

Abstract
Nicotine modulates neuroplasticity and improves cognitive functions in animals and humans. In the brain of smoking individuals, calcium-dependent plasticity induced by non-invasive brain stimulation methods such as transcranial direct current stimulation (tDCS) and paired associative stimulation (PAS) is impaired by nicotine withdrawal, but partially re-established after nicotine re-administration. In order to investigate the underlying mechanism further, we tested the impact of the α4β2-nicotinic receptor partial agonist varenicline on focal and non-focal plasticity in smokers during nicotine withdrawal, induced by PAS and tDCS, respectively. We administered low (0.3 mg) and high (1.0 mg) single doses of varenicline or placebo medication before stimulation over the left motor cortex of 20 healthy smokers under nicotine withdrawal. Motor cortex excitability was monitored by single-pulse transcranial magnetic stimulation-induced motor evoked potential amplitudes for 36 hours after plasticity induction. Stimulation-induced plasticity was absent under placebo medication, whereas it was present in all conditions under high dose. Low dose restituted only tDCS-induced non-focal plasticity, producing no significant impact on focal plasticity. High dose varenicline also prolonged inhibitory plasticity. These results are comparable to the impact of nicotine on withdrawal-related impaired plasticity in smokers and suggest that α4β2 nicotinic receptors are relevantly involved in plasticity deficits and restitution in smokers.
Introduction
Nicotine is the main component of tobacco responsible for the addictive properties of smoking. On the other hand, it positively impacts cognitive functions, such as working, episodic memory and attention1–3 in humans and animals. Clinical studies in patients with schizophrenia and Alzheimer’s disease have also revealed improvement of cognitive functions by nicotine4. However, other studies demonstrate negative or no effects of nicotine on cognition2, 5, which might partially be explained by baseline performance differences between participant groups6. The physiological foundation for these cognitive effects is hypothesized to be nicotinic modulation of intracellular Ca2+ concentration through α4β2 and α7 nicotinic acetylcholine receptors (nAChRs)7. These receptors are ligand-gated ion channels, involved in plasticity induction and cortical excitability modulation8, 9. In animal studies, with regard to plasticity, yet α4β2 and α7 nAChR activation yielded mixed results, producing enhancement of either LTP or LTD8, 10. Experiments in humans have shown that both, nicotine-induced non-selective activation of nAChRs, and targeted α4β2 and α7 receptor activation by respective agonists result in an increase of focal plasticity, while abolishing the effects of more diffuse plasticity induction protocols in non-smokers11, 12. In contrast, smokers under nicotine withdrawal display deficient facilitatory plasticity, probably caused by hypo-activation of desensitized nAChRs. In accordance, nicotine re-administration restitutes this impaired facilitatory plasticity13. The impact of nicotinic agents on glutamatergic plasticity is suggested to be mediated by nAChR-dependent neuronal calcium influx in non-smoking subjects. In accordance, administration of nicotine, and hereby enhancing calcium influx, reestablished LTP-like plasticity abolished by dextromethorphan, which blocks NMDA receptors with calcium channel properties14.
Plasticity in the above-mentioned studies was induced by non-invasive brain stimulation protocols, such as transcranial direct current stimulation (tDCS) and paired associative stimulation (PAS). Both techniques induce long-lasting Ca2+- and NMDA receptor-dependent shifts in cortical excitability15, 16. tDCS non-selectively affects neuronal populations under the relatively large stimulation electrodes via subthreshold resting membrane potential modulation, inducing non-focal plasticity16, 17, whereas PAS induces relatively focal and synapse-specific neuroplastic changes, affecting mainly synapses between motor and somatosensory neurons15. For tDCS, LTP-like plasticity is induced by anodal, and LTD-like plasticity by cathodal stimulation of the target area16, while for PAS synchrony of activation of somatosensory-motor cortical connections determines plasticity direction, resembling spike-timing dependent plasticity15.
Apart from disturbed facilitatory plasticity, numerous studies in humans have reported that abstention from nicotine in smokers leads to deficits in working memory and attention18, 19, which are partially restituted by re-administration of nicotine or a α4β2 nAChR agonists20–22. However, in smoking humans the role of specific nAChRs in the re-establishment of impaired plasticity has not yet been explored. The fact that the above-mentioned α4β2 nAChRs have calcium channel properties7 suggests their key role in the restitutive effect of nicotine on withdrawal-related impaired plasticity. Moreover, in non-smoking individuals, acute nicotine and varenicline (partial agonist to the α4β2 and full agonist to α7 nAChRs) administration produces fairly similar effects on stimulation-induced plasticity12, 23. Here, we aimed at exploring the effect of the α4β2 activation on impaired plasticity in smokers during nicotine abstinence. For this reason, we selected varenicline, due to its 4000–5000-fold greater affinity for the α4β2 compared to that for α7 nAChRs24. We expected that 0.3 mg and 1 mg dosages of varenicline should produce results similar to those of global nAChR activation and thus ameliorate nicotine withdrawal-related plasticity deficits in smokers.
Materials and Methods
Subjects
Twenty-six healthy smokers aged 24.5 ± 3.7 years (16 females/10 males) were recruited. All of them were students (BA, MSc and Ph.D.) of the University of Göttingen and naïve to the stimulation techniques. Six participants (4 females/2 males) from the initial group left the study after two or three experimental sessions: one participant (from the PAS experiment) cancelled the participation due to side effects and five participants (four from the tDCS, one from the PAS experiment) left the study due to their busy lesson schedules; data from these sessions were excluded from the analysis. From the remaining group of twenty participants, four (1 female/3 males) took part in both, the tDCS and PAS parts of the study, therefore a group of twelve subjects aged 25.4 ± 3.8 years (7 females/5 males) completed the tDCS and a group of twelve subjects aged 24 ± 3.4 years (6 females/6 males) completed the PAS experiment. No preliminary tests were conducted to check the responsiveness of the subjects to either tDCS or PAS. All subjects were right-handed according to the Edinburgh handedness inventory25. None of them took any medication, had a history of a neuropsychiatric or medical disease, present pregnancy, or metallic head implants. All subjects gave written informed consent and were compensated for participation. All subjects were smokers with a cigarette consumption of minimum 10 cigarettes a day for at least 5 years continuously and a Fagerström score of at least 1 point, indicating a light degree of nicotine dependence26. They were not allowed to smoke for 10 hours (3 to 4 half-lives of nicotine27) before and during the experimental sessions. The investigation was approved by the Ethics Committee of the University of Göttingen, and conforms to the principles laid down in the Declaration of Helsinki.
Transcranial Direct Current Stimulation
tDCS was administered by a battery-driven constant current stimulator (neuroConn GmbH, Ilmenau, Germany) through a pair of rubber electrodes (with the cable connector centered on the side of the rubber pad) covered with saline-soaked sponges (35 cm2, 5 × 7 cm). The motor cortex electrode was fixed over the area representing the right abductor digiti minimi muscle (ADM) and the return electrode above the contralateral supraorbital area. Subjects received 1 mA of either excitability-enhancing anodal tDCS for 13 minutes or excitability-diminishing cathodal tDCS for 9 minutes over the primary motor cortex, which induces motor cortex excitability alterations lasting for about 1 h after intervention28, 29.
Paired Associative Stimulation
The peripheral electric pulse over the right ulnar nerve at the level of the wrist at an intensity of 300% of the sensory perceptual threshold was delivered by a Digitimer D184 multipulse stimulator (Digitimer, Welwyn Garden City, United Kingdom) and was followed by a TMS pulse over the M1 representation of the abductor digiti minimi muscle (ADM) conducted by a Magstim 200 stimulator with an intensity to elicit single pulse MEPs of ~1 mV peak-to-peak amplitudes. In total, 90 paired pulses were delivered at a frequency of 0.05 Hz at ISIs of 10 ms (inhibitory PAS or PAS10) or 25 ms (facilitatory PAS or PAS25). During PAS, the participants were instructed to silently count the number of pulses to guarantee sufficient attention to the procedure, which has been shown to be crucial to obtain the desired after-effects30, 31.
Pharmacological Interventions
Low (0.3 mg) or high (1.0 mg) dosages of varenicline or 0.5 mg placebo were administered in two-piece non-transparent gelatin capsules (size 2.18 mm length, 6.35 mm external diameter) three hours before the start of the non-invasive brain stimulation protocol, allowing the verum drug to reach its maximum plasma level32. 1 mg varenicline is a usual single oral dosage administered in smokers twice per day to support cessation of cigarette consumption32–35 and both 0.3 and 1 mg doses of varenicline had a modulatory effect on both tDCS- and PAS-induced plasticity in a previous study of our group12.
Monitoring of motor cortical excitability
In order to measure excitability changes, MEPs were recorded from the right ADM by single-pulse TMS over the corresponding left primary motor cortex, conducted by a Magstim 200 magnetic stimulator (Magstim, Whiteland, Dyfed, United Kingdom) with a figure-of-eight magnetic coil (diameter of one winding −70 mm; peak magnetic field −2.2T). The coil was held tangentially to the skull, with the handle pointing posterior and laterally at 45° from the midline. The hotspot was defined as the coil placement, where the TMS pulse resulted consistently in the largest MEPs of the right ADM. Surface MEPs were recorded with Ag-AgCl electrodes in a belly-tendon montage. The signals were amplified, band-pass filtered (2 Hz to 2 kHz, sampling rate, 5 kHz) and digitized with a micro 1401 AD converter (Cambridge Electronic Design, Cambridge, UK), controlled by Signal Software (Cambridge Electronic Design, v. 2.13), and stored into a laboratory computer for offline analysis.
Experimental procedures
The participants were seated in a comfortable chair with head and arm rests. In the beginning, the motor cortex hotspot was identified by TMS and then the stimulation intensity was adjusted to elicit single pulse MEPs with peak-to-peak amplitudes of on average 1 mV. Then twenty-five MEPs were recorded for the determination of first baseline. To keep the EMG electrodes and TMS coil position constant throughout the session, their exact positions were marked with a waterproof pen. After first baseline recording, varenicline or placebo medication was administered. Three hours after intake of medication, a second baseline was recorded to monitor for a possible impact of the drug alone on cortical excitability (baseline 2), and TMS intensity was adjusted, if necessary (baseline 3). After determination of the second or third baseline, one of the plasticity induction protocols was applied (cathodal tDCS, anodal tDCS, PAS10 or PAS25) and twenty-five MEPs were recorded at time points of 0, 5, 10, 15, 20, 25, 30, 60, 90 and 120 minutes after tDCS. Further TMS measurements were conducted in the evening of the stimulation day (SE), next morning, at ~9:00 AM (NM), next noon, at ~12:00 PM (NN) and next evening, at ~6:00 PM (NE) (Fig. 1). To avoid interferences, the interval between two consecutive experimental sessions for a single subject was minimum seven days. Subjects were blinded for both, stimulation and medication conditions; the experimenter was blinded only for the medication condition.
Figure 1.
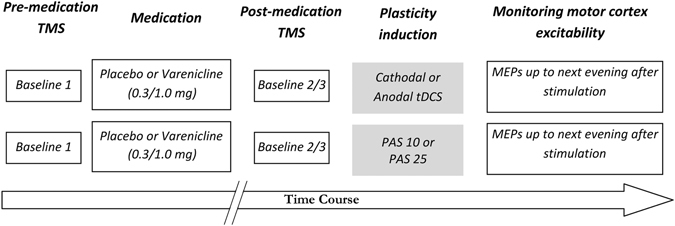
Course of the study. Participants were not allowed to smoke 10 h before and during the experimental session. In the beginning of each session, 25 baseline single pulse MEPs were recorded at an intensity to elicit MEPs with peak-to-peak amplitudes of on average ~1 mV before administration of varenicline (0.3 or 1 mg) or placebo medication. Three hours later, a second baseline was recorded to explore the effect of medication on cortical excitability, and the stimulation intensity was adjusted, if necessary (third baseline). Next, tDCS (cathodal or anodal) or PAS (PAS10 or PAS25) was administered and blocks of 25 MEPs were recorded at time points of 0, 5, 10, 15, 20, 25, 30, 60, 90 and 120 minutes after plasticity induction. Further TMS measurements were conducted in the evening of the same day (SE), morning (at ~9:00 AM, NM), noon (at ~12:00 PM, NN) and evening (at ~6:00 PM, NE) of the second day.
Analysis and statistics
The individual means of 25 MEP amplitudes were calculated at each time point for every subject. MEPs in which the muscle was not relaxed (i.e. observable EMG background activity present before the onset of the MEP) were excluded from the analysis.
Baselines
Individual baseline MEP amplitudes and maximum stimulator output (%MSO) percentage values for each session were analyzed with repeated measures analysis of variance (ANOVA) separately for tDCS and PAS experiments, using Baseline MEP or %MSO as dependent variable, respectively and session and medication (0.3 mg, 1.0 mg or placebo) as within-subject factors.
After-effects
post-stimulation mean MEP amplitudes were normalized to the respective mean baseline MEP amplitudes (quotient of post-stimulation vs pre-stimulation MEPs values: baseline 2, or, if TMS intensity was adjusted, baseline 3). Then the grand averages for each time point were calculated. A repeated measures ANOVA was performed on the above-mentioned data separately for tDCS and PAS experiments, using MEP amplitude as the dependent variable and medication, stimulation type and time course as within-subject factors.
The Mauchley’s test was used to evaluate the sphericity assumption and degrees of freedom were corrected using the Greenhouse-Geisser method when necessary. In case of significant results of the ANOVA, exploratory post hoc comparisons were performed using Student’s t-tests (paired samples, two-tailed, p < 0.05, not corrected for multiple comparisons) between the MEP amplitudes before and after intervention within one experimental condition and between the single time points (medication vs placebo) within the same stimulation condition.
Bonferroni-corrected t-tests were not used in the exploratory secondary analysis, because the experiment was powered for the primary statistical test, i.e. the ANOVA. Furthermore, since post hoc t-tests were also applied in the majority of our foregoing related studies11–14, 23, we used identical tests in order to improve inter-study comparability.
To compare main effects of different dosages of varenicline on plasticity, averaged MEPs for the first 30 minutes after stimulation were calculated for each subject per experimental session and normalized to baseline 2 (or 3, if TMS intensity was adjusted). These averaged MEP values for each dosage condition were tested via one-way ANOVA, exploratory post-hoc comparisons were conducted using Student’s t-tests (paired samples, two-tailed, p < 0.05, not corrected for multiple comparisons).
EMG measures were analyzed using custom python scripts based on the Stimfit library (version 0.14; available open source http://www.stimfit.org/doc/sphinx/)36. Statistical analysis was carried out with SPSS (version 23.0, IBM Corp.).
Results
All subjects tolerated the procedure well. Only one subject (from the PAS experiment) experienced dizziness, nausea, and vomiting under placebo medication and left the study.
On average, the interval between two consecutive experimental sessions was 11.8 ± 9.07 days.
Results of the Fagerström scores were 3.0 ± 1.8 (min 1; max 6) for the PAS and 3.5 ± 1.4 (min 1; max 6) for the tDCS group, indicating mild-to-moderate nicotine dependence37, with no significant differences as revealed by one-way-ANOVA (F(1, 22) = 0.559, p = 0.462) between stimulation groups.
Due to time constraints of participants, data were missing for the second day measurements (NM, NN and NE time points) for three subjects; in total, 8 time points in the tDCS and 3 in the PAS experiment (0.74% and 0.27% of the data, respectively). The expectation maximization (EM) imputation method was used to replace the missing values38, 39.
Effect of varenicline on motor cortex excitability
Varenicline and placebo alone did not have any impact on cortical excitability at any dosage, as revealed by the repeated measures ANOVAs conducted on the baseline data (independently on MEP amplitudes and %MSO) separately for tDCS and PAS experiments (for details see Table 1).
Table 1.
MEP amplitudes and stimulation intensity before and after varenicline administration.
| Stimulation | TMS Parameter | Medication condition | Baseline 1 | Baseline 2 | Baseline 3 |
|---|---|---|---|---|---|
| Cathodal tDCS | MEP | 0.3 mg | 0.97 ± 0.13 | 1.01 ± 0.19 | 0.93 ± 0.13 |
| 1.0 mg | 0.94 ± 0.11 | 0.81 ± 0.13 | 0.92 ± 0.06 | ||
| Placebo | 0.97 ± 0.13 | 1.06 ± 0.45 | 0.97 ± 0.12 | ||
| %MSO | 0.3 mg | 57.17 ± 14.48 | 57.08 ± 14.57 | ||
| 1.0 mg | 56.08 ± 14.15 | 56.67 ± 14.06 | |||
| Placebo | 56.50 ± 13.57 | 56.75 ± 14.07 | |||
| Anodal tDCS | MEP | 0.3 mg | 0.95 ± 0.08 | 0.95 ± 0.13 | 1.00 ± 0.11 |
| 1.0 mg | 0.99 ± 0.16 | 0.88 ± 0.15 | 0.93 ± 0.13 | ||
| Placebo | 1.00 ± 0.13 | 0.97 ± 0.15 | 0.99 ± 0.13 | ||
| %MSO | 0.3 mg | 56.83 ± 13.11 | 57.25 ± 13.07 | ||
| 1.0 mg | 57.08 ± 14.37 | 57.83 ± 14.83 | |||
| Placebo | 56.75 ± 14.00 | 57.17 ± 13.82 | |||
| PAS10 | MEP | 0.3 mg | 0.94 ± 0.10 | 1.05 ± 0.40 | 1.04 ± 0.15 |
| 1.0 mg | 0.99 ± 0.10 | 0.86 ± 0.12 | 0.90 ± 0.08 | ||
| Placebo | 1.01 ± 0.14 | 0.93 ± 0.18 | 0.97 ± 0.10 | ||
| %MSO | 0.3 mg | 57.08 ± 11.22 | 57.17 ± 11.63 | ||
| 1.0 mg | 57.75 ± 11.95 | 58.25 ± 11.93 | |||
| Placebo | 56.67 ± 12.35 | 57.08 ± 12.38 | |||
| PAS25 | MEP | 0.3 mg | 0.92 ± 0.11 | 0.91 ± 0.21 | 0.95 ± 0.12 |
| 1.0 mg | 0.97 ± 0.10 | 0.97 ± 0.19 | 0.98 ± 0.14 | ||
| Placebo | 1.01 ± 0.10 | 0.89 ± 0.15 | 0.94 ± 0.11 | ||
| %MSO | 0.3 mg | 56.83 ± 12.34 | 57.00 ± 12.45 | ||
| 1.0 mg | 57.17 ± 11.85 | 57.58 ± 11.95 | |||
| Placebo | 57.25 ± 11.42 | 57.58 ± 11.42 | |||
Shown are the mean MEP amplitudes ± S.D. and stimulation intensity (percentage of maximum stimulator output, %MSO) means ± S.D. of baselines 1, 2 and 3. The intensity of TMS was adjusted to elicit MEPs with peak-to-peak amplitude of ~1 mV (baseline 1). A second baseline (baseline 2) was recorded three hours after varenicline or placebo intake to determine the impact of the drug on cortical excitability and adjusted if necessary (baseline 3). RM-ANOVAs revealed no significant differences between %MSO values and MEP amplitudes.
Effect of varenicline on tDCS-induced plasticity
The RM-ANOVA revealed a significant MEDICATION × STIMULATION × TIME (F(28) = 1.877; p = 0.006) interaction (for details see Table 2).
Table 2.
Results of the repeated measures ANOVA.
| Experiment | Factor | Df | F | p |
|---|---|---|---|---|
| tDCS | Medication | 2 | 0.531 | 0.596 |
| Stimulation | 1 | 11.862 | 0.005* | |
| Time | 14 | 0.769 | 0.701 | |
| Medication × stimulation | 2 | 11.765 | <0.001* | |
| Medication × time | 28 | 0.518 | 0.981 | |
| Stimulation × time | 14 | 6.013 | <0.001* | |
| Medication × stimulation × time | 28 | 1.877 | 0.006* | |
| PAS | Medication | 2 | 0.486 | 0.621 |
| Stimulation | 1 | 27.095 | <0.001* | |
| Time | 14 | 0.907 | 0.553 | |
| Medication × stimulation | 2 | 6.718 | 0.005* | |
| Medication × time | 28 | 0.868 | 0.661 | |
| Stimulation × time | 14 | 2.897 | 0.001* | |
| Medication × stimulation × time | 28 | 1.136 | 0.294 |
*Significant results at p < 0.05.
Post-hoc Student’s t tests show that under placebo medication, tDCS induced no plasticity, as MEPs did not differ significantly from baseline values (the only exception being anodal tDCS at minute 60, where MEP size was significantly reduced compared to the baseline). Under both doses of varenicline, tDCS induced relevant excitability alterations. Cathodal tDCS induced a significant excitability diminution, lasting for 60 minutes after stimulation, whereas anodal tDCS induced an excitability enhancement, which was more prominent and stable under high dose varenicline (Fig. 2A,B).
Figure 2.
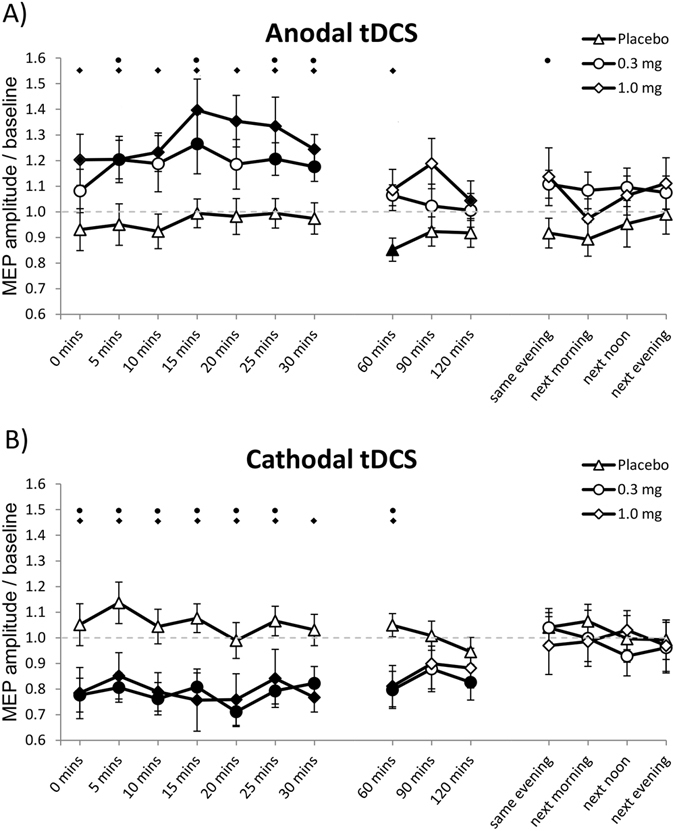
Impact of varenicline on tDCS-induced neuroplasticity. Shown are baseline-normalized MEP amplitudes after plasticity induction by anodal (A) and cathodal (B) tDCS under 0.3 mg, 1.0 mg varenicline or placebo medication conditions up to the evening of the post-stimulation day. (A) In smokers under placebo medication, anodal tDCS induced no excitability enhancement, while 0.3 mg and 1.0 mg varenicline resulted in enhanced MEP amplitudes after anodal tDCS. (B) In the placebo condition, cathodal tDCS failed to induce excitability alterations. In contrast, both low and high doses of varenicline led to significant inhibitory after-effects of tDCS. Error bars indicate S.E.M. Filled symbols indicate significant differences of post-stimulation MEP amplitudes from respective baseline values; Floating symbols indicate significant differences between the respective drug and placebo medication conditions at the same time points (Student’s t-test, two tailed, paired samples, p < 0.05).
For the effects of different dosages of varenicline on tDCS-induced plasticity, the grand average values calculated for the first 30 min after intervention were compared across groups by a one-way ANOVA and were significantly different (F(5, 66) = 11.951, p < 0.001). Both excitability-enhancing and -diminishing after-effects were present under low and high doses of varenicline compared to placebo (Student’s t test, paired samples, two-tailed, p < 0.01), under which no after-effects were observed (Fig. 3).
Figure 3.
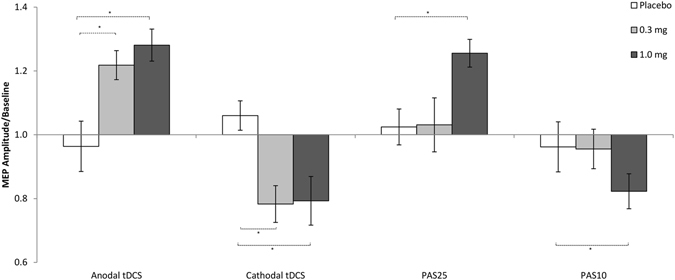
In smokers under placebo medication, tDCS- and PAS-induced plasticity is abolished. Furthermore, PAS also induced no excitability alterations under 0.3 mg varenicline. High dose varenicline restituted both tDCS and PAS-induced after-effects, while 0.3 mg varenicline only restored tDCS-induced plasticity. Each column represents the mean of baseline-normalized MEP ± S.E.M. amplitudes until 30 minutes after stimulation; Asterisks indicate significant differences between drug and placebo conditions (Student’s t-test, two tailed, paired samples, p < 0.05).
Effect of varenicline on PAS-induced plasticity
The RM-ANOVA revealed significant STIMULATION × TIME (F(14) = 2.897; p = 0.001) and MEDICATION × STIMULATION (F(2) = 6.718; p = 0.005) interactions (Table 2).
Post-hoc Student’s t - tests show that under placebo and low dose varenicline conditions, PAS failed to induce any excitability alterations. Here MEPs did not significantly differ from respective baseline values at any time point. Under high-dose varenicline, MEPs were significantly enhanced for 30 minutes after PAS25 and significantly reduced for 120 minutes after PAS10 (Fig. 4A,B).
Figure 4.
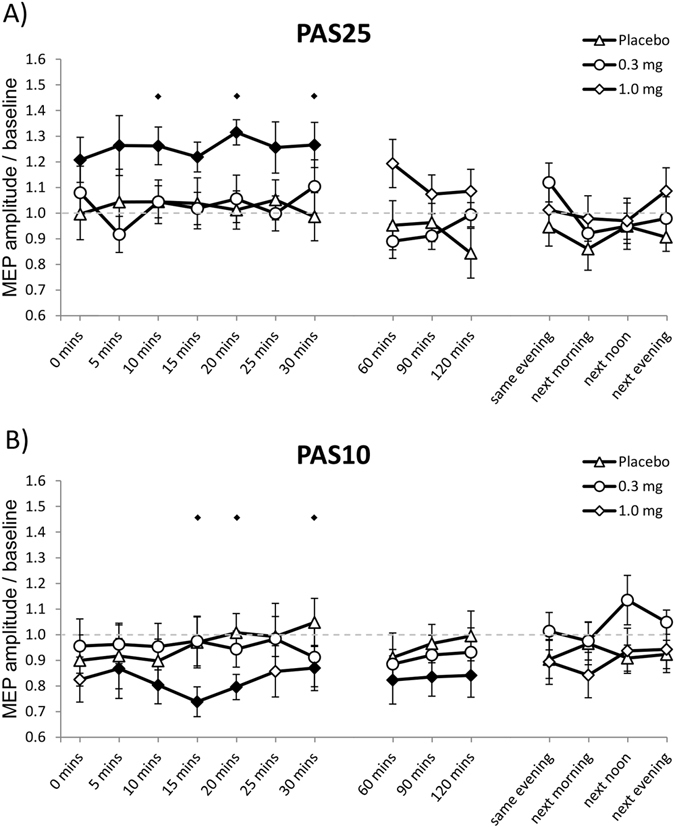
Impact of varenicline on PAS-induced neuroplasticity. Shown are baseline-normalized MEP amplitudes after plasticity induction by PAS25 (A) and PAS10 (B) under 0.3 mg, 1.0 mg varenicline or placebo medication conditions up to the evening of the post-stimulation day. Both, PAS25 and PAS10 induce no changes of MEP amplitudes in smokers under placebo or 0.3 mg varenicline conditions. (A) Cortical excitability was significantly enhanced for up to 30 minutes under high dose varenicline after PAS25 administration. (B) High dose varenicline restored inhibitory plasticity after PAS10. Error bars indicate S.E.M. Filled symbols indicate significant differences of post-stimulation MEP amplitudes from respective baseline values; floating symbols indicate significant differences between the respective drug and placebo medication conditions at the same time points (Student’s t-test, two tailed, paired samples, p < 0.05).
For the effects of different dosages of varenicline on PAS-induced plasticity, the one-way ANOVA conducted across groups on the grand average values calculated for the first 30 min after intervention was significant (F(5, 66) = 4.817, p = 0.001).
Low dose varenicline did not restitute PAS-induced plasticity, MEP sizes after both PAS10 and PAS25 were similar to those after in the placebo condition, as revealed by the respective Student’s t-tests (paired samples, two-tailed, p < 0.05). Under high dose varenicline, both PAS25 and PAS10 induced excitability enhancement and diminution respectively, compared to the placebo condition (Student’s t - test, paired samples, two-tailed, p < 0.05) (Fig. 3).
Discussion
The results of this study demonstrate that nicotine withdrawal and nAChR activation under withdrawal has a notable effect on neuroplasticity in smokers. Under nicotine withdrawal, both tDCS and PAS-induced after effects were abolished, but reinstated by high dose varenicline. High dose varencline also resulted in prolongation of tDCS-induced facilitatory and inhibitory after effects, whereas low dose medication succeeded in restitution of tDCS-induced after-effects, but was ineffective for PAS.
These results are partially similar to those acquired in a previous study, where a nicotine-withdrawal related abolishment of LTP-like facilitatory plasticity was reestablished by global nAChR activation13. Thus, we assume that the restitutive effect of nicotine on withdrawal-related impaired plasticity in smoking individuals is in large part caused by activation of α4β2 nAChRs. Moreover, similarly to another study of our group, conducted in non-smoking individuals, varenicline alone did not produce any effect on cortical excitability12.
Non-invasive brain stimulation protocols induce NMDA receptor- and Ca2+-dependent LTP- and LTD-like plasticity40, 41. Modulation of membrane penetrability to calcium ions impacts upon the induction of LTP and LTD42 and thus should be able to alter these after-effects. Nicotinic receptors that influence intracellular Ca2+ concentration are α4β2 and α7 nAChRs43. These receptors are relevant for LTP and LTD induction44, since they influence glutamatergic plasticity through intracellular calcium influx8, 9. Thus, administration of nicotine or varenicline leads to an elevated intracellular Ca2+ concentration via activation of respective subsets of nAChRs45, and affects stimulation-induced after - effects likely through this mechanism11, 12.
The lack of LTP-like plasticity in smokers can be explained by desensitization46, 47 and/or long-lasting inactivation of nAChRs under withdrawal after chronic nicotine consumption, the latter being observed even up to 5 hours after drug removal in rat brain tissue48, 49. Additionally, possibly due to the absence of the stimulatory effect of nicotine, a decrease of glutamate transmission is also observed during nicotine withdrawal50, 51. Failure of the facilitatory protocols to induce plasticity under nicotine withdrawal was also observed in a previous study of our group23, but here, both inhibitory PAS and cathodal tDCS still induced an excitability diminution. The reason for this discrepancy is unclear, as the groups are similar for age, gender distribution, Fagerstöm scores and withdrawal durations. Possible explanations include inter-group differences of the effects of chronic nicotine exposure on other neuromodulatory systems such as dopamine and serotonin52, 53, as well as Brain-derived neurotrophic factor (BDNF) levels54, which are shown to have an impact on stimulation-induced plasticity55–57. Varenicline administration probably activated α4β2 nAChRs, which are assumed to be desensitized by chronic nicotine exposure58, 59, thus facilitating intracellular calcium influx and therefore enabling either LTP or LTD-like after-effects after tDCS and PAS. Although varenicline is also a full agonist of α7 receptors, their impact on the restitutive effect of varenicline on impaired plasticity might be less relevant as (1) its affinity to α7 is 4000 to 5000 fold lower as compared to α4β2 24, (2) an α7 nAChR agonist failed to reduce nicotine withdrawal-associated cognitive deficits, as opposed to an α4β2 agonist, in another study60 and (3) in knockout mice nicotine withdrawal-related deficits in contextual fear conditioning involve β2, but not α7 subunit-containing nAChRs61.
The reason why low dosage varenicline succeeded in restitution of tDCS-induced plasticity, but failed in the PAS experiment can be explained by specific differences between these plasticity-inducing protocols. Since tDCS is assumed to induce plasticity by long-lasting tonic depolarization of relatively large neuronal populations underneath the stimulation electrodes, whereas PAS generates short depolarizations and affects only specific neuronal groups, tDCS might lead to stronger calcium increase, as compared to PAS, for a given individual. A somewhat similar effect was observed in an earlier study of our group, where in non-smoking participants both 0.3 and 1.0 mg varenicline abolished tDCS-induced after-effects, but only 1.0 mg had the same effect in the PAS condition12. These results are in agreement with the nicotine-induced calcium overflow in case of tDCS as demonstrated by our recent study14, where reduction of calcium influx blocks the conversion of anodal tDCS-induced after effects from excitability enhancement to diminution under nicotine (which is not present for PAS-induced LTP-like plasticity) and thus is compatible with larger calcium influx via tDCS, as compared to PAS.
The abolishment of plasticity in smokers under nicotine abstinence and the restorative impact of varenicline goes in line with respective results of cognitive studies in humans and animals, where α4β2 receptor-targeting pharmacologic agents ameliorated working memory and attention impairments related to withdrawal after chronic nicotine administration22, 62.
The results of this study demonstrate that withdrawal from nicotine leads to deficiency in both LTD- and LTP-like plasticity, but is re-installed after administration of a nAChR agonist which activates receptors with calcium channel properties. These results suggest a key role of α4β2 nAChRs in pathophysiological alterations under nicotine withdrawal, but also restitution of normal physiological mechanisms. Since nicotine withdrawal also negatively affects such cognitive processes as attention, learning, working memory and response inhibition21, results of this study deliver a probable physiological foundation for these cognitive deficits, which however should be explored more directly in future studies.
With regard to the clinical domain, an involvement of the cholinergic system in schizophrenia has repeatedly been suggested63, 64. Additionally, the rate of smoking and daily cigarette consumption in schizophrenia patients is significantly higher than in the general population65. This led to the assumption that nicotine consumption is a kind of self-medication to improve schizophrenia-related cognitive deficits (“the self-medication hypothesis”)66. Recent studies have demonstrated that tDCS and excitatory PAS-induced plasticity are impaired in schizophrenia patients67–69. Interestingly, excitability diminishing after-effects of cathodal tDCS are still present in smoking patients as compared to non-smokers70. Additionally, non-smoking schizophrenia patients were significantly more responsive to repeated fronto-temporal tDCS sessions, resulting in a decrease in auditory hallucinations compared to smokers71, suggesting a complex interaction between impaired plasticity and the cholinergic system. In principal accordance with these studies, demonstrating that neuroplasticity is compromised in schizophrenia, our results can at least partially explain why nicotinic agents may have positive effects in this condition72, 73. However, this explanation is hypothetical and should be systematically addressed in future studies.
This study targeted the impact of a single dose varenicline on neuroplasticity during acute (~10 h) withdrawal from nicotine. Plasticity changes related to an extended nicotine withdrawal syndrome, related to smoking cessation, which lasts several weeks74, could be significantly different. Additionally, chronic administration of varenicline might lead to discernible neuroplastic and/or cognitive changes as previously reported for nicotine75, as well as probable upregulation or desensitization of nicotinic receptors76, 77. Therefore, the effect of chronic administration of varenicline on nicotine withdrawal-related plasticity alterations should be addressed in future studies.
It should also be noted that our study explored primarily the impact of α4β2 receptors on plasticity, however, α7 nAChRs have also shown to play an important role for respective processes44, 78. For more than a decade, there has been an increased interest to develop high affinity α7 nAChR-agonists capable to cross the blood-brain barrier and induce or enhance neuroplastic changes in the brain79, 80 for treatment of schizophrenia and Alzheimer’s disease81 as well as nicotine addiction82. Thus, future studies should be designed to specifically explore the involvement of α7 receptors in nicotinic modulation of plasticity in humans.
The subjects recruited for our study were moderate smokers according to the Fagerström scale26. As was previously documented, an increase in the intensity of smoking in adults of all ages correlates with a decline in cognitive performance83–85, therefore plasticity changes in heavy smokers with stronger nicotine-dependence history may be qualitatively and quantitatively different. A systematic study involving different degrees of nicotine dependency could address this issue. Another limitation is the fact that neither blood nor breath CO tests were conducted to verify compliance of the subjects. Nevertheless, as the experimental sessions were mostly scheduled early in the morning and the participants were moderate smokers, we relied on their statements of compliance. Nicotine abstinence was also not verified after the 120th minute after intervention for the reason that the subjects were no longer present in the laboratory, thus it cannot be excluded that the late (SE-NE time points) neuroplastic effects of the stimulation were affected by nicotine consumption in some participants. Also tobacco craving (For example, Tobacco Craving Questionnaire86) and withdrawal (For example, Withdrawal Symptom Checklist87), which would have allowed to correlate physiological with behavioral data, was not assessed.
Between baseline 1 and 2 measurements the participants stayed in the local library, therefore it can be safely assumed that they were performing somewhat similar activities (e.g. reading, studying). However, in order to minimize the possible impact of the differences in these activities on the results of the experiment, an interim control would have been advantageous.
Obtaining tDCS and PAS-induced after-effect measures additionally during normal smoking behavior would have added potentially relevant data about plasticity in smokers under naturalistic conditions. This would have however added a substantial amount of experimental sessions, and we decided not to add these assessments, also in light of information we had obtained in a previous study about the impact of controlled nicotine administration in smokers13.
Furthermore, our experiment was limited to the motor cortex. Recently, an enhancement of cortical-evoked potentials after prefrontal PAS25 was demonstrated via TMS-EEG88. Since cognitive functions that are affected by nicotine abstinence, such as working memory, episodic memory and attention20 are related to the prefrontal cortex89–92 and can also be modulated by non-invasive brain stimulation protocols93, 94 as well as nicotinic agents1–3, it would be relevant and feasible to focus the future exploration of nicotine withdrawal-related plasticity shifts directly on prefrontal cortex physiology.
Finally, we did not perform cognitive tests in order to directly connect neurophysiologic results to cognition. Although the results of this study could explain the restitutive effect of varenicline on nicotine withdrawal-related cognitive deficits observed in several studies22, 62, 95, 96, this connection is still indirect and remains hypothetical. Thus, future studies should explore the direct relationship between changes in cortical excitability and cognition related to nicotine withdrawal and re-administration.
Acknowledgements
This work was supported by DFG grant NI 683/4-2. The publication of this article was supported by the Open Access Fund of the Leibniz Association.
Author Contributions
G.B. and M.N. designed the study. G.B. collected and analyzed the data. M.-F.K. and M.N. supervised the study. G.B. drafted the manuscript. All authors reviewed the manuscript.
Competing Interests
M.-F. Kuo, J. Grundey and G. Batsikadze received no financial support, and no compensation has been received from any individual or corporate entity over the past 3 years for research or professional service, and there are no personal financial holdings that could be perceived as constituting a potential conflict of interest. A. Hasan has received paid speakership by Desitin and Otsuka and BAK. He was member of a Roche and a Lundbeck Advisory Board. W. Paulus is member of the Advisory Boards of EBS Technologies and Precisis AG. M.A. Nitsche is member of the Advisory Board of Neuroelectronics.
Footnotes
M.-F. Kuo and M. A. Nitsche contributed equally to this work.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Froeliger B, Gilbert DG, McClernon FJ. Effects of nicotine on novelty detection and memory recognition performance: double-blind, placebo-controlled studies of smokers and nonsmokers. Psychopharmacology (Berl) 2009;205:625–633. doi: 10.1007/s00213-009-1571-y. [DOI] [PubMed] [Google Scholar]
- 2.Heishman SJ, Kleykamp BA, Singleton EG. Meta-analysis of the acute effects of nicotine and smoking on human performance. Psychopharmacology (Berl) 2010;210:453–469. doi: 10.1007/s00213-010-1848-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Mocking RJ, et al. Effects of short-term varenicline administration on emotional and cognitive processing in healthy, non-smoking adults: a randomized, double-blind, study. Neuropsychopharmacology. 2013;38:476–484. doi: 10.1038/npp.2012.205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sacco KA, Bannon KL, George TP. Nicotinic receptor mechanisms and cognition in normal states and neuropsychiatric disorders. J Psychopharmacol. 2004;18:457–474. doi: 10.1177/0269881104047273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sherwood N. Effects of Nicotine on Human Psychomotor Performance. Hum Psychopharm Clin. 1993;8:155–184. doi: 10.1002/hup.470080303. [DOI] [Google Scholar]
- 6.Niemegeers P, et al. The effects of nicotine on cognition are dependent on baseline performance. Eur Neuropsychopharmacol. 2014;24:1015–1023. doi: 10.1016/j.euroneuro.2014.03.011. [DOI] [PubMed] [Google Scholar]
- 7.Burnashev N. Calcium permeability of ligand-gated channels. Cell Calcium. 1998;24:325–332. doi: 10.1016/S0143-4160(98)90056-2. [DOI] [PubMed] [Google Scholar]
- 8.Placzek AN, Zhang TA, Dani JA. Nicotinic mechanisms influencing synaptic plasticity in the hippocampus. Acta Pharmacol Sin. 2009;30:752–760. doi: 10.1038/aps.2009.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Dani JA, Bertrand D. Nicotinic acetylcholine receptors and nicotinic cholinergic mechanisms of the central nervous system. Annu Rev Pharmacol Toxicol. 2007;47:699–729. doi: 10.1146/annurev.pharmtox.47.120505.105214. [DOI] [PubMed] [Google Scholar]
- 10.Yakel JL. Nicotinic ACh receptors in the hippocampus: role in excitability and plasticity. Nicotine Tob Res. 2012;14:1249–1257. doi: 10.1093/ntr/nts091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Thirugnanasambandam N, et al. Nicotinergic impact on focal and non-focal neuroplasticity induced by non-invasive brain stimulation in non-smoking humans. Neuropsychopharmacology. 2011;36:879–886. doi: 10.1038/npp.2010.227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Batsikadze G, Paulus W, Grundey J, Kuo MF, Nitsche MA. Effect of the Nicotinic alpha4beta2-receptor Partial Agonist Varenicline on Non-invasive Brain Stimulation-Induced Neuroplasticity in the Human Motor Cortex. Cereb Cortex. 2015;25:3249–3259. doi: 10.1093/cercor/bhu126. [DOI] [PubMed] [Google Scholar]
- 13.Grundey J, et al. Neuroplasticity in cigarette smokers is altered under withdrawal and partially restituted by nicotine exposition. J Neurosci. 2012;32:4156–4162. doi: 10.1523/JNEUROSCI.3660-11.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lugon MD, et al. Mechanisms of Nicotinic Modulation of Glutamatergic Neuroplasticity in Humans. Cereb Cortex. 2015 doi: 10.1093/cercor/bhv252. [DOI] [PubMed] [Google Scholar]
- 15.Carson RG, Kennedy NC. Modulation of human corticospinal excitability by paired associative stimulation. Front Hum Neurosci. 2013;7:823. doi: 10.3389/fnhum.2013.00823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nitsche MA, et al. Transcranial direct current stimulation: State of the art 2008. Brain stimulation. 2008;1:206–223. doi: 10.1016/j.brs.2008.06.004. [DOI] [PubMed] [Google Scholar]
- 17.Purpura DP, McMurtry JG. Intracellular Activities and Evoked Potential Changes during Polarization of Motor Cortex. J Neurophysiol. 1965;28:166–185. doi: 10.1152/jn.1965.28.1.166. [DOI] [PubMed] [Google Scholar]
- 18.Jacobsen LK, et al. Effects of smoking and smoking abstinence on cognition in adolescent tobacco smokers. Biol Psychiatry. 2005;57:56–66. doi: 10.1016/j.biopsych.2004.10.022. [DOI] [PubMed] [Google Scholar]
- 19.Wesnes KA, Edgar CJ, Kezic I, Salih HM, de Boer P. Effects of nicotine withdrawal on cognition in a clinical trial setting. Psychopharmacology (Berl) 2013;229:133–140. doi: 10.1007/s00213-013-3089-6. [DOI] [PubMed] [Google Scholar]
- 20.Grundey J, et al. Double dissociation of working memory and attentional processes in smokers and non-smokers with and without nicotine. Psychopharmacology (Berl) 2015;232:2491–2501. doi: 10.1007/s00213-015-3880-7. [DOI] [PubMed] [Google Scholar]
- 21.Ashare, R. L., Falcone, M. & Lerman, C. Cognitive function during nicotine withdrawal: Implications for nicotine dependence treatment. Neuropharmacology76 Pt B, 581–591, 10.1016/j.neuropharm.2013.04.034 (2014). [DOI] [PMC free article] [PubMed]
- 22.Loughead J, et al. Effects of the alpha4beta2 partial agonist varenicline on brain activity and working memory in abstinent smokers. Biol Psychiatry. 2010;67:715–721. doi: 10.1016/j.biopsych.2010.01.016. [DOI] [PubMed] [Google Scholar]
- 23.Grundey J, et al. Rapid effect of nicotine intake on neuroplasticity in non-smoking humans. Front Pharmacol. 2012;3:186. doi: 10.3389/fphar.2012.00186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mihalak KB, Carroll FI, Luetje CW. Varenicline is a partial agonist at alpha4beta2 and a full agonist at alpha7 neuronal nicotinic receptors. Mol Pharmacol. 2006;70:801–805. doi: 10.1124/mol.106.025130. [DOI] [PubMed] [Google Scholar]
- 25.Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- 26.Heatherton TF, Kozlowski LT, Frecker RC, Fagerstrom KO. The Fagerstrom Test for Nicotine Dependence: a revision of the Fagerstrom Tolerance Questionnaire. Br J Addict. 1991;86:1119–1127. doi: 10.1111/j.1360-0443.1991.tb01879.x. [DOI] [PubMed] [Google Scholar]
- 27.Benowitz NL, Jacob P, 3rd, Jones RT, Rosenberg J. Interindividual variability in the metabolism and cardiovascular effects of nicotine in man. J Pharmacol Exp Ther. 1982;221:368–372. [PubMed] [Google Scholar]
- 28.Nitsche, M. A. et al. Level of action of cathodal DC polarisation induced inhibition of the human motor cortex. Clin Neurophysiol114, 600–604, doi:S1388245702004121 (2003). [DOI] [PubMed]
- 29.Nitsche MA, Paulus W. Sustained excitability elevations induced by transcranial DC motor cortex stimulation in humans. Neurology. 2001;57:1899–1901. doi: 10.1212/WNL.57.10.1899. [DOI] [PubMed] [Google Scholar]
- 30.Stefan K, Kunesch E, Cohen LG, Benecke R, Classen J. Induction of plasticity in the human motor cortex by paired associative stimulation. Brain. 2000;123(Pt 3):572–584. doi: 10.1093/brain/123.3.572. [DOI] [PubMed] [Google Scholar]
- 31.Stefan K, Wycislo M, Classen J. Modulation of associative human motor cortical plasticity by attention. J Neurophysiol. 2004;92:66–72. doi: 10.1152/jn.00383.2003. [DOI] [PubMed] [Google Scholar]
- 32.Mohanasundaram UM, Chitkara R, Krishna G. Smoking cessation therapy with varenicline. Int J Chron Obstruct Pulmon Dis. 2008;3:239–251. doi: 10.2147/COPD.S1848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Obach RS, et al. Metabolism and disposition of varenicline, a selective alpha4beta2 acetylcholine receptor partial agonist, in vivo and in vitro. Drug Metab Dispos. 2006;34:121–130. doi: 10.1124/dmd.105.006767. [DOI] [PubMed] [Google Scholar]
- 34.Faessel HM, et al. A review of the clinical pharmacokinetics and pharmacodynamics of varenicline for smoking cessation. Clin Pharmacokinet. 2010;49:799–816. doi: 10.2165/11537850-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 35.Faessel HM, et al. Single-dose pharmacokinetics of varenicline, a selective nicotinic receptor partial agonist, in healthy smokers and nonsmokers. J Clin Pharmacol. 2006;46:991–998. doi: 10.1177/0091270006290669. [DOI] [PubMed] [Google Scholar]
- 36.Guzman SJ, Schlogl A, Schmidt-Hieber C. Stimfit: quantifying electrophysiological data with Python. Front Neuroinform. 2014;8:16. doi: 10.3389/fninf.2014.00016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Fagerstrom K, Russ C, Yu CR, Yunis C, Foulds J. The Fagerstrom Test for Nicotine Dependence as a predictor of smoking abstinence: a pooled analysis of varenicline clinical trial data. Nicotine Tob Res. 2012;14:1467–1473. doi: 10.1093/ntr/nts018. [DOI] [PubMed] [Google Scholar]
- 38.Musil CM, Warner CB, Yobas PK, Jones SL. A comparison of imputation techniques for handling missing data. West J Nurs Res. 2002;24:815–829. doi: 10.1177/019394502762477004. [DOI] [PubMed] [Google Scholar]
- 39.Schafer JL, Olsen MK. Multiple Imputation for Multivariate Missing-Data Problems: A Data Analyst’s Perspective. Multivariate Behav Res. 1998;33:545–571. doi: 10.1207/s15327906mbr3304_5. [DOI] [PubMed] [Google Scholar]
- 40.Nitsche MA, et al. Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J Physiol. 2003;553:293–301. doi: 10.1113/jphysiol.2003.049916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wolters A, et al. A temporally asymmetric Hebbian rule governing plasticity in the human motor cortex. J Neurophysiol. 2003;89:2339–2345. doi: 10.1152/jn.00900.2002. [DOI] [PubMed] [Google Scholar]
- 42.Lisman JE. Three Ca2+ levels affect plasticity differently: the LTP zone, the LTD zone and no man’s land. J Physiol. 2001;532:285–285. doi: 10.1111/j.1469-7793.2001.0285f.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Dajas-Bailador F, Wonnacott S. Nicotinic acetylcholine receptors and the regulation of neuronal signalling. Trends Pharmacol Sci. 2004;25:317–324. doi: 10.1016/j.tips.2004.04.006. [DOI] [PubMed] [Google Scholar]
- 44.Matsuyama S, Matsumoto A. Epibatidine induces long-term potentiation (LTP) via activation of alpha4beta2 nicotinic acetylcholine receptors (nAChRs) in vivo in the intact mouse dentate gyrus: both alpha7 and alpha4beta2 nAChRs essential to nicotinic LTP. J Pharmacol Sci. 2003;93:180–187. doi: 10.1254/jphs.93.180. [DOI] [PubMed] [Google Scholar]
- 45.He J, Deng CY, Chen RZ, Zhu XN, Yu JP. Long-term potentiation induced by nicotine in CA1 region of hippocampal slice is Ca(2+)-dependent. Acta Pharmacol Sin. 2000;21:429–432. [PubMed] [Google Scholar]
- 46.Picciotto MR, Addy NA, Mineur YS, Brunzell DH. It is not “either/or”: activation and desensitization of nicotinic acetylcholine receptors both contribute to behaviors related to nicotine addiction and mood. Prog Neurobiol. 2008;84:329–342. doi: 10.1016/j.pneurobio.2007.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wang H, Sun X. Desensitized nicotinic receptors in brain. Brain Res Brain Res Rev. 2005;48:420–437. doi: 10.1016/j.brainresrev.2004.09.003. [DOI] [PubMed] [Google Scholar]
- 48.Rowell PP, Duggan DS. Long-lasting inactivation of nicotinic receptor function in vitro by treatment with high concentrations of nicotine. Neuropharmacology. 1998;37:103–111. doi: 10.1016/S0028-3908(97)00193-7. [DOI] [PubMed] [Google Scholar]
- 49.Lukas RJ. Effects of chronic nicotinic ligand exposure on functional activity of nicotinic acetylcholine receptors expressed by cells of the PC12 rat pheochromocytoma or the TE671/RD human clonal line. J Neurochem. 1991;56:1134–1145. doi: 10.1111/j.1471-4159.1991.tb11403.x. [DOI] [PubMed] [Google Scholar]
- 50.D’Souza, M. S. & Markou, A. The “stop” and “go” of nicotine dependence: role of GABA and glutamate. Cold Spring Harb Perspect Med3, doi:10.1101/cshperspect.a012146 (2013). [DOI] [PMC free article] [PubMed]
- 51.Kenny PJ, Gasparini F, Markou A. Group II metabotropic and alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate (AMPA)/kainate glutamate receptors regulate the deficit in brain reward function associated with nicotine withdrawal in rats. J Pharmacol Exp Ther. 2003;306:1068–1076. doi: 10.1124/jpet.103.052027. [DOI] [PubMed] [Google Scholar]
- 52.Tan H, Bishop SF, Lauzon NM, Sun N, Laviolette SR. Chronic nicotine exposure switches the functional role of mesolimbic dopamine transmission in the processing of nicotine’s rewarding and aversive effects. Neuropharmacology. 2009;56:741–751. doi: 10.1016/j.neuropharm.2008.12.008. [DOI] [PubMed] [Google Scholar]
- 53.Semba J, Wakuta M. Chronic effect of nicotine on serotonin transporter mRNA in the raphe nucleus of rats: reversal by co-administration of bupropion. Psychiatry and clinical neurosciences. 2008;62:435–441. doi: 10.1111/j.1440-1819.2008.01822.x. [DOI] [PubMed] [Google Scholar]
- 54.Kivinummi T, Kaste K, Rantamaki T, Castren E, Ahtee L. Alterations in BDNF and phospho-CREB levels following chronic oral nicotine treatment and its withdrawal in dopaminergic brain areas of mice. Neuroscience letters. 2011;491:108–112. doi: 10.1016/j.neulet.2011.01.015. [DOI] [PubMed] [Google Scholar]
- 55.Kuo HI, et al. Chronic Enhancement of Serotonin Facilitates Excitatory Transcranial Direct Current Stimulation-Induced Neuroplasticity. Neuropsychopharmacology. 2016;41:1223–1230. doi: 10.1038/npp.2015.270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kuo MF, Paulus W, Nitsche MA. Boosting focally-induced brain plasticity by dopamine. Cereb Cortex. 2008;18:648–651. doi: 10.1093/cercor/bhm098. [DOI] [PubMed] [Google Scholar]
- 57.Podda MV, et al. Anodal transcranial direct current stimulation boosts synaptic plasticity and memory in mice via epigenetic regulation of Bdnf expression. Sci Rep. 2016;6:22180. doi: 10.1038/srep22180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Benowitz NL. Nicotine Addiction. New Engl J Med. 2010;362:2295–2303. doi: 10.1056/NEJMra0809890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Dani JA, Harris RA. Nicotine addiction and comorbidity with alcohol abuse and mental illness. Nat Neurosci. 2005;8:1465–1470. doi: 10.1038/nn1580. [DOI] [PubMed] [Google Scholar]
- 60.Yildirim E, Connor DA, Gould TJ. ABT-089, but not ABT-107, ameliorates nicotine withdrawal-induced cognitive deficits in C57BL6/J mice. Behav Pharmacol. 2015;26:241–248. doi: 10.1097/FBP.0000000000000111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Portugal GS, Kenney JW, Gould TJ. Beta2 subunit containing acetylcholine receptors mediate nicotine withdrawal deficits in the acquisition of contextual fear conditioning. Neurobiol Learn Mem. 2008;89:106–113. doi: 10.1016/j.nlm.2007.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Raybuck JD, Portugal GS, Lerman C, Gould TJ. Varenicline ameliorates nicotine withdrawal-induced learning deficits in C57BL/6 mice. Behav Neurosci. 2008;122:1166–1171. doi: 10.1037/a0012601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Adams CE, Stevens KE. Evidence for a role of nicotinic acetylcholine receptors in schizophrenia. Front Biosci. 2007;12:4755–4772. doi: 10.2741/2424. [DOI] [PubMed] [Google Scholar]
- 64.Leonard S, et al. Nicotinic receptors, smoking and schizophrenia. Restor Neurol Neurosci. 1998;12:195–201. [PubMed] [Google Scholar]
- 65.de Leon J, Diaz FJ. A meta-analysis of worldwide studies demonstrates an association between schizophrenia and tobacco smoking behaviors. Schizophr Res. 2005;76:135–157. doi: 10.1016/j.schres.2005.02.010. [DOI] [PubMed] [Google Scholar]
- 66.Kumari V, Postma P. Nicotine use in schizophrenia: The self medication hypotheses. Neurosci Biobehav R. 2005;29:1021-+. doi: 10.1016/j.neubiorev.2005.02.006. [DOI] [PubMed] [Google Scholar]
- 67.Hasan A, et al. Impaired long-term depression in schizophrenia: a cathodal tDCS pilot study. Brain stimulation. 2012;5:475–483. doi: 10.1016/j.brs.2011.08.004. [DOI] [PubMed] [Google Scholar]
- 68.Frantseva MV, et al. Evidence for impaired long-term potentiation in schizophrenia and its relationship to motor skill learning. Cereb Cortex. 2008;18:990–996. doi: 10.1093/cercor/bhm151. [DOI] [PubMed] [Google Scholar]
- 69.Hasan A, et al. Dysfunctional long-term potentiation-like plasticity in schizophrenia revealed by transcranial direct current stimulation. Behav Brain Res. 2011;224:15–22. doi: 10.1016/j.bbr.2011.05.017. [DOI] [PubMed] [Google Scholar]
- 70.Strube W, et al. Smoking restores impaired LTD-like plasticity in schizophrenia: a transcranial direct current stimulation study. Neuropsychopharmacology. 2015;40:822–830. doi: 10.1038/npp.2014.275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Brunelin J, Hasan A, Haesebaert F, Nitsche MA, Poulet E. Nicotine Smoking Prevents the Effects of Frontotemporal Transcranial Direct Current Stimulation (tDCS) in Hallucinating Patients With Schizophrenia. Brain stimulation. 2015;8:1225–1227. doi: 10.1016/j.brs.2015.08.002. [DOI] [PubMed] [Google Scholar]
- 72.Hong LE, et al. Effects of moderate-dose treatment with varenicline on neurobiological and cognitive biomarkers in smokers and nonsmokers with schizophrenia or schizoaffective disorder. Arch Gen Psychiatry. 2011;68:1195–1206. doi: 10.1001/archgenpsychiatry.2011.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Shim JC, et al. Adjunctive varenicline treatment with antipsychotic medications for cognitive impairments in people with schizophrenia: a randomized double-blind placebo-controlled trial. Neuropsychopharmacology. 2012;37:660–668. doi: 10.1038/npp.2011.238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Hughes JR, Higgins ST, Bickel WK. Nicotine withdrawal versus other drug withdrawal syndromes: similarities and dissimilarities. Addiction. 1994;89:1461–1470. doi: 10.1111/j.1360-0443.1994.tb03744.x. [DOI] [PubMed] [Google Scholar]
- 75.Levin ED, McClernon FJ, Rezvani AH. Nicotinic effects on cognitive function: behavioral characterization, pharmacological specification, and anatomic localization. Psychopharmacology (Berl) 2006;184:523–539. doi: 10.1007/s00213-005-0164-7. [DOI] [PubMed] [Google Scholar]
- 76.Buisson B, Bertrand D. Chronic exposure to nicotine upregulates the human (alpha)4((beta)2 nicotinic acetylcholine receptor function. J Neurosci. 2001;21:1819–1829. doi: 10.1523/JNEUROSCI.21-06-01819.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Fenster CP, Rains MF, Noerager B, Quick MW, Lester RA. Influence of subunit composition on desensitization of neuronal acetylcholine receptors at low concentrations of nicotine. J Neurosci. 1997;17:5747–5759. doi: 10.1523/JNEUROSCI.17-15-05747.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hunter BE, de Fiebre CM, Papke RL, Kem WR, Meyer EM. A novel nicotinic agonist facilitates induction of long-term potentiation in the rat hippocampus. Neuroscience letters. 1994;168:130–134. doi: 10.1016/0304-3940(94)90433-2. [DOI] [PubMed] [Google Scholar]
- 79.Ghiron C, et al. Novel alpha-7 nicotinic acetylcholine receptor agonists containing a urea moiety: identification and characterization of the potent, selective, and orally efficacious agonist 1-[6-(4-fluorophenyl)pyridin-3-yl]-3-(4-piperidin-1-ylbutyl) urea (SEN34625/WYE-103914) J Med Chem. 2010;53:4379–4389. doi: 10.1021/jm901692q. [DOI] [PubMed] [Google Scholar]
- 80.Roncarati R, et al. Procognitive and neuroprotective activity of a novel alpha7 nicotinic acetylcholine receptor agonist for treatment of neurodegenerative and cognitive disorders. J Pharmacol Exp Ther. 2009;329:459–468. doi: 10.1124/jpet.108.150094. [DOI] [PubMed] [Google Scholar]
- 81.Pohanka M. Alpha7 Nicotinic Acetylcholine Receptor Is a Target in Pharmacology and Toxicology. Int J Mol Sci. 2012;13:2219–2238. doi: 10.3390/ijms13022219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Brunzell DH, McIntosh JM. Alpha7 Nicotinic Acetylcholine Receptors Modulate Motivation to Self-Administer Nicotine: Implications for Smoking and Schizophrenia. Neuropsychopharmacology. 2012;37:1134–1143. doi: 10.1038/npp.2011.299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Vajravelu HR, Gnanadurai TK, Krishnan P, Ayyavoo S. Impact of Quantified Smoking Status on Cognition in Young Adults. J Clin Diagn Res. 2015;9:CC01–03. doi: 10.7860/JCDR/2015/16444.6867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Richards M, Jarvis MJ, Thompson N, Wadsworth ME. Cigarette smoking and cognitive decline in midlife: evidence from a prospective birth cohort study. Am J Public Health. 2003;93:994–998. doi: 10.2105/AJPH.93.6.994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kalmijn S, van Boxtel MP, Verschuren MW, Jolles J, Launer LJ. Cigarette smoking and alcohol consumption in relation to cognitive performance in middle age. Am J Epidemiol. 2002;156:936–944. doi: 10.1093/aje/kwf135. [DOI] [PubMed] [Google Scholar]
- 86.Heishman SJ, Singleton EG, Moolchan ET. Tobacco Craving Questionnaire: reliability and validity of a new multifactorial instrument. Nicotine Tob Res. 2003;5:645–654. doi: 10.1080/1462220031000158681. [DOI] [PubMed] [Google Scholar]
- 87.Hughes JR, Hatsukami D. Signs and symptoms of tobacco withdrawal. Arch Gen Psychiatry. 1986;43:289–294. doi: 10.1001/archpsyc.1986.01800030107013. [DOI] [PubMed] [Google Scholar]
- 88.Rajji TK, et al. PAS-induced potentiation of cortical-evoked activity in the dorsolateral prefrontal cortex. Neuropsychopharmacology. 2013;38:2545–2552. doi: 10.1038/npp.2013.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Lara AH, Wallis JD. The Role of Prefrontal Cortex in Working Memory: A Mini Review. Front Syst Neurosci. 2015;9:173. doi: 10.3389/fnsys.2015.00173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Rossi AF, Pessoa L, Desimone R, Ungerleider LG. The prefrontal cortex and the executive control of attention. Experimental brain research. 2009;192:489–497. doi: 10.1007/s00221-008-1642-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Fletcher PC, Shallice T, Dolan RJ. The functional roles of prefrontal cortex in episodic memory. I. Encoding. Brain. 1998;121(Pt 7):1239–1248. doi: 10.1093/brain/121.7.1239. [DOI] [PubMed] [Google Scholar]
- 92.Fletcher PC, Shallice T, Frith CD, Frackowiak RS, Dolan RJ. The functional roles of prefrontal cortex in episodic memory. II. Retrieval. Brain. 1998;121(Pt 7):1249–1256. doi: 10.1093/brain/121.7.1249. [DOI] [PubMed] [Google Scholar]
- 93.Roy LB, Sparing R, Fink GR, Hesse MD. Modulation of attention functions by anodal tDCS on right PPC. Neuropsychologia. 2015;74:96–107. doi: 10.1016/j.neuropsychologia.2015.02.028. [DOI] [PubMed] [Google Scholar]
- 94.Au J, et al. Enhancing Working Memory Training with Transcranial Direct Current Stimulation. Journal of cognitive neuroscience. 2016;28:1419–1432. doi: 10.1162/jocn_a_00979. [DOI] [PubMed] [Google Scholar]
- 95.Ashare RL, McKee SA. Effects of varenicline and bupropion on cognitive processes among nicotine-deprived smokers. Exp Clin Psychopharmacol. 2012;20:63–70. doi: 10.1037/a0025594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Patterson F, et al. Varenicline improves mood and cognition during smoking abstinence. Biol Psychiatry. 2009;65:144–149. doi: 10.1016/j.biopsych.2008.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]