

Abstract
Endometrial cancer is the most common gynaecological malignancy encountered in developed countries and the second most common in the developing world. The five-year survival rate of patients with endometrial cancer diagnosed at a late stage is <30%. Therefore, it is critical to develop a suitable chemotherapeutic regimen for late-stage endometrial cancer. Cisplatin (CDDP) is a first-line chemotherapeutic drug for endometrial cancer chemotherapy. The present study investigated the molecular mechanism underlying the effect of CDDP on endometrial cancer from the perspective of cell autophagy. Ishikawa cells were treated with 10, 20, 40 or 80 µg/ml CDDP for 12, 24, 48 and 72 h. The cells were then harvested and subjected to cell proliferation assays. Based on the results, 20 µg/ml CDDP was selected as the treatment used for 12 and 24 h for the assays. To detect the effect of CDDP on Ishikawa cell autophagy, autophagosome formation was observed using a transmission electron microscope, and the expression level of autophagy-related gene microtubule-associated protein 1 light chain 3α, was examined using immunofluorescence microscopy. The results demonstrated that CDDP treatment promoted cell autophagy in Ishikawa cells. In addition, the total and phosphorylated protein levels of phosphoinositide 3-kinase (PI3K) p85, protein kinase B (AKT) and mammalian target of rapamycin (mTOR), the key proteins of the PI3K/AKT/mTOR signalling pathway, were detected by western blot analysis. The results indicated that CDDP treatment inactivated the PI3K/AKT/mTOR signalling pathway. To further examine whether CDDP affects cell autophagy in Ishikawa cells via the PI3K/AKT/mTOR signalling pathway, the cells were co-treated with a PI3K activator, insulin-like growth factor-1 (IGF-1). The results demonstrated that IGF-1 co-treatment reversed the effect of CDDP on cell autophagy in Ishikawa cells. In brief, the present study hypothesized that CDDP may regulate cell autophagy in the Ishikawa endometrial cancer cell line via the PI3K/AKT/mTOR signalling pathway.
Keywords: endometrial cancer, autophagy, PI3K/AKT/mTOR, cisplatin
Introduction
Endometrial cancer is a cancer of the endometrium (the lining of the uterus), and is a common gynaecologic malignancy encountered in developed countries and the second most common in the developing world (1). The leading treatment method for endometrial cancer is abdominal hysterectomy (total surgical removal of the uterus), along with removal of the fallopian tubes and ovaries on both sides, which is known as a bilateral salpingo-oophorectomy (2). If the disease is diagnosed at an early stage, the outcome is favourable, and the overall five-year survival rate in the United States is >80% (3). However, the majority of cases of endometrial cancer are diagnosed at a later stage and the outcome is poor, with a five-year survival rate of <30% (3). Therefore, it is critical to develop suitable chemotherapeutic regimens for late-stage endometrial cancer. It is also important to demonstrate the molecular mechanism underlying the effect of chemotherapeutic drugs on endometrial cancer treatment.
Autophagy is a dynamic process involving the rearrangement of subcellular membranes to sequester the cytoplasm and organelles for delivery to the lysosome or vacuole, where the sequestered cargo is degraded and recycled (4). Autophagy has been implicated in the pathogenesis of cancer, and is commonly referred to as a ‘double-edged sword’ for its role in tumour progression and tumour suppression (5). Previous studies have revealed that chemotherapy drugs may regulate autophagy in the cells of a number of cancer subtypes (6–8). Cisplatin (CDDP) is the first-line chemotherapeutic drug for endometrial cancer chemotherapy (9). CDDP-induced autophagy has been reported in numerous types of cancer cells, including hepatocellular carcinoma (10), laryngeal cancer (11) and lung adenocarcinoma (12), though the association between CDDP and autophagy varied between types of cancer cell. Treatment with CDDP was demonstrated to activate autophagy in hepatocellular carcinoma and lung adenocarcinoma; however, it suppressed autophagy in laryngeal cancer (10–12). The present study investigated the association between CDDP and autophagy in endometrial cancer.
The activation of phosphoinositide 3-kinase (PI3K) or mammalian target of rapamycin (mTOR) are two of the most common events in the development of human cancer, including endometrial cancer (5,13). Alteration of the PI3K/protein kinase B (AKT)/mTOR signalling pathway is implicated in endometrial cancer pathogenesis (14). Previous studies have demonstrated that the PI3K/AKT/mTOR signalling pathway may repress autophagy in response to insulin-like and other growth factor signals (15,16). However, it is not clear whether CDDP regulates cell autophagy in endometrial cancer cells by the PI3K/AKT/mTOR signalling pathway. In the present study, it was considered whether CDDP regulated cell autophagy in the endometrial cancer cell line Ishikawa via the PI3K/AKT/mTOR signalling pathway, a hypothesis that was confirmed by the results.
Materials and methods
Materials and reagents
The Ishikawa human endometrial cancer cell line was purchased from the American Type Culture Collection (ATCC; Manassas, VA, USA). RPMI-1640 medium, fetal bovine serum (FBS), PBS, sodium pyruvate, trypsin and antibiotics were purchased from HyClone (GE Healthcare Life Sciences, Logan, UT, USA). CellTiter 96® AQueous One Solution Cell Proliferation Assay kit was purchased from Promega Corp. (Madison, WI, USA). Polyvinylidene fluoride (PVDF) membrane was purchased from EMD Millipore (Billerica, MA, USA). Molecular weight markers were purchased from Fermentas (Thermo Fisher Scientific, Inc., Pittsburgh, PA, USA). Bicinchoninic acid (BCA) protein assay kit and radioimmunoprecipitation assay (RIPA) buffer were obtained from Beyotime Institute of Biotechnology (Shanghai, China). Freeze-dried CDDP powder was purchased from Qilu Pharmaceutical Co., Ltd. (Jinan, China). Mouse monoclonal anti-PI3K p85 antibody (ab40776), mouse monoclonal anti-AKT1 antibody (ab89402), rabbit polyclonal anti-AKT1 (phospho-S473) antibody (ab66138), rabbit monoclonal anti-mTOR antibody (ab32028), rabbit monoclonal anti-mTOR (phospho-S2448) antibody (ab109268), rabbit polyclonal anti-microtubule-associated protein 1 light chain 3α (LC3) B antibody (ab63817) and rabbit polyclonal anti-GAPDH antibody (ab9485) were all purchased from Abcam (Cambridge, UK). Enhanced chemiluminescence (ECL) reagent was obtained from EMD Millipore. Alexa Fluor 488-conjugated donkey anti-rabbit immunoglobulin (Ig)G secondary antibodies were provided by Santa Cruz Biotechnology, Inc. (Dallas, TX, USA) (sc-3895). Horseradish peroxidase-conjugated goat anti-rabbit IgG was purchased from Boster Biological Technology (BA1055; Wuhan, China). Insulin-like growth factor-1 (IGF-1) was purchased from Sino Biological, Inc. (Beijing, China).
Cell culture and CDDP treatment
Ishikawa cells were maintained on culture dishes in 90% (v/v) RPMI-1640 medium supplemented with 2 mM L-glutamine and 1.5 g/l sodium bicarbonate with 10% (v/v) FBS. The cells were cultured in an atmosphere containing 5% CO2 at 37°C in an incubator. A stock solution of CDDP was prepared in dimethyl sulfoxide (DMSO) at 1 mg/ml and was further diluted to the final working concentrations (10, 20, 40 and 80 µg/ml) with antibiotic-free RPMI-1640 medium.
Cell proliferation assays
Cell proliferation was evaluated using the CellTiter 96® AQueous One Solution Cell Proliferation Assay kit, according to the manufacturer's protocol. Ishikawa cells (1×104 cells) were seeded onto 96-well plates. Following 4 h of incubation, 10 µl CDDP in 1% DMSO was added to the wells at 10, 20, 40 and 80 µg/ml, whereas 10 µl 1% DMSO was used as the negative control. Briefly, following CDDP treatment for 0, 6, 12, 24 and 48 h, 20 µl CellTiter 96® AQueous One Solution reagents were added to each well and incubated for 4 h at 37°C. The absorbance was determined at 490 nm using a Multiskan MK3 microplate reader (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The experiments were repeated three times, and the rate of cell proliferation inhibition was evaluated using the following formula: Inhibition rate (%) = 1 - (mean absorbance of treated cells/mean absorbance of the negative control) × 100.
Transmission electron microscopy
The autophagy of Ishikawa cells was evaluated by autophagosome screening under a JEM-1010 transmission electron microscope (Matsunaga Manufacturing, Co., Ltd., Gifu, Japan). At the end of the intervention, the cells were digested with 0.25% trypsin and collected in centrifuge tubes, followed by centrifugation at 100 × g for 10 min, then 80 × g for 10 min, at 4°C. The supernatant was discarded, and 2.5% glutaraldehyde was added to the tube to fix the cells. Following a 2-h period of fixation, dehydration, and embedding, ultra-thin sections (70 nm) were prepared using a microtome and each section was mounted on a copper grid. Samples were contrasted with 4% aqueous uranyl acetate (10 min) and then Reynolds lead citrate (2 min). The autophagosomes were observed under a transmission electron microscope and imaged.
Western blot analysis
Each group of Ishikawa cells was washed twice with ice-cold PBS, and resuspended in ice-cold RIPA buffer supplemented with 1 mmol/l phenylmethanesulfonyl fluoride and a cocktail of protease inhibitors (dilution, 1:100; Beyotime Institute of Biotechnology, Nantong, China). Samples were centrifuged at 4°C for 20 min at 800 × g. Supernatants were recovered and total protein was quantified using the aforementioned BCA protein assay kit. Equal amounts of protein were loaded and separated on 10% SDS-polyacrylamide gels and transferred to PVDF membranes. The membranes were blocked for 1 h at room temperature with 5% milk in TBS supplemented with 0.05% Tween-20 (TBST), incubated for 1 h with the corresponding primary antibody [anti-PI3K p85, dilution, 1:5,000; anti-phosphorylated (p)-AKT1, dilution, 1:2,000; anti-total (t)-AKT1, dilution, 1:1,000; anti-p-mTOR, dilution, 1:2,000; anti-t-mTOR, dilution, 1:5,000; and anti-GAPDH, dilution 1:5,000], washed three times with TBST, incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (dilution, 1:20,000; 40 min at 37°C), washed three times with TBST and visualized using the aforementioned ECL reagent. GAPDH served as an internal loading control.
Immunofluorescence microscopy
Ishikawa cells treated with 0, 10, 20, 40 or 80 µg/ml were grown on coverslips. After 0, 12 or 24 h, the cells were washed with PBS and fixed for 20 min with 4% paraformaldehyde at room temperature. The cells were then permeabilized for 5 min in 0.2% Triton X-100 and washed with PBS. In order to block non-specific background staining, the samples were incubated at 37°C in a solution containing 10% normal goat serum (Boster Biological Technology) for 30 min. Upon washing with PBS, the cells were incubated for 1 h at room temperature with primary rabbit polyclonal anti-LC3B antibody diluted in PBS supplemented with 1% bovine serum albumin (Boster Biological Technology). The cells were then rinsed with PBS and incubated with Alexa Fluor 488-conjugated donkey anti-rabbit IgG secondary antibodies for 60 min at room temperature. The nuclei were counterstained with DAPI for 5 min at room temperature. Upon washing with PBS, the stained cells were mounted with a fluorescent-mounting medium (Dako; Agilent Technologies, Inc., Santa Clara, CA, USA) and examined using a TCS SP2 AOBS confocal laser scanning microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Statistical analysis
All results are presented as the mean ± standard deviation, and were analysed using SPSS version 19.0 software (IBM SPSS, Armonk, NY, USA). A one-way analysis of variance was used to determine statistical significance. P<0.05 was considered to indicate a statistically significant difference.
Results
Effect of CDDP treatment on Ishikawa cell proliferation
Prior to investigating the effect of CDDP on Ishikawa cell autophagy, the effect of CDDP on Ishikawa cell proliferation was evaluated, and a suitable treatment concentration and time were determined by the results of the cell proliferation assays. Ishikawa cells were treated with 10, 20, 40 and 80 µg/ml CDDP for 12, 24, 48 and 72 h. The cells were then harvested and subjected to cell proliferation assays. As presented in Fig. 1, the proliferation inhibition of Ishikawa cells treated with CDDP demonstrated dose- and time-dependent results. For the 10 µg/ml CDDP treatment, the proliferation inhibition rates were 18.27±2.25, 24.88±3.58, 38.27±5.34 and 45.43±4.25 following 12, 24, 48 and 72 h of treatment, respectively. For the 20 µg/ml CDDP treatment, the proliferation inhibition rate was ≤20% at all time-points. For the 40 and 80 µg/ml CDDP treatment, the proliferation inhibition rate was <50% at all time points. Based on these results, 20 µg/ml CDDP treatment for 12 or 24 h was selected as the concentration and time periods for the subsequent assays.
Figure 1.
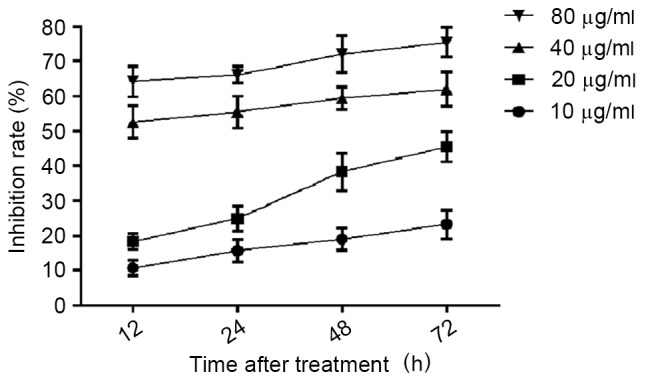
Effect of CDDP treatment on Ishikawa cell proliferation. Ishikawa cells were treated with 10, 20, 40 and 80 µg/ml CDDP for 12, 24, 48 and 72 h. Cell proliferation assay was then performed using the CellTiter 96® AQueous One Solution Cell Proliferation Assay kit, according to the manufacturer's protocol. CDDP; cisplatin.
Promotion of cell autophagy induced by CDDP treatment in Ishikawa cells
To detect the effect of CDDP on Ishikawa cell autophagy, the formation of autophagosomes was observed using a transmission electron microscope. As presented in Fig. 2A, the number of intracellular autophagosomes following 20 µg/ml CDDP treatment for 12 h increased compared with that of the control group (no treatment). In addition, the number of intracellular autophagosomes following 20 µg/ml CDDP treatment for 24 h was higher than that following 20 µg/ml CDDP treatment for 12 h. Furthermore, in order to determine the effect of CDDP treatment on cell autophagy, the expression level of the autophagy-related gene (ATG) LC3 (17), was examined using immunofluorescence microscopy. The LC3 protein expression levels following 20 µg/ml CDDP treatment for 12 h were increased compared with those of the group that did not receive treatment. The LC3 protein expression level following 20 µg/ml CDDP treatment for 24 h was higher than that following 20 µg/ml CDDP treatment for 12 h. In brief, CDDP treatment may promote cell autophagy in a time-dependent manner in Ishikawa cells. Based on these results, 20 µg/ml CDDP treatment for 24 h was selected as the concentration and time to be used for the following assays.
Figure 2.
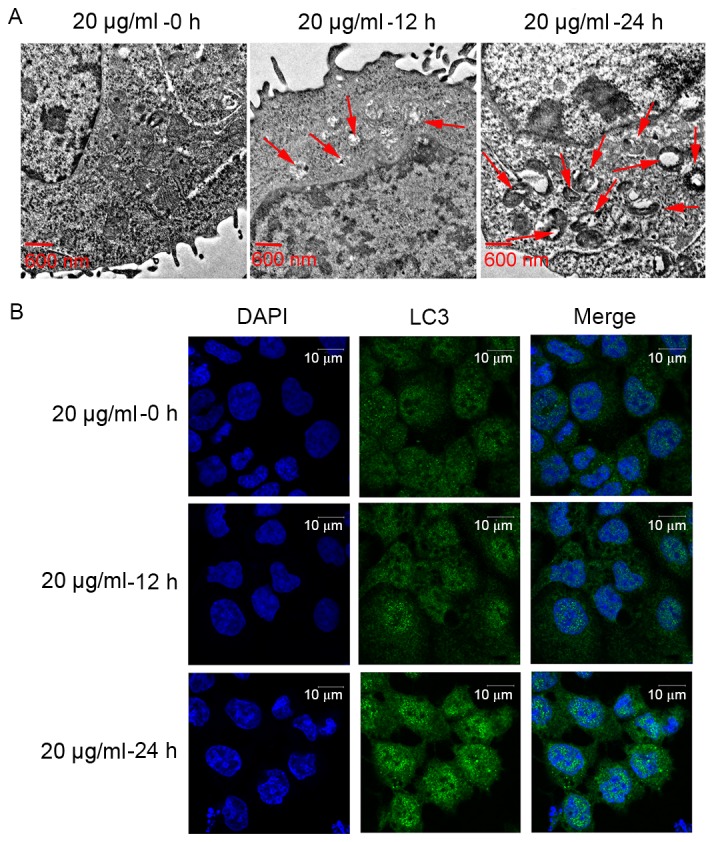
CDDP treatment promotes cell autophagy in Ishikawa cells. (A) CDDP treatment enhanced the formation of intracellular autophagosomes in Ishikawa cells. Representative transmission electron microscope images from 20 µg/ml CDDP-treated cells for 0, 12 and 24 h are presented (arrows indicate intracellular autophagosomes). (B) The expression level of the autophagy-associated gene LC3 was examined using immunofluorescence microscopy. CDDP; cisplatin; LC3, microtubule-associated protein 1 light chain 3α.
Inactivation of the PI3K/AKT/mTOR signalling pathway by CDDP treatment
To investigate the mechanism underlying the effect of CDDP treatment on autophagy in Ishikawa cells, the protein expression level of PI3K p85, p-AKT1, t-AKT1, p-mTOR and t-mTOR were evaluated by the western blot analysis. As presented in Fig. 3, the t-AKT1 and t-mTOR expression levels remained constant; however, the expression levels of p-Akt and p-mTOR were reduced in cells treated with 20 µg/ml CDDP for 24 h. Reduction in the expression level of PI3K p85 was also observed.
Figure 3.
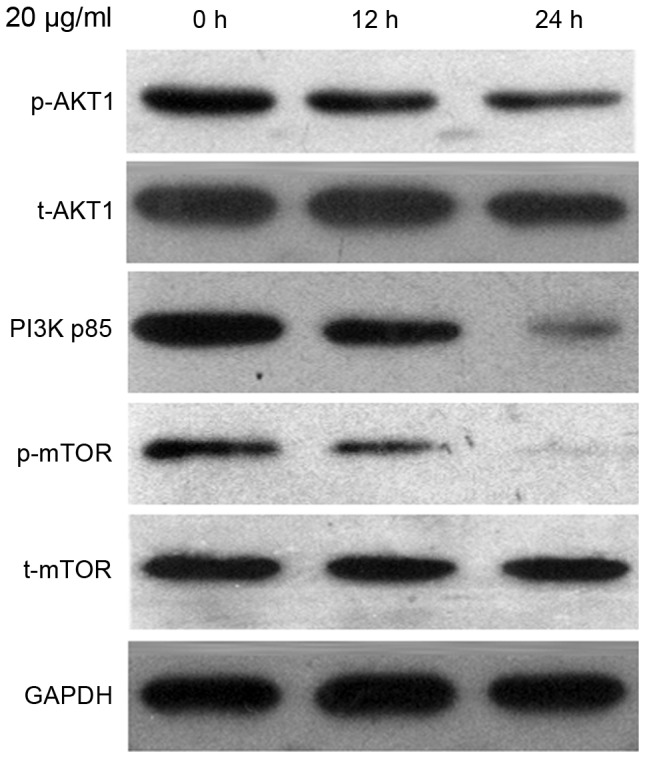
The protein expression level of PI3K p85, p-AKT1, t-AKT1, p-mTOR and t-mTOR were detected using western blot analysis. Ishikawa cells were treated with 20 µg/ml CDDP for 12 and 24 h, followed by harvesting for western blot analysis. PI3K, phosphoinositide 3-kinase; p-, phosphorylated; t-, total; AKT, protein kinase B; mTOR, mammalian target of rapamycin; CDDP, cisplatin.
IGF-1 co-treatment reverses the effect of CDDP on cell autophagy in Ishikawa cells
To further evaluate the effect of CDDP on cell autophagy in Ishikawa cells via the PI3K/AKT/mTOR signalling pathway, the cells were co-treated with a PI3K activator, IGF-1, at 100 ng/ml. As presented in Fig. 4, the number of intracellular autophagosomes following treatment with 20 µg/ml CDDP and IGF-1 co-treatment for 24 h was decreased compared with that in the 20 µg/ml CDDP treatment group. However, the number of intracellular autophagosomes following 20 µg/ml CDDP and IGF-1 co-treatment was higher than that in the cells treated with DMSO alone. Furthermore, the expression level of the ATG LC3 was examined using immunofluorescence microscopy. The LC3 protein expression levels following 20 µg/ml CDDP and IGF-1 co-treatment were decreased compared with those in the 20 µg/ml CDDP treatment group. However, the LC3 protein expression level following 20 µg/ml CDDP and IGF-1 co-treatment was higher than that in the DMSO-only group. In brief, IGF-1 co-treatment may partially reverse the effect of CDDP on cell autophagy in Ishikawa cells.
Figure 4.
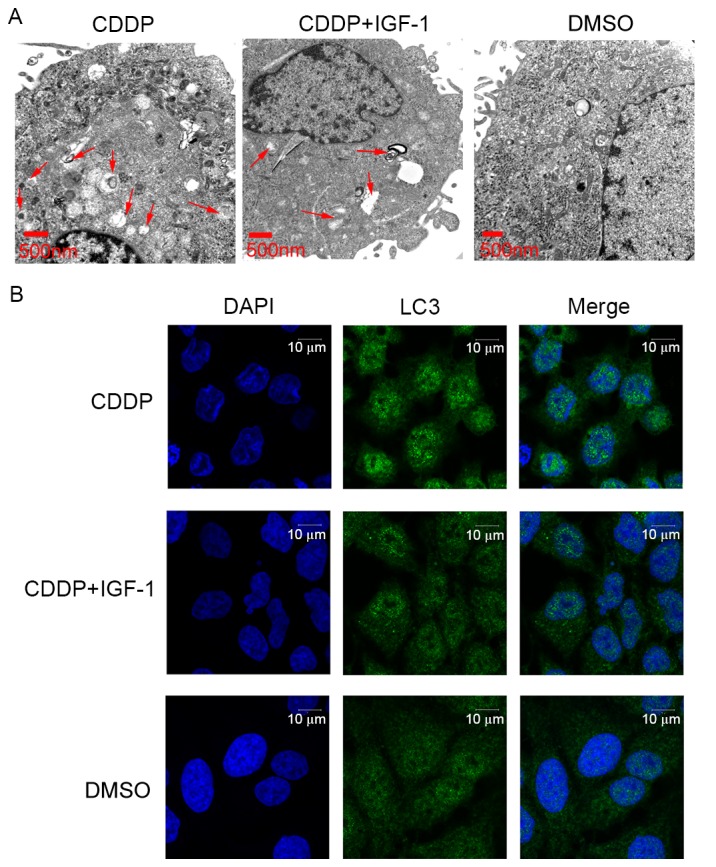
IGF-1 co-treatment reverses the effect of CDDP on cell autophagy in Ishikawa cells. (A) CDDP and IGF-1 co-treatment reduced the formation of intracellular autophagosomes in Ishikawa cells. Representative transmission electron microscope images from the 20 µg/ml CDDP treatment group, 20 µg/ml CDDP and IGF-1 co-treatment group and the control cell group are presented (arrows indicate intracellular autophagosomes). (B) The expression level of the autophagy-associated gene LC3 was examined using immunofluorescence microscopy. CDDP, cisplatin; IGF-1, insulin-like growth factor 1; LC3, microtubule-associated protein 1 light chain 3α.
Discussion
CDDP is a highly effective chemotherapeutic agent used for the treatment of human solid tumours, including endometrial cancer (18). However, the cytotoxicity induced by CDDP is an important obstacle in its utility and therapeutic profile. CDDP and other platinum-based compounds are considered as cytotoxic drugs that kill cancer cells by damaging the DNA, inhibiting DNA synthesis and mitosis, and inducing apoptotic cell death (19). CDDP chemotherapy is also associated with substantial side effects, including hepatotoxic, nephrotoxic, cardiotoxic, neurotoxic and/or hematotoxic damage (19). The molecular mechanism underlying the cytotoxic effects induced by CDDP was widely discussed in previous studies, and numerous molecular mechanisms are explained, including the induction of oxidative stress, which is characterized by reactive oxygen species production and lipid peroxidation; induction of p53 signalling and cell cycle arrest; downregulation of proto-oncogenes and anti-apoptotic proteins; and activation of intrinsic and extrinsic pathways of apoptosis (19–22). In previous studies, autophagy, a highly regulated self-degradation process of eukaryotic cells, was identified as one of the molecular mechanisms underlying CDDP cytotoxicity in certain types of cancer cells and in normal cells (10,23). Autophagy is a context-dependent tumour-suppressing mechanism that can also promote tumour cell survival under stress conditions and treatment resistance (24). The present study aimed to discuss the association between autophagy and CDDP in endometrial cancer. Therefore, the current results will aid in the explanation of molecular mechanisms underlying CDDP cytotoxicity in endometrial cancer.
The dynamic process of autophagy involves membrane formation and fusion, including autophagosome formation, autophagosome-lysosome fusion and the degradation of intra-autophagosomal contents by lysosomal hydrolases (25). The present study used transmission electron microscopy to detect the formation of intracellular autophagosomes in Ishikawa cells following CDDP treatment. The results revealed that CDDP treatment may increase the number of intracellular autophagosomes in a time-dependent manner in Ishikawa cells. To further determine the effect of CDDP treatment on cell autophagy, the expression level of the ATG LC3 (15) was examined using immunofluorescence microscopy. The results demonstrated that CDDP treatment may upregulate LC3 protein expression level. Together, these results revealed that CDDP treatment promoted cell autophagy in the Ishikawa endometrial cancer cells. The results of the present study are in accord with previous studies (10,12). In Huh7 and HepG2 hepatocellular carcinoma cells, CDDP treatment activated autophagy (10), and in human lung adenocarcinoma cells, LC3-II was increased in the A549/DDP CDDP-resistant cells compared with that in A549 cells (12).
PI3K/AKT/mTOR signalling pathway activation is heavily implicated in endometrial cancer pathogenesis (14). The results of the present study revealed that CDDP treatment may inactivate the PI3K/AKT/mTOR signalling pathway. In addition co-treatment with a PI3K activator, IGF-1, may reverse the effect of CDDP on cell autophagy in Ishikawa cells. The association between CDDP and the PI3K/AKT/mTOR signalling pathway was also discussed in other cells. In non-small cell lung cancer cells, platycodin-D induced autophagy in NCI-H460 and A549 cells via inhibition of the PI3K/AKT/mTOR signalling pathway (26), while in human cervical cancer cells, licochalcone A induced autophagy via inactivation of the PI3K/AKT/mTOR signalling pathway (27). Based on these results, the present study hypothesized that CDDP may regulate cell autophagy in the Ishikawa endometrial cancer cells by the PI3K/AKT/mTOR signalling pathway.
Acknowledgements
The present study was supported by the General Guidance Project of Guangzhou City Health Bureau (grant no. 20151A010106).
References
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Magrina JF, Mutone NF, Weaver AL, Magtibay PM, Fowler RS, Cornella JL. Laparoscopic lymphadenectomy and vaginal or laparoscopic hysterectomy with bilateral salpingo-oophorectomy for endometrial cancer: Morbidity and survival. Am J Obstet Gynecol. 1999;181:376–381. doi: 10.1016/S0002-9378(99)70565-X. [DOI] [PubMed] [Google Scholar]
- 3.National Cancer Institute: Cancer Stat Facts: Endometrial Cancer. 2015 [Google Scholar]
- 4.Klionsky DJ, Emr SD. Autophagy as a regulated pathway of cellular degradation. Science. 2000;290:1717–1721. doi: 10.1126/science.290.5497.1717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.White E, DiPaola RS. The double-edged sword of autophagy modulation in cancer. Clin Cancer Res. 2009;15:5308–5316. doi: 10.1158/1078-0432.CCR-07-5023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Du F, Feng Y, Fang J, Yang M. MicroRNA-143 enhances chemosensitivity of Quercetin through autophagy inhibition via target GABARAPL1 in gastric cancer cells. Biomed Pharmacother. 2015;74:169–177. doi: 10.1016/j.biopha.2015.08.005. [DOI] [PubMed] [Google Scholar]
- 7.Liu YP, Dong FX, Chai X, Zhu S, Zhang BL, Gao DS. Role of autophagy in capsaicin-induced apoptosis in U251 Glioma Cells. Cell Mol Neurobiol. 2016;36:737–743. doi: 10.1007/s10571-015-0254-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Nazim UM, Park SY. Genistein enhances TRAIL-induced cancer cell death via inactivation of autophagic flux. Oncol Rep. 2015;34:2692–2698. doi: 10.3892/or.2015.4247. [DOI] [PubMed] [Google Scholar]
- 9.Zhang Y, Jiang F, Bao W, Zhang H, He X, Wang H, Wan X. SOX17 increases the cisplatin sensitivity of an endometrial cancer cell line. Cancer Cell Int. 2016;16:29. doi: 10.1186/s12935-016-0304-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Xu N, Zhang J, Shen C, Luo Y, Xia L, Xue F, Xia Q. Cisplatin-induced downregulation of miR-199a-5p increases drug resistance by activating autophagy in HCC cell. Biochem Biophys Res Commun. 2012;423:826–831. doi: 10.1016/j.bbrc.2012.06.048. [DOI] [PubMed] [Google Scholar]
- 11.Kang R, Wang ZH, Wang BQ, Zhang CM, Gao W, Feng Y, Bai T, Zhang HL, Huang-Pu H, Wen SX. Inhibition of autophagy-potentiated chemosensitivity to cisplatin in laryngeal cancer Hep-2 cells. Am J Otolaryngol. 2012;33:678–684. doi: 10.1016/j.amjoto.2012.05.005. [DOI] [PubMed] [Google Scholar]
- 12.Ren JH, He WS, Nong L, Zhu QY, Hu K, Zhang RG, Huang LL, Zhu F, Wu G. Acquired cisplatin resistance in human lung adenocarcinoma cells is associated with enhanced autophagy. Cancer Biother Radiopharm. 2010;25:75–80. doi: 10.1089/cbr.2009.0701. [DOI] [PubMed] [Google Scholar]
- 13.Chen J, Zhao KN, Li R, Shao R, Chen C. Activation of PI3K/Akt/mTOR pathway and dual inhibitors of PI3K and mTOR in endometrial cancer. Curr Med Chem. 2014;21:3070–3080. doi: 10.2174/0929867321666140414095605. [DOI] [PubMed] [Google Scholar]
- 14.Slomovitz BM, Coleman RL. The PI3K/AKT/mTOR pathway as a therapeutic target in endometrial cancer. Clin Cancer Res. 2012;18:5856–5864. doi: 10.1158/1078-0432.CCR-12-0662. [DOI] [PubMed] [Google Scholar]
- 15.Lum JJ, Bauer DE, Kong M, Harris MH, Li C, Lindsten T, Thompson CB. Growth factor regulation of autophagy and cell survival in the absence of apoptosis. Cell. 2005;120:237–248. doi: 10.1016/j.cell.2004.11.046. [DOI] [PubMed] [Google Scholar]
- 16.Levine B, Kroemer G. Autophagy in the pathogenesis of disease. Cell. 2008;132:27–42. doi: 10.1016/j.cell.2007.12.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chen Y, Li X, Wu X, He C, Guo L, Zhang S, Xiao Y, Guo W, Tan B. Autophagy-related proteins LC3 and Beclin-1 impact the efficacy of chemoradiation on esophageal squamous cell carcinoma. Pathol Res Pract. 2013;209:562–567. doi: 10.1016/j.prp.2013.06.006. [DOI] [PubMed] [Google Scholar]
- 18.Youn CK, Kim J, Park JH, Do NY, Cho SI. Role of autophagy in cisplatin-induced ototoxicity. Int J Pediatr Otorhinolaryngol. 2015;79:1814–1819. doi: 10.1016/j.ijporl.2015.08.012. [DOI] [PubMed] [Google Scholar]
- 19.Dasari S, Tchounwou PB. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur J Pharmacol. 2014;740:364–378. doi: 10.1016/j.ejphar.2014.07.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jiang Y, Guo C, Vasko MR, Kelley MR. Implications of apurinic/apyrimidinic endonuclease in reactive oxygen signaling response after cisplatin treatment of dorsal root ganglion neurons. Cancer Res. 2008;68:6425–6434. doi: 10.1158/0008-5472.CAN-08-1173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.DeHaan RD, Yazlovitskaya EM, Persons DL. Regulation of p53 target gene expression by cisplatin-induced extracellular signal-regulated kinase. Cancer Chemother Pharmacol. 2001;48:383–388. doi: 10.1007/s002800100318. [DOI] [PubMed] [Google Scholar]
- 22.Zhu H, Luo H, Zhang W, Shen Z, Hu X, Zhu X. Molecular mechanisms of cisplatin resistance in cervical cancer. Drug Des Devel Ther. 2016;10:1885–1895. doi: 10.2147/DDDT.S106412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kaushal GP, Kaushal V, Herzog C, Yang C. Autophagy delays apoptosis inrenal tubular epithelial cells in cisplatin cytotoxicity. Autophagy. 2008;4:710–712. doi: 10.4161/auto.6309. [DOI] [PubMed] [Google Scholar]
- 24.Kubisch J, Turei D, Földvári-Nagy L, Dunai ZA, Zsákai L, Varga M, Vellai T, Csermely P, Korcsmáros T. Complex regulation of autophagy in cancer-integrated approaches to discover the networks that hold a double-edged sword. Semin Cancer Biol. 2013;23:252–261. doi: 10.1016/j.semcancer.2013.06.009. [DOI] [PubMed] [Google Scholar]
- 25.Tanida I. Autophagosome formation and molecular mechanism of autophagy. Antioxid Redox Signal. 2011;14:2201–2214. doi: 10.1089/ars.2010.3482. [DOI] [PubMed] [Google Scholar]
- 26.Zhao R, Chen M, Jiang Z, Zhao F, Xi B, Zhang X, Fu H, Zhou K. Platycodin-D induced autophagy in non-small cell lung cancer cells via PI3K/Akt/mTOR and MAPK Signaling Pathways. J Cancer. 2015;6:623–631. doi: 10.7150/jca.11291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tsai JP, Lee CH, Ying TH, Lin CL, Hsueh JT, Hsieh YH. Licochalcone A induces autophagy through PI3K/Akt/mTOR inactivation and autophagy suppression enhances Licochalcone A-induced apoptosis of human cervical cancer cells. Oncotarget. 2015;6:28851–28866. doi: 10.18632/oncotarget.4767. [DOI] [PMC free article] [PubMed] [Google Scholar]