

Abstract
The β-barrel assembly machinery (BAM) is a multi-component complex responsible for the biogenesis of β-barrel outer membrane proteins (OMPs) in Gram-negative bacteria, with conserved systems in both mitochondria and chloroplasts. Given its importance in the integrity of the outer membrane and in the assembly of surface exposed virulence factors, BAM is an attractive therapeutic target against pathogenic bacteria, particularly multi-drug resistant strains. While the mechanism for how BAM functions remains elusive, previous structural studies have described each of the individual components of BAM, offering only a few clues to how the complex functions. Recently, a number of structures have been reported of complexes, including that of fully assembled BAM in differing conformational states. These studies have provided the molecular blueprint detailing the atomic interactions between the components and have revealed new details about BAM, which suggest a dynamic mechanism that may use conformational changes to assist in the biogenesis of new OMPs.
Keywords: β-barrel assembly machinery, BAM complex, β-barrel membrane proteins, outer membrane protein, membrane protein, protein folding, membrane biogenesis, X-ray crystallography, lateral gate, MD simulations
Graphical Abstract
The β-barrel assembly machinery (BAM) is a multi-component complex responsible for the biogenesis of β-barrel outer membrane proteins (OMPs) in Gram-negative bacteria. Recently, a number of structures have been reported of BAM in differing conformational states, revealing new details suggesting a dynamic mechanism that may use conformational changes to facilitate the biogenesis of new OMPs.
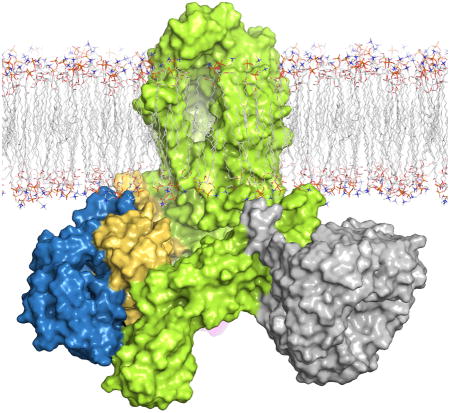
Introduction
While α-helical membrane proteins can be found in nearly all membranes, β-barrel membrane proteins can only be found in the outer membranes (OM) of Gram-negative bacteria, mitochondria, apicoplasts, and chloroplasts [1–3]. Here, these β-barrel outer membrane proteins (OMPs) can serve many functions for the cell including nutrient import and signaling. The membrane domain of OMPs consists of a linear pattern of typically an even number of antiparallel β-strands forming an elongated β-sheet, where the first and last strands interact at a junction site to form a barrel-like shape [4, 5]. While little is known about the biogenesis of OMPs, which can vary in size from 8–26 strands, much of what is known comes from studies in Gram-negative bacteria [6–8]. Here, a multi-component membrane-embedded complex called the β-barrel assembly machinery (BAM) orchestrates the folding and insertion of new OMPs into the OM. In E. coli, where most of the studies have been done, BAM consists of five components called BamA, BamB, BamC, BamD, and BamE [9–11]. BamA is an OMP itself while BamB-E are all lipoproteins anchored to the inner leaflet of the OM via an N-terminal lipid anchor. While the composition of BAM varies by species, the essential core is well conserved and consists of BamA and BamD. The other Bam proteins have been shown to be required for optimal stability and efficiency of BAM [11–14].
In Gram-negative bacteria, OMPs are first synthesized in the cytoplasm before being transported across the inner membrane (IM) into the periplasm by the Sec translocon [6, 7]. Once in the periplasm, chaperones such as SurA and/or Skp stabilize the nascent OMPs (nOMPs) and further escort them through the periplasm and peptidoglycan to the OM, where they are handed off to the BAM complex for folding and insertion. Importantly, unlike at the IM, there is no known energy source at the OM to assist in OMP biogenesis. Despite all the functional and structural studies reported to date, the exact mechanism for how BAM functions has remained elusive. However, these studies have collectively offered some clues about how BAM might operate as a molecular machine, by undergoing cyclic conformational changes that drive folding and insertion of nOMPs into the OM [15–17]. In the past decade, crystal structures of BamA and each of the four accessory proteins BamB, BamC, BamD, and BamE were reported [9, 18–34]. Together these structures revealed the architecture of the individual Bam proteins and provided the first insight about the important role of BamA, its unique structural properties, and its unprecedented conformationally dynamic behavior within a lipid membrane. Additionally, structures of two-component complexes have been reported including BamCD and a periplasmic fragment of BamA in complex with BamB and separately with BamD [35–37]. And recently, crystal structures of fully assembled BAM were reported, providing structural snapshots of several conformational states and further supporting the hypothesis of a dynamic complex [38–40].
Given the essential nature of BAM and the fact that it is found at the surface of bacteria, it is an attractive therapeutic target, particularly against pathogenic strains of Gram-negative bacteria that have acquired multi-drug resistance. Potential antimicrobial compounds could directly interact with BAM at the surface of the bacteria and would not have to cross membranes to access the intended target. While we will describe here the intricate details of the molecular structure of BAM and how it may function in OMP biogenesis, the ultimate goal is to eventually utilize these findings to help combat multi-drug resistant strains of pathogenic bacteria.
The structures of the Bam components
BamA is the largest BAM component with an N-terminal periplasmic domain composed of five polypeptide transport-associated (POTRA) domains and a 16-stranded C-terminal β-barrel transmembrane domain. Since 2007, several crystal structures of BamA from different bacterial species have been reported, including the POTRA domains and β-barrel domains individually (PDB IDs 4C4V, 4N75, 3OG5, 3Q6B, 3EFC, 2V9H, 2QCZ, 2QDF, 4K3C), as well as a full length structure of BamA from Neisseria gonorrhoeae (PDB ID 4K3B) (Fig. 1A) [18–22, 24–29, 32–34]. Interestingly, the barrel domain of all reported BamA structures exhibits a novel architecture not observed in other OMPs, where there is a significant thinning of the barrel near the junction site where strands β1 and β16 close the barrel [18, 41]. Recent studies have shown that this narrowed side of the barrel would thin and destabilize the local membrane bilayer, which may be a key feature in how BamA is used to promote OMP folding/insertion [18, 41–43]. Further, an exit pore at the extracellular side of the barrel was identified and has been proposed to mediate passage of extracellular loops of new OMPs to the extracellular surface, some of which can be as large as 100 residues [18, 30, 42]. Possibly the most revealing observation from this set of studies was that strand β16 of the β-barrel domain of BamA was found unzipped from strand β1 and pointing into the lumen of the barrel, leaving only a few hydrogen bonds tethering the barrel together. This observation suggested that the barrel domain of BamA contains a lateral gate which could mediate the insertion of new strands directly into the OM (Fig. 1A) [18, 41]. Subsequent molecular dynamics simulations and disulfide crosslinking studies are consistent with the essential role of this lateral gate of the barrel domain and the exit pore in the function of BamA [18, 30].
Figure 1. Structures of the individual Bam proteins.

A. The crystal structure of BamA from Neisseria gonorrhoeae, showing opposite faces with the junction site indicated on the right between strand β1 and β16 (highlighted in red). The barrel domain is show in green with POTRA1-5 (P1–P5) shown in different colors. A zoomed view of POTRA1 details the βααββ fold of each POTRA domain. B. The structures of BamB (gray), BamC (violet), BamD (gold), and BamE (magenta). BamB is an eight-bladed β-propeller. BamC contains three domains including an unstructured N-terminal domain of ~75 residues followed by tandem helix-grip domains. BamD is composed of five TPR motifs and BamE contains a βααβββ fold.
The N-terminal periplasmic domain of BamA contains five tandem POTRA motifs. Although the mechanistic role of these POTRA domains is still not well understood, early studies suggested that they act as a structural scaffold for binding other Bam components and for recruiting nOMPs and chaperones such as SurA (Fig. 1A) [11, 16, 44, 45]. The structure of each POTRA motif consists of a three stranded β-sheet positioned against two antiparallel helices in βααββ topology [32, 33]. Despite the lack of significant sequence similarity, each of the five POTRAs are structurally similar with the largest difference found in POTRA3, which has an extended loop between α1-α2 of unknown significance [32, 33]. The full length structure of BamA from N. gonorrhoeae shows POTRA5 located at the periphery of the bottom of the barrel, however, other structures show POTRA5 positioned away from the barrel, providing further evidence in favor of BamA being conformationally dynamic [31]. A short loop connects POTRA4 to POTRA5 via a 90° turn with the other POTRA domains extending downward from POTRA4 in a right-handed spiral (Fig. 1A) [18]. Comparison of POTRA motifs from various structures suggests an innate flexibility, particularly in the loop connecting the β-barrel to POTRA5 and in the loop that links POTRA2 and POTRA3 [34, 41]. Additionally, early studies identified that BamB and BamD each bind to the POTRA motifs at non-overlapping binding sites, and it has been suggested that binding of these accessory proteins may be used to regulate flexibility between POTRA motifs [12, 41, 46].
Crystal structures of BamB (PDB IDs 3Q7M, 3Q7N, 3Q7O, 3P1L, 3PRW, 3Q54, 2YH3, 4HDJ, 4IMM) reveal an eight-bladed β-propeller fold containing WD40-like motifs (Fig. 1B) [19–23]. While its importance has been well established in vivo and in vitro, the exact role of BamB is unknown, but has been hypothesized to be linked to substrate recruitment by possibly serving as a scaffolding protein [12, 19, 23, 47, 48]. A recent structure of a fusion protein of BamB with POTRA234 of BamA (PDB IDs 4PK1, 4XGA) is consistent with studies that indicated BamB binds to POTRA3 near the flexible linker between POTRA2 and POTRA3 [11, 12, 36]. In this position, BamB is ideally positioned to regulate conformational flexibility between POTRA2 and POTRA3, as suggested by crystal structures of BamA/B and mutagenesis studies with BamB [12].
Individual structures of BamC, BamD, and BamE have also been reported (Fig. 1B) [9, 49]. BamC contains three domains with a large unstructured N-terminal domain of ~75 residues followed by tandem helix grip domains (PDB IDs 3SNS, 2YH5, 2YH6, 2LAE, 2LAF). Interestingly, whole cell fluorescence studies have shown that the helix grip domains of BamC are presented on the extracellular surface in E. coli, however the functional significance of this is unknown [50]. BamD (PDB IDs 2YHC, 3Q5M, 3QKY) consists of five tetratricopeptide repeat (TPR) domains which are found throughout biology and are commonly used for mediating protein-protein interactions [22, 24, 51]. Likewise, BamD has been shown to directly bind both BamA and BamC, and to some client nOMPs [11, 13, 52]. A crystal structure of BamD bound to BamC lacking the C-terminal helix grip domain (PDB ID 3TGO) revealed that the unstructured N-terminus of BamC binds almost exclusively to BamD via an elongated interacting interface [35]. Using a fusion protein approach again, a crystal structure of BamD bound to POTRA45 of BamA from Rhodothermus marinus was also reported (PDB ID 5EFR), providing an early glimpse of the binding interface between BamA and BamD [37]. The structure of BamE was solved by both X-ray crystallography and NMR spectroscopy (PDB IDs 2YH9, 2KM7, 2KXX), showing an ααβββ fold [22, 28, 29]. Although BamE has been shown to enhance the binding of BamD to BamA, whether this is its primary role within BAM is unclear [14].
The structures of BAM
A significant advance in understanding BAM function has stemmed from four recent crystal structures of fully assembled BAM from E. coli [38–40]. Two of the reported structures contain all four lipoproteins (PDB IDs 5D0O, 5AYW; same space group and cell parameters), while two structures lack BamB (PDB IDs 5EKQ, 5D0Q) (Fig. 2A, B). These structures revealed the molecular interactions between the individual BAM components, confirming many that were already described either structurally and/or functionally, and uncovering novel ones. In all structures of BAM, the general domain organization and binding interfaces between individual Bam components are mostly unchanged. In each BAM structure, the periplasmic POTRA motifs of BamA extend downward below the barrel in a right-handed spiral that forms a semi-circle shape parallel to the membrane. BamD was found with its long axis also parallel to the membrane, directly interacting with both POTRA5 (primary interaction) and POTRA1 and 2, forming a ring-like structure below the β-barrel domain of BamA within the periplasm. Studies have suggested that the BamA POTRA domains and BamD may interact with nOMP substrates, therefore it was proposed that nOMPs may be recruited to this periplasmic ring as a holding/folding chamber prior to membrane insertion [39]. In the BamABCDE crystal structures, BamB binds to one side of the periplasmic ring where it interacts primarily with POTRA3 near the linker with POTRA2, almost exactly as was observed in the structure of the fusion protein between BamA and BamB [36]. Flanking the opposite side of the periplasmic ring is BamC, which was found to bind BamD similar to what was previously observed in the BamCD structure [35]. Notably, the two helix grip domains of BamC were only observed in one BamACDE structure (PDB ID 5D0Q) and not positioned within reasonable distance to be exposed on the cell surface as suggested by fluorescence studies. Rather, the C-terminal helix-grip domain interacts with the β-sheet of POTRA2, helping to further close and stabilize the ring-like conformation of the periplasmic domains of BAM. As expected, BamE was found to bind BamD, interacting primarily along TPR4 and 5. Unexpectedly, BamE also interacts with POTRA5 of BamA, which supports previous reports indicating BamE can influence the interaction of BamD with BamA [14].
Figure 2. Structural snapshots of BAM.

A. The X-ray crystal structure of BAM with BamB (PDB IDs 5D0O, 5AYW). BamA is shown in green, BamB in gray, BamC in purple, BamD in gold and BamE in magenta. B. The X-ray crystal structure of BAM lacking BamB (PDB IDs 5EKQ, 5D0Q). C. Two major differences were observed in the X-ray crystal structures in the absence or presence of BamB. The first was the large unprecedented conformational change in the barrel domain of BamA, where the first 7–8 strands undergo a 45° rotation, leading to full opening of the exit pore (‘outward open’ state of the barrel domain). And second was the positional shift of the periplasmic ring-like portion of BAM, by ~35 Å, along a plane parallel the membrane bilayer. Since there is no energy source at the outer membrane, these conformational changes may contribute to directly ushering OMPs into the membrane or by further priming the local membrane for OMP biogenesis. D. More recently, the cryoEM structure of the BAM complex with BamB was reported (PDB ID 5LJO). The overall conformation was very similar to the X-ray crystal structures lacking BamB, where the barrel domain of BamA was in an ‘outward open’ state, yet BamB was present. Several movements of the POTRA domains were observed as indicated by the black arrows. Together, these structures indicate that the conformational state of the barrel domain of BamA likely is not regulated just by the absence or presence of BamB.
The major differences observed between the two types of complexes is a positional shift of the periplasmic ring-like structure (in reference to the barrel domain) and a large conformational change in the shape of the β-barrel domain of BamA (Fig. 2C). In the crystal structures with BamB, POTRA5 associates with periplasmic turns T1 and T2 at the periphery of the β-barrel domain and the periplasmic ring is positioned directly below the barrel with open access between the periplasmic ring cavity and the lumen of the barrel. In contrast, in the crystal structures lacking BamB, the periplasmic ring-like structure is shifted such that the bottom of the β-barrel is completely occluded by POTRA5 and the periplasmic ring is offset from the barrel, with the lumen inaccessible from the periplasm. The repositioning of POTRA5 below the barrel domain is stabilized by a number of additional interactions with, periplasmic turns T1–T3 on one side and with T4, T6, and T7 on the opposite side of the barrel. Further, the shift of the periplasmic ring observed in the absence of BamB is accompanied by an unprecedented conformational change in the barrel domain of BamA, where the first seven β-strands on one side of the β-barrel (β1-β7) are rotated by ~45° with the bottom of the β-strands moving inward towards the center of the barrel and the top half of the β-strands rotated outward, starting most prominently with strand β1 and diminishing at strand β8, where strands β9-β16 are unchanged. Extracellular loops 1–3 are also flipped outward 180° to a fully open conformation along the putative exit pore. A significant consequence of this conformational change in the first half of the barrel domain of BamA is that the junction site is completely distorted, such that strands β1 and β16 no longer interact to form a sheet to close the barrel. Notably, this ‘outward open’ conformation was only observed in the crystal structures of BAM lacking BamB and supported by in vivo crosslinking studies [39, 42].
Most recently, a cryoEM structure of the BAM complex with BamB was reported with an overall architecture highly similar to the BamACDE crystal structures, where the barrel domain of BamA was also found in an ‘outward open’ conformation (PDB ID 5LJO) [53]. Compared to the X-ray crystal structures, two structural differences within the POTRA domains were observed with the first being a shift in the angle between POTRA4 and 5, which elevates the periplasmic ring slightly towards the membrane (Figure 2D). And in the second, there is a positional difference in POTRA1 with some slight movement also observed within the angle between POTRA2 and POTRA3. Importantly, the cryoEM structure indicates that gating of BamA does not depend solely on the presence of BamB, although the extent to which BamB or the other Bam accessory proteins are involved in modulating conformational states of BamA remains to be determined.
Mechanistic insights for BAM-mediated OMP insertion
Molecular dynamics (MD) simulations have played a major role in sampling the conformational plasticity of BAM in order to extract clues to exactly how BAM may function in the biogenesis of OMPs at the OM [18, 39, 42, 54]. With the structure of BamA first being reported, MD simulations revealed the possible formation of both a lateral gate along the barrel junction site and a surface exposed exit pore [42]. The importance of both the lateral gate and the exit pore were then further supported by crosslinking experiments as being essential for BamA function. Furthermore, MD simulations also indicated that loop 6 of BamA is quite stable, which conflicted with structural comparisons with FhaC, where loop 6 was found in a conformation extending towards the periplasm. However, a more recent report corrects the position of loop 6 in FhaC, which now agrees with that observed both in BamA, and TamA, a related OMP [55]. In the end, both MD simulations and subsequent crosslinking experiments verified that loop 6 does not sample multiple conformations [42]. Recent MD simulations have indicated that the POTRA domains of BamA may interact directly with the inner leaflet of the OM, which is an attractive idea since this may assist in shuttling or ushering nOMPs directly into the membrane [54]. Further, MD simulations on fully assembled BAM revealed that the full complex is most stable with all components, and that removal of components such as BamB can lead to increased dynamics, suggesting that the role of the accessory proteins may be to regulate the conformation of BamA within BAM [39].
Aligning with the recent structural reports, studies have previously indicated that BamA may need to be activated, possibly by BamD, for BAM to interact with substrate OMPs and to function properly [16]. It has also been shown that the accessory proteins BamD and BamE may regulate the conformational cycling of BamA which is required for BAM function [17]. And more recently, it was shown in vitro that BamA alone can mediate folding in a cyclic fashion via a catalytic mechanism, which has been suggested to be mostly attributed to BamA’s role in destabilizing the local membrane for OMP insertion [15, 53].
Conclusions and future directions
Together, structural, computational, and functional studies indicate that BAM likely undergoes conformational changes in a cyclic manner. Despite the distinct conformations observed for BamA in the recent structures of the BAM complex, more work is needed to determine exactly how these unprecedented conformational changes are regulated. Further, it is still unclear exactly what role these conformational changes may play. Could they be to help usher nOMPs into the membrane or possibly to further prime the local membrane for nOMP insertion? Could the differences between the X-ray and cryoEM structures just be an artefact of the detergents used for each method? Since it has been shown in several studies that BAM functions in a cyclic process, could these structural changes be responsible for the earlier observations? Further, does the ring-like structure of the periplasmic portion of BAM play a significant role in OMP biogenesis, and if so, how? Aside from the possibility that the conformational changes in BAM provide energy to usher nOMPs into the membrane, it is difficult to envision the importance of this quaternary assembly itself just by the correlation to the circular shape of an OMP, which can vary in size from 8–26 strands. And, while the proposed mechanisms may address monomeric OMPs, they don’t necessarily account for multimeric OMPs such as YadA or TolC. Could these OMPs follow a different pathway as has been proposed or could BAM clusters orchestrate their assembly? Another longstanding question is at what step in the biogenesis pathway do OMPs really begin folding? It is feasible to think that an OMP could begin folding as soon as it enters the periplasm and reaches out to BAM at the OM. This would be efficient and would prevent accumulation of nOMPs in the periplasm. However, it has generally been accepted that the C-terminal strand of an OMP (termed the β-signal) is likely the recognition signal, possibly even initiating biogenesis into the OM. But if this is true, all nOMPs would need to be fully translocated into the periplasm and stabilized by chaperones until the β-signal arrives to initiate biogenesis by BAM. One obvious advantage of this idea is that by requiring the β-signal to initiate biogenesis, only those nOMPs that are fully translated and translocated into the periplasm ever make it to the OM. This may serve as a checkpoint to prevent misfolded or incomplete OMPs from being inserted into the OM, which could make the cell vulnerable to host defenses and environmental hazards.
The past decade has seen an explosion of structural and functional studies investigating the BAM complex and the biogenesis pathway of OMPs. In the past year, new structures of fully assembled BAM have pushed the field even further by providing the molecular details for how each of the components interacts with the others. These structures have also revealed unprecedented conformational changes in BamA, which may play a significant role in how BAM is able to mediate folding/insertion of nOMPs into the OM. While there is certainly more structural work to be done, the arsenal of structures currently available will certainly synergize current and future functional and mechanistic studies to determine exactly how BAM operates. Further, these studies will continue to offer clues about the conserved machineries responsible for the biogenesis of OMPs in both mitochondria and chloroplasts.
Acknowledgments
JB and NN are supported by the Department of Biological Sciences at Purdue University, a Showalter Trust Award, Pilot Funding provided by the Indiana Clinical and Translational Sciences Institute, and by the National Institute of Allergy and Infectious Diseases (1K22AI113078-01); SKB is supported by the Intramural Research Program of the National Institutes of Health, National Institute of Diabetes and Digestive and Kidney Diseases.
Abbreviations
- BAM
β-barrel assembly machinery
- OMP
β-barrel outer membrane protein
- POTRA
polypeptide-transport associated
- TPR
tetratricopeptide repeat
- nOMP
nascent OMP
Footnotes
Conflicts of Interest
The authors declare no conflicts of interest.
Author Contributions
J.B., S.K.B., and N.N. contributed to writing the paper.
References
- 1.Gabriel K, Buchanan SK, Lithgow T. The alpha and the beta: protein translocation across mitochondrial and plastid outer membranes. Trends Biochem Sci. 2001;26:36–40. doi: 10.1016/s0968-0004(00)01684-4. [DOI] [PubMed] [Google Scholar]
- 2.Schleiff E, Soll J. Membrane protein insertion: mixing eukaryotic and prokaryotic concepts. EMBO Rep. 2005;6:1023–7. doi: 10.1038/sj.embor.7400563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Misra R. Assembly of the beta-Barrel Outer Membrane Proteins in Gram-Negative Bacteria, Mitochondria, and Chloroplasts. ISRN molecular biology. 2012;2012:708203. doi: 10.5402/2012/708203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fairman JW, Noinaj N, Buchanan SK. The structural biology of β-barrel membrane proteins: a summary of recent reports. Curr Opin Struct Biol. 2011;21:523–531. doi: 10.1016/j.sbi.2011.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schulz GE. beta-Barrel membrane proteins. Curr Opin Struct Biol. 2000;10:443–7. doi: 10.1016/s0959-440x(00)00120-2. [DOI] [PubMed] [Google Scholar]
- 6.Knowles TJ, Scott-Tucker A, Overduin M, Henderson IR. Membrane protein architects: the role of the BAM complex in outer membrane protein assembly. Nat Rev Microbiol. 2009;7:206–14. doi: 10.1038/nrmicro2069. [DOI] [PubMed] [Google Scholar]
- 7.Ricci DP, Silhavy TJ. The Bam machine: A molecular cooper. Biochim Biophys Acta. 2012;1818:1067–84. doi: 10.1016/j.bbamem.2011.08.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hagan CL, Silhavy TJ, Kahne D. beta-Barrel membrane protein assembly by the Bam complex. Annu Rev Biochem. 2011;80:189–210. doi: 10.1146/annurev-biochem-061408-144611. [DOI] [PubMed] [Google Scholar]
- 9.O’Neil PK, Rollauer SE, Noinaj N, Buchanan SK. Fitting the Pieces of the beta-Barrel Assembly Machinery Complex. Biochemistry. 2015;54:6303–11. doi: 10.1021/acs.biochem.5b00852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Rollauer SE, Sooreshjani MA, Noinaj N, Buchanan SK. Outer membrane protein biogenesis in Gram-negative bacteria. Philos Trans R Soc Lond B Biol Sci. 2015:370. doi: 10.1098/rstb.2015.0023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wu T, Malinverni J, Ruiz N, Kim S, Silhavy TJ, Kahne D. Identification of a Multicomponent Complex Required for Outer Membrane Biogenesis in Escherichia coli. Cell. 2005;121:235–245. doi: 10.1016/j.cell.2005.02.015. [DOI] [PubMed] [Google Scholar]
- 12.Vuong P, Bennion D, Mantei J, Frost D, Misra R. Analysis of YfgL and YaeT interactions through bioinformatics, mutagenesis, and biochemistry. J Bacteriol. 2008;190:1507–17. doi: 10.1128/JB.01477-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Malinverni JC, Werner J, Kim S, Sklar JG, Kahne D, Misra R, Silhavy TJ. YfiO stabilizes the YaeT complex and is essential for outer membrane protein assembly in Escherichia coli. Mol Microbiol. 2006;61:151–64. doi: 10.1111/j.1365-2958.2006.05211.x. [DOI] [PubMed] [Google Scholar]
- 14.Sklar JG, Wu T, Gronenberg LS, Malinverni JC, Kahne D, Silhavy TJ. Lipoprotein SmpA is a component of the YaeT complex that assembles outer membrane proteins in Escherichia coli. Proceedings of the National Academy of Sciences. 2007;104:6400–6405. doi: 10.1073/pnas.0701579104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Plummer AM, Fleming KG. BamA Alone Accelerates Outer Membrane Protein Folding In Vitro through a Catalytic Mechanism. Biochemistry. 2015;54:6009–11. doi: 10.1021/acs.biochem.5b00950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ricci DP, Hagan CL, Kahne D, Silhavy TJ. Activation of the Escherichia coli beta-barrel assembly machine (Bam) is required for essential components to interact properly with substrate. Proc Natl Acad Sci U S A. 2012;109:3487–91. doi: 10.1073/pnas.1201362109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Rigel NW, Ricci DP, Silhavy TJ. Conformation-specific labeling of BamA and suppressor analysis suggest a cyclic mechanism for beta-barrel assembly in Escherichia coli. Proc Natl Acad Sci U S A. 2013;110:5151–6. doi: 10.1073/pnas.1302662110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Noinaj N, Kuszak AJ, Gumbart JC, Lukacik P, Chang H, Easley NC, Lithgow T, Buchanan SK. Structural insight into the biogenesis of beta-barrel membrane proteins. Nature. 2013;501:385–390. doi: 10.1038/nature12521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Noinaj N, Fairman JW, Buchanan SK. The Crystal Structure of BamB Suggests Interactions with BamA and Its Role within the BAM Complex. J Mol Biol. 2011;407:248–60. doi: 10.1016/j.jmb.2011.01.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Heuck A, Schleiffer A, Clausen T. Augmenting beta-Augmentation: Structural Basis of How BamB Binds BamA and May Support Folding of Outer Membrane Proteins. J Mol Biol. 2011;406:659–66. doi: 10.1016/j.jmb.2011.01.002. [DOI] [PubMed] [Google Scholar]
- 21.Kim KH, Paetzel M. Crystal Structure of Escherichia coli BamB, a Lipoprotein Component of the beta-Barrel Assembly Machinery Complex. J Mol Biol. 2011;406:667–78. doi: 10.1016/j.jmb.2010.12.020. [DOI] [PubMed] [Google Scholar]
- 22.Albrecht R, Zeth K. Structural basis of outer membrane protein biogenesis in bacteria. J Biol Chem. 2011;286:27792–803. doi: 10.1074/jbc.M111.238931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jansen KB, Baker SL, Sousa MC. Crystal structure of BamB from Pseudomonas aeruginosa and functional evaluation of its conserved structural features. PLoS One. 2012;7:e49749. doi: 10.1371/journal.pone.0049749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sandoval CM, Baker SL, Jansen K, Metzner SI, Sousa MC. Crystal structure of BamD: an essential component of the beta-Barrel assembly machinery of gram-negative bacteria. J Mol Biol. 2011;409:348–57. doi: 10.1016/j.jmb.2011.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Dong C, Hou HF, Yang X, Shen YQ, Dong YH. Structure of Escherichia coli BamD and its functional implications in outer membrane protein assembly. Acta Crystallogr D Biol Crystallogr. 2012;68:95–101. doi: 10.1107/S0907444911051031. [DOI] [PubMed] [Google Scholar]
- 26.Kim KH, Aulakh S, Tan W, Paetzel M. Crystallographic analysis of the C-terminal domain of the Escherichia coli lipoprotein BamC. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011;67:1350–8. doi: 10.1107/S174430911103363X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Warner LR, Varga K, Lange OF, Baker SL, Baker D, Sousa MC, Pardi A. Structure of the BamC two-domain protein obtained by Rosetta with a limited NMR data set. J Mol Biol. 2011;411:83–95. doi: 10.1016/j.jmb.2011.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kim KH, Kang HS, Okon M, Escobar-Cabrera E, McIntosh LP, Paetzel M. Structural characterization of Escherichia coli BamE, a lipoprotein component of the beta-barrel assembly machinery complex. Biochemistry. 2011;50:1081–90. doi: 10.1021/bi101659u. [DOI] [PubMed] [Google Scholar]
- 29.Knowles TJ, Browning DF, Jeeves M, Maderbocus R, Rajesh S, Sridhar P, Manoli E, Emery D, Sommer U, Spencer A, Leyton DL, Squire D, Chaudhuri RR, Viant MR, Cunningham AF, Henderson IR, Overduin M. Structure and function of BamE within the outer membrane and the beta-barrel assembly machine. EMBO Rep. 2011;12:123–8. doi: 10.1038/embor.2010.202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ni D, Wang Y, Yang X, Zhou H, Hou X, Cao B, Lu Z, Zhao X, Yang K, Huang Y. Structural and functional analysis of the β-barrel domain of BamA from Escherichia coli. The FASEB Journal. 2014;28:2677–2685. doi: 10.1096/fj.13-248450. [DOI] [PubMed] [Google Scholar]
- 31.Albrecht R, Schutz M, Oberhettinger P, Faulstich M, Bermejo I, Rudel T, Diederichs K, Zeth K. Structure of BamA, an essential factor in outer membrane protein biogenesis. Acta Crystallographica Section D. 2014;70:1779–1789. doi: 10.1107/S1399004714007482. [DOI] [PubMed] [Google Scholar]
- 32.Gatzeva-Topalova PZ, Walton TA, Sousa MC. Crystal Structure of YaeT: Conformational Flexibility and Substrate Recognition. Structure. 2008;16:1873–1881. doi: 10.1016/j.str.2008.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kim S, Malinverni JC, Sliz P, Silhavy TJ, Harrison SC, Kahne D. Structure and Function of an Essential Component of the Outer Membrane Protein Assembly Machine. Science. 2007;317:961–964. doi: 10.1126/science.1143993. [DOI] [PubMed] [Google Scholar]
- 34.Gatzeva-Topalova PZ, Warner LR, Pardi A, Sousa MC. Structure and flexibility of the complete periplasmic domain of BamA: the protein insertion machine of the outer membrane. Structure. 2010;18:1492–501. doi: 10.1016/j.str.2010.08.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim KH, Aulakh S, Paetzel M. Crystal structure of beta-barrel assembly machinery BamCD protein complex. J Biol Chem. 2011;286:39116–21. doi: 10.1074/jbc.M111.298166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jansen KB, Baker SL, Sousa MC. Crystal structure of BamB bound to a periplasmic domain fragment of BamA, the central component of the beta-barrel assembly machine. J Biol Chem. 2015;290:2126–36. doi: 10.1074/jbc.M114.584524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bergal HT, Hopkins AH, Metzner SI, Sousa MC. The Structure of a BamA-BamD Fusion Illuminates the Architecture of the beta-Barrel Assembly Machine Core. Structure. 2016;24:243–51. doi: 10.1016/j.str.2015.10.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bakelar J, Buchanan SK, Noinaj N. The structure of the beta-barrel assembly machinery complex. Science. 2016;351:180–6. doi: 10.1126/science.aad3460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Gu Y, Li H, Dong H, Zeng Y, Zhang Z, Paterson NG, Stansfeld PJ, Wang Z, Zhang Y, Wang W, Dong C. Structural basis of outer membrane protein insertion by the BAM complex. Nature. 2016;531:64–9. doi: 10.1038/nature17199. [DOI] [PubMed] [Google Scholar]
- 40.Han L, Zheng J, Wang Y, Yang X, Liu Y, Sun C, Cao B, Zhou H, Ni D, Lou J, Zhao Y, Huang Y. Structure of the BAM complex and its implications for biogenesis of outer-membrane proteins. Nat Struct Mol Biol. 2016;23:192–6. doi: 10.1038/nsmb.3181. [DOI] [PubMed] [Google Scholar]
- 41.Noinaj N, Rollauer SE, Buchanan SK. The beta-barrel membrane protein insertase machinery from Gram-negative bacteria. Curr Opin Struct Biol. 2015;31:35–42. doi: 10.1016/j.sbi.2015.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Noinaj N, Kuszak Adam J, Balusek C, Gumbart James C, Buchanan Susan K. Lateral Opening and Exit Pore Formation Are Required for BamA Function. Structure. 2014;22:1055–1062. doi: 10.1016/j.str.2014.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sinnige T, Weingarth M, Renault M, Baker L, Tommassen J, Baldus M. Solid-State NMR Studies of Full-Length BamA in Lipid Bilayers Suggest Limited Overall POTRA Mobility. J Mol Biol. 2014 doi: 10.1016/j.jmb.2014.02.007. [DOI] [PubMed] [Google Scholar]
- 44.Knowles TJ, Jeeves M, Bobat S, Dancea F, McClelland D, Palmer T, Overduin M, Henderson IR. Fold and function of polypeptide transport-associated domains responsible for delivering unfolded proteins to membranes. Mol Microbiol. 2008;68:1216–27. doi: 10.1111/j.1365-2958.2008.06225.x. [DOI] [PubMed] [Google Scholar]
- 45.Bennion D, Charlson ES, Coon E, Misra R. Dissection of beta-barrel outer membrane protein assembly pathways through characterizing BamA POTRA 1 mutants of Escherichia coli. Mol Microbiol. 2010;77:1153–71. doi: 10.1111/j.1365-2958.2010.07280.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Wu T, McCandlish AC, Gronenberg LS, Chng S-S, Silhavy TJ, Kahne D. Identification of a protein complex that assembles lipopolysaccharide in the outer membrane of Escherichia coli. Proceedings of the National Academy of Sciences. 2006;103:11754–11759. doi: 10.1073/pnas.0604744103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hagan CL, Kim S, Kahne D. Reconstitution of Outer Membrane Protein Assembly from Purified Components. Science. 2010;328:890–892. doi: 10.1126/science.1188919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Roman-Hernandez G, Peterson JH, Bernstein HD. Reconstitution of bacterial autotransporter assembly using purified components. Elife. 2014;3:e04234. doi: 10.7554/eLife.04234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kim KH, Aulakh S, Paetzel M. The bacterial outer membrane beta-barrel assembly machinery. Protein Sci. 2012;21:751–68. doi: 10.1002/pro.2069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Webb CT, Selkrig J, Perry AJ, Noinaj N, Buchanan SK, Lithgow T. Dynamic association of BAM complex modules includes surface exposure of the lipoprotein BamC. J Mol Biol. 2012;422:545–55. doi: 10.1016/j.jmb.2012.05.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Blatch GL, Lassle M. The tetratricopeptide repeat: a structural motif mediating protein-protein interactions. Bioessays. 1999;21:932–9. doi: 10.1002/(SICI)1521-1878(199911)21:11<932::AID-BIES5>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 52.Hagan CL, Wzorek JS, Kahne D. Inhibition of the beta-barrel assembly machine by a peptide that binds BamD. Proc Natl Acad Sci U S A. 2015;112:2011–6. doi: 10.1073/pnas.1415955112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Iadanza MG, Higgins AJ, Schiffrin B, Calabrese AN, Brockwell DJ, Ashcroft AE, Radford SE, Ranson NA. Lateral opening in the intact beta-barrel assembly machinery captured by cryo-EM. Nature communications. 2016;7:12865. doi: 10.1038/ncomms12865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fleming PJ, Patel DS, Wu EL, Qi Y, Yeom MS, Sousa MC, Fleming KG, Im W. BamA POTRA Domain Interacts with a Native Lipid Membrane Surface. Biophys J. 2016;110:2698–709. doi: 10.1016/j.bpj.2016.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Maier T, Clantin B, Gruss F, Dewitte F, Delattre AS, Jacob-Dubuisson F, Hiller S, Villeret V. Conserved Omp85 lid-lock structure and substrate recognition in FhaC. Nature communications. 2015;6:7452. doi: 10.1038/ncomms8452. [DOI] [PMC free article] [PubMed] [Google Scholar]