation between general intelligence and academic performance is approximately 0.58, 9, which varies considerably depending on the variability of the measures and samples. Furthermore, several studies have shown that general intelligence plays a causal role in academic performance10–13. In this research, we sought to explore the neuroanatomical correlates of academic performance and the role of general intelligence in the association between brain anatomy and academic performance by performing structural magnetic resonance imaging (S-MRI).
Although academic performance is a popular research topic in the fields of psychology and education14, 15, the association between academic performance and the brain remains largely unknown. Neuroscientific researchers have recently begun to examine the neurobiological basis underlying measures of academic performance. An electroencephalography study first reported that, during working memory tasks, high school students with higher academic performances exhibited greater frontal energy in the theta and delta frequencies16. Furthermore, as one of the most important facilities for psychoradiology (https://radiopaedia.org/articles/psychoradiology) 17, 18, Magnetic Resonance Imaging (MRI) has been widely used to investigate the human brain. Using a longitudinal functional MRI (fMRI) study design, Horowitz-Kraus et al. (2015) found that students’ college preparedness test scores can be predicted by the activities in the prefrontal cortex (PFC) (e.g., the dorsolateral prefrontal cortex [DLPFC] and the anterior cingulate cortex [ACC]) while performing narrative-comprehension tasks at 5–7 years of age19. In addition, a recent voxel-based morphometry (VBM) study based on region of interest (ROI) analysis reported a positive association between academic performance and regional gray matter volume (rGMV) of the frontal lobe in adolescents and children aged 4 to 22 years20. In summary, these findings consistently show that the PFC, which is known to be essential for complex learning, emotion regulation and inhibition control21, 22, may predict individual differences in academic performance.
The PFC is also commonly regarded as the neural center of individual differences in general intelligence23–25. First, many investigations of individuals with deficits in intelligent behavior and cognitive function caused by brain lesions have reported that the functioning of PFC regions plays an essential role in general intelligence26–29. Moreover, in healthy subjects, general intelligence has also been found to be associated with the functional and structural variations in PFC regions, which include the DLPFC, the ventrolateral PFC (VLPFC), the medial PFC (MPFC), the ACC and the supplementary motor area (SMA) (see a meta-analysis)30. Given the causal role of general intelligence in academic performance and the relationships between the PFC regions and academic performance, general intelligence might mediate the influence of the PFC on academic performance.
To carry out our investigations, we used the real-world academic performance, the standard measurement of general intelligence and the methodology of VBM. VBM analysis is one of the most popular and valid methods for evaluating the amount of gray matter in different brain areas31. Due to its low-cost and task-free condition characteristics, VBM analysis has been popularly used to investigate the brain correlates of the human mind and behavior32, 33. Considering previously reported brain findings regarding academic performance, we speculated the structural variations in PFC regions might predict individual differences in academic performance. Specially, to ensure adequate statistical power for a whole-brain analysis, we investigated the association of academic performance with brain structures in a large sample of adolescent students (N = 214) within a narrow age range because it is hard to identify individual differences across a broad age range34. In addition, in light of the causal role of general intelligence in academic performance and the relationships between PFC regions and general intelligence, we further hypothesized that general intelligence might play a mediating role in the associations of the structural variations in the PFC regions with academic performance. Finally, we tested the specificity of our findings by excluding the effects of family socioeconomic status (SES).
Results
Brain structure of academic performance
Table 1 presents the descriptive statistics of the measures, including the mean, standard deviation, range, skewness, and kurtosis. As is customary35, the scores for the measures were normally distributed because the skewness and kurtosis values ranged from −0.51 to 0.11, with the exception of age. Age was not correlated with CNCEE score (r = −0.04, p = 0.572). Male students demonstrated higher CNCEE scores than female students [t (212) = 4.10, p < 0.001]. After controlling for gender and age, we found that the total gray matter volume (GMV) was significantly correlated with CNCEE score (r = 0.21, p = 0.003). Next, we examined the neural substrates underlying academic performance.
Table 1.
Descriptive statistics of participant-level variables.
| Variable | Mean | SD | Range | Skewness | Kurtosis |
|---|---|---|---|---|---|
| Age | 18.49 | 0.55 | 16–20 | 0.49 | 1.68 |
| Total GMV | 0.67 | 0.06 | 0.53–0.82 | 0.10 | −0.51 |
| CNCEE | 521.32 | 70.13 | 301–645 | −0.46 | 0.06 |
| Term examination | 544.38 | 45.61 | 401–633 | −0.51 | 0.11 |
| RAPM | 24.16 | 5.64 | 6–36 | −0.28 | 0.01 |
| Family SES | 5.29 | 1.49 | 1–9 | −0.14 | −0.15 |
Note: SD = standard deviation; GMV = gray matter volume; CNCEE = Chinese National College Entrance Examination; RAPM = Raven’s Advanced Progressive Matrix; SES = socioeconomic status. The values of term examination were based on 129 participants, other values were based on 214 participants.
To identify the brain areas associated with academic performance, a whole-brain multiple regression analysis was conducted between individuals’ CNCEE scores and their regional gray matter density (rGMD) values in each voxel. The results revealed that CNCEE score was positively related to the rGMD of the left DLPFC (the middle frontal gyrus) (r = 0.38, p < 0.001) (see Table 2 and Fig. 1), after controlling for gender, age and the total GMV. We observed no other significant results. To examine the stability of the association of brain structure with academic performance, we first extracted the mean rGMD values of the region identified from the whole-brain analyses and then implemented a machine learning approach with four-fold balanced cross-validation procedures. The results showed that the rGMD of the left DLPFC can reliably predict individuals’ CNCEE scores [r(predicted, observed) = 0.35, p < 0.001], after adjusting for gender, age and the total GMV.
Table 2.
Brain regions where gray matter density was associated with academic performance.
| Region | BA | Peak MNI coordinate | Peak T score | Cluster size (mm3) | ||
|---|---|---|---|---|---|---|
| x | y | z | ||||
| Correlation with CNCEE | ||||||
| Left DLPFC | 9/10 | −30 | 58 | 28 | 4.99 | 1552 |
| Correlation with term examination | ||||||
| Left DLPFC | 9/10 | −30 | 58 | 26 | 5.04 | 2224 |
Note: DLPFC = dorsolateral prefrontal cortex; BA = Brodmann’s area; MNI = Montreal Neurological Institute; CNCEE = Chinese National College Entrance Examination. We set the threshold for significant regions to p < 0.05 at the cluster level with an underlying p < 0.001 at the voxel level (non-stationary cluster correction).
Figure 1.
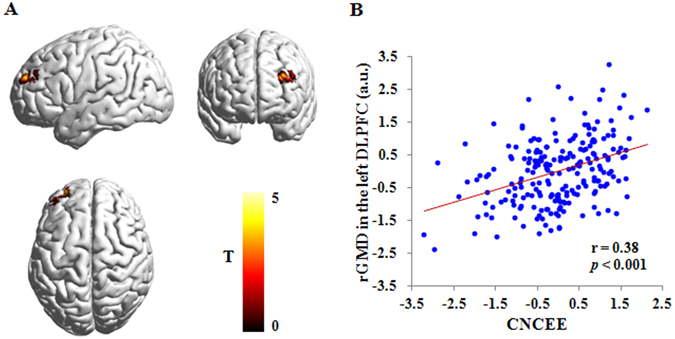
Brain regions that are related to CNCEE. (A) Brain images showing the positive association between the rGMD of the left DLPFC and CNCEE. (B) Scatter plot depicting the correlation between CNCEE and the rGMD of the left DLPFC (r = 0.38, p < 0.001). The score of horizontal axis represents the standardized residual of the CNCEE score (average standardized score of four curriculum subjects) after adjusting for age, gender and total gray matter volume. The score of vertical axis represents the standardized residual of the rGMD in the left DLPFC after adjusting for age, gender and total gray matter volume. The color bar represents the Student’s T-test (T) scores in each voxel, 0 represents the minimum T score and 5 represents the maximum T score. rGMD = regional gray matter density; DLPFC = dorsolateral prefrontal cortex; a.u. = arbitrary unit; CNCEE = Chinese National College Entrance Examination.
To test the stability of the academic performance across time, we collected term examination score in part of our participants (129 students). Behaviorally, this term examination score was highly correlated with CNCEE score (r = 0.71, p < 0.001). Neurally, the rGMD in the left DLPFC related to CNCEE score could significantly predict term examination score (r = 0.38, p < 0.001). More importantly, whole-brain regression analyses showed that term examination score was related to the rGMD in the same region (the left DLPFC; r = 0.33, p < 0.001; see Table 2 and Fig. 2), after adjusting for gender, age and the total GMV. We observed no other significant results in the whole-brain regression analyses. Then, we performed a prediction analysis to test the stability of the association between brain structure and term examination score. The results showed that the rGMD of the left DLPFC can reliably predict individuals’ term examination scores [r(predicted, observed) = 0.30, p < 0.001], after adjusting for gender, age and the total GMV. Considering the high correlation between CNCEE score and term examination score and the same brain region observed between CNCEE score and term examination score, we used only CNCEE score as the measure of academic performance in subsequent analyses.
Figure 2.
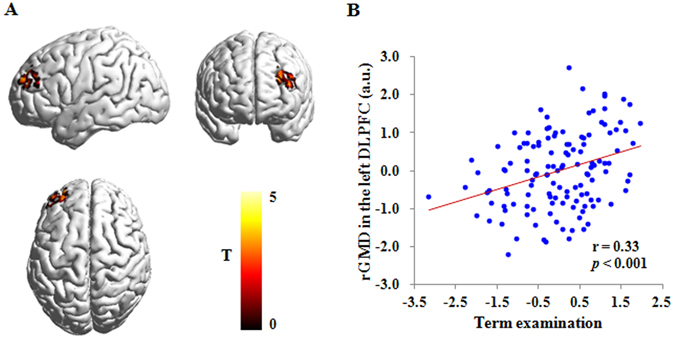
Brain regions that are related to term examination. (A) Brain images showing the positive association between the rGMD of the left DLPFC and term examination. (B) Scatter plot depicting the correlation between term examination and the rGMD of the left DLPFC (r = 0.33, p < 0.001). The score of horizontal axis represents the standardized residual of the term examination score (average standardized score of nine curriculum subjects) after adjusting for age, gender and total gray matter volume. The score of vertical axis represents the standardized residual of the rGMD in the left DLPFC after adjusting for age, gender and total gray matter volume. The color bar represents the Student’s T-test (T) scores in each voxel, 0 represents the minimum T score and 5 represents the maximum T score. rGMD = regional gray matter density; DLPFC = dorsolateral prefrontal cortex; a.u. = arbitrary unit.
Finally, to confirm the specificity of the association between academic performance and left DLPFC density, we first created a spherical ROI (radius = 10 mm) in the left DLPFC by using the coordinate of peak (−30, 58, 28) in the significant region associated with CNCEE score that was detected from the whole-brain regression analyses. Correspondingly, we created another spherical ROI (radius = 10 mm) in the right DLPFC by using the coordinate of peak (30, 58, 28). Then, we extracted the mean rGMD values in these ROIs and explored their associations with academic performance. As depicted in Fig. 3, CNCEE score was correlated with the rGMD of the left DLPFC (r = 0.36, p < 0.001) but not with the rGMD of the right DLPFC (r = 0.10, p = 0.12).
Figure 3.
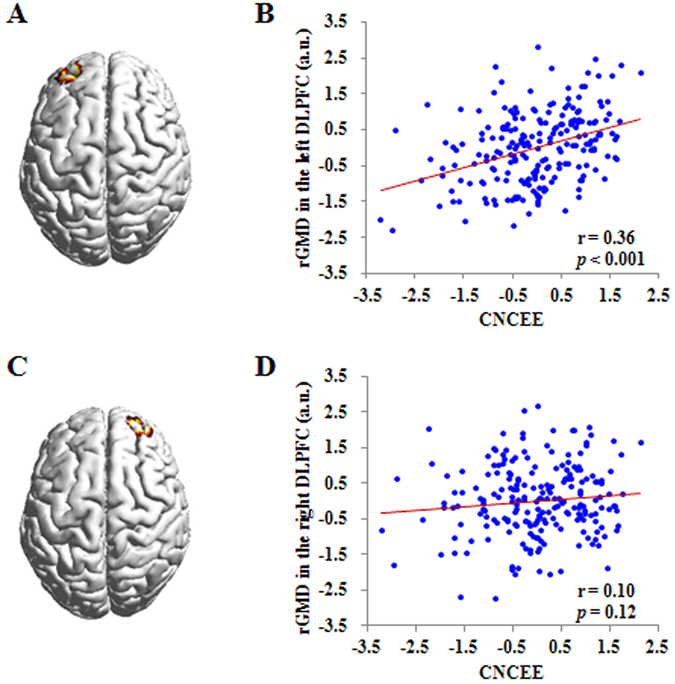
Correlation between CNCEE and the rGMD in the left DLPFC and right DLPFC. (A) Brain image showing the spherical region of interest (ROI, radius = 10 mm) in the left DLPFC, which was created by using the coordinate of peak (−30, 58, 28) in the significant region related to CNCEE from the whole-brain regression analyses. (B) Scatter plot depicting the correlation between CNCEE and the rGMD of the left DLPFC (r = 0.36, p < 0.001). The score of horizontal axis represents the standardized residual of the CNCEE score (average standardized score of four curriculum subjects) after adjusting for age, gender and total gray matter volume. The score of vertical axis represents the standardized residual of the rGMD in the left DLPFC after adjusting for age, gender and total gray matter volume. (C) Brain image showing the spherical ROI (radius = 10 mm) in the right DLPFC, which was created by using the coordinate of peak (30, 58, 28) in contrast to the left DLPFC. (D) Scatter plot depicting the correlation between CNCEE and the rGMD of the right DLPFC (r = 0.10, p = 0.12). The score of horizontal axis represents the standardized residual of the CNCEE score (average standardized score of four curriculum subjects) after adjusting for age, gender and total gray matter volume. The score of vertical axis represents the standardized residual of the rGMD in the right DLPFC after adjusting for age, gender and total gray matter volume. rGMD = regional gray matter density; DLPFC = dorsolateral prefrontal cortex; a.u. = arbitrary unit; CNCEE = Chinese National College Entrance Examination.
The relationship between brain structure and academic performance was mediated by general intelligence
To explore the role of general intelligence in the relation of brain structure and academic performance, we collected Raven’s Advanced Progressive Matrix (RAPM) data in our sample. First, we confirmed a significant correlation between general intelligence and CNCEE score (in our dataset: r = 0.38, p < 0.001). Moreover, general intelligence explained additional variance in CNCEE score (ΔR 2 = 8.7%, p < 0.001) beyond that explained by gender, age and the total GMV. Second, we tested whether the rGMD of the left DLPFC that was related to CNCEE score could predict general intelligence. The results revealed that the rGMD of the left DLPFC was significantly related to general intelligence (r = 0.34, p < 0.001). Furthermore, the left DLPFC density explained additional variance in general intelligence (ΔR 2 = 3.2%, p < 0.001) beyond that explained by gender, age and the total GMV.
These results suggested that there were close relationships among brain anatomy, academic performance and general intelligence, although the exact associations among them remain unknown. To investigate whether general intelligence could mediate the relation of brain structure and academic performance, we carried out mediation analyses with gender, age and the total GMV as controlling variables. We found that, after including general intelligence as an intermediate variable, the association of the rGMD of the left DLPFC with CNCEE score reduced, although it was still significant (see Fig. 4). The 5,000 bootstrap simulations further demonstrated that general intelligence mediated the influence of the left DLPFC on CNCEE score (indirect effect = 0.045, 95% CI = [0.01, 0.10], p < 0.05).
Figure 4.
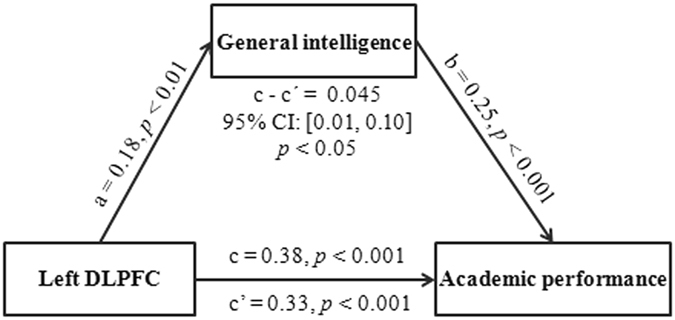
General intelligence mediates the relationship between the left DLPFC and academic performance. The panel shows that the left DLPFC affects academic performance through general intelligence. Here, we used the standardized regression coefficients to indicate the paths between variables, and all paths (a, b, c and c’) were significant. The indirect effect (c − c’ or a × b) was significant. Academic performance was measured using the Chinese National College Entrance Examination (CNCEE) score. Gender, age and the total gray matter volume were controlled for. DLPFC = dorsolateral prefrontal cortex.
Finally, we included family SES as a covariate to test the specificity of our findings. First, we checked the impact of SES on the neural correlates of academic performance. After adjusting for SES, CNCEE score was still related to the rGMD of the left DLPFC (r = 0.38, p < 0.001). Second, we investigated the influence of SES on the association of brain structure with general intelligence. The results demonstrated that the rGMD of the left DLPFC was still associated with general intelligence (r = 0.17, p = 0.01) even after adjusting for SES. Third, we examined whether SES affected the indirect effect of general intelligence on the relationship between brain structure and academic performance. The results revealed that, after controlling for SES, general intelligence still accounted for the common variance between the rGMD of the left DLPFC and CNCEE score (indirect effect = 0.043, 95% CI = [0.01, 0.09], p < 0.05). Gender, age and the total GMV were controlled for in these analyses. In summary, these results suggested that our findings were not affected by family SES.
Discussion
The current study investigated the association between academic performance and brain structure and the role of general intelligence in this association in a sample of healthy high school students. We observed two main findings. First, whole-brain regression analyses revealed that higher academic performance was correlated with greater rGMD of the left DLPFC. Second, mediation analyses demonstrated that general intelligence mediated the influence of the rGMD of the left DLPFC on academic performance. Our results persisted even after adjusting for the effect of family SES, indicating the specificity of these effects. According to our knowledge, this is the first investigation to directly examine the relationship between academic performance and rGMD in adolescent students. In brief, our study provides initial evidence for the neuroanatomical correlates of academic performance and reveals that general intelligence plays a mediating role in the association between brain structure and academic performance.
Confirming our first hypothesis, we observed that academic performance was significantly related to the rGMD of the left DLPFC. This positive association was consistent with prior studies that reported relationships between the structure and function of the DLPFC and measures of academic performance16, 20. Indeed, the DLPFC has been consistently considered as a neural hub related to psychological attributes in the intersection of cognitive and non-cognitive capacities21, 36, 37. On one hand, many prior studies have reported that the DLPFC is involved in a variety of cognitive capacities such as executive function38, 39, working memory40, 41, top-down attention42, 43, and reasoning and problem-solving44, 45. On the other hand, the DLPFC has also been linked with many non-cognitive capacities such as planning46, 47, emotional regulation48, 49, personality trait of conscientiousness50, self-regulation51, and self-esteem52. Behaviorally, it has also been revealed that academic performance apparently relies on these cognitive and non-cognitive factors3, 53–56. Therefore, the rGMD of the left DLPFC might be associated with academic performance through its relation to the cognitive and non-cognitive functions reviewed above.
In addition, our study showed that left DLPFC but not right DLPFC can predict individual differences in academic performance. One possible explanation for this result is that the measures of academic performance used in this study (i.e., CNCEE score and term examination score) mainly required participants’ language ability compared with non-language ability. This feature of the CNCEE score and term examination score may cause the result of the significant association between academic performance and the rGMD of the left DLPFC, which partly overlapped with the Broca’s area, a well known brain region for language processing in the left PFC57. Moreover, the present study only observed significant association with the left DLPFC but not with the other brain regions (e.g., temporal lobe and occipital lobe), which are found to be critical for measures of academic performance20, 34. Thus, future studies may consider using other measures of academic performance (e.g., GPA) to examine the brain basis of academic performance.
Confirming our second hypothesis, the influence of the rGMD of the left DLPFC on academic performance was mediated by general intelligence. Behaviorally, the stable relationship between general intelligence and academic performance has been repeatedly reported in previous investigations6–13, which fit well with the correlation observed in our dataset (r = 0.38, p < 0.001). Moreover, some investigators even questioned whether other psychological factors could account for additional variance in academic performance beyond the variance accounted for by general intelligence because the influence of general intelligence is so strong58. Therefore, general intelligence is likely a dominant factor in shaping the academic performance of individuals. In addition, the present study revealed a positive association of DLPFC density with general intelligence, which well corroborates previous findings that revealed relationships between the structure and function of the DLPFC and general intelligence in healthy individuals (see a meta-analysis)30. The literature reviewed above has shown that the DLPFC is implicated in many higher-order cognitive functions including executive function, working memory, top-down attention, and reasoning and problem-solving38–45. These cognitive functions have also been verified to be crucial for performing well on standard intelligence tests59–62. Taken together, our findings highlight that general intelligence might act as an underlying mechanism for explaining the relation between the structural variations in the DLPFC and academic performance.
The present study has several limitations that deserve consideration in future studies. First, we could not determine the direction of causality regarding the relationships among academic performance, general intelligence and brain structure because of the cross-sectional design used in this study. Future investigators might consider using longitudinal designs to examine the causal direction of these associations. Second, the participants included in this study were a group of high school students with a narrow age range. Therefore, future research could extend upon our study to include more diverse populations, such as primary school students and undergraduates. Third, because we used only the rGMD as the measure of brain structure, future researchers may consider employing other measures of brain structure and function to investigate the neural basis of academic performance. In particular, functional activations during task conditions (i.e., task-based fMRI) and task-free conditions (i.e., resting-state fMRI) are different from the characteristics of brain structure. It is well known that brain regions work as a functional network in the processing of corresponding psychological ability63. Given academic performance consists of various complex cognitive and non-cognitive abilities, future studies are encouraged to explore the associations between academic performance and brain networks by using task-based fMRI and resting-state fMRI.
In conclusion, we explored the brain correlates of academic performance using a VBM approach with S-MRI. We found that higher academic performance was related to greater rGMD of the left DLPFC, which sheds light on the underlying brain basis that determines academic performance. Furthermore, general intelligence mediated the impact of DLPFC density on academic performance, revealing a potential mechanism that may explain the common variance between brain structure and academic performance. These results remained significant even after adjusting for family SES, suggesting the robustness of these effects. Overall, our findings present the first evidence that rGMD serves as the neural basis of academic performance and reveal the role of general intelligence in the association between brain structure and academic performance.
Methods
Participants
Two hundred thirty-four healthy adolescent students who reported no history of psychiatric or neurological illness were recruited to participate in this study. However, twenty students with no behavioral testing scores or abnormal brain structures were excluded. Thus, a total of two hundred fourteen students (114 females; Mage = 18.49 years, SD = 0.55) were included in the data analyses. We recruited the participants from an ongoing project with primary goals of investigating the determinants of academic achievement, well-being and social cognition among adolescents in Chengdu, China. All participants were right-handed according to the Edinburgh Handedness Inventory64 and had recently graduated in June 2015 from several local public high schools. Experiments were conducted from June 2015 to September 2015, and each participant provided written informed consent before testing. This study was approved by the local research ethics committee of the West China Hospital of Sichuan University. The study protocols were performed in accordance with the approved guidelines and regulations.
Behavioral measures
Academic performance
We used the average standardized scores of CNCEE, which consists of tests of four curriculum subjects (Chinese, English, Mathematics and Comprehensive Ability), to measure participants’ academic performances. The scores of the CNCEE range from 0 to 750, which comprehensively reflect the learning processes and outcomes during three years of high school in Chinese students. The participants took the CNCEE on June 7 and June 8, 2015. We obtained the CNCEE scores from the Chengdu Education Institute database.
To ensure the stability of the academic performance across time, we collected the data of a term examination in a subsample of our participants (73 females, 56 males; Mage = 18.45 years, SD = 0.54), which are part of our larger longitudinal project. This unified term examination consists of tests of nine curriculum subjects, including Chinese, English, Mathematics, Physics, Chemistry, Biology, Politics, History, and Geography. The participants took this term examination on June 2014 to monitor learning progress. We used the average standardized scores of the nine curriculum subjects as another measure of academic performance in our dataset. These scores have been used in previous study65.
General intelligence
To assess individuals’ general intelligence, the RAPM66, which is one of the most popular and sound instruments for evaluating intelligence, was administered. This measure includes 36 non-verbal items that refer to abstract reasoning. During testing, participants were instructed to choose the missing figure for each graphical matrix within 30 minutes67. The participants completed RAPM and several questionnaires after their MRI scans. All of these tests were completed in a quiet room at the West China Hospital of Sichuan University. Two research assistants who were blind to this study supervised these testing sessions, and schoolteachers were not present. Therefore, the participants were confident about the confidentiality of their responses. RAPM scores were obtained by summing the number of correct answers, with higher scores representing higher levels of general intelligence. In our sample, this test exhibited an adequate internal reliability (Cronbach’s α = 0.82).
Family SES
To rule out the potential influence of family SES on the association between academic performance and brain structure34, 68–70, we employed a one-item scale as a graph of a ladder with ten rungs71. The participants were asked to rank the levels of their parents’ occupational prestige, income and education with ranging from 1 (lowest rank) to 10 (highest rank). Compared to objective SES, prior evidence has suggested that subjective SES exhibits a greater association with health-related measures71.
MRI data acquisition and preprocessing
Data acquisition
MRI experiments were performed using a Germany Siemens-Trio Erlangen 3.0 T MRI scanner located at the West China Hospital of Sichuan University and equipped with a 12-channel head coil. We used a magnetization-prepared rapid gradient echo sequence to obtain the T1-weighted anatomical images with the following scanning parameters: 176 slices; slice thickness, 1 mm; flip angle, 9°; matrix size, 256 × 256; repetition time, 1900 ms; inversion time, 900 ms; echo time, 2.26 ms; voxel size, 1 mm × 1 mm × 1 mm.
Data preprocessing
We preprocessed the MRI data with the Statistical Parametric Mapping program (SPM8, Wellcome Department of Cognitive Neurology, London, UK). First, a medical radiologist who was blinded to the study design visually inspected each image. Three participants were excluded because of the abnormal brain structures (e.g., unusual cyst). Second, we manually set the origin of the images to the anterior commissure for better registration. Third, we used the new segmentation in SPM8 to segment the images into white matter and gray matter. Next, we conducted registration, normalization and smoothness analyses by using Diffeomorphic Anatomical Registration Through Exponentiated Lie algebra (DARTEL) in SPM872. To do so, we aligned and resampled the gray matter images to 2 mm × 2 mm × 2 mm and then normalized them to a study specific template in the MNI152 space. Finally, we applied an 8-mm full-width at half-maximum Gaussian kernel to smooth the normalized gray matter images. The resulting images, which represent the rGMD, were used in the following analyses.
Here, we used rGMD but not rGMV as the index of gray matter. While rGMV reflects the absolute volume of the gray matter, rGMD reflects the relative concentration of the gray matter. Although the implications of the differences between rGMD and rGMV are not well known, both have been widely employed in previous structural investigations, and there is a high similarity in the results observed based on the two parameters31, 73. However, rGMD has been more frequently used in developmental studies than rGMV because the gray matter in brain areas such as the PFC is thinning during normal development74–76. Given that the participants of the current study were adolescents, which is considered as a critical period for cortical thinning77, 78, we selected rGMD as the gray matter index.
Statistical analyses
VBM analysis
To examine the associations of brain structures with academic performance, we conducted a whole-brain multiple regression analysis with the rGMD in each voxel as the dependent variable, the CNCEE score as the independent variable, and gender, age and total GMV as confounding variables. Moreover, we conducted another whole-brain multiple regression analysis with the rGMD in each voxel as the dependent variable, the term examination score as the independent variable, and gender, age and total GMV as confounding variables. In these analyses, we applied an absolute threshold mask of 0.2 to remove the edge effects between the white matter and gray matter. To determine the regions of significance, we used non-stationary cluster correction based on the random field theory79. Specifically, we set the cluster-level threshold at p < 0.05 combined with the underlying voxel-level at p < 0.001. The non-isotropic cluster-size test has been widely employed in prior studies that have analyzed VBM data80–82. We performed these analyses using SPM8 software.
Prediction analysis
We employed a machine learning approach to investigate the stability of the association between brain structure and academic performance. This approach is based on balanced cross-validation employing linear regression80, 83, 84. In the analysis, we input academic performance as the dependent variable and the rGMD of the region(s) as the independent variable. The predictive ability of the independent variable on the dependent variable was measured by r(predicted, observed), which was evaluated using a four-fold balanced cross-validation procedure. First, we divided the data into four folds to ensure that the distributions of the independent variable and the dependent variable across the folds were balanced. Second, we used three folds to build a linear regression model, with the fourth fold left out. Then, we employed the model to predict the data of the fourth fold. We repeated this procedure four times to calculate a final r(predicted, observed), which represents the association between the observed data and the data predicted by the regression model. Here, we employed a nonparametric testing method to determine the statistical significance of the model. Specially, we generated 1000 surrogate datasets to estimate the null distribution of r(predicted, observed), where the null hypothesis corresponds to no relationship between academic performance and rGMD. Then, we permuted the labels of the observed data points to generate each surrogate dataset Di of size equal to the observed dataset. Next, we used the predicted labels with the four-fold balanced cross-validation procedure and the actual labels of Di to calculate the r(predicted, observed) of Di [i.e., r(predicted, observed)i]. Finally, we counted the number of r(predicted, observed)i values greater than r(predicted, observed) and then divided that count by the number of Di datasets (1000). The resulting value was treated as the statistical significance (p value).
Mediation analysis
To confirm whether general intelligence can mediate the impact of brain anatomy on academic performance, we performed mediation analyses with the SPSS macro program85. In the standard three-variable mediation model, we included academic performance as the dependent variable, brain anatomy as the independent variable and general intelligence as the mediator variable. According to the conventions86, path c is the association of the independent variable with the dependent variable (total effect), path c’ is the association of the independent variable with the dependent variable after adjusting for the mediator variable (direct effect), and the indirect effect is equal to path a (relation of the independent variable and the mediator variable) × path b (relation of the mediator variable and the dependent variable after controlling for the independent variable) or path c - path c’. The significance of the indirect effect was determined by using the bootstrapping procedure85. Here, we used 5,000 samplings to generate 95% confidence intervals (CIs). If a CI did not contain zero, then the association between the independent variable and the dependent variable was significantly explained by the mediator variable (p < 0.05).
Acknowledgements
This study was supported by the National Natural Science Foundation (Grant Nos. 81621003, 8161101403, 81220108013, 81227002 and 81030027), Program for Changjiang Scholars and Innovative Research Team in University (PCSIRT, Grant No. IRT16R52) and the Changjiang Scholar Professorship Award (Award No. T2014190) of China. Drs. Meiyun Wang and Qiyong Gong contributed equally to playing the role of corresponding author, and in particular, Dr Qiyong Gong acknowledges the support from his China Medical Board (CMB) Distinguished Professorship Award administered by the Institute of International Education, USA.
Author Contributions
Q.Y.G. and S.W. conceived the project. S.W. and M.Z. designed the protocol and wrote the main manuscript. S.W., M.Z., T.L.C., X.Y., G.X.C. and M.Y.W. obtained the data. S.W. and M.Z. analysed the results. All authors reviewed and approved the final version of the manuscript for submission.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Song Wang and Ming Zhou contributed equally to this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Larmer, B. Inside a Chinese test-prep factory. New York Times (2014).
- 2.Zhao, Y. Who’s Afraid of the Big Bad Dragon: Why China Has the Best (and Worst) Education System in the World (John Wiley & Sons, 2014).
- 3.Richardson M, Abraham C, Bond R. Psychological Correlates of University Students’ Academic Performance: A Systematic Review and Meta-Analysis. Psychological bulletin. 2012;138:353–387. doi: 10.1037/a0026838. [DOI] [PubMed] [Google Scholar]
- 4.Fan XT, Chen M. Parental involvement and students’ academic achievement: A meta-analysis. Educ Psychol Rev. 2001;13:1–22. doi: 10.1023/A:1009048817385. [DOI] [Google Scholar]
- 5.Seipp B. Anxiety and academic performance: A meta-analysis of findings. Anxiety Research. 1991;4:27–41. doi: 10.1080/08917779108248762. [DOI] [Google Scholar]
- 6.Kuncel NR, Hezlett SA, Ones DS. Academic performance, career potential, creativity, and job performance: Can one construct predict them all? J Pers Soc Psychol. 2004;86:148–161. doi: 10.1037/0022-3514.86.1.148. [DOI] [PubMed] [Google Scholar]
- 7.Sternberg RJ, Grigorenko EL, Bundy DA. Merrill Palmer Quart. 2001. The predictive value of IQ; pp. 1–41. [Google Scholar]
- 8.Jensen, A. R. The g factor: The science of mental ability (Praeger, Westport, CT, 1998).
- 9.Neisser U, et al. Intelligence: Knowns and unknowns. Am Psychol. 1996;51:77–101. doi: 10.1037/0003-066X.51.2.77. [DOI] [Google Scholar]
- 10.Deary IJ, Strand S, Smith P, Fernandes C. Intelligence and educational achievement. Intelligence. 2007;35:13–21. doi: 10.1016/j.intell.2006.02.001. [DOI] [Google Scholar]
- 11.Freberg ME, Vandiver BJ, Watkins MW, Canivez GL. Significant factor score variability and the validity of the WISC-III Full Scale IQ in predicting later academic achievement. Appl Neuropsychol. 2008;15:131–139. doi: 10.1080/09084280802084010. [DOI] [PubMed] [Google Scholar]
- 12.Mcgrew KS, Pehl J. Prediction of Future Achievement by the Woodcock-Johnson Psycho-Educational Battery and the Wisc-R. J School Psychol. 1988;26:275–281. doi: 10.1016/0022-4405(88)90006-4. [DOI] [Google Scholar]
- 13.Watkins MW, Lei PW, Canivez GL. Psychometric intelligence and achievement: A cross-lagged panel analysis. Intelligence. 2007;35:59–68. doi: 10.1016/j.intell.2006.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lavin, D. E. The Prediction of Academic Performance: A Theoretical Analysis and Review of Research (Russell Sage Foundation, New York, 1965).
- 15.Rivkin SG, Hanushek EA, Kain JF. Teachers, schools, and academic achievement. Econometrica. 2005;73:417–458. doi: 10.1111/j.1468-0262.2005.00584.x. [DOI] [Google Scholar]
- 16.Aguirre-Perez DM, Otero-Ojeda GA, Pliego-Rivero FB, Ferreira-Martinez AA. Relationship of working memory and EEG to academic performance: A study among high school students. Int J Neurosci. 2007;117:869–882. doi: 10.1080/00207450600910077. [DOI] [PubMed] [Google Scholar]
- 17.Lui S, Zhou XJ, Sweeney JA, Gong Q. Psychoradiology: The Frontier of Neuroimaging in Psychiatry. Radiology. 2016;281:357–372. doi: 10.1148/radiol.2016152149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kressel HY. Setting Sail: 2017. Radiology. 2017;282:4–6. doi: 10.1148/radiol.2016162471. [DOI] [PubMed] [Google Scholar]
- 19.Horowitz-Kraus T, et al. Predicting better performance on a college preparedness test from narrative comprehension at the age of 6 years: An fMRI study. Brain research. 2015;1629:54–62. doi: 10.1016/j.brainres.2015.10.008. [DOI] [PubMed] [Google Scholar]
- 20.Hair NL, Hanson JL, Wolfe BL, Pollak SD. Association of Child Poverty, Brain Development, and Academic Achievement. Jama Pediatr. 2015;169:878–878. doi: 10.1001/jamapediatrics.2015.1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fuster JM. The prefrontal cortex - An update: time is of the essence. Neuron. 2001;30:319–333. doi: 10.1016/S0896-6273(01)00285-9. [DOI] [PubMed] [Google Scholar]
- 22.Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annual Review of Neuroscience. 2001;24:167–202. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- 23.Deary IJ, Penke L, Johnson W. The neuroscience of human intelligence differences. Nat Rev Neurosci. 2010;11:201–211. doi: 10.1038/nrn2793. [DOI] [PubMed] [Google Scholar]
- 24.Gray JR, Chabris CF, Braver TS. Neural mechanisms of general fluid intelligence. Nature neuroscience. 2003;6:316–322. doi: 10.1038/nn1014. [DOI] [PubMed] [Google Scholar]
- 25.Jung RE, Haier RJ. The Parieto-Frontal Integration Theory (P-FIT) of intelligence: Converging neuroimaging evidence. Behav Brain Sci. 2007;30:135–154. doi: 10.1017/S0140525X07001185. [DOI] [PubMed] [Google Scholar]
- 26.Barbey AK, et al. An integrative architecture for general intelligence and executive function revealed by lesion mapping. Brain. 2012;135:1154–1164. doi: 10.1093/brain/aws021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Duncan J. Frontal lobe function and general intelligence: Why it matters. Cortex. 2005;41:215–217. doi: 10.1016/S0010-9452(08)70896-7. [DOI] [PubMed] [Google Scholar]
- 28.Duncan J, Burgess P, Emslie H. Fluid Intelligence after Frontal-Lobe Lesions. Neuropsychologia. 1995;33:261–268. doi: 10.1016/0028-3932(94)00124-8. [DOI] [PubMed] [Google Scholar]
- 29.Roca M, et al. Executive function and fluid intelligence after frontal lobe lesions. Brain. 2010;133:234–247. doi: 10.1093/brain/awp269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Basten U, Hilger K, Fiebach CJ. Where smart brains are different: A quantitative meta-analysis of functional and structural brain imaging studies on intelligence. Intelligence. 2015;51:10–27. doi: 10.1016/j.intell.2015.04.009. [DOI] [Google Scholar]
- 31.Mechelli A, Price CJ, Friston KJ, Ashburner J. Voxel-based morphometry of the human brain: Methods and applications. Curr Med Imaging Rev. 2005;1:105–113. doi: 10.2174/1573405054038726. [DOI] [Google Scholar]
- 32.Kanai R, Rees G. OPINION The structural basis of inter-individual differences in human behaviour and cognition. Nat Rev Neurosci. 2011;12:231–242. doi: 10.1038/nrn3000. [DOI] [PubMed] [Google Scholar]
- 33.Mar RA, Spreng RN, DeYoung CG. How to produce personality neuroscience research with high statistical power and low additional cost. Cogn Affect Behav Ne. 2013;13:674–685. doi: 10.3758/s13415-013-0202-6. [DOI] [PubMed] [Google Scholar]
- 34.Mackey AP, et al. Neuroanatomical Correlates of the Income-Achievement Gap. Psychol Sci. 2015;26:925–933. doi: 10.1177/0956797615572233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Marcoulides, G. A. & Hershberger, S. L. Multivariate statistical methods: A first course. (Psychology Press, 1997).
- 36.Petrides M. Lateral prefrontal cortex: architectonic and functional organization. Philos T Roy Soc B. 2005;360:781–795. doi: 10.1098/rstb.2005.1631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Ray RD, Zald DH. Anatomical insights into the interaction of emotion and cognition in the prefrontal cortex. Neurosci Biobehav R. 2012;36:479–501. doi: 10.1016/j.neubiorev.2011.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Alvarez JA, Emory E. Executive function and the frontal lobes: A meta-analytic review. Neuropsychol Rev. 2006;16:17–42. doi: 10.1007/s11065-006-9002-x. [DOI] [PubMed] [Google Scholar]
- 39.Koechlin E, Summerfield C. An information theoretical approach to prefrontal executive function. Trends in cognitive sciences. 2007;11:229–235. doi: 10.1016/j.tics.2007.04.005. [DOI] [PubMed] [Google Scholar]
- 40.Owen, A. M., McMillan, K. M., Laird, A. R. & Bullmore, E. N-back working memory paradigm: a meta-analysis of normative functional neuroimaging studies. Human brain mapping25 (2005). [DOI] [PMC free article] [PubMed]
- 41.Petrides M. The role of the mid-dorsolateral prefrontal cortex in working memory. Exp Brain Res. 2000;133:44–54. doi: 10.1007/s002210000399. [DOI] [PubMed] [Google Scholar]
- 42.Milham MP, Banich MT, Barada V. Competition for priority in processing increases prefrontal cortex’s involvement in top-down control: an event-related fMRI study of the stroop task. Cognitive Brain Res. 2003;17:212–222. doi: 10.1016/S0926-6410(03)00108-3. [DOI] [PubMed] [Google Scholar]
- 43.Silton RL, et al. The time course of activity in dorsolateral prefrontal cortex and anterior cingulate cortex during top-down attentional control. Neuroimage. 2010;50:1292–1302. doi: 10.1016/j.neuroimage.2009.12.061. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kroger JK, et al. Recruitment of anterior dorsolateral prefrontal cortex in human reasoning: a parametric study of relational complexity. Cereb Cortex. 2002;12:477–485. doi: 10.1093/cercor/12.5.477. [DOI] [PubMed] [Google Scholar]
- 45.Newman SD, Carpenter PA, Varma S, Just MA. Frontal and parietal participation in problem solving in the Tower of London: fMRI and computational modeling of planning and high-level perception. Neuropsychologia. 2003;41:1668–1682. doi: 10.1016/S0028-3932(03)00091-5. [DOI] [PubMed] [Google Scholar]
- 46.Kaller CP, Rahm B, Spreer J, Weiller C, Unterrainer JM. Dissociable Contributions of Left and Right Dorsolateral Prefrontal Cortex in Planning. Cereb Cortex. 2011;21:307–317. doi: 10.1093/cercor/bhq096. [DOI] [PubMed] [Google Scholar]
- 47.Tanji J, Shima K, Mushiake H. Concept-based behavioral planning and the lateral prefrontal cortex. Trends in cognitive sciences. 2007;11:528–534. doi: 10.1016/j.tics.2007.09.007. [DOI] [PubMed] [Google Scholar]
- 48.Golkar A, et al. Distinct Contributions of the Dorsolateral Prefrontal and Orbitofrontal Cortex during Emotion Regulation. PloS one. 2012;7:e48107. doi: 10.1371/journal.pone.0048107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kanske P, Heissler J, Schonfelder S, Bongers A, Wessa M. How to Regulate Emotion? Neural Networks for Reappraisal and Distraction. Cereb Cortex. 2011;21:1379–1388. doi: 10.1093/cercor/bhq216. [DOI] [PubMed] [Google Scholar]
- 50.DeYoung CG, et al. Testing Predictions From Personality Neuroscience: Brain Structure and the Big Five. Psychol Sci. 2010;21:820–828. doi: 10.1177/0956797610370159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kelley WM, Wagner DD, Heatherton TF. In Search of a Human Self-Regulation System. Annu Rev Neurosci. 2015;38:389–411. doi: 10.1146/annurev-neuro-071013-014243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Agroskin D, Klackl J, Jonas E. The Self-Liking Brain: A VBM Study on the Structural Substrate of Self-Esteem. PloS one. 2014;9:e86430. doi: 10.1371/journal.pone.0086430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Duckworth AL, Seligman MEP. Self-discipline outdoes IQ in predicting academic performance of adolescents. Psychol Sci. 2005;16:939–944. doi: 10.1111/j.1467-9280.2005.01641.x. [DOI] [PubMed] [Google Scholar]
- 54.Higgins DM, Peterson JB, Pihl RO, Lee AGM. Prefrontal cognitive ability, intelligence, big five personality, and the prediction of advanced academic and workplace performance. J Pers Soc Psychol. 2007;93:298–319. doi: 10.1037/0022-3514.93.2.298. [DOI] [PubMed] [Google Scholar]
- 55.Poropat AE. A Meta-Analysis of the Five-Factor Model of Personality and Academic Performance. Psychological bulletin. 2009;135:322–338. doi: 10.1037/a0014996. [DOI] [PubMed] [Google Scholar]
- 56.Rohde TE, Thompson LA. Predicting academic achievement with cognitive ability. Intelligence. 2007;35:83–92. doi: 10.1016/j.intell.2006.05.004. [DOI] [Google Scholar]
- 57.Friederici AD. The brain basis of language processing: From structure to function. Physiol Rev. 2011;91:1357–1392. doi: 10.1152/physrev.00006.2011. [DOI] [PubMed] [Google Scholar]
- 58.Gagne F, St Pere F. When IQ is controlled, does motivation still predict achievement? Intelligence. 2002;30:71–100. doi: 10.1016/S0160-2896(01)00068-X. [DOI] [Google Scholar]
- 59.Ardila A, Pineda D, Rosselli M. Correlation between intelligence test scores and executive function measures. Arch Clin Neuropsych. 2000;15:31–36. doi: 10.1093/arclin/15.1.31. [DOI] [PubMed] [Google Scholar]
- 60.Engle RW, Tuholski SW, Laughlin JE, Conway ARA. Working memory, short-term memory, and general fluid intelligence: A latent-variable approach. J Exp Psychol Gen. 1999;128:309–331. doi: 10.1037/0096-3445.128.3.309. [DOI] [PubMed] [Google Scholar]
- 61.Schweizer K, Moosbrugger H. Attention and working memory as predictors of intelligence. Intelligence. 2004;32:329–347. doi: 10.1016/j.intell.2004.06.006. [DOI] [Google Scholar]
- 62.Sternberg, R. J. Reasoning, Problem Solving, and Intelligence In Sternberg, R. J. (Ed.), Handbook of Human Intelligence pp. 225–307 (Cambridge University Press, New York, 1982).
- 63.Bullmore ET, Sporns O. The economy of brain network organization. Nat Rev Neurosci. 2012;13:336–349. doi: 10.1038/nrn3214. [DOI] [PubMed] [Google Scholar]
- 64.Oldfield RC. The Assessment and Analysis of Handedness: The Edinburgh Inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- 65.Li, J. et al. Psychometric assessment of the short grit scale among Chinese adolescents. Journal of Psychoeducational Assessment, doi:10.1177/0734282916674858 (2016).
- 66.Raven J. The Raven’s Progressive Matrices: Change and stability over culture and time. Cognitive Psychology. 2000;41:1–48. doi: 10.1006/cogp.1999.0735. [DOI] [PubMed] [Google Scholar]
- 67.Li WF, et al. Brain structure links trait creativity to openness to experience. Social Cognitive and Affective Neuroscience. 2015;10:191–198. doi: 10.1093/scan/nsu041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Hackman DA, Farah MJ. Socioeconomic status and the developing brain. Trends in cognitive sciences. 2009;13:65–73. doi: 10.1016/j.tics.2008.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Noble KG, et al. Family income, parental education and brain structure in children and adolescents. Nature neuroscience. 2015;18:773–778. doi: 10.1038/nn.3983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sirin SR. Socioeconomic status and academic achievement: A meta-analytic review of research. Rev Educ Res. 2005;75:417–453. doi: 10.3102/00346543075003417. [DOI] [Google Scholar]
- 71.Adler NE, Epel ES, Castellazzo G, Ickovics JR. Relationship of subjective and objective social status with psychological and physiological functioning: Preliminary data in healthy white women. Health Psychol. 2000;19:586–592. doi: 10.1037/0278-6133.19.6.586. [DOI] [PubMed] [Google Scholar]
- 72.Ashburner J. A fast diffeomorphic image registration algorithm. Neuroimage. 2007;38:95–113. doi: 10.1016/j.neuroimage.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 73.Good CD, et al. Cerebral asymmetry and the effects of sex and handedness on brain structure: A voxel-based morphometric analysis of 465 normal adult human brains. Neuroimage. 2001;14:685–700. doi: 10.1006/nimg.2001.0857. [DOI] [PubMed] [Google Scholar]
- 74.Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo evidence for post-adolescent brain maturation in frontal and striatal regions. Nature neuroscience. 1999;2:859–861. doi: 10.1038/13154. [DOI] [PubMed] [Google Scholar]
- 75.Sowell ER, Thompson PM, Tessner KD, Toga AW. Mapping continued brain growth and gray matter density reduction in dorsal frontal cortex: Inverse relationships during postadolescent brain maturation. J Neurosci. 2001;21:8819–8829. doi: 10.1523/JNEUROSCI.21-22-08819.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Sowell ER, et al. Mapping cortical change across the human life span. Nature neuroscience. 2003;6:309–315. doi: 10.1038/nn1008. [DOI] [PubMed] [Google Scholar]
- 77.Fuhrmann D, Knoll LJ, Blakemore SJ. Adolescence as a Sensitive Period of Brain Development. Trends in cognitive sciences. 2015;19:558–566. doi: 10.1016/j.tics.2015.07.008. [DOI] [PubMed] [Google Scholar]
- 78.Konrad K, Firk C, Uhlhaas PJ. Brain Development During Adolescence Neuroscientific Insights Into This Developmental Period. Deutsches Arzteblatt international. 2013;110:425–431. doi: 10.3238/arztebl.2013.0425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hayasaka S, Phan KL, Liberzon I, Worsley KJ, Nichols TE. Nonstationary cluster-size inference with random field and permutation methods. Neuroimage. 2004;22:676–687. doi: 10.1016/j.neuroimage.2004.01.041. [DOI] [PubMed] [Google Scholar]
- 80.Kong, F., Chen, Z. C., Xue, S., Wang, X. & Liu, J. Mother’s but not father’s education predicts general fluid intelligence in emerging adulthood: Behavioral and neuroanatomical evidence. Human brain mapping36 (2015). [DOI] [PMC free article] [PubMed]
- 81.Takeuchi, H. et al. Regional Gray Matter Density Associated With Emotional Intelligence: Evidence From Voxel-Based Morphometry. Human brain mapping32 (2011). [DOI] [PMC free article] [PubMed]
- 82.Li HJ, et al. Examining brain structures associated with perceived stress in a large sample of young adults via voxel-based morphometry. Neuroimage. 2014;92:1–7. doi: 10.1016/j.neuroimage.2014.01.044. [DOI] [PubMed] [Google Scholar]
- 83.Supekar K, et al. Neural predictors of individual differences in response to math tutoring in primary-grade school children. Proc Natl Acad Sci USA. 2013;110:8230–8235. doi: 10.1073/pnas.1222154110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kong F, Wang X, Hu SY, Liu J. Neural correlates of psychological resilience and their relation to life satisfaction in a sample of healthy young adults. Neuroimage. 2015;123:165–172. doi: 10.1016/j.neuroimage.2015.08.020. [DOI] [PubMed] [Google Scholar]
- 85.Preacher KJ, Hayes AF. Asymptotic and resampling strategies for assessing and comparing indirect effects in multiple mediator models. Behav Res Methods. 2008;40:879–891. doi: 10.3758/BRM.40.3.879. [DOI] [PubMed] [Google Scholar]
- 86.Baron RM, Kenny DA. The Moderator Mediator Variable Distinction in Social Psychological-Research - Conceptual, Strategic, and Statistical Considerations. J Pers Soc Psychol. 1986;51:1173–1182. doi: 10.1037/0022-3514.51.6.1173. [DOI] [PubMed] [Google Scholar]