

Abstract
Chronic pancreatitis (CP) is a fibro-inflammatory disease leading to pain, maldigestion, and pancreatic insufficiency. No therapeutic options exist due to a limited understanding of the biology of CP pathology. Recent findings implicate pancreatic stellate cells (PSC) as prominent mediators of inflammatory and fibrotic processes during CP. Here, we utilized primary and immortalized PSC obtained from mice and patients with CP or pancreatic cancer to examine the effect of Jak/STAT and MAPK pathway inhibition in vitro. The well-characterized caerulein model of CP was used to assess the therapeutic efficacy of Jak1/2 inhibition in vivo. Treatment of cultured PSC with the Jak1/2 inhibitor ruxolitinib reduced STAT3 phosphorylation, cell proliferation, and expression of alpha-smooth muscle actin (α-SMA), a marker of PSC activation. Treatment with the MAPK inhibitor, MEK162, had less consistent effects on PSC proliferation and no impact on activation. In the caerulein-induced murine model of CP, administration of ruxolitinib for one week significantly reduced biomarkers of inflammation and fibrosis. These data suggest that the Jak/STAT pathway plays a prominent role in PSC proliferation and activation. In vivo treatment with the Jak1/2 inhibitor ruxolitinib reduced the severity of experimental CP, suggesting that targeting Jak/STAT signaling may represent a promising therapeutic strategy for CP.
Introduction
Chronic pancreatitis (CP) is characterized by persistent inflammation and fibrosis of the pancreas. Patients with CP often experience recurrent abdominal pain, nausea, and maldigestion that progress to exocrine insufficiency, fat-soluble vitamin deficiency, metabolic bone disease, and diabetes mellitus1–6. Depending on etiology, CP patients also have an approximate 3–5-fold increased risk of developing pancreas cancer7–12. To date, no clinical therapy is available to reverse the inflammatory damage associated with CP. Instead, management is focused on treatment of associated symptoms and complications. Thus, identifying novel interventions for this disease represents a high priority and would fill an unmet medical need to improve quality of life, reduce risk of malignancy, and limit medical costs associated with long-term care of these patients13, 14.
The fibro-inflammatory response associated with CP is hypothesized to result from premature activation of pancreatic enzymes within the gland, resulting in auto-digestion of parenchyma. Subsequent acute inflammatory events result in release of pro-inflammatory mediators that promote both immune cell infiltration and activation of local fibroblasts termed pancreatic stellate cells (PSC). Once active, PSC promote inflammation and fibrosis through secretion of cytokines and chemokines as well as matrix metalloproteinases (MMPs), tissue inhibitor of metalloproteinases (TIMPs), and collagen15–17. Transient PSC activation occurs during instances of acute pancreatic inflammation, however the onset of CP is characterized by PSC that display a constitutively active phenotype to promote a state of chronic fibro-inflammation18, 19. The proportion of patients with acute pancreatitis (AP) that will progress to CP varies by etiology. Specifically, the development of CP is more common among those with acute alcoholic pancreatitis compared to acute gallstone pancreatitis. This difference may be due, in part, to the reduced viability of PSC following exposure to bile acids during acute gallstone pancreatitis20. This suggests the importance of PSC activity to the transition from acute inflammation to CP. Activated PSC are also observed in pancreatic ductal adenocarcinoma (PDAC), where they support growth and invasiveness of tumors21–23. Accordingly, PSC may represent a viable therapeutic target in the context of CP to reduce inflammation, fibrosis, and risk of malignancy.
PSC secrete high levels of several immunomodulatory factors, including interleukin-6 (IL-6), tumor necrosis factor alpha (TNFα), monocyte chemoattractant protein-1 (MCP-1), and others24. Many of these factors act in an autocrine and paracrine manner to orchestrate continued PSC survival, cellular activation, and proliferation while driving fibro-inflammatory processes that contribute to CP pathology25–28. IL-6 and other cytokines exert their effects via the transmembrane receptor gp130 to activate Jak/STAT signaling, notably Jak2/STAT3. Once activated, STAT3 positively regulates several pro-survival and pro-inflammatory gene signatures. The Jak/STAT pathway also cross-talks with other signal transduction pathways including MAPK and NF-kB to amplify expression of inflammatory genes29–32. Data from animal models and human patients suggest that IL-6 signaling is of particular importance in the context of CP. In murine models of disease, genetic ablation of IL-6 reduces susceptibility to caerulein-induced pancreatitis and associated lung injury33. Serum levels of IL-6 are also often found to be elevated in human CP patients34–37. Although acquisition of human pancreatic tissue across the spectrum of CP disease stages is not feasible, several studies have explored the role of this pathway in the context of PDAC. IHC analysis of human PDAC tumors revealed robust staining of IL-6 localized to the stromal compartment, which includes PSC, immune cells, and others38. Furthermore, murine models of PDAC have demonstrated cooperation between STAT3 signaling and activated KRas within the pancreas to drive cancer progression39. Thus, stromal-derived IL-6/Jak2/STAT3 signaling appears to play a prominent role in PSC activity, CP pathology, and PDAC development.
To our knowledge there are currently no clinical trials and only limited in vitro or in vivo studies targeting soluble IL-6 or the Jak/STAT pathway in the context of CP40. Although development of clinically suitable STAT3 inhibitors is lacking, considerable advances have been made in the development of small molecule inhibitors of the upstream Jak proteins41, 42. These agents are well tolerated by patients and are FDA-approved for treatment of other inflammatory disorders including rheumatoid arthritis, myelofibrosis and polycythemia vera43–45. However, Jak inhibitors have never been formally tested in patients with CP.
We sought to characterize the activation of pro-inflammatory STAT3 and MAPK pathways in PSC from both CP and PDAC, and to assess the ability of targeted inhibition to limit pathologic PSC activity. We hypothesized that inhibition of Jak/STAT or MAPK signaling would reduce PSC activity and limit the severity of caerulein-induced CP. Our results demonstrate that both the STAT3 and MAPK pathways are activated in cultured mouse and human PSC from the setting of CP and PDAC. Inhibition of Jak1/2 resulted in decreased proliferation of PSC, which was associated with diminished cellular activation. The impact of MEK inhibition was more variable, depending on the cell culture assayed. In a proof-of-concept study using the caerulein-induced murine model of CP, short-term treatment with ruxolitinib, a Jak1/2 inhibitor, led to partial resoration of serum lipase levels and reduced acinar cell loss and fibrosis. These findings suggest that Jak/STAT inhibition may limit the pathology observed in caerulein-induced pancreatitis.
Results
α-SMA+ PSC display STAT3 and MAPK signaling and secrete immunomodulatory factors
PSC lines (Table 1) were purchased or PSC cell cultures were isolated from mouse or human pancreatic tissue as described and were characterized24. In culture, PSC exhibited a pseudoquiescent phenotype when incubated with 10 μM all-trans retinoic acid (ATRA), as evidenced by intracellular Oil-Red O positive lipid droplets (Fig. 1a ). Under normal culture conditions, PSC typically assume an activated state, and are thus negative for Oil-Red O staining, while positive for α-SMA, a marker of stellate cell activation (Fig. 1b,c ). These active PSC were evident in pancreata from the caerulein-induced murine model of CP and increased with disease severity (Fig. 1d,e ). Lysates prepared from activated PSC grown to 70–80% confluence demonstrated constitutive phosphorylation of STAT3 and ERK proteins in all cell lines tested, indicating activation of the pro-inflammatory, pro-survival STAT3 and MAPK pathways (Fig. 2a,b ). Supernatants from these cells further demonstrated elevated levels of several immunomodulatory factors, including IL-6, MCP-1, and CXCL10, as compared to a human pancreas-derived fibroblast line (HPF), which served as a control (Fig. 2c,d).
Table 1.
Human and murine pancreatic stellate cell lines utilized in vitro. Details, including cell type, species of origin, immortalization status, and disease of origin, are provided for each cell line (PDAC indicates pancreatic ductal adenocarcinoma, GEMM indicates genetically engineered mouse model).
| # | Name | Cell Type | Species | Immortalization Status | Disease of origin/Genotype |
|---|---|---|---|---|---|
| 1 | HPF | fetal pancreatic fibroblast | human | non-immortalized | Normal |
| 2 | PaSC | PSC | mouse | immortalized | Normal |
| 3 | m-PSC-PDAC-1 | PSC | mouse | non-immortalized | PDAC GEMM: Mist1(KRAS G12D/+) |
| 4 | m-PSC-PDAC-2 | PSC | mouse | non-immortalized | PDAC GEMM: Mist1(KRAS G12D/+) |
| 5 | m-PSC-PDAC-3 | PSC | mouse | non-immortalized | PDAC GEMM: (Brca1 flox2/flox2; Kras LSL-G12D/+; p53 LSL-R27OH/+; Pdx1-cre) |
| 6 | h-PSC-PDAC-1 | PSC | human | non-immortalized | PDAC |
| 7 | h-PSC-PDAC-2 | PSC | human | non-immortalized | PDAC |
| 8 | h-iPSC-PDAC-1 | PSC | human | immortalized | PDAC |
| 9 | h-PSC-CP-1 | PSC | human | non-immortalized | chronic pancreatitis |
| 10 | RLT-PSC | PSC | human | immortalized | chronic pancreatitis |
Figure 1.
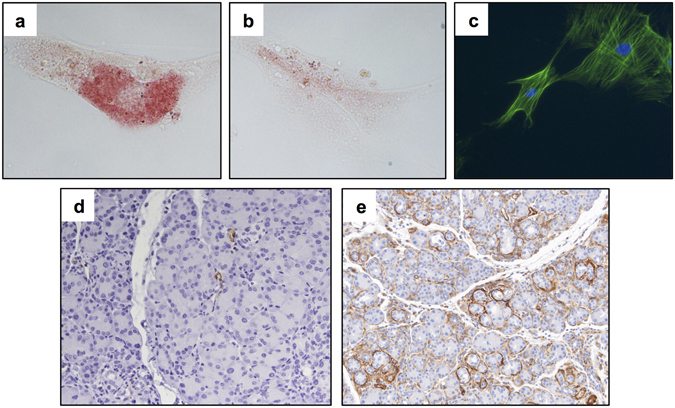
PSC display an activated phenotype in culture and in a murine model of CP. PaSC were treated with (a) 10 μM all-trans retinoic acid (ATRA) or (b) vehicle control for 48 hours and stained for Oil-Red O. Cells were analyzed by light microscopy at 40X magnification. (c) Untreated PaSC were stained for α-SMA (green) by fluorescent microscopy following 48 hours of incubation (DAPI counterstain, 40X magnification). (d) Formalin fixed paraffin embedded (FFPE) pancreatic tissue from mice with caerulein-induced pancreatitis after (d) 1 week and (e) 5 weeks of treatment were stained for α-SMA (20X magnification). Representative images from n = 5 mice per group.
Figure 2.
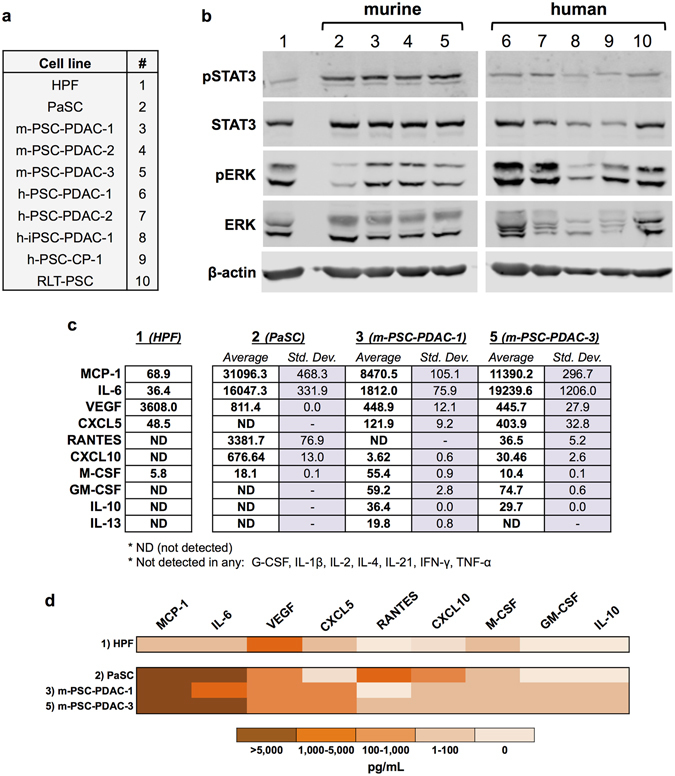
PSC display constitutive activation of STAT3 and MAPK signaling and secrete pro-inflammatory factors. (a) Summary of all PSC and PSC cell cultures used. (b) Cells were grown in DMEM and lysed once they reached 70–80% confluence for western blot analysis. pSTAT3, STAT3, pERK, and ERK were analyzed by immunoblot. β-actin served as a loading control. (c) Supernatants from 3 murine PSC cell lines and the human control pancreas-derived fibroblast line (HFP) (detailed in Table 1) were collected from 70% confluent cells grown in DMEM. A panel of cytokines and chemokines was analyzed in these supernatants by Luminex assay. Values are listed as an average of two replicates in pg/mL with the standard deviation. ND indicates analytes were assayed, but the levels were detected beneath the lowest standard curve reference value. (d) Selected results from the Luminex assay, presented as a heat map.
Effects of the Jak1/2 inhibitor, ruxolitinib, and the MEK inhibitor, MEK162, on proliferation of PSC in vitro
Because the inflammatory, pro-survival Jak-STAT and MAPK pathways were activated in PSC, we investigated the effects of inhibiting these pathways using small molecule kinase inhibitors. Treatment of representative murine (PaSC) and human (h-iPSC-PDAC-1) PSC with the Jak1/2 inhibitor ruxolitinib reduced STAT3 phosphorylation and decreased cell proliferation (Fig. 3a,b). The absence of PARP cleavage, as assessed by immunoblot, and retention of cell adherence observed by light microscopy, suggest that these cells did not undergo apoptosis in response to Jak1/2 inhibition. Treatment of cells with the MEK inhibitor MEK162 produced more variable results. For instance, while treatment of murine PaSC cells with MEK162 did not alter cell proliferation, as assessed by MTT and light microscopy (Fig. 3c), there was a modest but statistically significant reduction in proliferation of human PSC (Fig. 3d) in response to MEK162. Notably, a trend toward increased activation of the STAT3 pathway was observed following treatment with MEK162, although this did not reach statistical significance. This compensatory survival mechanism has been previously described in pancreatic and color cancer cell lines46. Similarly, increased MAPK pathway activation was seen following treatment with the Jak1/2 inhibitor ruxolitinib in PaSC cells (Fig. 3a).
Figure 3.
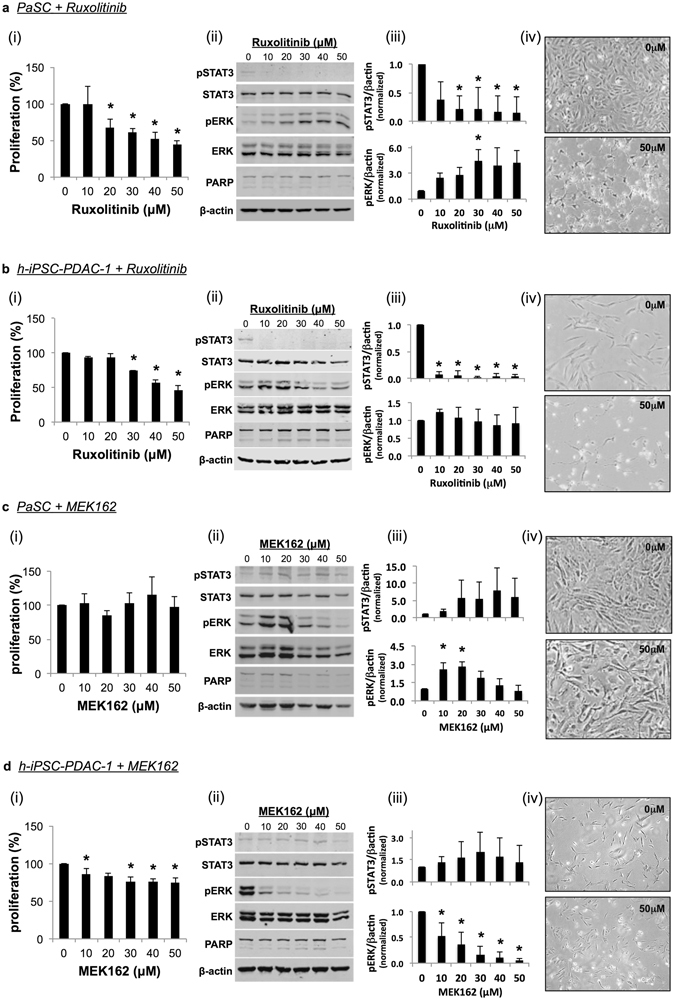
Effect of Jak/STAT and MEK inhibition on PSC proliferation in vitro. (a,c) PaSC or (b,d) h-iPSC-PDAC-1 were treated with ruxolitinib (a,b) or MEK162 (c,d). (i) After 24 hours of incubation, cell proliferation was analyzed by MTT assay. (ii) Lysates were also taken at 24 hours and analyzed by western blot. β-actin served as a loading control. (iii) Results were quantified via densitometry and normalized to β-actin. Graphs display mean ± STD from 3 biological replicates (* indicates p < 0.05). (iv) Light microscopy images were taken of treated cells following 72 hours of incubation (40X magnification).
Treatment with ruxolitinib, but not MEK162, reduces PSC activation in vitro
Because PSC display reduced cellular proliferation without apparent cell death in response to ruxolitinib, we examined the effect of Jak1/2 or MEK inhibition on biomarkers of cellular activation. By immunoblot, PaSC treated with ruxolitinib exhibited decreased α-SMA expression (Fig. 4a). This trend was not observed in cells treated with MEK162. To confirm these results, PaSC were stained with Oil-Red O, a marker of PSC quiescence or pseudoquiescence, following treatment. Light microscopy results revealed Oil-Red O positive lipid droplets in ATRA (positive control) and ruxolitinib-treated cells. In contrast, untreated and MEK162-treated cells remained Oil-Red O negative (Fig. 4b,c). Taken together, these phenotypic profiles indicate that ruxolitinib treatment reduced PSC activation in vitro.
Figure 4.
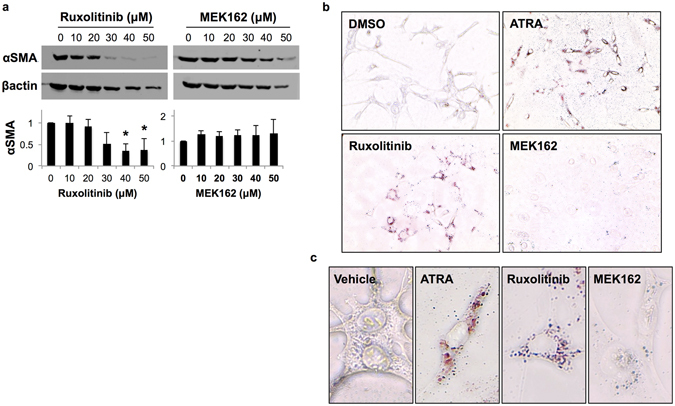
Jak/STAT inhibition reduces PSC activation in vitro. (a) Lysates of PaSC were taken after 72 hours of treatment with either ruxolitinib or MEK162. α-SMA expression was assessed by immunoblot and quantified by densitometric analysis. Graphs display mean ± SD of 3 biological replicates (* indicates p < 0.05). (b,c) PaSC were stained with Oil-Red O to examine quiescence or pseudoquiescence following incubation with vehicle, 10 μM ATRA (positive control), 10 μM ruxolitinib, or 10 μM MEK162 for 48 hours. (b) Light microscopy images were taken at 20X magnification. (c) Increased magnification of 20X images are shown below at 40X.
In vivo treatment with ruxolitinib reduces disease severity in a murine model of chronic pancreatitis
To examine the impact of ruxolitinib on biomarkers relevant to CP in vivo, we utilized the well-characterized murine model of caerulein-induced chronic pancreatitis (Fig. 5a). In this proof of concept study, oral administration of ruxolitinib for one week in mice with established pancreatitis led to reduced pSTAT3 in the pancreata as determined by IHC (Fig. 5b). This short-term treatment also resulted in a trend toward restored serum lipase levels (Fig. 5c). By IHC analysis, formalin-fixed pancreata showed significantly reduced acinar cell loss and fibrosis. Assessment of CD3+ cells by IHC as a biomarker of inflammation also indicated a trend toward a decrease in ruxolitinib-treated mice (Fig. 5d), although no difference in α-SMA staining was observed between groups (data not shown).
Figure 5.
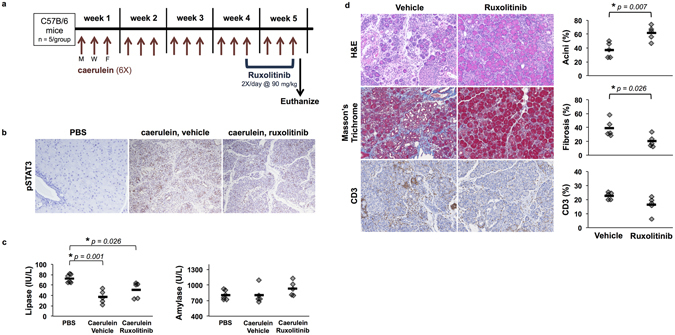
Ruxolitinib reduces disease severity in a murine model of chronic pancreatitis. (a) Treatment schedule for in vivo experiments. Mice were administered caerulein or PBS (control) via intraperitoneal injection 6X/day, 3 days/week, for 5 weeks. Ruxolitinib was given 2X/day via oral gavage during the final week. (b) IHC analysis of pSTAT3 from healthy control mice, pancreatitis-induced mice, and pancreatitis-induced mice treated with ruxolitinib or vehicle control. (c) Serum lipase and amylase levels were analyzed upon euthanasia (* indicates p < 0.05). (d) FFPE pancreata from caerulein-injected mice treated with either ruxolitinib or vehicle were stained for H&E, Masson’s Trichrome, CD3, and α-SMA. Images are representative of 5 mice per group. Slides were quantified using Vectra automated quantitative pathology imaging system and InForm analysis. Graphs display percent acini, fibrosis, CD3, and α-SMA (* indicates p < 0.05).
Discussion
We have demonstrated that PSC display constitutive activation of both the Jak/STAT3 and MAPK pathways and secrete an abundance of several immunomodulatory factors, including IL-6 and MCP-1. When treated with the Jak1/2 inhibitor, ruxolitinib, these cells displayed diminished phosphorylation of STAT3 and reduced cell proliferation. This functional phenotype corresponded with a trend toward quiescence or pseudoquiescense, as evidenced by increased Oil-Red O staining and reduced α-SMA positivity. In contrast, treatment with the MEK inhibitor, MEK162, did not alter activation of PSC and had variable effects on cell proliferation. Finally, in a well-characterized in vivo murine model of chronic pancreatitis, short-term treatment with ruxolitinib led to preservation of acini and reduced fibrosis by IHC as compared to control animals. These results suggest that disruption of Jak/STAT signaling deserves further investigation as a potential therapeutic strategy in CP. This information is important, considering the lack of any clinically effective strategy to reduce inflammation and fibrosis associated with CP.
In the development of therapeutic strategies for CP, PSC have been a prominent target due to the ability of these cells to promote inflammatory and fibrotic processes during disease. Several studies have shown that PSC-targeted therapies can reduce the severity of CP in vivo. Xiao et al. demonstrated that, in mice with caerulein-induced CP, treatment with retinoic acid reduced PSC activation and disease severity47. Tsang et al. showed similar results using an anthraquinone derivative, rhein. In this study, treatment with rhein resulted in reduced PSC activation and fibrosis in caerulein-induced CP48. Thus, the available pre-clinical data indicate that strategies to reduce PSC activity may limit the pathological changes associated with CP.
The observed success of ruxolitinib in reducing severity of caerulein-induced CP is promising, as this type of therapy has not been formally evaluated in CP patients to date. Indeed, our results with ruxolitinib are consistent with another pre-clinical study using the AG490 Jak inhibitor. In this report, AG490 reduced secretion of IL-1β by acinar cells and limited inflammatory changes associated with CP in a caerulein-induced rodent model40. However, the impact of AG490 on PSC was not evaluated in this prior study.
To date, blockade of pro-inflammatory cytokines and their downstream signaling pathways has not been explored rigorously in human clinical trials of pancreatitis. This may be due to a lack of qualifying pre-clinical data with these agents. Furthermore, preventative or therapeutic clinical trials for CP remain difficult for several reasons, including the heterogeneous nature of the disease and lack of standardized clinical endpoints. Considering these challenges, pre-clinical models remain an important component in advancing treatment options for this disease to ensure that interventions studied in humans have a high likelihood of success.
Although these initial data are promising, it is important to acknowledge some limitations. First, all in vitro experiments were conducted in immortalized and non-immortalized cultured PSC. Studies by Gryshchenko et al. have demonstrated differences in the threshold of bradykinin-induced Ca2+ signaling of cultured PSC compared to freshly isolated PSC within pancreatic lobules49. This suggests that differences may exist in the cellular properties of PSC cultured in vitro as compared to cells residing in the pancreatic microenvironment. Additionally, interactions between PSC, acinar cells, and immune cells in the pancreas are likely important to PSC biology and are not fully represented by cultured PSC. Accordingly, testing therapeutic concepts in animal models is critical.
Our in vivo experiments demonstrated the ability of ruxolitinib to reduce disease severity in caerulein-induced CP. One limitation of this study was the use of a single in vivo model. The caerulein-induced murine model is a common and well-characterized model of CP and has been shown to recapitulate several aspects of the pathology present in human CP50. However, like any animal model, there are disadvantages to this system. Namely, this model relies upon continuous administration of supramaximal doses of the CCK-analog caerulein, which is distinct from the mechanisms of CP development in human patients and may reduce the clinical relevance of caerulein-induced CP. In mice, caerulein appears to produce a transient up-regulation of STAT3 in the pancreas, not only in PSC, but also within other cells of the parenchyma. For this reason, it is possible that the effects of Jak/STAT inhibition in this model are the result of inhibiting caerulein-dependent events within the pancreas that may or may not be relevant to human disease. However, others have hypothesized that cytokines produced during pancreatic inflammation promote STAT3 signaling within the pancreas which may then cooperate with mutant KRAS to drive acinar-to-ductal metaplasia and PanIN formation39. This may indicate that inhibition of STAT3 during pancreatic inflammation could limit progression to malignancy, in addition to its potential role in limiting inflammation and fibrosis.
Additionally, our study design examined only short-term dosing with ruxolitinib. Thus, we have not identified the potential long-term effects of this treatment in the context of induced CP. Because the damage caused by caerulein-induced CP is reversible upon cessation of caerulein administration, other clinically-relevant, irreversible animal models should be considered for future studies to assess the impact of long-term inhibition of the Jak/STAT or other pathways in CP51. One irreversible model, the recently characterized duct ligation model, mimics the course of human CP caused by pancreatic duct blockage. This or other models may provide additional avenues to evaluate potential therapeutic interventions for CP in the future52.
In summary, inhibition of Jak/STAT signaling reduces PSC activation in vitro and limits the severity of CP in vivo. Thus, this treatment strategy could be adapted for further pre-clinical testing to limit disease progression in CP.
Methods
Cell lines and reagents
The PaSC and H-iPSC-CP-1 cell lines were kindly provided by Dr. Hanno Steen53–55. The HPF line was purchased from Vitro Biopharma. All other PSC cultures were isolated as described below. All cells were grown in Dulbecco’s Modified Eagle Medium (Gibco, Waltham, MA) with 10% FBS (Gibco) and antibiotics (Gibco). Ruxolitinib (S1378) and MEK162 (S7007) were purchased from Selleck Chemicals (Houston, TX). MTT reagent was purchased from ATCC Bioproducts (Manassas, VA).
Stellate cell isolation and culture
Human pancreatic stellate cells were obtained in accordance with human subjects research guidelines of The Ohio State University under an Institutional Review Board (IRB)-approved protocol following informed consent and cultured as previously described24. Briefly, fresh pancreatic tissue was dissected into 0.5–1 mm3 pieces and plated in uncoated culture wells with Dulbecco’s Modified Eagle Medium (Gibco 11965-092) with 10% FBS (Gibco 26140-079) and antibiotics (Gibco 15240-062). These wells were incubated at 37 °C for 2–3 weeks until PSC were observed growing out of the tissue. Following 2 passages, pure PSC cultures were verified via morphology and by α-SMA and GFAP staining. The h-iPSC-PDAC-1 cell line was processed as described above and was then immortalized via transfection with a lentivirus containing SV40 large T-antigen (GenTarget Inc.).
Microscopic analysis of in vitro PSC activation
PSC were plated on chamber slides (Nalgene Nunc International, Rochester, NY) and incubated at 37 °C for 24 hours, after which fresh media containing ruxolitinib, MEK162, or all-trans retinoic acid (R2625, Sigma Aldrich) was added. After 72 hours, slides were stained with Oil-Red O (O0625, Sigma-Aldrich) or α-SMA antibody (ab5694, Abcam). For Oil-Red O staining, cells were fixed with 4% formalin for 30 minutes at room temperature followed by 60% isopropanol for 5 minutes. The Oil-Red O working solution (diluted and filtered according to manufacturer instructions) was added directly to slides for 5 minutes. Wells were rinsed with tap water and slides were mounted with mounting medium (Life Technologies, Carlsbad, CA). Slides were imaged by light microscopy.
For α-SMA staining, wells were fixed with 4% formalin for 20 minutes followed by 0.2% Triton X-100 for 10 minutes. Slides were then blocked with 4% BSA for 60 minutes. Antibody against α-SMA, diluted in 4% BSA was then added to wells and incubated at room temperature for 1 hour. Wells were then mounted using an anti-fade DAPI mounting medium (P36931, Life Technologies). Slides were allowed to cure overnight and then viewed on a fluorescent microscope.
Analysis of soluble factors in PSC supernatants
Supernatants were collected from PSC cultures at 70–80% confluence and analyzed by bioplex assay (Procarta Cytokine Assay Kit, Affymetrix, Santa Clara, CA). Values were quantified based on a unique standard curve for each analyte.
Immunoblot and densitometric analysis
Immunoblot was performed as described56. Primary antibodies for pSTAT3-Y705 (9145 L) STAT3 (4904 S), pERK (4370 S), ERK (4695 S), PARP (9542 L), and β-actin (4967 S) were purchased from Cell Signaling Technology, Inc. Primary antibody for α-SMA was purchased from Abcam (ab5694). Secondary antibody was purchased from LI-COR (926–32211). Blots were imaged on a LI-COR infrared imager. Densitometry was performed using ImageJ software. All densitometric calculations represent an average of 3 biological replicates.
MTT assay
MTT assays were performed as recommended by the manufacturer. Briefly, PaSC and h-iPSC-PDAC-1 cells cultures were grown in 96 well plates and treated with ruxolitinib, MEK162, or vehicle control. After 72 hours, MTT reagent (ATCC) was added for 2 hours at 37 °C. Absorbance was measured by a plate reader at 450 nM.
Caerulein-induced murine model of pancreatitis
All animal studies were conducted in accordance with the guidelines set forth by The Ohio State University Institutional Animal Care and Use Committee (IACUC) under a protocol approved by the Ohio State University Institutional Review Board. In 4–6 week old female C57BL/6 mice, caerulein was administered at 50 μg/kg by intraperitoneal injection. Six hourly injections were performed three days a week for five weeks57. During the final week of injections, ruxolitinib was administered twice daily at 90 mg/kg by oral gavage. Mice were sacrificed and blood and pancreata were collected for analysis. Serum levels of amylase and lipase were detected by spectrophotometric analysis using the Alfa Wassermann VetAce analyzer (West Caldwell NJ). Pancreata were formalin-fixed and paraffin-embedded before being stained for H&E (Leica 560 MX, Wetzlar, Germany), CD3 (Aligent Technologies, Santa Clara, CA), and Masson’s Trichrome (Polyscientific Inc., Bay Shore, NY).
Quantification of histological acinar loss, fibrosis, and inflammation
All analyses of histological specimens were performed using the Vectra and InForm analysis systems (PerkinElmer). For acinar cell loss, 10 images per mouse were taken from Masson’s Trichrome stained slides. InForm was used to train tissue segmentation to calculate the percent of each image that contained acini. Areas of fibrosis or edema were not included. For fibrosis, the same images were used and the tissue segmenter trained to recognize only areas of fibrosis (blue) and calculate the percentage of each image represented by this tissue category. For inflammation, slides stained with anti-CD3 antibody were quantified by first detecting all nuclei in the field and then calculating the percent of nuclei stained with DAB (CD3+). For all analyses, the scores from each of 10 photos per slide were averaged to create an average per pancreas. These values, along with the overall average per treatment group, were graphed.
Statistics
For immunoblot densitometric analysis, values were normalized to the loading control (β-actin) and the no-treatment control for each experiment. Statistical significance for densitometric analysis, MTT, and serum amylase and lipase was determined using a one-way ANOVA with post-hoc Tukey HSD analysis (p < 0.05). Quantification of IHC, including percentage of acini, fibrosis, and CD3 in pancreata, was analyzed by a simple T-test (p < 0.05).
Acknowledgements
We would like to thank the Comparative Pathology and Mouse Phenotyping Shared Resource (CPMPSR) (The Ohio State University, Columbus, OH) for histological staining and serum enzyme analysis. This work was supported by the National Pancreas Foundation (Lesinski), NIH 1R21AI124687-01 (Lesinski and Ostrowski), NIH 1 U01 DK108327-01 (Conwell and Hart), the Division of Gastroenterology, Hepatology and Nutrition (GHN) at The Ohio State University and the Division of Hematology and Medical Oncology at The Winship Cancer Institute of Emory University. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author Contributions
H.M.K. wrote the main manuscript text, prepared the figures, and performed the in vitro and in vivo experiments with assistance from G.S., C.K., and E.S. T.A.M. provided cultured P.S.C., offered expertise in P.S.C. biology, and assisted in in vivo studies. M.J. analyzed histological slides and offered guidance for in vivo studies. M.C.Y. and C.S.C. immortalized P.S.C. cells. M.B. provided the surgical specimens from C.P. and P.D.A.C. patients. M.C.O. provided expertise in the caerulein mouse model. P.A.H., D.L.C., and G.B.L. provided guidance for the project and helped prepare the text and figures. G.B.L. is the corresponding author.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Mann ST, Stracke H, Lange U, Klör HU, Teichmann J. Alterations of bone mineral density and bone metabolism in patients with various grades of chronic pancreatitis. Metabolism. 2003;52:579–585. doi: 10.1053/meta.2003.50112. [DOI] [PubMed] [Google Scholar]
- 2.Ewald N, Hardt PD. Diagnosis and treatment of diabetes mellitus in chronic pancreatitis. World J Gastroenterol. 2013;19:7276–7281. doi: 10.3748/wjg.v19.i42.7276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pezzilli R. Chronic pancreatitis: maldigestion, intestinal ecology and intestinal inflammation. World J Gastroenterol. 2009;15:1673–1676. doi: 10.3748/wjg.15.1673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Haas S, Krins S, Knauerhase A, Löhr M. Altered bone metabolism and bone density in patients with chronic pancreatitis and pancreatic exocrine insufficiency. JOP. 2015;16:58–62. doi: 10.6092/1590-8577/2898. [DOI] [PubMed] [Google Scholar]
- 5.Lankisch PG, Löhr-Happe A, Otto J, Creutzfeldt W. Natural course in chronic pancreatitis. Pain, exocrine and endocrine pancreatic insufficiency and prognosis of the disease. Digestion. 1993;54:148–155. doi: 10.1159/000201029. [DOI] [PubMed] [Google Scholar]
- 6.Hart, P., Bellin, M., Andersen, D. et al. on behalf of the Consortium for the Study of Chronic Pancreatitis, Diabetes, and Pancreatic Cancer (CPDPC) Type 3c (pancreatogenic) diabetes mellitus secondary to chronic pancreatitis and pancreatic cancer. Lancet Gastroenerol Hepatol1, 226–237 (2016). [DOI] [PMC free article] [PubMed]
- 7.Lowenfels, A. B., Maisonneuve, P. & Lankisch, P. G. Chronic pancreatitis and other risk factors for pancreatic cancer. Gastroenterol Clin North Am28, 673–685, x (1999). [DOI] [PubMed]
- 8.Lowenfels AB, Maisonneuve P. Epidemiology and risk factors for pancreatic cancer. Best Pract Res Clin Gastroenterol. 2006;20:197–209. doi: 10.1016/j.bpg.2005.10.001. [DOI] [PubMed] [Google Scholar]
- 9.Raimondi S, Lowenfels AB, Morselli-Labate AM, Maisonneuve P, Pezzilli R. Pancreatic cancer in chronic pancreatitis; aetiology, incidence, and early detection. Best Pract Res Clin Gastroenterol. 2010;24:349–358. doi: 10.1016/j.bpg.2010.02.007. [DOI] [PubMed] [Google Scholar]
- 10.Bang UC, Benfield T, Hyldstrup L, Bendtsen F, Beck Jensen JE. Mortality, cancer, and comorbidities associated with chronic pancreatitis: a Danish nationwide matched-cohort study. Gastroenterology. 2014;146:989–994. doi: 10.1053/j.gastro.2013.12.033. [DOI] [PubMed] [Google Scholar]
- 11.Howes N, Neoptolemos JP. Risk of pancreatic ductal adenocarcinoma in chronic pancreatitis. Gut. 2002;51:765–766. doi: 10.1136/gut.51.6.765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kolodecik T, Shugrue C, Ashat M, Thrower EC. Risk factors for pancreatic cancer: underlying mechanisms and potential targets. Front Physiol. 2013;4:415. doi: 10.3389/fphys.2013.00415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ting J, et al. Direct Costs of Acute Recurrent and Chronic Pancreatitis in Children in the INSPPIRE Registry. J Pediatr Gastroenterol Nutr. 2016;62:443–449. doi: 10.1097/MPG.0000000000001057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Wilson, G. C., Ahmad, S. A., Schauer, D. P., Eckman, M. H. & Abbott, D. E. Cost-effectiveness of total pancreatectomy and islet cell autotransplantation for the treatment of minimal change chronic pancreatitis. J Gastrointest Surg19, 46–54, discussion 54–45, doi:10.1007/s11605-014-2612-3 (2015). [DOI] [PubMed]
- 15.Masamune A, Watanabe T, Kikuta K, Shimosegawa T. Roles of pancreatic stellate cells in pancreatic inflammation and fibrosis. Clin Gastroenterol Hepatol. 2009;7:S48–54. doi: 10.1016/j.cgh.2009.07.038. [DOI] [PubMed] [Google Scholar]
- 16.Etemad B, Whitcomb DC. Chronic pancreatitis: diagnosis, classification, and new genetic developments. Gastroenterology. 2001;120:682–707. doi: 10.1053/gast.2001.22586. [DOI] [PubMed] [Google Scholar]
- 17.Phillips PA, et al. Rat pancreatic stellate cells secrete matrix metalloproteinases: implications for extracellular matrix turnover. Gut. 2003;52:275–282. doi: 10.1136/gut.52.2.275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kuroda N, et al. The distribution and role of myofibroblasts and CD34-positive stromal cells in normal pancreas and various pancreatic lesions. Histol Histopathol. 2004;19:59–67. doi: 10.14670/HH-19.59. [DOI] [PubMed] [Google Scholar]
- 19.Shimizu K. Mechanisms of pancreatic fibrosis and applications to the treatment of chronic pancreatitis. J Gastroenterol. 2008;43:823–832. doi: 10.1007/s00535-008-2249-7. [DOI] [PubMed] [Google Scholar]
- 20.Ferdek PE, Jakubowska MA, Gerasimenko JV, Gerasimenko OV, Petersen OH. Bile acids induce necrosis in pancreatic stellate cells dependent on calcium entry and sodium-driven bile uptake. J Physiol. 2016;594:6147–6164. doi: 10.1113/JP272774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Apte MV, et al. Desmoplastic reaction in pancreatic cancer: role of pancreatic stellate cells. Pancreas. 2004;29:179–187. doi: 10.1097/00006676-200410000-00002. [DOI] [PubMed] [Google Scholar]
- 22.Xu Z, et al. Role of pancreatic stellate cells in pancreatic cancer metastasis. Am J Pathol. 2010;177:2585–2596. doi: 10.2353/ajpath.2010.090899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Apte MV, Wilson JS. Dangerous liaisons: pancreatic stellate cells and pancreatic cancer cells. J Gastroenterol Hepatol. 2012;27(Suppl 2):69–74. doi: 10.1111/j.1440-1746.2011.07000.x. [DOI] [PubMed] [Google Scholar]
- 24.Mace TA, et al. Pancreatic cancer-associated stellate cells promote differentiation of myeloid-derived suppressor cells in a STAT3-dependent manner. Cancer research. 2013;73:3007–3018. doi: 10.1158/0008-5472.CAN-12-4601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Apte MV, et al. Pancreatic stellate cells are activated by proinflammatory cytokines: implications for pancreatic fibrogenesis. Gut. 1999;44:534–541. doi: 10.1136/gut.44.4.534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schneider E, et al. Identification of mediators stimulating proliferation and matrix synthesis of rat pancreatic stellate cells. Am J Physiol Cell Physiol. 2001;281:C532–543. doi: 10.1152/ajpcell.2001.281.2.C532. [DOI] [PubMed] [Google Scholar]
- 27.Mews P, et al. Pancreatic stellate cells respond to inflammatory cytokines: potential role in chronic pancreatitis. Gut. 2002;50:535–541. doi: 10.1136/gut.50.4.535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Michalski CW, et al. Mononuclear cells modulate the activity of pancreatic stellate cells which in turn promote fibrosis and inflammation in chronic pancreatitis. J Transl Med. 2007;5:63. doi: 10.1186/1479-5876-5-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang W, Liu HT. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002;12:9–18. doi: 10.1038/sj.cr.7290105. [DOI] [PubMed] [Google Scholar]
- 30.Ogata A, et al. IL-6 triggers cell growth via the Ras-dependent mitogen-activated protein kinase cascade. J Immunol. 1997;159:2212–2221. [PubMed] [Google Scholar]
- 31.Fan Y, Mao R, Yang J. NF-κB and STAT3 signaling pathways collaboratively link inflammation to cancer. Protein Cell. 2013;4:176–185. doi: 10.1007/s13238-013-2084-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Grivennikov SI, Karin M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010;21:11–19. doi: 10.1016/j.cytogfr.2009.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhang H, et al. IL-6 trans-signaling promotes pancreatitis-associated lung injury and lethality. J Clin Invest. 2013;123:1019–1031. doi: 10.1172/JCI64931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Talar-Wojnarowska R, et al. Clinical significance of interleukin-6 (IL-6) gene polymorphism and IL-6 serum level in pancreatic adenocarcinoma and chronic pancreatitis. Dig Dis Sci. 2009;54:683–689. doi: 10.1007/s10620-008-0390-z. [DOI] [PubMed] [Google Scholar]
- 35.Miron N, Miron MM, Milea VG, Cristea V. Proinflammatory cytokines: an insight into pancreatic oncogenesis. Roum Arch Microbiol Immunol. 2010;69:183–189. [PubMed] [Google Scholar]
- 36.Duggan SN, et al. An association between abnormal bone turnover, systemic inflammation, and osteoporosis in patients with chronic pancreatitis: a case-matched study. Am J Gastroenterol. 2015;110:336–345. doi: 10.1038/ajg.2014.430. [DOI] [PubMed] [Google Scholar]
- 37.Park J, et al. Interleukin-6 is associated with obesity, central fat distribution, and disease severity in patients with acute pancreatitis. Pancreatology. 2015;15:59–63. doi: 10.1016/j.pan.2014.11.001. [DOI] [PubMed] [Google Scholar]
- 38.Mace, T. A. et al. IL-6 and PD-L1 antibody blockade combination therapy reduces tumour progression in murine models of pancreatic cancer. Gut, doi:10.1136/gutjnl-2016-311585 (2016). [DOI] [PMC free article] [PubMed]
- 39.Corcoran RB, et al. STAT3 plays a critical role in KRAS-induced pancreatic tumorigenesis. Cancer Res. 2011;71:5020–5029. doi: 10.1158/0008-5472.CAN-11-0908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yu JH, Kim KH, Kim H. Suppression of IL-1beta expression by the Jak 2 inhibitor AG490 in cerulein-stimulated pancreatic acinar cells. Biochem Pharmacol. 2006;72:1555–1562. doi: 10.1016/j.bcp.2006.07.008. [DOI] [PubMed] [Google Scholar]
- 41.Lesinski GB. The potential for targeting the STAT3 pathway as a novel therapy for melanoma. Future Oncol. 2013;9:925–927. doi: 10.2217/fon.13.83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Johnston PA, Grandis JR. STAT3 signaling: anticancer strategies and challenges. Mol Interv. 2011;11:18–26. doi: 10.1124/mi.11.1.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Harrison C, et al. JAK inhibition with ruxolitinib versus best available therapy for myelofibrosis. N Engl J Med. 2012;366:787–798. doi: 10.1056/NEJMoa1110556. [DOI] [PubMed] [Google Scholar]
- 44.Vannucchi AM, et al. Ruxolitinib versus standard therapy for the treatment of polycythemia vera. N Engl J Med. 2015;372:426–435. doi: 10.1056/NEJMoa1409002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Mesa RA. Ruxolitinib, a selective JAK1 and JAK2 inhibitor for the treatment of myeloproliferative neoplasms and psoriasis. IDrugs. 2010;13:394–403. [PubMed] [Google Scholar]
- 46.Zhao C, et al. Rational combination of MEK inhibitor and the STAT3 pathway modulator for the therapy in K-Ras mutated pancreatic and colon cancer cells. Oncotarget. 2015;6:14472–14487. doi: 10.18632/oncotarget.3991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Xiao W, et al. Retinoic Acid Ameliorates Pancreatic Fibrosis and Inhibits the Activation of Pancreatic Stellate Cells in Mice with Experimental Chronic Pancreatitis via Suppressing the Wnt/β-Catenin Signaling Pathway. PLoS One. 2015;10:e0141462. doi: 10.1371/journal.pone.0141462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Tsang SW, et al. Rhein, a natural anthraquinone derivative, attenuates the activation of pancreatic stellate cells and ameliorates pancreatic fibrosis in mice with experimental chronic pancreatitis. PLoS One. 2013;8:e82201. doi: 10.1371/journal.pone.0082201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Gryshchenko O, Gerasimenko JV, Gerasimenko OV, Petersen OH. Ca(2+) signals mediated by bradykinin type 2 receptors in normal pancreatic stellate cells can be inhibited by specific Ca(2+) channel blockade. J Physiol. 2016;594:281–293. doi: 10.1113/JP271468. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Reed, A. & Gorelick, F. In Pancreapedia: Exocrine Pancreas Knowledge Base (2014).
- 51.Mayerle, J., Sendler, M. & Lerch, M. M. Secretagogue (Caerulein) induced pancreatitis in rodents (2013).
- 52.Sendler, M. et al. Complement Component 5 Mediates Development of Fibrosis, via Activation of Stellate Cells, in 2 Mouse Models of Chronic Pancreatitis. Gastroenterology149, 765-776 e710, doi:10.1053/j.gastro.2015.05.012 (2015). [DOI] [PMC free article] [PubMed]
- 53.Paulo JA, Urrutia R, Banks PA, Conwell DL, Steen H. Proteomic analysis of an immortalized mouse pancreatic stellate cell line identifies differentially-expressed proteins in activated vs nonproliferating cell states. J Proteome Res. 2011;10:4835–4844. doi: 10.1021/pr2006318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Paulo JA, Kadiyala V, Banks PA, Conwell DL, Steen H. Mass spectrometry-based quantitative proteomic profiling of human pancreatic and hepatic stellate cell lines. Genomics Proteomics Bioinformatics. 2013;11:105–113. doi: 10.1016/j.gpb.2013.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Jesnowski R, et al. Immortalization of pancreatic stellate cells as an in vitro model of pancreatic fibrosis: deactivation is induced by matrigel and N-acetylcysteine. Lab Invest. 2005;85:1276–1291. doi: 10.1038/labinvest.3700329. [DOI] [PubMed] [Google Scholar]
- 56.Bill A, et al. Anti-proliferative effect of cytohesin inhibition in gefitinib-resistant lung cancer cells. PLoS One. 2012;7:e41179. doi: 10.1371/journal.pone.0041179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Yamada T, et al. Adiponectin deficiency enhanced the severity of cerulein-induced chronic pancreatitis in mice. J Gastroenterol. 2010;45:742–749. doi: 10.1007/s00535-010-0205-9. [DOI] [PubMed] [Google Scholar]