

Abstract
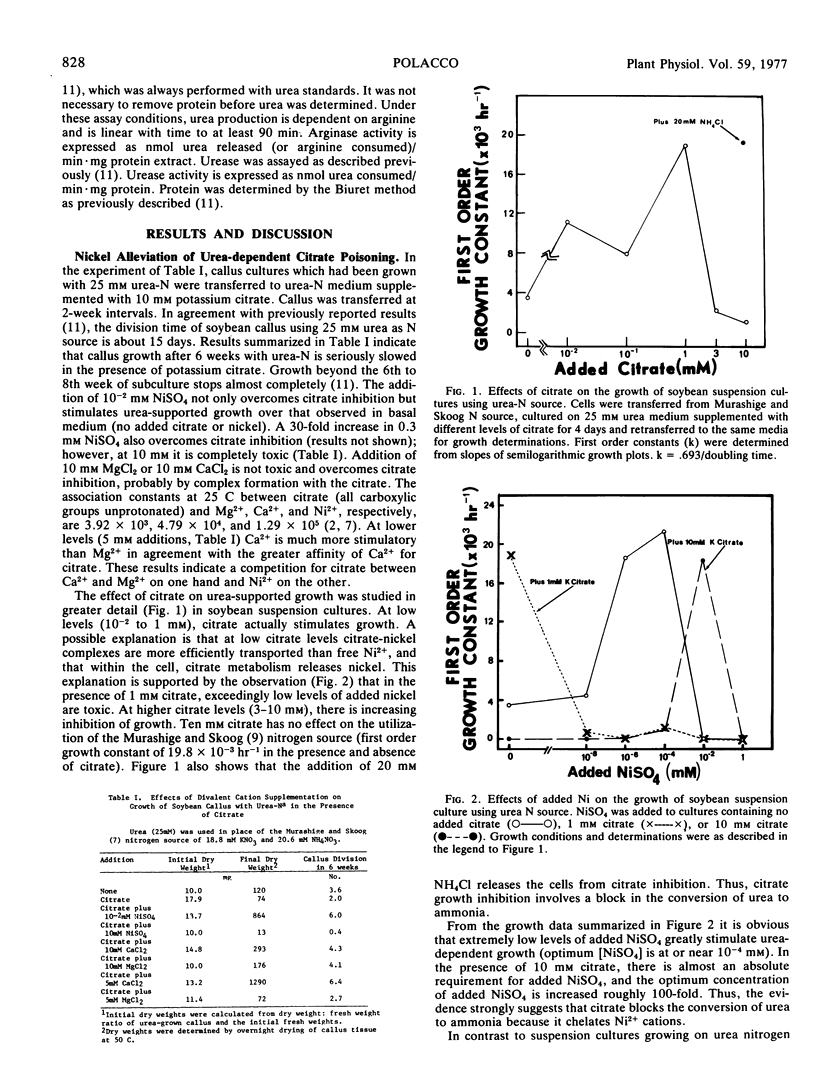
Potassium citrate (10 mM, pH 6) inhibits the growth of cultured (Glycine max L.) cells when urea is the sole nitrogen source. Ureadependent citrate toxicity is overcome by three separate additions to the growth medium: (a) NH4Cl (20 mM); (b) high levels of MgCl2 (10 mM) or CaCl2 (5-10 mM); (c) low levels of NiSO4 (10−2 mM). Additions of 10−2 mM NiSO4 not only overcome citrate growth inhibition but the resultant growth is usually better than urea-supported growth in basal medium (neither added citrate nor added nickel). In the absence of added citrate, exceedingly low levels of NiSO4 (10−4 mM) strongly stimulate urea-supported growth in suspension cultures.
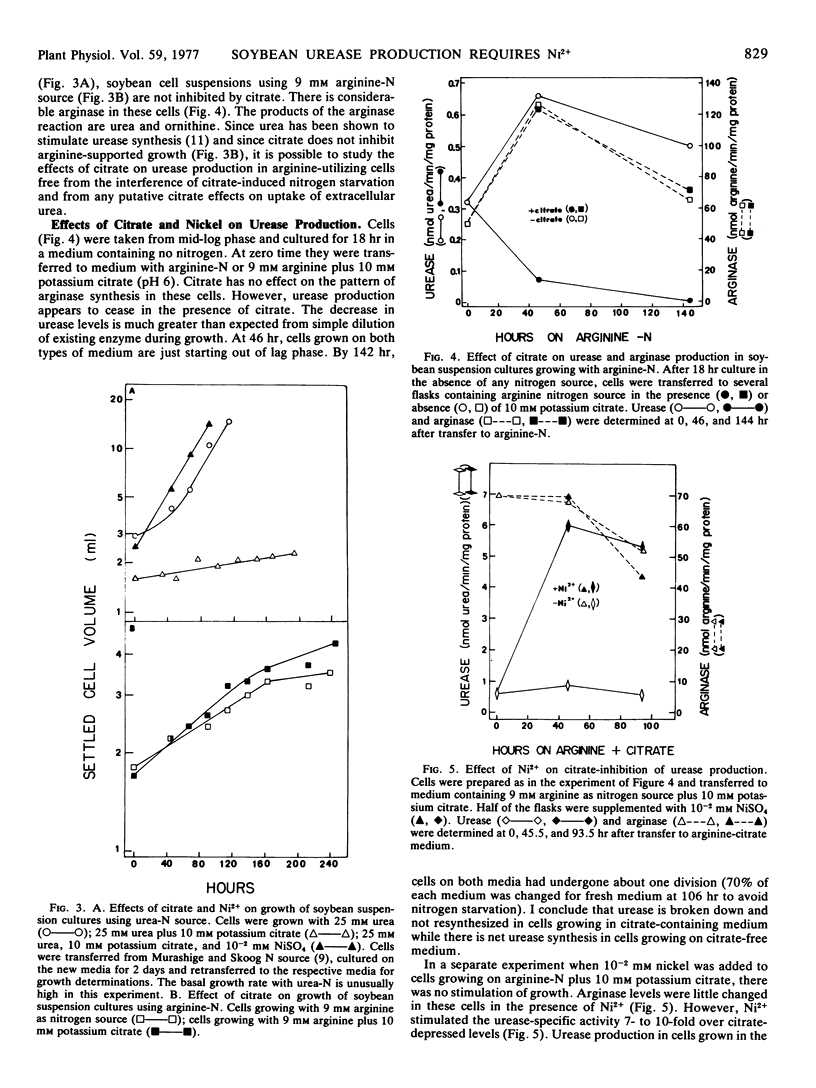
Citrate does not inhibit growth when arginine is sole nitrogen source. However, cells using arginine have no net urease synthesis in the presence of 10 mM potassium citrate. When 10−2 mM NiSO4 is added to this medium, urease specific activity is 10 times that observed in basal medium lacking both citrate and added nickel.
Citrate is a chelator of divalent cations. That additional Mg2+ or Ca2+ alleviates urea-dependent citrate toxicity indicates that citrate is acting by chelation, probably of another trace divalent cation; this is probably Ni2+ since at 10−2 mM it overcomes citrate toxicity and at 10−4 mM it stimulates urea-supported growth in the absence of citrate. That ammonia overcomes citrate toxicity indicates that the trace Ni2+ is essential specifically for the conversion of urea to ammonia. Ni2+ stimulation of urease levels in arginine-grown cells supports this contention.
In basal medium, soybean cells grow slowly with urea nitrogen source presumably because the trace amounts of Ni2+ present (≤10−6 mM) are growth-limiting.
Full text
PDF

Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- Behrend J., Mateles R. I. Nitrogen Metabolism in Plant Cell Suspension Cultures: II. Role of Organic Acids during Growth on Ammonia. Plant Physiol. 1976 Oct;58(4):510–512. doi: 10.1104/pp.58.4.510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dixon N. E., Gazzola T. C., blakeley R. L., Zermer B. Letter: Jack bean urease (EC 3.5.1.5). A metalloenzyme. A simple biological role for nickel? J Am Chem Soc. 1975 Jul 9;97(14):4131–4133. doi: 10.1021/ja00847a045. [DOI] [PubMed] [Google Scholar]
- Gamborg O. L., Shyluk J. P. The culture of plant cells with ammonium salts as the sole nitrogen source. Plant Physiol. 1970 May;45(5):598–600. doi: 10.1104/pp.45.5.598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kollöffel C., van Dijke H. D. Mitochondrial Arginase Activity from Cotyledons of Developing and Germinating Seeds of Vicia faba L. Plant Physiol. 1975 Mar;55(3):507–510. doi: 10.1104/pp.55.3.507. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polacco J. C. Nitrogen metabolism in soybean tissue culture: I. Assimilation of urea. Plant Physiol. 1976 Sep;58(3):350–357. doi: 10.1104/pp.58.3.350. [DOI] [PMC free article] [PubMed] [Google Scholar]