

Abstract
The phylum Rotifera consists of minuscule, nonsegmented animals with a unique body plan and an unresolved phylogenetic position. The presence of pharyngeal articulated jaws supports an inclusion in Gnathifera nested in the Spiralia. Comparison of Hox genes, involved in animal body plan patterning, can be used to infer phylogenetic relationships. Here, we report the expression of five Hox genes during embryogenesis of the rotifer Brachionus manjavacas and show how these genes define different functional components of the nervous system and not the usual bilaterian staggered expression along the anteroposterior axis. Sequence analysis revealed that the lox5-parapeptide, a key signature in lophotrochozoan and platyhelminthean Hox6/lox5 genes, is absent and replaced by different signatures in Rotifera and Chaetognatha, and that the MedPost gene, until now unique to Chaetognatha, is also present in rotifers. Collectively, our results support an inclusion of chaetognaths in gnathiferans and Gnathifera as sister group to the remaining spiralians.
Rotifers are microscopic animals with an unusual, nonsegmented body plan consisting of a head, trunk and foot. Here, Fröbius and Funch investigate the role of Hox genes—which are widely used in animal body plan patterning—in rotifer embryogenesis and find non-canonical expression in the nervous system.
Introduction
Originally discovered in Drosophila melanogaster 1, 2, Hox gene transcription factors have been researched extensively in a phylogenetically diverse range of animals over the last 30 years. Being present in the genomes of nearly all animals with exception of Porifera, Ctenophora, and Placozoa, this family of highly conserved transcriptional regulators controls some of the most fundamental processes of embryonic development. During morphogenesis of all triploblastic metazoans studied, Hox genes exhibit expression in body regions along the antero-posterior axis of the embryo at some point. Regional identities are imprinted by either single or combined expression of different Hox genes, referred to as “Hox code”. This is clearly seen in the segmented taxa Arthropoda3, Annelida4, 5, and Chordata6, 7 where the exact identity of body segments is regulated by the “Hox code”. Here, deviations from the appropriate expression profiles result in homeotic transformations of the body regions involved. Amazingly, even across distantly related taxa, body regions comparable to each other (anterior, median, posterior) are patterned by orthologous Hox genes. Thus, evolutionary changes of Hox gene expression may have led to evolutionary shifts accounting for the emergence of new body plans8. Having arisen by tandem duplication, Hox genes are often physically linked in genomic clusters. A correlation of the spatial order of Hox gene expression along a primary body axis and sometimes even temporal order of Hox gene activation during embryogenesis with the order of the genes in the Hox cluster, referred to as colinearity9, has been observed in a broad range of metazoans. In some bilaterian taxa, however, the Hox cluster has been rearranged10 or even dispersed up to the relocation of all Hox genes to different locations of the genome11, 12. Degradation of the Hox cluster, however, does not necessarily result in destruction of the Hox code. In some cases, specification of appropriate body regions is maintained in taxa where Hox genes are no longer closely linked13, 14.
Within Protostomia, Hox gene expression has been studied in a variety of ecdysozoan and spiralian taxa. However, Hox gene activation during embryonic development has not been studied in gnathiferans so far. Based on morphological characters, it has been proposed that the phylum Rotifera including the parasitic Acanthocephala, along with Gnathostomulida and Micrognathozoa, form the clade Gnathifera15–17. Rotifers are microscopic, ecologically important aquatic animals comprising ca. 2200 described species. Their embryonic development and sexual dimorphic adult body plans exhibit special features. Protected within an egg shell, development starts with a unique cleavage pattern involving an exceptional type of D-quadrant cleavage18–20. Lacking a larval form, direct development with early determination gives rise to eutelic animals typically with a tripartite, pseudocoelomic body plan, consisting of a head with a ciliated corona, a trunk and a post-cloacal foot. Several rotiferan tissues are syncytial but musculature and nervous system are cellular21. Muscles are mostly formed by few cells directly innervated by nerves formed by simple chains of single neurons22. The excretory system consists of paired protonephridia with both cellular and syncytial sections21.
Phylogenomic approaches show that rotiferan sequences exhibit very high evolutionary rates resulting in unstable positions within calculated trees23–25. Likewise, Platyhelminthes and some ecdysozoans display fast evolving sequences. As a result, Rotifera are prone to long-branch-attraction (LBA) artifacts by grouping with e.g., Platyhelminthes. In addition, taxonomic sampling within Gnathifera is sparse. This has lead to the controversial hypothesis of a clade “Platyzoa” uniting Gnathifera with Platyhelminthes and Gastrotricha within Spiralia (Supplementary Fig. 1)26, 27. Recent phylogenomic approaches focusing on resolving spiralian phylogeny, though, support paraphyly of Platyzoa and monophyly of Gnathifera placing Gnathifera basally near the Spiralia/Ecdysozoa split as sister to Lophotrochozoa and Rouphozoa (including Platyhelminthes and Gastrotricha)28–30.
Based on the presence of specific amino-acid residues and peptide motifs within the homeodomain and its flanking regions—referred to as signatures—Hox genes can be assigned to various paralogous groups (PG1-15)31, 32. These paralogous groups are quite clearly defined for anterior class PG1-2, PG3 and median class PG4-5 Hox genes. Evolution of these Hox genes predates the divergence of Protostomia and Deuterostomia. Due to independendly duplicated median and posterior class Hox genes in different bilaterian lineages later on, the exact paralogy status of the remaining median genes (PG6-8) and posterior class genes (PG9-14) is more difficult to determine given the lack of diagnostic position and overall phylogenetic signal. However, these duplication events and subsequent selection present us with Hox gene orthologues along with their conserved amino-acid signatures characteristic for these specific lineages in the clades existing today, allowing us to examine phylogenetic relationships based on Hox gene complements and the Hox signatures within33.
In this study, we isolate and examine genes of the Hox complement of the monogonont rotifer Brachionus manjavacas. Our analysis of Hox gene expression during embryogenesis of amictic females shows non-canonical expression patterns in the developing nervous system consistent with an original role of Hox genes in neurogenesis. Our sequence analyses show the presence of a new signature in the Hox6 paralog of B. manjavacas, shared by chaetognath Hox6 genes only. Furthermore, one of the rotifer Hox genes possesses median- and posterior-like amino-acid residues, exhibiting similarity to chaetognath MedPost genes. These results provide evidence for inclusion of both Rotifera and Chaetognatha in Gnathifera and also support a basal phylogenetic position of Gnathifera as sister group to the remaining Spiralia.
Results
Rotiferan Hox genes and metazoan phylogeny
We isolated single copies of five Hox genes from the monogonont rotifer Brachionus manjavacas. Based on phylogenetic analyses of the homeodomain and diagnostic amino-acid motifs, we assigned orthology of these genes to the anterior class Hox gene PG2 (Bm-Hox2), a PG3 gene (Bm-Hox3) and central class genes PG4 (Bm-Hox4) and PG6 (Bm-Hox6) (Supplementary Figs. 2, 3, 4, and 5). The fifth Hox gene isolated from Brachionus manjavacas surprisingly clusters with MedPost genes from the chaetognaths Flaccisagitta enflata and Spadella cephaloptera (Fig. 1a, Supplementary Figs. 3, 4, and 5) and is strongly supported with a posterior probability of 100% in Bayesian analysis. Maximum-likelyhood (ML) bootstrap support for this grouping is only 63%, this, however, is comparable to the ML support of grouping of the ecdysozoan AbdB genes (67%) or all Saccoglossus kowalevskii Hox11-13 genes analyzed (61%) and even higher than the support for grouping of all Lox5-genes undoubtedly related (< 50%) (Supplementary Fig. 5). Mean statistical support from ML analyses for Hox gene orthology assignments usually is significantly lower due to the highly conserved nature of the homeodomain34. While it could be argued that accelerated evolution could have led to phylogenetic artefacts as LBA, phylogenetic analyses did not reveal branch lengths for the grouping of the chaetognath and rotifer MedPost genes significantly larger than those observed for some posterior class Hox genes in general.
Fig. 1.
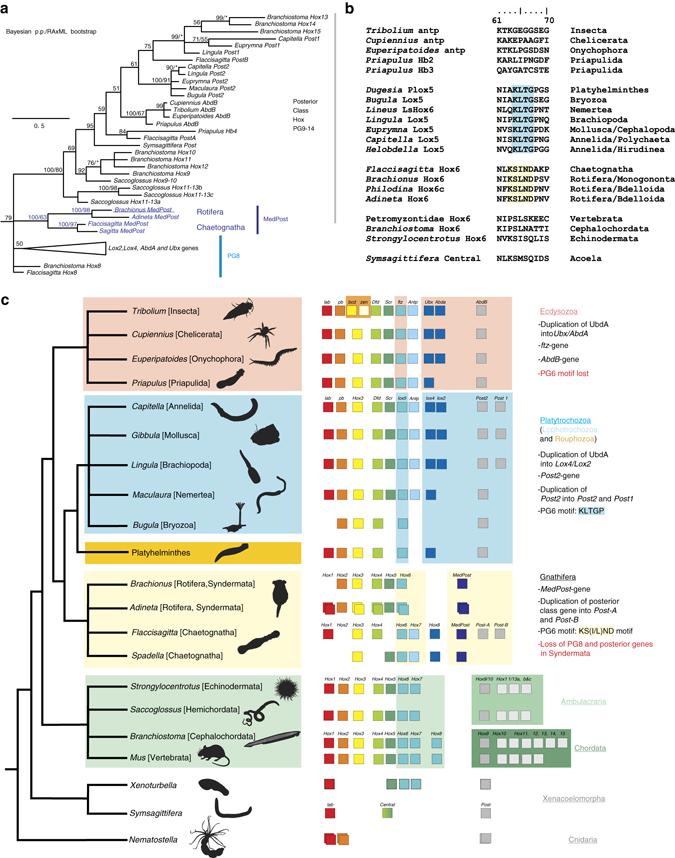
Hox gene data places rotifers and chaetognaths in Gnathifera within Spiralia. a Phylogenetic tree depicting the relationship of MedPost genes to PG8 and posterior class Hox genes. Tree topology is from Bayesian analysis. Bayesian posterior probabilities based on 400,000 trees from 40,000,000 generations and ML support values from 1000 iterations are shown above branches. Single values represent Bayesian posterior probabilities only. Asterisks denote ML support below 50%. b Alignment of ten amino acids of the carboxy flanking region to the homeodomain of PG6 genes. Sequences highlighted with yellow contain the new signature found in rotifers and chaetognaths. Blue highlighting marks the lox5-parapeptide of lophotrochozoan genes. Neither is found in Ecdysozoa, Ambulacraria, Chordata, or Xenacoelomorpha. c Summary of representative characteristics of the Hox cluster within different metazoan taxa. The tree to the left represents bilaterian phylogeny with Cnidaria as an outgroup. Boxes in the middle depict Hox gene contingents (color coded according to the assignment of the Hox genes to the different paralogous groups) isolated from representative species. The right hand column summarizes characteristic Hox gene evolution and duplication events along with presence of special Hox signatures resulting in Hox genes characterizing the respective groups
A careful examination of the homeodomain alignment of MedPost genes with either central class Hox genes or posterior class Hox genes (Supplementary Fig. 3) illustrates both, similarities and differences between rotiferan and chaetognath MedPost genes. Both groups share nine of eleven central class diagnostic amino acids: Q (position 6 of the homeodomain), LTR(R/K)RR (26–31) and E (59). Previous work on MedPost genes of Flaccisagitta enflata 35 and Spadella cephaloptera 36 defined diagnostic posterior class residues characteristic for chaetognath MedPost genes: K(3), A(14), R(18), Y(20) and V(21). Only two of these posterior class diagnostic residues, K(3) and Y(20) are shared between chaetognaths and rotifers, but clearly, they represent plesiomorphic characters since they are also present in posterior Hox genes of all major bilaterian clades and interestingly also in chaetognath PostA and PostB genes (Supplementary Fig. 3). The other amino-acid residues A(14), R(18), and V(21) present in chaetognaths but not in rotifers are neither found in posterior class Hox genes of Ecdysozoa nor in those of Deuterostomia, but surprisingly in Post1 genes typical of Lophotrochozoa (Supplementary Fig. 3). Amino-acid residues supposedly characteristic for a new gene class thus have to be re-evaluated once new genes sharing these characteristics have been isolated. MedPost genes in general might be defined by only some of the posterior class specific amino-acid positions while others might be specific for one of the taxa only. Hox genes belonging to the same paralogous group often exhibit diversity at some positions of the homeodomain (Supplementary Fig. 2). The presence of MedPost could also be interpreted as a possible ancestral character, but if it was lost in the Lophotrochozoan + Rouphozoan lineage chaetognaths could still be affiliated to gnathiferans.
The possible close relationship between Rotifera and Chaetognatha was further supported when the homeodomain and 3′ flanking sequences of the PG6 gene Bm-Hox6 were analyzed (Fig. 1b). Lophotrochozoans and Platyhelminthes possess some central class Hox genes containing amino-acid motifs not observed in ecdysozoan or deuterostome taxa. These genes have been named Lox5 (PG6/7), Lox2, and Lox4 (both PG8). Lox5 orthologs possess the motif “KLTGP” in the carboxy flanking region of the homeodomain. (position 64–68)31, 37, and it seems likely that this motif was present in a common ancestor of Lophotrochozoa and a newly proposed clade Rouphozoa30 consisting of Platyhelminthes and Gastrotricha. Importantly, the PG6 gene of B. manjavacas not only lacks this “Lox5-parapeptide”, it possesses a new Hox signature “KS(I/L)ND” at position 63–67 also identified in PG6 genes of the bdelloid rotifers Philodina roseola and Adineta vaga, and the chaetognath Flaccisagitta enflata (Fig. 1b). This motif is not entirely identical among rotifers and chaetognaths, however, Hox signatures exhibit some variability as known from the variant Lox5 signature of Myzostomida38 and some Platyhelminthes39 or the Ubd-A peptide of Spiralia35. Recent phylogenomic approaches support Ecdysozoa as sister group to Spiralia, and neither ecdysozoan nor deuterostome PG6 genes possess a Hox signature in the carboxy flanking region to the homeodomain. Moreover PG6 Hox genes are absent in Xenacoelomorpha40–42 sister group to Nephrozoa (Deuterostomia, Ecdysozoa, and Spiralia)43, 44. Thus both signatures, the “Lox5-parapeptide” and the “KS(I/L)ND” signature could have evolved independently after the split of Ecdysozoa and Spiralia. The alternative hypothesis that the new signature is plesiomorphic has weaker support.
Consistent with their absence in the publically available genome of the bdelloid rotifer Adineta vaga 45, PG8 genes (Lox2/Lox4/Ubx/AbdA) and posterior Hox genes (PG9-14) were not recovered from the monogonont rotifer B. manjavacas suggesting that these genes are missing in Rotifera (Fig. 1c). Another central class gene (PG5) has recently been recovered from the genome of Brachionus manjavacas while the presence of the second anterior class gene (PG1) as identified in A. vaga 45 could not be confirmed (D. M. Welch, personal communication). Interestingly a single PG8 gene with similarity to ecdysozoan and lophotrochozoan PG8 genes and two posterior Hox genes sharing key residues with lophotrochozoans, ecdysozoans, and deuterostomes, have been isolated from the chaetognath F. enflata 35. Posterior class Hox genes are also present in the cnidarian Nematostella vectensis 34 and have been shown to exhibit extraordinary flexibility leading to possible independent evolution in different lineages46. Thus, the most parsimonous explanation is that both PG8 Hox genes and the posterior genes (PG9-14) have been lost in rotifers (Fig. 1c).
Some phylogenomic analyses found support for a group called Platyzoa consisting of Gnathifera, Gastrotricha, and Platyhelminthes23, 24, 47, but morphological characteristics supporting this group have never been strong. All platyzoans are non-coelomate, ciliated animals with worm-like appearance without specialized respiratory or vascular systems27. However, most platyzoans are microscopic and aquatic making diffusion an effective transport mechanism and vascular systems and respiratory organs are unnecessary. Developmental features uniting Platyhelminthes, namely spiral cleavage, resulting cell-lineage and the characteristic Müller’s and Götte’s larvae regarded to be modified trochophores can neither be found in gnathiferan taxa nor in gastrotrichs. Overall morphological synapomorphies supporting Platyzoa are hard to find.
The presence of a MedPost gene and the differing signature in the PG6 gene of rotifers and chaetognaths points to a close relationship, which is supported by several shared morphological and developmental traits (Table 1). Gnathifera, which is well supported by phylogenomic studies, is named after the presence of complex chitinous jaws used for feeding, which is found in Rotifera, Micrognathozoa, and Gnathostomulida15, 48, 49.
Table 1.
Morphological, developmental, and special characteristics of the Hox cluster of spiralian taxa combined provide an informative basis for the phylogenetic relationship of rotifers and chaetognaths
| Chaetognatha | Rotifera | Micrognathozoa | Gnathostomulida | Gastrotricha | Platyhelminthes | Lophotrochozoa | |
|---|---|---|---|---|---|---|---|
| Morphological characteristics | |||||||
| Tripartite body plan with anus terminal of medial body region | + | + | –No anus | –No anus | − | –No anus | −a |
| Stomatogastric nerve plexi | + | + | − | + | − | − | − |
| Additional major nerve plexus in the trunk | + | + | − | − | − | − | − |
| Lateral sensory antennae | + | + | − | − | − | − | − |
| Trunk exterior cilitated | − | − | + | + | + | + | + |
| Complex chitinous structures associated with feeding | + | + | + | + | (−)b | –(No chitin) | (+)c |
| Protonephridia | − | + | + | + | + | + | + |
| Developmental characteristics | |||||||
| D-quadrant cleavage | + | + | ? | + | + | + | + |
| Spiral cleavage | − | − | ? | + | − | + | + |
| 4d mesentoblast | − | − | ? | − | + | + | |
| PGC specification | Preformation | Preformation | ? | Epigenesis | Epigenesis/preformation | Mostly epigenesis | |
| Trochophora larvae | − | −d | ? | ? | − | + | + |
| Hox cluster characteristics | |||||||
| PG6 Hox signature KS(I/L)ND | + | + | ? | ? | ? | − | − |
| MedPost class Hox gene | + | + | ? | ? | ? | − | − |
aPhoronids feature a tripartite body plan with terminal anus
bGastrotrichs possess chitinous pharygeal cuticle
cThe radula of molluscs is chitinous. Being an autapomorphy of this phylum such structures are exceptional among Lophotrochozoa
dThe body plan of a planktotrophic rotifer resembles a neotenic larva similar to trochophores of Lophotrochozoa; however, there are no separate larval and adult stages
Even though chaetognaths do not possess internal structures quite comparable to the jaws of gnathiferans, both the high chitin content of the spines and teeth and the structure of the chitinous cuticle of the chaetognath head could be homologous to the chitinous parts and membranes of the pharynx in gnathiferans. Both chaetognaths and rotifers feature a tripartite body plan consisting of head, trunk without external motile cilia and foot/tail region. In contrast to Lophotrochozoa and Gastrotricha, the anus is not located terminally but instead near the posterior border of the medial region. Also the trunk regions of Lophotrochozoa and Gastrotricha are with motile cilia. The nervous system includes additional nerve plexi: the caudal ganglion in rotifers and the ventral ganglion in chaetognaths. Both also feature lateral sensory antennae connected to the nervous system. The corona, the ciliary organ of rotifers used for downstream collection of food particles, consists of compound cilia while the corona of chaetognaths is formed by a band of monociliate cells. Despite these structural differences, both are innervated by two coronal nerves. The mastax ganglion of rotifers is connected to the brain via two nerves. In chaetognaths, the suboesophageal ganglion is connected to the brain in a similar fashion with two small separate vestibular ganglia integrated in the nerves connecting to the brain. An additional pharyngeal ganglion has also been reported for Gnathostomulida50, 51.
Embryonic development of Chaetognatha and Rotifera shares some important characteristics. Spiral cleavage, prevalent in Lophotrochozoa and Platyhelminthes, is absent in Rotifera and Chaetognatha. The latter two groups exhibit D-quadrant cleavage but do not form a 4d mesentoblast. Primordial germ cells (PGCs) of rotifers and chaetognaths are specified by preformation only, in contrast to the specification of PGCs of Platyhelminthes and Lophotrochozoa by mostly epigenesis52. Unfortunately, hardly anything is known about embryonic development of Micrognathozoa and only the earliest embryonic development has been described for a gnathostomulid species once, indicating possible presence of spiralian cleavage in this group53. Early cleavage patterns of chaetognaths and rotifers differ from each other. In chaetognaths, cleavage is total and equal forming a blastula. A typical invagination gastrula can be observed. This basic cleavage pattern had originally been mistaken as radial cleavage54, 55. Rotifer development involves total and unequal first cleavages. Subsequently columns of cells descending from the A-C quadrant are formed by cleavage with mitotic spindles parallel to the primary axis. The 2D blastomere is then internalized by epibolic gastrulation19. These differences do not necessarily contradict the hypothesis of unison of Chaetognatha and Gnathifera. Even within the morphologically and phylogenetically well supported Gnathifera very different cleavage modes can be observed. The development of the parasitic thorny-headed worms Acanthocephala is different from that reported for monogonont rotifers though both are in included in Syndermata56. The cleavage modes of rotifers and chaetognaths could, therefore, be interpreted as steps in a transformation series towards spiral cleavage with chaetognaths showing a more basal pattern.
These findings are consistent with the results of phylogenetic analyses based on EST data, mitochondrial genomes and tropomyosin where Chaetognatha is sister group to Lophotrochozoa57 or sister group to Protostomia47, 58. Moreover, a phylogenomic study based on EST sequences of 197 genes from 66 metazoan species including both Rotifera and Chaetognatha but unfortunately lacking other gnathiferan groups supports this grouping, albeit weakly25. Newer phylogenetic studies that took LBA artefacts into account placed Gnathifera as sister to Lophotrochozoa and Rouphozoa but excluded chaetognaths from the analysis. Thus, both Rotifera and Chaetognatha were placed at the same position in different phylogenomic studies, indicating a possible close relationship of these taxa. Intriguingly, the newest phylogenomic study including Gnathostomulida, Rotifera, and Chaetognatha shows strong support for a clade including Gnathifera and Chaetognatha as sister to all lophotrochozoans after Bayesian analysis (posterior probability = 1.0) and mediocre support for a clade formed by Gnathifera and Chaetognatha alone (pp = 0.69)29.
Non-canonical expression of rotifer Hox genes
The most fascinating and highly conserved feature of Hox genes is the correlation of spatial expression along the anteroposterior axis with the structure of the genomic Hox cluster. This spatial collinearity results in the formation of nested Hox expression domains by shifting anterior borders of expression in the developing nervous system and other tissues4. Unique combinations of Hox genes activated within a body region specify that region’s identity (Hox-code). Here, we report Hox gene expression patterns in Rotifera. During embryogenesis of the monogonont rotifer Brachionus manjavacas all five isolated Hox genes are expressed in parts of the nervous system and display unique expression patterns unrelated to anteroposterior axis formation (Figs. 2 and 3a).
Fig. 2.
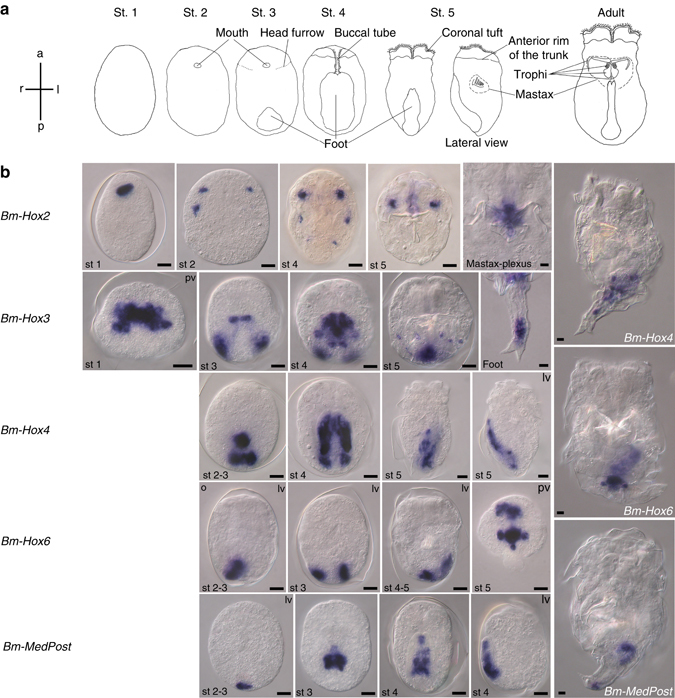
Expression of Hox genes during embryogenesis of Brachionus manjavacas. a Schematic of embryonic stages of Brachionus manjavacas with morphological characteristics used for staging. b Whole-mount in situ hybridization on amictic female embryos. Adults are only shown for genes with expression persisting into the adult stage. Anterior to the top. Mostly ventral views are shown. pv, posterior view, dorsal side up; lv, lateral view, ventral to the left. Scale bar, 10 µm
Fig. 3.

Body plans and nervous systems in Rotifera and Chaetognatha. a Diagram of Hox gene expression in the nervous system of Brachionus manjavacas. b Comparison of rotiferan and chaetognath body plans with respect to the structure of the nervous system. Both groups have a dorsal brain and additional nerve plexi: mastax nerves and ganglia in rotifers and vestibular, and esophageal ganglia in chaetognaths as well as a caudal ganglion in rotifers and a ventral nerve centre in chaetognaths with the latter possibly incorporating functional subsets, these are still separated from the caudal ganglion in rotifers, e.g., innervation of sensory lateral antennae
The anterior class Hox gene Brachionus manjavacas Hox2 (Bm-Hox2) is expressed in cells forming the main ventrolateral nerves connecting the brain to the caudal ganglion, a secondary nerve centre at the base of the foot. Initially upregulated in a pair of cells on the ventral side of the embryo near the anterior pole at the beginning of morphogenesis, these Bm-Hox2-positive cells move laterally and undergo cell divisions in an anterior-to-posterior fashion resulting in paired nerve cords consisting of three interconnected neurons each (Fig. 2 and Supplementary Fig. 6a). Interestingly, anteroposterior patterning of the main longitudinal nerves by nested expression of several Hox genes does not occur. Faint expression of Bm-Hox2 is also detectable in the mastax plexus after hatching (Fig. 2 and Supplementary Fig. 6d, e), consistent with the observation of anterior Hox gene expression in structures associated with the stomatogastric nervous system in other taxa4. Expression of Hox genes in the rotifer brain or in the coronal nerves (Supplementary Fig. 6b, c) has not been detected.
Strikingly, most Hox orthologues (Bm-Hox3, Bm-Hox4, Bm-Hox6, and Bm-MedPost) are expressed in a neurogenic region of the embryo near the posterior pole giving rise to morphological innovations: e.g., the caudal ganglion and foot primordium (Fig. 2). The central class gene Bm-Hox6 is expressed in the developing disc-shaped caudal ganglion originating from a single expression domain shifting inwards from the posterior pole of the embryo. Protruding laterally and ventrally the caudal ganglion resembles a clover leaf. Bm-Hox3 also participates in patterning of this secondary nerve centre. Bilateral symmetrical domains expressing Bm-Hox3 at the base of the forming foot fuse during morphogenesis and are integrated into the caudal ganglion. This ganglion serves as the control hub for the posterior part of the trunk providing neuromuscular control of the pedal muscles and innervation of the foot. Bm-Hox3 is also involved in patterning of the pedal nerves. An expression domain in the distal part of the foot primordium gives rise to six cells with neural morphology in the trunk region connecting to the caudal ganglion. The position of the labeled pericarya is consistent with the six horns of the pedal muscles in Brachionus. Remarkably, Bm-Hox6 was recruited to pattern an additional functional domain of the nervous system, forming an asymmetrical nerve loop on the dorsal left side of the animal connecting the single germovitellarium to the caudal ganglion (Supplementary Fig. 6a). This observation indicates neuronal control of the germovitellarium by or via the caudal ganglion.
The rotiferan foot is a remarkable structure enabling transient attachment to surfaces via a glue-like secretion. Expression of Bm-Hox4 during foot formation marks cells in the proximal and central parts of the foot, giving rise to the caudal nerves and their connection to the caudal ganglion. Cells clustered at the tip of the foot have long been regarded to be simple gland cells21. Bm-Hox3, however, is strongly expressed in 12–14 cells connected to the caudal nerves and both Bm-Hox4 and Bm-MedPost exhibit overlapping expression in one or two cells at the base of the “toes”. In deuterostomes, posterior class Hox genes play key roles in patterning of the postanal tail46. In rotifers, posterior class genes are missing, leaving the key role of modeling of the nervous system of the postanal foot to central and MedPost class Hox genes. Judging from the innervation of this body region with FMRFamide- and serotonin (5HT)-positive nerve cells (Supplementary Fig. 6f, g) and expression of several Hox genes in the foot, we conclude that secretion of the “glue” is under neuronal control and the cells in the foot might represent an additional nerve plexus.
As most rotifer tissues are syncytial, and nerves consist of only a few interconnected neurons21, 22, code-like Hox expression is somewhat limited. This and newly evolved neuronal structures might have led to Hox gene regulation being adapted to modulate functional subsets of the rotifer nervous system (Fig. 3a). That the evolution of new and individual regulatory elements allowed the uncoupling of Hox expression from the constraints of collinearity is supported by the dispersed Hox cluster structure reported in the bdelloid rotifer Adineta vaga 45. However, the genomic structure of a monogonont rotifer Hox cluster has not yet been published. Surprisingly, Hox gene expression in Brachionus manjavacas does not seem to violate spatial colinearity completely as the patterns observed exhibit shifting anterior borders of expression to some extent. Temporal colinearity has been reported for some taxa4, a correlation of the temporal order of Hox gene activation during embryogenesis with the order of Hox genes in a cluster. The rather rapid morphogenesis in rotifers complicates determination of the order of Hox gene activation in this case. In B. manjavacas, transcription of the anterior Hox geneBm-Hox2 and Bm-Hox3 is indeed upregulated earlier (stage 1) than transcription of the other Hox genes (stage 2). The central class Hox genes analyzed are likely to be activated more or less simultaneously, but are all involved in patterning of the caudal ganglion and the nerves of the foot. Differing onsets of transcription may, therefore, be based on the order of morphological processes these genes are involved in rather than correlation with Hox cluster structure.
Discussion
Though not typically used for reconstructing phylogenies, analyses of Hox genes for diagnostic residues and conserved motifs give important phylogenetic clues. Patterning of animal body plans during ontogenesis is linked to the Hox gene complement in a very unique way. Major evolutionary changes of body plans have been accompanied and most probably have been made possible by changes of the Hox cluster structure6, 8, 59. Hox gene duplications enabled imprinting of additional positional information and thus evolution of additional body regions along the antero-posterior axes of animals. Reflected by possession of conserved amino-acid residues encoded in Hox genes these evolutionary changes and gene duplications seem to have happened independently in different clades. Hox genes play key roles in axial patterning and segment identity in many taxa. Our findings, however, suggest a different original role of Hox genes in metazoan evolution. In the diploblast metazoan Nematostella vectensis, Hox genes are predominantly expressed asymmetrically on one side of the body column of the polyp exhibiting slightly staggered epithelial expression patterns with large overlap34. Nematostella vectensis Hox genes are predominantly expressed in the endodermal layer. Neural expression has not been reported. Within triploblast metazoans, recent phylogenomic studies revealed a sister group relationship of Xenacoelomorpha (consisting of Acoela, Nemertodermatida, and Xenoturbella) and Nephrozoa (Deuterostomia and Protostomia)43, 44. Analyses of Hox gene expression in the acoel Convolutriloba longifissura indicate possible participation of acoel Hox genes in axial patterning of the nervous system due to subepidermal localization and coexpression with neural markers60. Consistent with morphological characteristics, phylogenomic analyses place Gnathifera near the base of Spiralia28–30. Rotifers possess a miniature but rather complex nervous system. Here Hox genes have been recruited to pattern the nervous system in a non-canonical way. Amazingly, rotiferan Hox genes show expression domains specifying functional subsets, with a strong bias in patterning of the caudal ganglion and the postanal foot as morphological novelties rather than exhibiting canonical Hox code expression along the anteroposterior axis. Unfortunately, Hox expression analysis has been reported for only a single chaetognath Hox gene61; however, expression of Spadella Hox4 in the ventral ganglion seems to be comparable to Bm-Hox4 expression in the caudal ganglion. These results might suggest an original role of Hox genes in neurogenesis. Consequently, their function in bauplan development would have been co-opted for other tissues during evolution.
Morphological or molecular ambiguities often lead to difficulties in phylogenetic placement of taxa. Several morphological traits strongly support Gnathifera. The support of a grouping of Rotifera and Chaetognatha based solely on either morphological traits (Fig. 3b) or developmental features may be weak; the combined analyses of morphology, development, and Hox sequences, however, provide an informative basis for this relationship. In addition, the newest and most comprehensive phylogenomic studies show some albeit moderate support of a close relationship of Gnathifera and Chaetognatha consistent with our results29. Though Hox gene information from Gnathostomulida and Micrognathozoa is currently not available, we expect these groups to show Hox characteristics consistent with this study. Exhibiting ambiguous characteristics indicating a possible close relationship to either Platyhelminthes or Gnathifera, placement of Gastrotricha has always been problematic. Phylogenomic approaches support both, a relationship with Platyhelminthes in Rouphozoa or a close relationship to Rotifera. Based on this study, we suggest an inclusion of chaetognaths in gnathiferans and Gnathifera as sister group to the remaining spiralians. The rather unusual expression of Hox genes in Brachionus manjavacas is additional evidence of this proposed phylogeny.
Methods
Collection of embryos
The rotifer Brachionus manjavacas (Florida Aqua Farms) was cultured in 15 ppm artificial sea water (ASW, Tropic Marin Classic) at 24 °C and fed Nannochloropsis microalgae culture in ASW (Florida Aqua Farms) twice a day. Animals were collected by sifting through a 50 µm nylon mesh and washed briefly with fresh artificial seawater. After being anesthetized in 0.5 mM Bupivacain in ASW for 12 min, animals and embryos were subjected to prefixation in 0.5, 1, 1.5, 2, 3% formaldehyde in PBS pH 7.4 for 2 min each followed by 3.7% for 10 min at room temperature (RT). For permeabilization of the egg shell, specimens were sonicated in glass test tubes for 40 s. Final fixation took place for 30 min at RT thereafter. Fixative was removed by washing 3–4 times with PTw (1× PBS pH 7.4, 0.1% Tween-20) for 5 min each, tissue was subsequently dehydrated by washing 3–4 times in methanol for 5 min each and stored at –32 °C until use.
In situ hybridization
Fixed rotifers and embryos were rehydrated briefly in a series of 75, 50, 25% methanol in PTw followed by four washes in PTw for 5 min each. Tissue was permeabilized by treatment with 0.01 mg ml−1 proteinase K in PTw for 10 min on a shaker. Digestion was stopped by two 5 min washes with 2 mg ml−1 glycine in PTw. After transfer to 1% triethanolamine (TEA) in PTw, specimens were subjected to two treatments with 0.3% acetic anhydride in 1% TEA for 5 min each. After brief washes in PTw the tissue was refixed in 3.7% formaldehyde in PTw for 30 min. Five washes with PTw were followed by a short preincubation in hybridization solution (HYBE: 50% formamide, 5 × SSC pH 4.5, 50 µg ml−1 heparin, 0.1% Tween-20, 1% SDS and 100 µg ml−1 salmon sperm DNA in diethyl pyrocarbonate (DEPC)-treated water) for 10 min at RT. Prehybridization in fresh hybridization solution was carried out over night at 65 °C. Tissues were hybridized with anti-sense riboprobes (1–3 ng µl−1) at 65 °C for 60 h. Subsequently tissues underwent post-hybridization by washing with HYBE twice for 10 and 20 min at 65 °C, followed for 10 min each in 75% HYBE and 25% 2 × SSC, 50% HYBE and 50% 2 × SSC, 25% HYBE and 75% 2 × SSC and 100% 2 × SSC at 65 °C. Two 30 min washes in 0.05 × SSC at 65 °C concluded posthybrization. Tissues were washed for 10 min each in 75% 0.05 × SSC and 25% PTw, 50% 0.05 × SSC and 50% PTw, 25% 0.05 × SSC and 75% PTw and 100% PTw. Blocking was performed by washing five times for 10 min in PBT (1 × PBS, pH 7.4, 0.2 % Triton X-100, 0.1 % bovine serum albumin) and 1 h in 1 × blocking buffer (Roche) in maleic acid buffer (100 mM maleic acid, 150 mM NaCl, pH 7.5) at RT. For detection of the riboprobes tissues were incubated in anti-digoxygenin-AP Fab fragments (Roche) diluted 1:5000 in blocking buffer for 16 h over night at 6 °C on a shaker followed by ten washes for 10 min in PBT at RT. Expression patterns were visualized by three washes in AP buffer (100 mM NaCl, 50 mM MgCl2, 100 mM Tris pH 9.5, 0.5% Tween-20) and detection with NBT/BCIP in AP-buffer as substrate. Specimens were analyzed using differential interference contrast optics on an Olympus BX-51 microscope. Digital photomicrographs were captured with a Nikon Coolpix 4500 digital camera (4.0 megapixel).
Cloning of Brachionus manjavacasHox genes
Initially, small fragments of the homeodomain region of Brachionus manjavacas Hox genes were amplified by degenerate primer PCR. Different primer sets more or less specific for Hox genes in general or Hox genes belonging to specific PGs in particular (Supplementary Table 1) were used to isolate Hox gene fragments from either mixed stage complementary DNA (cDNA) or genomic DNA (gDNA). In the latter case, presence of introns within the homeodomain was taken into account. gDNA was isolated with an ArchivePure gDNA kit (5prime), cDNA was obtained by RNA isolation (RNeasy kit, Qiagen) followed by reverse transcription (RevertAid First Strand cDNA Synthesis Kit, Thermo Fisher Scientific). Genes were preliminarily identified by BLASTX search (NCBI). Large fragments of cDNAs suitable for phylogenetic analysis and riboprobe synthesis were obtained by RACE (rapid amplification of cDNA ends) with gene specific primers using the SmartRACE Kit (Clontech). All fragments were cloned into pGEM-Teasy vector (Promega) and sequenced at Macrogen Inc (South Korea) or StarSeq (Germany).
Riboprobe synthesis
Digoxigenin-labeled riboprobes were generated by in vitro transcription using MEGAscript High Yield SP6 or T7 transcription kits (Ambion) with PCR products of suitables clones flanked by SP6- or T7 RNA polymerase promotor sites as templates.
Orthology assignment and phylogenetic analyses
Assignment to paralog groups (PG) was based on the phylogenetic analyses as well as the presence or absence of diagnostic amino-acid residues or motifs in homeodomain or flanking region of Hox genes, commonly regarded as apomorphies for specific PGs and even specific taxonomic groups.
For phylogenetic analyses sequences including the homeodomain and 12 amino acids of the carboxy flanking region next to the homeodomain were aligned using MacVector 8.0. Genome accession numbers of Adineta vaga Hox genes and Genbank accession numbers of all other Hox gene sequences used in phylogenetic analyses are given in Supplementary Table 2. The most suitable amino-acid substitution model LG + Γ + I was determined by ProtTest 3.462. Bayesian phylogenetic analyses were conducted with MrBayes V3.2.663, 64 on the tera-grid accessible via the CIPRES science gateway V3.365. LG with invgamma was selected with 100% posterior probability with four independent runs of 10,000,000 generations sampled every 100 generations and four chains each. A summary tree was generated from the final 300,000 trees. ML bootstrap analysis was conducted with RAxML-HPC v8.2.966 on XSEDE via the CIPRES science gateway V3.3 with 1000 iterations using the LG + Γ + I model of protein evolution. Final trees were drawn using Figtree 1.4.2 (http://tree.bio.ed.ac.uk/software/figtree/) and CorelDraw 12. Nexus alignments are available upon request.
Data availability
Additional data associated with this study are available in the Supplementary Information of this publication. Assembled sequences for all Hox genes isolated from Brachionus manjavacas have been deposited with GenBank under accession numbers KT989538 (Bm-Hox2), KT989539 (Bm-Hox3), KT989540 (Bm-Hox4), KT989541 (Bm-Hox6), and KT989542 (Bm-MedPost). The amino-acid sequence of Bm-Hox5 is available in the Figshare Repository under the identifier 10.6084/m9.figshare.4616125.
Electronic supplementary material
Supplementary Figures, Supplementary Tables and Supplementary References
Acknowledgements
We wish to thank David S. Richardson for valuable comments on the manuscript. We gratefully acknowledge Andreas Vilcinskas and Adriaan W.C. Dorresteijn for support of the project.
Author contributions
P.F. initially conceived the project. A.C.F. oversaw the project and performed acquisition, analysis, and interpretation of the data including cloning of the Hox genes, staging of morphogenesis of Brachionus, analysis of gene expressing patterns and phylogenetic analyses. A.C.F. wrote the first draft of the paper and generated the figures. P.F. made substantial contributions to revisions of the draft of the article. Both authors discussed the results and commented on the manuscript.
Competing interests
The authors declare no competing financial interests.
Footnotes
Electronic supplementary material
Supplementary Information accompanies this paper at doi:10.1038/s41467-017-00020-w.
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Lewis EB. A gene complex controlling segmentation in Drosophila. Nature. 1978;276:565–570. doi: 10.1038/276565a0. [DOI] [PubMed] [Google Scholar]
- 2.Dessain S, Gross CT, Kuziora MA, McGinnis W. Antp-type homeodomains have distinct DNA binding specificities that correlate with their different regulatory functions in embryos. EMBO. J. 1992;11:991–1002. doi: 10.1002/j.1460-2075.1992.tb05138.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Averof M, Akam M. Hox genes and the diversification of insect and crustacean body plans. Nature. 1995;376:420–423. doi: 10.1038/376420a0. [DOI] [PubMed] [Google Scholar]
- 4.Fröbius AC, Matus DQ, Seaver EC. Genomic organization and expression demonstrate spatial and temporal Hox gene colinearity in the lophotrochozoan Capitella sp. I. PLoS. ONE. 2008;3:e4004. doi: 10.1371/journal.pone.0004004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kourakis MJ, Martindale MQ. Hox gene duplication and deployment in the annelid leech Helobdella. Evol. Dev. 2001;3:145–153. doi: 10.1046/j.1525-142x.2001.003003145.x. [DOI] [PubMed] [Google Scholar]
- 6.Burke AC, Nelson CE, Morgan BA, Tabin C. Hox genes and the evolution of vertebrate axial morphology. Development. 1995;121:333–346. doi: 10.1242/dev.121.2.333. [DOI] [PubMed] [Google Scholar]
- 7.Carroll SB. Homeotic genes and the evolution of arthropods and chordates. Nature. 1995;376:479–485. doi: 10.1038/376479a0. [DOI] [PubMed] [Google Scholar]
- 8.Averof M, Patel NH. Crustacean appendage evolution associated with changes in Hox gene expression. Nature. 1997;388:682–686. doi: 10.1038/41786. [DOI] [PubMed] [Google Scholar]
- 9.Boncinelli E, Simeone A, Acampora D, Gulisano M. Homeobox genes in the developing central nervous system. Ann. Genet-Paris. 1993;36:30–37. [PubMed] [Google Scholar]
- 10.Cameron RA, et al. Unusual gene order and organization of the sea urchin hox cluster. J. Exp. Zool. B. 2006;306:45–58. doi: 10.1002/jez.b.21070. [DOI] [PubMed] [Google Scholar]
- 11.Pierce RJ, et al. Evidence for a dispersed Hox gene cluster in the platyhelminth parasite Schistosoma mansoni. Mol. Biol. Evol. 2005;22:2491–2503. doi: 10.1093/molbev/msi239. [DOI] [PubMed] [Google Scholar]
- 12.Edvardsen RB, et al. Remodelling of the homeobox gene complement in the tunicate Oikopleura dioica. Curr. Biol. 2005;15:R12–R13. doi: 10.1016/j.cub.2004.12.010. [DOI] [PubMed] [Google Scholar]
- 13.Struhl G. Splitting the bithorax complex of Drosophila. Nature. 1984;308:454–457. doi: 10.1038/308454a0. [DOI] [Google Scholar]
- 14.Von Allmen G, et al. Splits in fruitfly Hox gene complexes. Nature. 1996;380:116. doi: 10.1038/380116a0. [DOI] [PubMed] [Google Scholar]
- 15.Kristensen RM, Funch P. Micrognathozoa: a new class with complicated jaws like those of Rotifera and Gnathostomulida. J. Morphol. 2000;246:1–49. doi: 10.1002/1097-4687(200010)246:1<1::AID-JMOR1>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 16.Funch P, Sørensen MV, Obst M. On the phylogenetic position of Rotifera - have we come any further? Hydrobiologia. 2005;546:11–28. doi: 10.1007/s10750-005-4093-6. [DOI] [Google Scholar]
- 17.Sørensen MV. Further structures in the jaw apparatus of Limnognathia maerski (Micrognathozoa), with notes on the phylogeny of the Gnathifera. J. Morphol. 2003;255:131–145. doi: 10.1002/jmor.10038. [DOI] [PubMed] [Google Scholar]
- 18.Jennings HS. The early development of Asplanchna herrickii de Guerne. A contribution to developmental mechanics. Bull. Mus. Comp. Zool. Harv. 1896;30:1–117. [Google Scholar]
- 19.Lechner M. Untersuchungen zur Embryonalentwicklung des Rädertieres Asplanchna girodi de Guerne. Roux’s Arch. Entwicklungsmech. Org. 1966;157:117–173. doi: 10.1007/BF00577929. [DOI] [PubMed] [Google Scholar]
- 20.Nachtwey R. Untersuchungen über die Keimbahn, Organogenese und Anatomie von Asplanchna priodonta Gosse. Z. Wiss. Zool. 1925;126:239–492. [Google Scholar]
- 21.Clément, P. & Wurdak, E in Microscopic Anatomy of Invertebrates Vol. 4 (eds Harrison, F. W. & Ruppert, E. E.) 219-297 (Wiley-Liss, 1991).
- 22.Clément P, Amsellem J. The skeletal-muscles of rotifers and their innervation. Hydrobiologia. 1989;186:255–278. doi: 10.1007/BF00048921. [DOI] [Google Scholar]
- 23.Dunn CW, et al. Broad phylogenomic sampling improves resolution of the animal tree of life. Nature. 2008;452:745–749. doi: 10.1038/nature06614. [DOI] [PubMed] [Google Scholar]
- 24.Hejnol A, et al. Assessing the root of bilaterian animals with scalable phylogenomic methods. Proc. R. Soc. B. 2009;276:4261–4270. doi: 10.1098/rspb.2009.0896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Philippe H, et al. Acoelomorph flatworms are deuterostomes related to Xenoturbella. Nature. 2011;470:255–258. doi: 10.1038/nature09676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Giribet G, Distel DL, Polz M, Sterrer W, Wheeler WC. Triploblastic relationships with emphasis on the acoelomates and the position of Gnathostomulida, Cycliophora, Plathelminthes, and Chaetognatha: a combined approach of 18S rDNA sequences and morphology. Syst. Biol. 2000;49:539–562. doi: 10.1080/10635159950127385. [DOI] [PubMed] [Google Scholar]
- 27.Cavalier-Smith T. A revised six-kingdom system of life. Biol. Rev. Camb. Philos. Soc. 1998;73:203–266. doi: 10.1017/S0006323198005167. [DOI] [PubMed] [Google Scholar]
- 28.Laumer CE, et al. Spiralian phylogeny informs the evolution of microscopic lineages. Curr. Biol. 2015;25:2000–2006. doi: 10.1016/j.cub.2015.06.068. [DOI] [PubMed] [Google Scholar]
- 29.Kocot, K. M. et al. Phylogenomics of Lophotrochozoa with consideration of systematic error. Syst. Biol. doi:10.1093/sysbio/syw079 (2016). [DOI] [PubMed]
- 30.Struck TH, et al. Platyzoan paraphyly based on phylogenomic data supports a noncoelomate ancestry of Spiralia. Mol. Biol. Evol. 2014;31:1833–1849. doi: 10.1093/molbev/msu143. [DOI] [PubMed] [Google Scholar]
- 31.de Rosa R, et al. Hox genes in brachiopods and priapulids and protostome evolution. Nature. 1999;399:772–776. doi: 10.1038/21631. [DOI] [PubMed] [Google Scholar]
- 32.Telford MJ. Turning Hox “signatures” into synapomorphies. Evol. Dev. 2000;2:360–364. doi: 10.1046/j.1525-142x.2000.00075.x. [DOI] [PubMed] [Google Scholar]
- 33.Balavoine G, de Rosa R, Adoutte A. Hox clusters and bilaterian phylogeny. Mol. Phylogenet. Evol. 2002;24:366–373. doi: 10.1016/S1055-7903(02)00237-3. [DOI] [PubMed] [Google Scholar]
- 34.Ryan JF, et al. Pre-bilaterian origins of the Hox cluster and the Hox code: evidence from the sea anemone, Nematostella vectensis. PLoS ONE. 2007;2:e153. doi: 10.1371/journal.pone.0000153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Matus DQ, Halanych KM, Martindale MQ. The Hox gene complement of a pelagic chaetognath, Flaccisagitta enflata. Integr. Comp. Biol. 2007;47:854–864. doi: 10.1093/icb/icm077. [DOI] [PubMed] [Google Scholar]
- 36.Papillon D, Perez Y, Fasano L, Le Parco Y, Caubit X. Hox gene survey in the chaetognath Spadella cephaloptera: evolutionary implications. Dev. Genes. Evol. 2003;213:142–148. doi: 10.1007/s00427-003-0306-z. [DOI] [PubMed] [Google Scholar]
- 37.Passamaneck YJ, Halanych KM. Evidence from Hox genes that bryozoans are lophotrochozoans. Evol. Dev. 2004;6:275–281. doi: 10.1111/j.1525-142X.2004.04032.x. [DOI] [PubMed] [Google Scholar]
- 38.Bleidorn C, Lanterbecq D, Eeckhaut I, Tiedemann R. A PCR survey of Hox genes in the myzostomid Myzostoma cirriferum. Dev. Genes. Evol. 2009;219:211–216. doi: 10.1007/s00427-009-0282-z. [DOI] [PubMed] [Google Scholar]
- 39.Orii H, et al. The planarian HOM HOX homeobox genes (Plox) expressed along the anteroposterior axis. Dev. Biol. 1999;210:456–468. doi: 10.1006/dbio.1999.9275. [DOI] [PubMed] [Google Scholar]
- 40.Fritzsch G, et al. PCR survey of Xenoturbella bocki Hox genes. J. Exp. Zool. B, Mol. Develop. evol. 2008;310:278–284. doi: 10.1002/jez.b.21208. [DOI] [PubMed] [Google Scholar]
- 41.Jimenez-Guri E, Paps J, Garcia-Fernandez J, Salo E. Hox and ParaHox genes in Nemertodermatida, a basal bilaterian clade. Int. J. Dev. Biol. 2006;50:675–679. doi: 10.1387/ijdb.062167ej. [DOI] [PubMed] [Google Scholar]
- 42.Cook CE, Jimenez E, Akam M, Salo E. The Hox gene complement of acoel flatworms, a basal bilaterian clade. Evol. Dev. 2004;6:154–163. doi: 10.1111/j.1525-142X.2004.04020.x. [DOI] [PubMed] [Google Scholar]
- 43.Cannon JT, et al. Xenacoelomorpha is the sister group to Nephrozoa. Nature. 2016;530:89–93. doi: 10.1038/nature16520. [DOI] [PubMed] [Google Scholar]
- 44.Rouse GW, Wilson NG, Carvajal JI, Vrijenhoek RC. New deep-sea species of Xenoturbella and the position of Xenacoelomorpha. Nature. 2016;530:94–97. doi: 10.1038/nature16545. [DOI] [PubMed] [Google Scholar]
- 45.Flot JF, et al. Genomic evidence for ameiotic evolution in the bdelloid rotifer Adineta vaga. Nature. 2013;500:453–457. doi: 10.1038/nature12326. [DOI] [PubMed] [Google Scholar]
- 46.Ferrier DE, Minguillon C, Holland PW, Garcia-Fernandez J. The amphioxus Hox cluster: deuterostome posterior flexibility and Hox14. Evol. Dev. 2000;2:284–293. doi: 10.1046/j.1525-142x.2000.00070.x. [DOI] [PubMed] [Google Scholar]
- 47.Marletaz F, et al. Chaetognath transcriptome reveals ancestral and unique features among bilaterians. Genome Biol. 2008;9:R94. doi: 10.1186/gb-2008-9-6-r94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ahlrichs WH. Epidermal ultrastructure of Seison nebaliae and Seison annulatus, and a comparison of epidermal structures within the Gnathifera. Zoomorphology. 1997;117:41–48. doi: 10.1007/s004350050028. [DOI] [Google Scholar]
- 49.Ahlrichs, W. H. Ultrastruktur und Phylogenie von Seison nebaliae (Grube 1859) und Seison annulatus (Claus 1876) (Dissertation, Georg-August-University, Göttingen, Cuvillier Verlag, 1995).
- 50.Kristensen RM, Nørrevang A. On the fine structure of Rastrognathia macrostoma gen. et sp. n. placed in Rastrognathiidae fam. n. (Gnathostomulida) Zool. Scr. 1977;6:27–41. doi: 10.1111/j.1463-6409.1977.tb00757.x. [DOI] [Google Scholar]
- 51.Müller MCM, Sterrer W. Musculature and nervous system of Gnathostomula peregrina (Gnathostomulida) shown by phalloidin labeling, immunohistochemistry, and cLSM, and their phylogenetic significance. Zoomorphology. 2004;123:169–177. [Google Scholar]
- 52.Extavour CG. Evolution of the bilaterian germ line: lineage origin and modulation of specification mechanisms. Integr. Comp. Biol. 2007;47:770–785. doi: 10.1093/icb/icm027. [DOI] [PubMed] [Google Scholar]
- 53.Riedl RJ. Gnathostomulida from America. Science. 1969;163:445–452. doi: 10.1126/science.163.3866.445. [DOI] [PubMed] [Google Scholar]
- 54.Shimotori T, Goto T. Establishment of axial properties in the arrow worm embryo, Paraspadella gotoi (Chaetognatha): Developmental fate of the first two blastomeres. Zool. Sci. 1999;16:459–469. doi: 10.2108/zsj.16.459. [DOI] [Google Scholar]
- 55.Shimotori T, Goto T. Developmental fates of the first four blastomeres of the chaetognath Paraspadella gotoi: Relationship to protostomes. Dev. Growth Differ. 2001;43:371–382. doi: 10.1046/j.1440-169x.2001.00583.x. [DOI] [PubMed] [Google Scholar]
- 56.Schmidt, G. D. in Biology of the Acanthocephala (eds Crompton, D.W.T. & Nickol B. B.) 273–305 (Cambridge University Press, 1985).
- 57.Matus DQ, et al. Broad taxon and gene sampling indicate that chaetognaths are protostomes. Curr. Biol. CB. 2006;16:R575–R576. doi: 10.1016/j.cub.2006.07.017. [DOI] [PubMed] [Google Scholar]
- 58.Marletaz F, et al. Chaetognath phylogenomics: a protostome with deuterostome-like development. Curr. Biol. CB. 2006;16:R577–R578. doi: 10.1016/j.cub.2006.07.016. [DOI] [PubMed] [Google Scholar]
- 59.Lee PN, Callaerts P, De Couet HG, Martindale MQ. Cephalopod Hox genes and the origin of morphological novelties. Nature. 2003;424:1061–1065. doi: 10.1038/nature01872. [DOI] [PubMed] [Google Scholar]
- 60.Hejnol A, Martindale MQ. Coordinated spatial and temporal expression of Hox genes during embryogenesis in the acoel Convolutriloba longifissura. BMC. Biol. 2009;7:65. doi: 10.1186/1741-7007-7-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Papillon D, Perez Y, Fasano L, Le Parco Y, Caubit X. Restricted expression of a median Hox gene in the central nervous system of chaetognaths. Dev. Genes Evol. 2005;215:369–373. doi: 10.1007/s00427-005-0483-z. [DOI] [PubMed] [Google Scholar]
- 62.Darriba D, Taboada GL, Doallo R, Posada D. ProtTest 3: fast selection of best-fit models of protein evolution. Bioinformatics. 2011;27:1164–1165. doi: 10.1093/bioinformatics/btr088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–1574. doi: 10.1093/bioinformatics/btg180. [DOI] [PubMed] [Google Scholar]
- 64.Huelsenbeck JP, Ronquist F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics. 2001;17:754–755. doi: 10.1093/bioinformatics/17.8.754. [DOI] [PubMed] [Google Scholar]
- 65.Miller, M. A., Pfeiffer, W. & Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. in Proceedings of the Gateway Computing Environments Workshop (GCE). New Orleans, LA, 1–8 (2010).
- 66.Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–1313. doi: 10.1093/bioinformatics/btu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figures, Supplementary Tables and Supplementary References
Data Availability Statement
Additional data associated with this study are available in the Supplementary Information of this publication. Assembled sequences for all Hox genes isolated from Brachionus manjavacas have been deposited with GenBank under accession numbers KT989538 (Bm-Hox2), KT989539 (Bm-Hox3), KT989540 (Bm-Hox4), KT989541 (Bm-Hox6), and KT989542 (Bm-MedPost). The amino-acid sequence of Bm-Hox5 is available in the Figshare Repository under the identifier 10.6084/m9.figshare.4616125.