

Abstract
Cyanobacterial blooms are undesirable for ecological and health reasons. While Woronichinia naegeliana is a cyanobacterial species that appears frequently in freshwater, information about it is limited. An evaluation of its toxicity was conducted via tests based on the crustaceans Thamnocephalus platyurus and Daphnia pulex. The greatest effect of the aqueous extract obtained from W. naegeliana cells was observed for T. platyurus. The denoted semi-lethal concentration (LC50) after 24 h of exposure was 0.99 mg of dry weight (d.w.) mL−1. A lower toxicity was displayed for D. pulex, although it grew with time. Among the 18 fractions separated from cyanobacterial extract, only one containing the microginin FR3 (MG-FR3) displayed biological activeness against T. platyurus. The remaining products synthesized by W. naegeliana displayed an absence or a low level of toxicity making it impossible to determine the LC50 value. Detailed research revealed that MG-FR3 did not affect the activity of enzymes such as trypsin, chymotrypsin, elastase and thrombin, which indicates another mode of action. The results demonstrated that blooms of W. naegeliana showed toxic activity towards invertebrate zooplankton.
Keywords: Bioassays, Cyanobacteria, Cyanopeptides, Microginins
Introduction
The intensification of cyanobacterial mass occurrences in various aquatic environments is observed worldwide. Cyanobacterial blooms are serious problems, particularly for reservoirs that are a source for potable water. Cyanobacteria are known for the production of a wide range of secondary metabolites of various structure and biological activity. Among them are compounds affecting the taste and odour of water (e.g. 2-methylisoborneol [MIB] and geosmin), toxins (e.g. alkaloids, cyclic peptides) and oligopeptides, whose biological function is unclear (see references in Smith et al. 2008). Although cyanobacteria are capable of simultaneous synthesis of several compounds, only a few are dominant. Generally, cyanobacteria are recognized as toxic if they possess the ability to produce neurotoxins (e.g. anatoxin-a), dermatotoxins (e.g. aplysiatoxin) or hepatotoxins (e.g. microcystins) (Burkholder and Gilbert 2006). This classification does not concern the synthesis of some oligopeptides that contain unusual amino acids in their structures and are considered ‘non-toxic’, although some of them have been recognized as protease inhibitors. The strong inhibitory activity of cyanobacterial peptides against animal digestive proteases (e.g. elastase, chymotrypsin, trypsin) might indicate their function as anti-grazing factors, whereas the inhibition of thrombin activity, final coagulation protease, might be useful from a medical point of view in designing of new drugs (see references in Chlipala et al. 2011).
The monitoring of potential threats caused by cyanobacterial blooms relies on confirmation of the presence of toxins using mostly analytical methods. However, the application of chemical methods is often complicated due to the lack of appropriate equipment, operational procedures or commercially available standards for compounds. In addition, based only on the results obtained from chemical analyses, it is difficult to exclude the synergic effects of several toxic and non-toxic compounds released by cyanobacteria. Jungmann (1992) showed that fractions isolated from the extract of Microcystis aeruginosa cells that did not contain hepatotoxic microcystin-LR appeared to be more toxic to Daphnia sp. than those containing the compound. These may be the most probable reasons why an application of bioassays for evaluation of the bioactivity of all secondary metabolites synthesized by cyanobacteria increases in time. Additionally, relatively simple procedures keep the costs of these routinely performed analyses relatively low (Törökné et al. 2007).
Out of all the cyanobacterial species commonly occurring in water reservoirs, the most widely characterized are those frequently described as toxic (Sivonen and Jones 1999). Although blooms of the cyanobacterium Woronichinia naegeliana belonging to the Chroococcales also appear more frequently in the phytoplankton of freshwater worldwide; information about this species is limited. This is likely due to difficulties in the cultivation of this species under laboratory conditions (Rajaniemi-Wacklin et al. 2005). Bober et al. (2011) reported determination of compounds produced by W. naegeliana. The majority of its identified secondary metabolites belong to three classes of oligopeptides: microginins, cyanopeptolins and anabaenopeptins. The extract also contained trace amounts of microcystin-LR. Some studies of W. naegeliana report contradictory data about its toxicity (Rajaniemi–Wacklin et al. 2005, Willame et al. 2005, Baudin et al. 2006, Oberholster et al. 2006). However, in these studies, the main criteria for cyanobacterial biological activity were the presence of microcystins or genes responsible for its production.
Due to potentially significant consequences following the occurrence of W. naegeliana blooms in freshwater and the limited information about its biological activity, this study has been undertaken to determine its toxicity. The toxicity tests used here were based on the crustaceans Thamnocephalus platyurus and Daphnia pulex. These assays are commonly used to evaluate the toxicity of many compounds and are particularly sensitive to cyanobacterial toxins. In addition, assays on separated fractions isolated from cyanobacterial cell extract were performed to determine which compounds are responsible for the biological response. More detailed research including various protease inhibition assays was conducted on the microginin FR3, the dominant compound synthesized by W. naegeliana.
Material and methods
Cyanobacteria
Woronichinia naegeliana (Unger) Elenkin was collected from Dobczyce reservoir in southern Poland in September 2013. Identification and isolation were done according to Bober et al. (2011).
Sample preparation
The cell material was lyophilized and divided into two portions of 1 g dry weight (d.w.) each. One portion was extracted with 100 mL Milli-Q water and then filtered through GF/C glass-fibre filters (Whatman, UK). The second portion was used for separation of crude cell extract into fractions containing secondary metabolites according to the procedure described previously (Bober et al. 2011). Briefly, lyophilized cells were extracted with 100 mL 100% methanol under constant shaking and then filtered through a GF/C filter. After evaporation to dryness in a nitrogen atmosphere at room temperature, the sample was dissolved in 100 mL Milli-Q water and concentrated by solid phase extraction with a C18 silica cartridge (Baker Bond, USA). The cartridge was conditioned with 10 mL 100% methanol followed by 10 mL methanol: 10% acetic acid (4:1 v/v). The aqueous sample was passed through at 5 mL min−1 flow rate. The cartridge was then washed with 10 mL portions of 10, 20 and 30% methanol. The eluate, obtained using 10 mL 80% methanol, was evaporated to dryness in a nitrogen atmosphere at room temperature and dissolved in 1 mL Milli-Q water for analysis using a Waters Inc. (USA) high-performance liquid chromatography system containing a 600E gradient pump, 717 plus autosampler, a Jetstream 2 plus column thermostat, 996 photodiode array detector and Millenium32 SS Software with PDA option. Separation of extract was achieved using a Symmetry C18 column (4.6 × 250 mm; 5 μm; Waters, USA) maintained at 23 °C and gradient mobile phases consisting of (a) water/trifluoroacetic acid (0.05%, v/v) and (b) acetonitrile/trifluoroacetic acid (0.05%, v/v). The elution gradient was changed from 70 to 35% of eluent A over 35 min at a flow rate 0.7 mL min−1. Chromatograms were monitored at 220 nm. The fractions separated were evaporated to dryness in a nitrogen atmosphere at room temperature and diluted with Milli-Q water to the volume of the extract that was separated; therefore, their concentrations were equivalent to W. naegelina biomass concentration. The fractions were analysed using the ULPC-MS/MS system consisting of a Waters ACQUITY UPLC (Waters Inc., USA) coupled with a Waters TQD mass spectrometer (electrospray ionization mode ESI-tandem quadrupole). Chromatographic analyses were performed using the Acquity UPLC BEH (bridged ethyl hybrid) C18 column (2.1 × 100 mm; 1.7 μm) equipped with the Acquity UPLC BEH C18 VanGuard pre-column (2.1 × 5 mm; 1.7 μm). The column was maintained at 40 °C and eluted using mobile phases consisting of (a) water/formic acid (0.1%, v/v) and (b) acetonitrile/formic acid (0.1%, v/v). Gradient conditions changed from 95 to 0% of eluent A over 10 min at a flow rate of 0.3 mL min−1. The MS operational conditions were as follows: source temperature of 150 °C, desolvation temperature of 350 °C, desolvation gas flow rate of 600 L h−1, cone gas flow of 100 L h−1, capillary potential of 3.00 kV and cone potential of 20 V. Nitrogen was used for both the nebulizing and drying gas. The data were obtained in a scan mode ranging from 50 to 2000 m/z. Target ions selected for MS/MS analyses were fragmented with a collision-activated dissociation energy of 40 eV. The ion spectra were obtained by scanning from the 50 to 1100 m/z range. The MassLynx V 4.1 (Waters Inc., USA) software was used for data evaluation. The identities of the compounds presented in each fraction were determined according to Bober et al. (2011).
Toxicity assays
The dilution series of the aqueous crude extract and separated fractions were prepared with appropriate exposure media in the range of 0.1 to 10 mg d.w. mL−1. The evaluation of cyanobacterial extract toxicity was performed through the application of commercially available acute toxicity assays based on the crustaceans Thamnocephalus platyurus (THAMNOTOXKIT F) and Daphnia pulex (DAPHNOTOXKIT F) following the respective standard operational procedures (Microbiotests Inc., Belgium). The quantitative importance of the toxic effects was calculated as a 50% effective concentration. Mortality (LC50) of T. platyurus larvae was recorded after exposure to the cyanobacterial extract for 24 h, whereas dead and immobile (EC50) D. pulex were estimated after 24 and 48 h. In both assays, appropriate exposure media were used as a control. The bioassays were considered valid if the mortality or immobilization of test organisms in the controls did not exceed 10%. The toxicity of separated fractions was performed only against T. platyurus.
Enzyme inhibition assays
Detailed analysis of the toxicity mechanism used for the following enzymes, trypsin, chymotrypsin, elastase and thrombin, was performed only for the fraction, which gave a positive result in the crustaceans’ bioassays. This fraction was dissolved in ethanol and then diluted in appropriate buffers to perform the respective enzyme assays.
Trypsin inhibition assays
Inhibition assays of trypsin (EC 3.4.21.4) were performed according to the modified method of Reshef and Carmeli (2001). Trypsin was dissolved in 50 mM TRIS-HCl/100 mM NaCl/1 mM CaCl2 pH 7.5 to prepare 1 mg mL−1 solutions. This buffer was also used for the preparation of the 2 mM substrate solution of N-benzoyl-d,l-arginine-p-nitroanilide. The reaction mixture, which contained 100 μL of buffer, 10 μL of enzyme solution and 10 μL of test solution, was added to a microtiter plate well and pre-incubated at 37 °C for 5 min followed by the addition of 100 μL substrate solution. The absorbance of the solutions was measured immediately at 405 nm and after 30-min incubation at 37 °C.
Chymotrypsin inhibition assays
Inhibition assays of chymotrypsin (EC 3.4.21.1) were performed according to the method of Kisugi and Okino (2009). Briefly, chymotrypsin was dissolved in 50 mM TRIS-HCl pH 7.6 to prepare a 15 U mL−1 solution. This buffer was used for the preparation of the 1 mg mL−1 substrate solution of N-succinyl-l-phenylalanyl-p-nitroanilide. The reaction mixture, which contained 30 μL of buffer, 50 μL of enzyme solution and 10 μL of test solution, was added to a microtiter plate well and pre-incubated at 37 °C for 5 min, followed by the addition of 100 μL substrate solution. The absorbance of the solutions was measured immediately at 405 nm and after 30-min incubation at 37 °C.
Elastase inhibition assay
This inhibition assay was determined by modifying the method described by Grach-Pogrebinsky et al. (2003). The reaction mixture, which contained 150 μL of 0.2 M TRIS-HCl buffer (pH 8.0), 10 μL of elastase (EC 3.4.21.36) (0.0075 mg/mL in 0.2 M TRIS-HCl buffer (pH 8.0)) and 10 μL of test solution, was added to each microtiter plate well. Then, the mixture was pre-incubated at 30 °C for 20 min, and 30 μL of N-succinyl-Ala-Ala-Ala-p-nitroanilide (2 mM in 0.2 M TRIS-HCl buffer (pH 8.0)) was added. The absorbance of the solutions was read immediately at 405 nm and after incubation at 30 °C for 30 min.
Thrombin inhibition assay
Thrombin (EC 3.4.21.5) inhibitory activity was measured according to Anas (2012). Briefly, the reaction mixture, which contained 90 μL of thrombin from bovine plasma (1.3 U mL−1 in 0.15 M TRIS-imidazole (pH 8.2)) and 20 μL of test sample, was pre-incubated at 37 °C for 5 min. Then, 90 μL of Bz-Phe-Val-Arg-pNa.HCl (200 μg mL−1 in 0.2 M TRIS-HCl buffer (pH 8.0)) was added to begin the reaction. The absorbance was measured immediately and after incubation at 37 °C for 30 min.
Chemicals
All reagents were analytical, HPLC or MS grade and were from Sigma-Aldrich (USA) or delivered within crustacean toxicity screening tests (Microbiotests Inc., Belgium). Ultrapure grade water (Milli-Q water) was obtained from Millipore (USA).
Statistical analysis
Calculations of toxicity assays based on crustaceans were made using probit analysis (U.S. EPA 1985). Briefly, the percentages of toxic effects were plotted against log concentration, and the 50% effective concentrations were calculated using analysis of regression. Data obtained from enzyme assays were analysed using a sigmoidal dose-response curve and BioDataFit 1.02 software (Chang Bioscience Inc.). All data are expressed as the mean ± standard deviation (SD) of five replicates.
Results
Toxicity of W. naegeliana cell extract
Among the test organisms, the greatest biological sensitivity to the effect of the aqueous extract from W. naegeliana cells was displayed by T. platyurus (Table 1). The denoted lethal concentration on 50% of individuals after 24-h exposure to cyanobacterial extract was 0.99 mg d.w. mL−1. The effective concentration of 5.21 mg d.w. mL−1 caused death or immobilization of D. pulex. However, two times longer exposure to cyanobacterial extract (48 h) caused reduction of the effective concentration by 1.77 times.
Table 1.
Toxicity of the extract from Woronichinia naegeliana cells assessed with crustaceans’ bioassays expressed as 50% effective concentrations (LC50—mortality concentration; EC50—mobility inhibitory concentration). Data are expressed as mean ± SD (n = 5)
| Extract exposure | |||
|---|---|---|---|
| Organism | Time (h) | Response type | Concentration (mg dry weight mL−1) |
| Thamnocephalus platyurus | 24 | LC50 | 0.99 ± 0.25 |
| Daphnia pulex | 24 | EC50 | 5.21 ± 0.38 |
| 48 | EC50 | 2.94 ± 0.20 | |
Toxicity of fractions
The extract obtained from W. naegeliana cells was separated into 18 fractions (Fig. 1) that were used to determine which secondary metabolites are responsible for the toxicity of W. naegeliana. The bioactivity assays of separated fractions were based on the crustacean T. platyurus that was found to be more sensitive on cyanobacterial extract than D. pulex. One of the 18 fractions isolated from the cell extract exhibited biological activity in the tested concentration range, and the determined LC50 was equal to its content extracted from 5.34 mg d.w. mL−1 W. naegeliana cells. The remaining fractions containing secondary metabolites displayed a very low or no toxicity making it impossible to determine the LC50 value (Table 2). The bioactive fraction A (Fig. 2) contained pure microginin FR3 (MG-FR3) (purity 99%) for which determined LC50 was equal to 7.78 μg mL−1.
Fig. 1.

Chromatogram of extract obtained from Woronichinia naegeliana cells monitored at 220 nm. Letters match the fractions described in Table 2
Table 2.
Percent of dead Thamnocephalus platyurus larvae exposed for 24 h to separate fractions extracted from 10 mg d.w. mL−1 Woronichinia naegeliana cells. Data are expressed as mean ± SD (n = 5)
| Fraction | Fraction compositiona | Retention time (min) | (%) of dead T. platyurus larvae |
|---|---|---|---|
| A | Microginin FR3 | 9.2 | 100.0 ± 0.0 |
| B | Microginin FR4 | 10.7 | 14.0 ± 5.5 |
| C | Micropeptin T2 | 11.4 | 22.0 ± 8.4 |
| D | Micropeptin 478-B | 12.3 | 16.0 ± 11.4 |
| E | Micropeptin 88D | 12.8 | 30.0 ± 7.1 |
| F | Cyanopeptolin 880 | 13.6 | 20.0 ± 0.0 |
| G | Unknown compound 791 Da | 17.0 | 24.0 ± 5.5 |
| H | Microginin 757 | 18.1 | 16.0 ± 5.5 |
| I | Cyanopeptolin 914, unknown compound 874 Da | 19.0 | 6.0 ± 5.5 |
| J | Unknown compounds—1013 and 973 Da | 19.7 | 24.0 ± 8.9 |
| K | Micropeptin SD999, unknown compound 1039 Da | 20.4 | 22.0 ± 8.4 |
| L | Unknown compound 1047 Da | 21.8 | 32.0 ± 4.5 |
| M | Microcystin-LR, microginin 91E, planktopeptin BL1061, unknown compound 1123 Da | 23.1 | 26.0 ± 5.5 |
| N | Cyanopeptolin 908, oscillamide B | 24.0 | 10.0 ± 0.0 |
| O | Cyanopeptolin B, unknown compound 888 Da | 24.4 | 8.0 ± 4.5 |
| P | Cyanopeptolin C, unknown compound 902 Da | 27.2 | 40.0 ± 7.1 |
| Q | Microginin 51A, cyanopeptolin D, unknown compounds—1061, 1021 and 812 Da | 28.2 | 8.0 ± 4.5 |
| R | Microginin 478 | 30.0 | 8.0 ± 4.5 |
aFraction composition was determined according to Bober et al. 2011
Fig. 2.
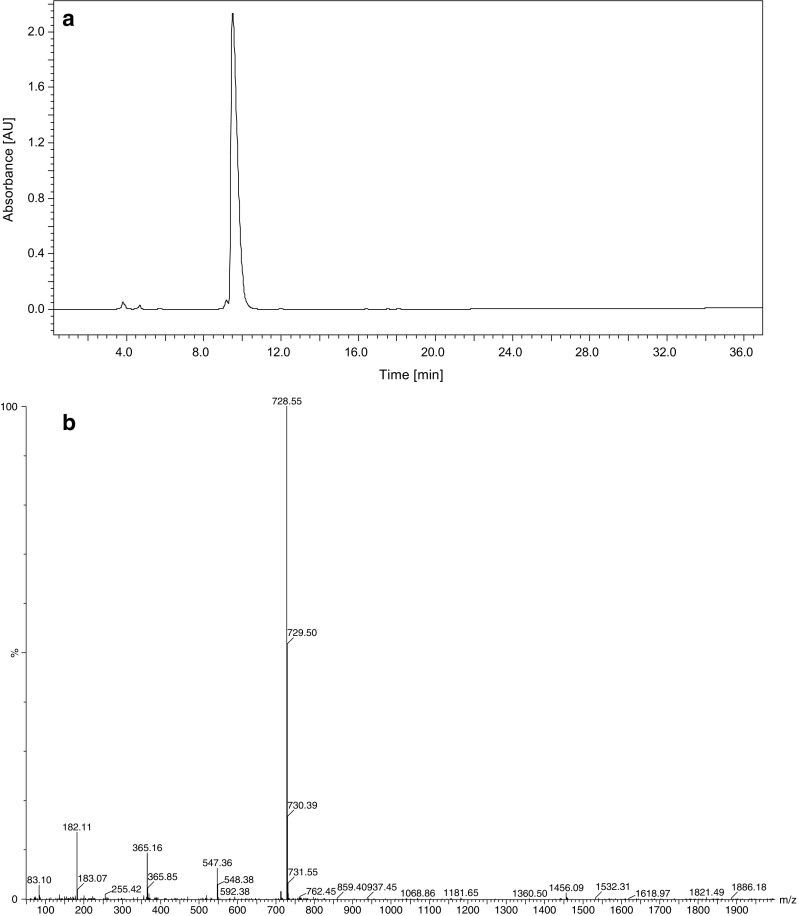
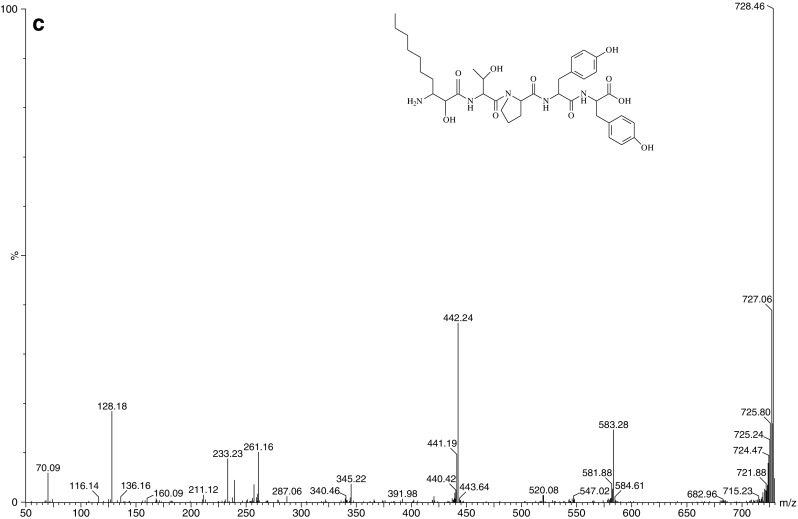
a Chromatogram of fraction A contained microginin FR3 monitored at 220 nm, b ion mass spectrum of microginin FR3 and c MS/MS product ion spectrum of microginin FR3 (m/z 728.5 Da) inside the structure of microginin FR3
The enzyme inhibitory activity of MG-FR3
Protease assays including trypsin, chymotrypsin, elastase and thrombin were performed only for the MG-FR3, which gave positive result in bioassays with T. platyurus. The MG-FR3 did not affect the activity of tested enzymes (Table 3).
Table 3.
The enzyme inhibitory activities of MG-FR3 (IC50—50% inhibitory concentration)
| Enzyme | IC50 (μg mL−1) |
|---|---|
| Trypsin | >40 |
| Chymotrypsin | >40 |
| Elastase | >40 |
| Thrombin | >100 |
Discussion
Results showed different sensitivities of tested crustaceans to aqueous extract from W. naegeliana cells. A higher biological response to cyanobacterial extract was observed for T. platyurus rather than for D. pulex (Table 1). These results are consistent with the findings of Marsálek and Bláha (2004), who found that T. platyurus revealed the highest sensitivity to cyanobacterial samples among all tested commercially available biotests. A lower sensitivity to cyanobacterial extract was observed for D. pulex, although its toxicity increased with time. Woronichinia naegeliana has been described as a potential source of toxin by Scott (1991), but to the best of our knowledge, there are no published quantitative data relating to its toxicity. We compared our results with the values presented by Marsálek and Bláha (2004), who applied similar bioassays in the assessment of Microcystis-dominated water blooms. The determined toxicity of W. naegeliana cells was classified as average (Table 4). In studies presenting data and analyses of samples collected from blooms where W. naegeliana was the dominant species among secondary metabolites, different variants of microcystins have been detected (Willame et al. 2005, Baudin et al. 2006). Although a trace amount of microcystin-LR was also detected in the studied material, its toxicity values were higher than those obtained for M. aeruginosa cells containing a high amount of this hepatotoxin. Additionally, the neurotoxicity of samples collected from Finnish lakes was statistically associated with the presence of Anabaena lemmermanni, Anabaena flos-aquae and W. naegeliana cells in the samples (Sivonen et al. 1990). Neurotoxins were not detected in the studied W. naegeliana cells. However, the possibility that the extraction procedure used and the conditions of the liquid chromatography analyses were not optimal for the detection of the neurotoxins cannot be excluded. The divergences in these results might also be due to difficulties in the separation of particular cyanobacterial species from the collected samples.
Table 4.
Comparison of the biological sensitivity of Thamnocephalus platyurus and Daphnia pulex exposed to aqueous extracts from various species of cyanobacteria. Data are presented as was described in Table 1
| Cyanobacterial extract | Thamnocephalus platyurus 24-h LC50 (mg dry weight mL−1) | Daphnia pulex 24-h EC50 (mg dry weight mL−1) |
|---|---|---|
| Woronichinia naegeliana a (99%)c | 0.99 ± 0.25 | 5.21 ± 0.38 |
| Microcystis aeruginosa b (98%)c | 0.11 ± 0.3 | 1.1 ± 1.2 |
| Microcystis ichtyoblabe (75%)c and M. aeruginosa b (20%)c | 0.35 ± 0.1 | 2.1 ± 1.2 |
| Microcystis wesenbergii b (98%)c | 3.7 ± 1.9 | 12.3 ± 2.6 |
aData obtained in this study
bData published by Maršálek and Bláha (2004)
cPercent in parenthesis represents the content of a given species in the studied cyanobacterial biomass
In the majority of the toxicity tests conducted for the secondary metabolites produced by W. naegeliana, their biological impact on T. platyurus has not been determined. The exception is for microginin FR3 (Table 2). The difference between the LC50 of separated fractions and that obtained for the total W. naegeliana extract might be the result of the synergic effect of all the compounds. Most of the metabolites synthesized by W. naegeliana (Bober et al. 2011, 2014) belong to several classes of oligopeptides, which non-ribosomal synthesis is a significant part of cell metabolism (Welker and von Döhren 2006). Although their ecological role has not been fully elucidated, their inhibitory activity against proteases indicates that they are able to affect other organisms, even causing cell lysis. As a consequence, they might be a part of a protection strategy in the aquatic ecosystem (see references in Chlipala et al. 2011).
MG-FR3 belongs to the microginins, linear peptides that contain four to six amino acid residues with a characteristic N-terminal residue 3-amino-2-hydroxydecanoic acid (Ahda) and frequently two tyrosine units at the C-terminus (Welker and von Döhren 2006). Some of these oligopeptides have been recognized as leucine aminopeptidase (LAP) inhibitors due to the 2S configuration of the Ahda group (Ishida et al. 1997; Neumann et al. 1997). The structure N-Me-Tyr-Tyr at the C-terminus seemed to be crucial for the inhibition of angiotensin-converting enzyme (ACE) (Ishida et al. 2000). The inhibitory activity of some microginins for ACE was considered for the production of blood pressure-decreasing drugs (Kraft et al. 2006). However, some microginins did not show any inhibition activity against trypsin, chymotrypsin, papain, elastase or protein phosphatase 1A (Ishida et al. 1997).
MG-FR3, which was detected in the extract from W. naegeliana cells, induced a biological response in crustaceans and therefore was used in assays including digestive proteases such as trypsin, chymotrypsin and elastase. The results are consistent with reports of other microginins showing that MG-FR3 did not affect the activity of tested enzymes (Table 3). No inhibition of thrombin was observed. Therefore, the observed toxicity of W. naegeliana towards planktonic invertebrates seems to be connected to a mode of action other than serine protease inhibition or may be a result of the synergic effect of all the produced cyanopeptides. To date, the significance of these oligopeptides to cyanobacteria metabolism and their biological function is unclear. There are several hypotheses concerning cyanopeptides, including the mechanism of the inhibition of vital enzymes, allelopathic activity on other phytoplankton organisms, chemical grazing protection and involvement in cell metabolism (see references in Halstvedt et al. 2008). However, the modes of action require further investigation.
In conclusion, the biological activity of the poorly studied freshwater bloom forming cyanobacteria W. naegeliana towards planktonic invertebrates was studied. The biological effects on planktonic crustaceans were related to a fraction containing MG-FR3. Effective assessment of the environmental risk connected to the occurrence of cyanobacterial blooms should also take into account others than those commonly known to produce toxic secondary metabolites.
Acknowledgements
The Faculty of Biochemistry, Biophysics and Biotechnology of Jagiellonian University is a partner of the Leading National Research Center (KNOW) supported by the Ministry of Science and Higher Education.
References
- Anas ARJ (2012) Studies on linear peptides from the freshwater cyanobacteria of genus Anabaena. PhD thesis, Hokkaido University
- Baudin I, Cagnard O, Grandguillaume JJ, Do-Quang Z. Algae and associated toxins & metabolites: methodology for risk assessment and risk management. Wat Pract Technol. 2006;1:1–12. [Google Scholar]
- Bober B, Lechowski Z, Bialczyk J. Determination of some cyanopeptides synthesized by Woronichinia naegeliana (Chroococcales, Cyanophyceae) Phycol Res. 2011;59:286–294. doi: 10.1111/j.1440-1835.2011.00628.x. [DOI] [Google Scholar]
- Bober B, Kaminski A, Chrapusta E, Bialczyk J. Stability of some microginins synthesized by the cyanobacterium Woronichinia naegeliana (Unger) Elenkin. Phycol Res. 2014;62:228–231. doi: 10.1111/pre.12058. [DOI] [Google Scholar]
- Burkholder JM, Glibert PM. Intraspecific variability: an important consideration in forming generalizations about toxigenic algal species. South Afr J Mar Sci. 2006;28:177–180. doi: 10.2989/18142320609504143. [DOI] [Google Scholar]
- Chlipala GE, Mo S, Orjala J. Chemodiversity in freshwater and terrestrial cyanobacteria—a source for drug discovery. Curr Drug Targets. 2011;12:1654–1673. doi: 10.2174/138945011798109455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grach-Pogrebinsky O, Sedmak B, Carmeli S. Protease inhibitors from Lake Bled toxic waterbloom of the cyanobacterium Planktothrix rubescens. Tetrahedron. 2003;59:8329–8336. doi: 10.1016/j.tet.2003.09.006. [DOI] [Google Scholar]
- Halstvedt CB, Rohrlack T, Ptacnik R, Edvardsen B. On the effect of abiotic environmental factors on production of bioactive oligopeptides in field populations of Planktothrix spp. (Cyanobacteria) J Plankton Res. 2008;5:607–617. doi: 10.1093/plankt/fbn025. [DOI] [Google Scholar]
- Ishida K, Matsuda H, Murakami M, Yamaguchi K. Microginins 299-A and -B, leucine aminopeptidase inhibitors from the cyanobacterium Microcystis aeruginosa (NIES-299) Tetrahedron. 1997;30:10281–10288. doi: 10.1016/S0040-4020(97)00684-4. [DOI] [Google Scholar]
- Ishida K, Kato T, Murakami M, Wanatabe M, Wanatabe MF. Microginins, zinc metalloproteases inhibitors from the cyanobacterium Microcystis aeruginosa. Tetrahedron. 2000;56:8643–8656. doi: 10.1016/S0040-4020(00)00770-5. [DOI] [Google Scholar]
- Jungmann D. Toxic compounds isolated from Microcystis PCC 7806 that are more active against Daphnia than two microcystins. Limnol Oceanogr. 1992;37:1777–1783. doi: 10.4319/lo.1992.37.8.1777. [DOI] [Google Scholar]
- Kisugi T, Okino T. Micropeptins from the freshwater cyanobacterium Microcystis aeruginosa (NIES-100) J Nat Prod. 2009;72:777–781. doi: 10.1021/np800631t. [DOI] [PubMed] [Google Scholar]
- Kraft M, Schleberger C, Weckesser J, Schulz GE. Binding structure of the leucine aminopeptidase inhibitor microginin FR1. FEBS Lett. 2006;580:6943–6947. doi: 10.1016/j.febslet.2006.11.060. [DOI] [PubMed] [Google Scholar]
- Marsálek B, Bláha L. Comparison of 17 biotests for detection of cyanobacterial toxicity. Environ Toxicol. 2004;19:310–317. doi: 10.1002/tox.20020. [DOI] [PubMed] [Google Scholar]
- Neumann U, Forchert A, Flury T, Weckesser J. Microginin FR1, a linear peptide from a water bloom of Microcystis species. FEMS Microbiol Lett. 1997;153:475–478. doi: 10.1111/j.1574-6968.1997.tb12612.x. [DOI] [Google Scholar]
- Oberholster PJ, Botha A-M, Cloete TE. Toxic cyanobacterial blooms in a shallow, artificially mixed urban lake in Colorado, USA. Lakes Reservoirs Res Manag. 2006;11:111–123. doi: 10.1111/j.1440-1770.2006.00297.x. [DOI] [Google Scholar]
- Rajaniemi-Wacklin P, Rantala A, Mugnai MA, Turicchia S, Ventura S, Komárková J, Lepistö L, Sivonen K. Correspondence between phylogeny and morphology of Snowella spp. and Woronichinia naegeliana, cyanobacteria commonly occurring in lakes. J Phycol. 2005;42:226–232. doi: 10.1111/j.1529-8817.2006.00179.x. [DOI] [Google Scholar]
- Reshef V, Carmeli S. Protease inhibitors from a water bloom of the cyanobacterium Microcystis aeruginosa. Tetrahedron. 2001;57:2885–2894. doi: 10.1016/S0040-4020(01)00141-7. [DOI] [PubMed] [Google Scholar]
- Scott WE. Occurence and significance of toxic cyanobacteria in southern Africa. Wat Sci Tech. 1991;23:175–180. [Google Scholar]
- Sivonen K, Jones G. Cyanobacterial toxins. In: Chorus I, Bartram J, editors. Toxic cyanobacteria in water: a guide to their public health consequences. London: Monitoring and Management. E&FN Spon; 1999. pp. 41–111. [Google Scholar]
- Sivonen K, Niemelä SI, Niemi RM, Lepistö L, Luoma TH, Räsänen LA. Toxic cyanobacteria (blue-green algae) in Finnish fresh and coastal waters. Hydrobiologia. 1990;3:256–275. [Google Scholar]
- Smith JL, Boyer GL, Zimba PV. A review of cyanobacterial odorous and bioactive metabolites: impacts and management alternatives in aquaculture. Aquaculture. 2008;280:5–20. doi: 10.1016/j.aquaculture.2008.05.007. [DOI] [Google Scholar]
- Törökné A, Vasdinnyei R, Asztalos M. A rapid microbiotest for the detection of cyanobacterial toxins. Environ Toxicol. 2007;22:64–68. doi: 10.1002/tox.20235. [DOI] [PubMed] [Google Scholar]
- U.S. EPA (1985) Method for calculation of LC50 values using probit analysis. US-EPA/600/485/013.
- Welker M, von Döhren H. Cyanobacterial peptides—nature’s own combinatorial biosynthesis. FEMS Microbiol Rev. 2006;30:530–563. doi: 10.1111/j.1574-6976.2006.00022.x. [DOI] [PubMed] [Google Scholar]
- Willame R, Jurczak T, Iffly J-F, Kull T, Meriluoto J, Hoffmann L. Distribution of hepatotoxic cyanobacterial blooms in Belgium and Luxembourg. Hydrobiologia. 2005;551:99–117. doi: 10.1007/s10750-005-4453-2. [DOI] [Google Scholar]