

Abstract
Background
The aims of this study were to investigate Salmonella contamination in broiler chicken farms and slaughterhouses, to assess the antibiotic resistance profile in avian and human Salmonella isolates, and to evaluate the relationship between avian and human Extended Spectrum β-Lactamase (ESBL)-producing isolates. Salmonella was screened in different sample matrices collected at thirty-two chicken farms and five slaughterhouses. The human isolates were recovered from clinical specimens at the University Teaching Hospital of Constantine (UTH). All suspected colonies were confirmed by MALDI-TOF (Matrix Assisted Laser Desorption Ionization Time OF light) and serotyped. Susceptibility testing against 13 antibiotics including, amoxicillin/clavulanic acid, ticarcillin, cefoxitin, cefotaxime, aztreonam, imipenem, ertapenem, gentamicin, amikacin, ciprofloxacin, colistin, trimethoprim/sulfamethoxazole and fosfomycin, was performed using the disk diffusion method on Mueller-Hinton agar. ESBL-production was screened by the double-disk synergy test and confirmed by molecular characterization using PCR (polymerase chain reaction) amplification and sequencing of ESBL encoding genes. Clonality of the avian and human strains was performed using the Multi Locus Sequencing Typing method (MLST).
Results
Forty-five isolated avian Salmonella strains and 37 human collected ones were studied. Five S. enterica serotypes were found in avian isolates (mainly Kentucky) and 9 from human ones (essentially Infantis). 51.11% and 26.6% of the avian isolates were resistant to ciprofloxacin and cefotaxime, respectively, whereas human isolates were less resistant to these antibiotics (13.5% to ciprofloxacin and 16.2% to cefotaxime). Eighteen (12 avian and 6 human) strains were found to produce ESBLs, which were identified as bla CTX-M-1 (n = 12), bla CTX-M-15 (n = 5) and bla TEM group (n = 8). Interestingly, seven of the ESBL-producing strains (5 avian and 2 human) were of the same ST (ST15) and clustered together, suggesting a common origin.
Conclusion
The results of the combined phenotypic and genotypic analysis found in this study suggest a close relationship between human and avian strains and support the hypothesis that poultry production may play a role in the spread of multidrug-resistant Salmonella in the human community within the study region.
Electronic supplementary material
The online version of this article (doi:10.1186/s12917-017-1050-3) contains supplementary material, which is available to authorized users.
Keywords: Salmonella, poultry, human, serotype, antimicrobial resistance, resistance genes, clonality
Background
Salmonella infections are a major public health problem with a significant social and economic impact. Many animal species are potential reservoirs for this bacterium, especially chickens, pigeons and reptiles [1]. Humans can commonly acquire the infection through the food chain [2]. Young and immunocompromised patients are the most exposed to dangerous complications which are generally treated with fluoroquinolones and extended-spectrum cephalosporins that are largely used in veterinary medicine [3].
In Algeria, the poultry industry has grown remarkably since 1980. However, as a result of the deleterious hygienic conditions, many infectious diseases, such as Salmonellosis, were detected in broilers and constitute a risk to the human health [4]. Due to a lack of surveillance programs, information on the prevalence of Salmonella and other food pathogens is incomplete. For instance, a study by the Pasteur Institute of Algeria revealed that 11% of food poisoning cases were caused by Salmonella spp. in 2011 [5]. 47% of these cases were mainly related to the consumption of chicken meat, as reported by Mouffok et al., [5]. Eggs and ovoproducts are also among the major sources of human infections, which require a thorough assessment and control measures for Salmonella spp. in the poultry industry [6]. These measures are well detailed in the Official Journal of the Algerian Republic (No. 36 of June 8, 2003); but were only applied to the poultry industry facilities in the public sector as well as to private poultry production units and facilities [7].
Selective pressure due to the misuse of antibiotics in humans and domestic livestock is one of the many factors that has led to the emergence of antibiotic resistance in commensal and pathogenic bacteria; thus, multidrug-resistant (MDR) Salmonella have increasingly been isolated from various food products worldwide [8].
Drug resistance is growing and has affected critically important classes of antibiotics, such as the β-lactams, which are among the most significant bactericidal antibiotics used to treat bacterial infections in humans [9]. Extended spectrum β-lactamases (ESBLs) were identified following the introduction of extended-spectrum oxyimino-cephalosporins in the 1980s for the treatment of severe human infections [10]. In veterinary medicine, a variety of these drugs are currently authorized for use, resulting in the emergence of ESBL-producing Gram-negative bacteria [11, 12].
TEM, SHV and CTX-M are the most prevalent ESBL types. Over the last decade, rates of CTX-M producing bacteria have increased worldwide in comparison with TEM and SHV [13]. This situation is rendered more complicated as these enzymes confer co-resistance to other drug classes [13, 14].
In Algeria, the first ESBLs were detected in non-typhoidal Salmonella in 1994 from humans [15], but until now there has been no information on the magnitude of this problem in animals and humans. This situation drove us to identify the contamination status of Salmonella serotypes in chicken farms and slaughterhouses, to assess their sensitivity to antimicrobials and ESBL-production and, finally, to evaluate their clonality with human pathogenic strains.
Methods
Study area
The present study was carried out between December 2011 and May 2013. Thirty-two chicken farms and five chicken slaughterhouses located in the province of Skikda, Algeria, took part in this survey. The choice of this study area was motivated by the size of the poultry industry and the frequent occurrence of infectious gastrointestinal pathologies as reported by local veterinary practitioners [16].
Twenty-seven poultry houses had concrete walls and floors and corrugated metal sheet roofs while the remaining five houses had earth floors with walls and roofs made of straw and reeds covered with plastic foil. None of the farming sites was fenced, allowing free access for domestic and wild animals. Their rearing capacities vary from 3500 to 20,000 birds per house.
All slaughterhouses had concrete floors with earthenware walls. The slaughtering capacity ranges from 2000 to 7000 chickens per day and the broilers are brought from different poultry farms located in several neighboring provinces. The number of chickens and slaughterhouses is statistically representative of the study region and governed by the capacity of the laboratory processing the samples.
The human Salmonella strains were kindly provided by the University Teaching Hospital (UTH) of Constantine and included in the study.
Study design
For technical reasons, including access to the sampling sites, a total of 1194 samples were collected during two sampling periods (between December 2011 and September 2012 and between December 2012 and May 2013). The poultry houses were visited at two periods (when the birds were aged 15–30 days and 45–60 days).
From the chicken farms, a total of 320 samples of water were taken from drinking vessels, 160 samples of feed were taken from the feeding vessels, 330 cloacal swabs were taken and 320 droppings were collected and placed in sterile containers. 64 surface wipes (25 cm × 25 cm, AES Chemunex, Combourg, France) were also obtained from a height of 30 cm from the ground over a 400 cm2 area of the four walls of the poultry house and placed into sterile Stomacher bags.
The five poultry slaughterhouses were visited once for sampling. Due to limited financial resources, in each poultry slaughterhouse, we pooled individual samples to minimize study costs. The samples were randomly taken from three organs from five chickens (5 g of 5 caeca, 5 g of 5 livers, 5 g of 5 neck skins), and from the environment (one sample of carcass rinsing water, one swab from a sticking knife and one wipe from the walls). The slaughtered animals were brought from several neighboring provinces.
All samples were transported to the laboratory, on ice packs within a period not exceeding two hours, to be treated on the same day or kept in the refrigerator overnight.
Salmonella isolation and identification
Bacteriological analyses were performed according to the EN/ISO 6579–2002/Amd1:2007 protocol for Salmonella detection in food and animal feedstuffs. 25 g of samples (droppings, feed, liver, caeca, neck skin) were individually pre-enriched with 225 mL of buffered peptone water broth (PWB) (Fluka, Sigma Aldrich, France). The swabs were individually placed in 10 mL PWB, while 100 mL of drinking and carcass rinsing water was individually mixed with 100 mL of double strength PWB for pre-enrichment according to NF U 47–101 Standard (2005) [17]. All samples were incubated at 37 °C for 18–20 h. From each pre-enrichment solution, 1 mL and 0.1 mL were respectively transferred into 10 mL of enrichment Muller-Kauffmann tetrathionate/novobiocin broth (AES Chemunex Combourg, France) and 10 mL of Rappaport Vassiliadis broth (Merck Darmstadt, Germany), incubated respectively at 37 °C and 42 °C for 24 h. Both enriched samples were then streaked on XLD (Fluka analytical Steinheim, Switzerland) and Hektoen agars (Pasteur Institute of Algeria) and incubated at 37 °C for 24 h [18]. Suspected colonies were first identified with the API 20E System (bioMérieux, France), then with MALDI-TOF (Bruker Daltonics GmbH, Germany) [19]. Confirmed Salmonella isolates were serotyped according to Kauffmann-White-Le Minor’s scheme [20].
Human clinical strains
We selected 37 (non-repetitive) strains recovered from clinical specimens at the UTH of Constantine over a decade (2005–2015). These were isolated from stool samples collected from different wards and among which 26 strains derived from the diarrheic stools of infants admitted to the neonatology ward. The Main characteristics of patients are shown in Additional file 1.
The strains were confirmed using MALDI-TOF mass spectrometry for prescreening Salmonella species and sub-species and to identify epidemiologically important serovars which were further tested with conventional serotyping method.
Antimicrobial susceptibility testing and ESBL detection
All human and avian strains were submitted to susceptibility testing against antibiotics using the disk diffusion method on Mueller-Hinton (MH) agar, and the results were interpreted according to the European Comitee on Antimicrobial Suceptibility Testing (EUCAST) [21] (Additional file 2). Thirteen antibiotics (Bio-Rad, France) were tested: amoxicillin/clavulanic acid AMC (20/10 μg), ticarcillin TIC (75 μg), cefoxitin FOX (30 μg), cefotaxime CTX (5 μg), aztreonam ATM (30 μg), imipenem IPM (10 μg), ertapenem ETP (10 μg), gentamicin GEN (10 μg), amikacin AK (30 μg), ciprofloxacin CIP (5 μg), colistin CT (50 μg), trimethoprim/sulfamethoxazole SXT (1.25 μg/23.75 μg) and fosfomycin FF (50 μg).
Extended spectrum β-lactamase production was screened by the double-disc synergy test (DDST) [22].
PCR detection of ESBL genes
Total nucleic acids were extracted using a BioRobot EZ1 Advanced XL instrument (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions.
Detection of β -lactamase genes (including bla TEM, bla SHV, and bla CTX-M) was carried out by polymerase chain reaction (PCR) using specific primers: bla CTX-M-1 group [23], bla CTX-M-9 group [24], bla TEM group [25] and bla SHV [26] and the master mix QuantiTect Probe PCR Kit (QIAGEN, Hilden, Germany).
Amplification products were detected by electrophoresis using agarose gels containing SYBR safe (Invitrogen, Leek, the Netherlands), along with a DNA molecular weight marker (BenchTop pGEM®DNA Marker, Promega, Madison, Wisconsin, USA). Visualization of gels was carried out using the BenchTop pGEM®DNA Marker (Promega, Madison, Wis- consin, USA) under ultraviolet illumination.
ESBL genes sequencing and multilocus sequence typing
ESBL-positive PCR products were purified using the NucleoFast 96 PCR plate (Machery-Nagel EURL, France) and sequenced using the BigDye terminator chemistry on an ABI3730 automated sequencer (Applied Biosystems, Foster City, California, USA). The obtained sequences were analyzed with the ARG-ANNOT database [27].
Multilocus sequence typing (MLST) is useful in assessing the role of specific STs in human and animal disease and assessing overlap between these hosts. It was carried out by PCR amplification and sequencing of seven housekeeping genes: thrA (aspartokinase + homoserine dehydrogenase), purE (phosphoribosylaminoimidazole carboxylase), sucA (alpha ketoglutarate dehydrogenase), hisD (histidinol dehydrogenase), aroC (chorismate synthase), hemD (uroporphyrinogen III cosynthase), and dnaN (DNA polymerase III beta subunit), as described by Kidgell et al. 2002 [28].
Briefly, template DNA prepared from bacterial isolates was amplified by PCR with the use of an oligonucleotide sequence for seven housekeeping genes (available in the MLST database: http://mlst.warwick.ac.uk/mlst/dbs/Senterica). Sequencing with the same automated sequencer of the PCR product was carried out by the dideoxynucleotide chain termination method using the master mix QuantiTect Probe PCR Kit (QIAGEN, Hilden, Germany). Forward and reverse DNA sequences were assembled, trimmed, edited, and analyzed for each gene fragment using the ARG-ANNOT database [27]. Allelic profile and sequence type determinations were assigned according to the Salmonella MLST database: http://mlst.warwick.ac.uk/mlst/dbs/Senterica.
Clonality analysis
Protein mass profiles were obtained using a Microflex LT MALDI-TOF mass spectrometer (Bruker Daltonics, Germany), with Flex Control software (Bruker Daltonics). The spectrum profiles obtained were visualized with Flex analysis v.3.3 software and exported to ClinProTools software v.2.2 and MALDI-Biotyper v.3.0 (Bruker Daltonics, Germany) for data processing (smoothing, baseline subtraction and spectra selection) and evaluation with cluster analysis.
The phyloproteomic analysis of ESBL-positive Salmonella strains from human and poultry origins was assessed through construction and comparison of their characteristic reference spectra (main spectra) with the MALDI-Biotyper v.3.0 software (Bruker Daltonics, Germany). Cluster analysis was performed based on pairwise comparisons of specific main spectra (MSP: mean spectra projection dendrogram) of the different strains to generate a dendrogram of similarities among spectra profiles using the software default correlation function. A distance level of 560 was selected for clustering evaluation of the isolates.
Statistical analysis
Differences in contamination levels of poultry houses at two sampling periods (15–30 days versus 45–60 days), and the antimicrobial resistance patterns between avian and human Salmonella strains, were assessed by the Chi square test (at 95% CI and p < 0.05) or Fisher’s exact test if N is less than 20 and one expected cell is less than or equal 5. All statistical analyses were performed using IBM SPSS Statistics version 24 software (2016).
Results
Frequency of isolation of Salmonella serotypes in broilers, slaughterhouses and human samples
Forty-five Salmonella enterica from slaughterhouses and poultry farms and 37 of human clinical origin were studied. 34.37% of the poultry farms and all slaughterhouses were contaminated with Salmonella and the isolation rate varied depending on the sampling matrix. The samples taken at the age of 15–30 days were more contaminated than those collected at 45–60 days; however, the difference was not significant (p > 0.05).
The isolated Salmonella strains belonged mainly to two serotypes: Kentucky and Heidelberg, and the remaining strains were Enteritidis, Virginia and Newport. There was an evident heterogeneous distribution of serotypes in poultry farms and slaughterhouses (Table 1).
Table 1.
Frequency of isolation of Salmonella enterica subsp. enterica serotypes in different sample matrices from poultry farms and slaughterhouses
| Serotype | Poultry farms and slaughterhouses | Samples | ||
|---|---|---|---|---|
| N° of positive (ID n°) | (%) | N° of positive samples | (%) | |
| Kentucky | 7 Poultry farms (F1, F6, F13, F14, F18, F20, F32) | 21.87 | 5 cloacal swabs 6 droppings 2 wipes 3 water samples |
1.51 1.87 3.12 0.93 |
| 4 Slaughterhouses (S1, S2, S3, S4) | 80 | 1 sticking knife 2 caeca 1 liver 1 wipe |
20 8.0 4.0 20 |
|
| Heidelberg | 4 Poultry farms (F12, F13, F15, F27) | 12.5 | 6 cloacal swabs 4 droppings 2 water samples 1 wipe |
1.81 1.25 0.62 1.56 |
| (0) Slaughterhouse | 0 | - | - | |
| Virginia | 1 Poultry farm (F31) | 3.12 | 1 water sample | 0.31 |
| 2 Slaughterhouses (S1,S4) | 40 | 1 wipe 2 neck skins 1caeca |
20 40 4.0 |
|
| Enteritidis | 3 Poultry farms (F18, F20, F27) | 9.37 | 2 cloacal swabs 1 wipe |
0.61 1.56 |
| 1 Slaughterhouse (S3) | 20 | 1 rinse water sample | ||
| Newport | 1 Poultry farm (F15) | 3.12 | 1 wipe | 1.56 |
| (0) Slaughterhouse | 0 | - | - | |
| Total | 11 Poultry farm (F1, F6, F12, F13, F14, F15, F18, F20, F27, F31, F32) | 34.7 | 13 cloacal swabs 10 droppings 5 wipes 6 water samples |
3.93 3.12 7.81 1.87 |
| 5 Slaughterhouse (S1, S2, S3, S4, S5) | 100 | 1 sticking knife 2 neck skins 1 rinse water sample 3 caeca 1 liver 2 wipe |
20 8.0 4.0 12 4.0 40 |
|
F Farm, S Slaughterhouse
Among the human Salmonella strains, Infantis was the most frequent serotype, followed by Senftenberg, Enteritidis, Kedougou, Tyhimurium, Heidelberg, Kentucky, Ohio and Arizona. Most of these strains were from infants and the others were from adult diarrheic stools (especially Salmonella enterica serotypes Enteritidis and Typhimurium).
Drug resistance patterns of the isolated Salmonella strains
A high frequency of resistance to ciprofloxacin (51.1%) was noted in Salmonella isolates from both chicken farms and slaughterhouses. These strains were resistant to cephalosporins (26.6% to cefotaxime), aztreonam (26.6%), ticarcillin (46.6%) and gentamicin (22.2%). ESBL-production was found in 26.6% of these avian isolates (11 Salmonella ser. Heidelberg and one Salmonella ser. Newport).
The human Salmonella isolates were highly resistant to ticarcillin (56.75%), amoxicillin (45.94%) and a less extent to trimethoprim/sulfamethoxazole (24.3%), gentamicin (24.3%), aztreonam (18.9%), cefotaxime (16.2%), ciprofloxacin (13.5%) and amikacin (10.8%). Six ESBL-positive strains were detected (Fig. 1).
Fig. 1.
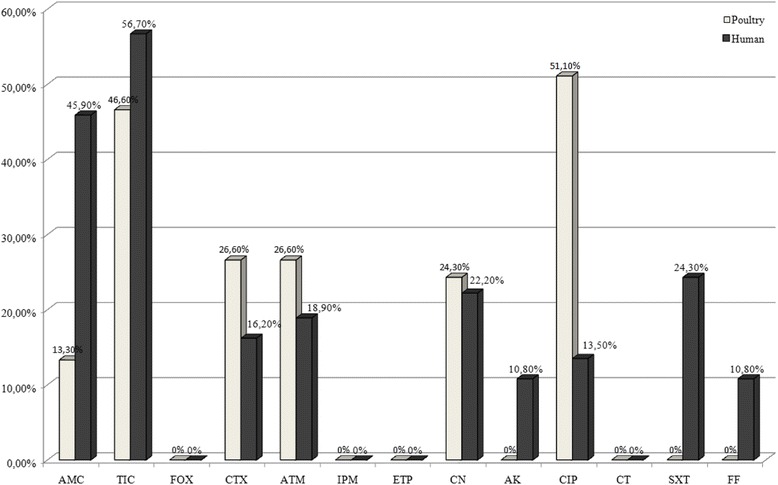
Distribution of resistance to antibiotics among Salmonella isolates from poultry and humans
The difference in resistance to antibiotics, between poultry strains and human ones was significant for: amoxicillin/clavulanic acid (P < 0.05), gentamicin (P < 0.05), ciprofloxacin (P < 0.05) and fosfomycin (P < 0.05). However, it was not significant for cefotaxime, ticarcillin, aztreonam and amikacin (P > 0.05).
The characteristics of the 18 ESBL-producing strains isolated from chicken farms/slaughterhouses and patients are shown in Table 2. Eleven of the Salmonella ser. Heidelberg strains harbored bla CTX-M-1 genes. The bla TEM group was identified in one avian Salmonella ser. Newport isolate as well as in S. Heidelberg, in which it was coupled with the bla CTX-M-1 gene. Human strains harbored the bla CTX-M-15 gene in association with bla TEM, except one strain which carried instead the bla CTX-M-1. None of the strains were positive for bla CTX-M-9 and bla SHV genes.
Table 2.
Antimicrobial resistance and resistant genes profiles of ESBLs producing Salmonella enterica strains isolated from poultry and humans
| Strain ID N° |
Origin | Antimicrobial Resistance Pattern | Serotype | B-Lactamase | ST |
|---|---|---|---|---|---|
| 162 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 163 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 164 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 165 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 167 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 169 | Poultry | TIC, CTX, ATM, | Newport | TEM | 198 |
| 170 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 171 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 172 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1, TEM | 15 |
| 174 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 177 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 178 | Poultry | TIC, CTX, ATM, | Heidelberg | CTX-M-1 | 15 |
| 305 | Human | AMC,TIC,CTX, ATM, GEN,FF | Senftenberg | CTX-M-15 | 14 |
| 476 | Human | AMC, TIC, CTX, ATM,GEN,AK,SXT | Infantis | CTX-M-15, TEM | 38 |
| 883 | Human | CTX, ATM, GEN, AK, SXT | Heidelberg | CTX-M-15, TEM | 15 |
| 884 | Human | AMC,TIC,CTX, ATM, GEN, AK, SXT | Heidelberg | CTX-M-15, TEM | 15 |
| YFA | Human | AMC, TIC, CTX,GEN | Infantis | CTX-M-1 | 32 |
| 1577 | Human | AMC,TIC,CTX, GEN, AK, SXT | Infantis | CTX-M-15, TEM | 16 |
AMC Amoxicillin/Clavulanic Acid, TIC Ticarcillin, FOX Cefoxitin, CTX Cefotaxime, ATM Aztreonam, IPM Imipenem, ETP Ertapenem, GEN Gentamicin, AK Amikacin, CIP Ciprofloxacin, CT Colistin, SXT Trimethoprim/Sulfamethoxazole, FF Fosfomycin
The MLST analysis showed that the salmonella isolates belonged to six different sequence types (ST) including, ST14, ST15, ST16, ST32, ST38 and ST198 (Fig. 2). The results demonstrate that ST15 represents the predominant clone. Indeed, this ST was found in 13 Salmonella ser. Heidelberg strains (two human and 11 avian) (Fig. 2). In addition, the phylogenetic tree shows that seven Salmonella ser. Heidelberg strains (two human and five avian) clustered together and belonged to the same sequence type ST15 (Fig. 2), which suggests a possible crossing of this serotype, and particularly this ST between the poultry and the human community in northeastern Algeria.
Fig. 2.
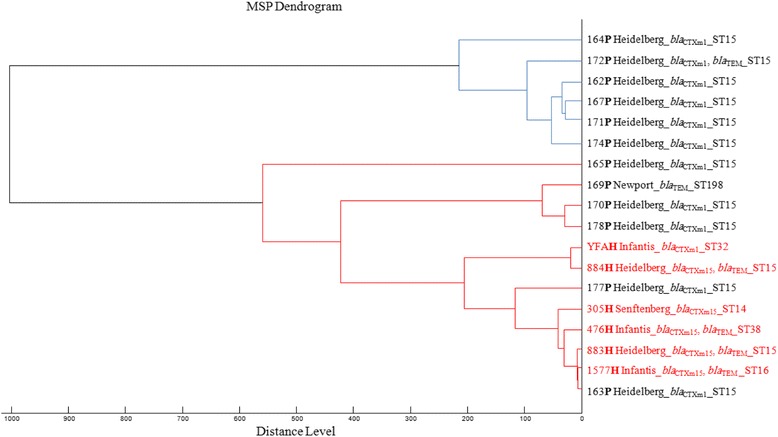
MALDI-TOF MS-based phylogenetic tree of ESBL-positive Salmonella enterica strains from humans (H) and poultry (P)
Discussion
This study involved thirty-two poultry farms and five slaughter houses. It covered most of the districts in the Skikda province. The selection of the sites was based on the managers’ willingness to cooperate with the study and to spend a significant amount of time and effort to perform and collect the various samples. This work provides epidemiological data on Salmonella serotype contamination in poultry farms and slaughterhouses in the region of Skikda, in order to investigate the molecular mechanisms of ß-lactam resistance in avian and human ESBL-producing S. enterica and to analyze the genetic relatedness of avian and human isolates.
The recorded prevalence rates (34.37% for poultry farms and 100% for slaughterhouses) are in accordance with those reported in Constantine and Batna provinces (northeastern Algeria): 36.6 and 60%, respectively [29, 30], but were higher than those reported in several European countries (Italy (9.2%), France (3.4%), Germany (2.7%), Spain (1.02%)) and Morocco (24%) [31–33]. This high prevalence can be attributed to the absence of a Salmonella infection-control plan (especially in healthy chicken flocks) [31] and to the poor hygienic state in poultry farms, where Salmonella can persist during several grow-outs [34]. Furthermore, our findings are in accordance with those of Gardel et al. (2003), who found that samples taken at the third week of the grow-out to be more contaminated by Salmonella spp. than those collected at advanced ages [35]. According to many surveys, contamination of poultry products with Salmonella may take place at different stages of the production process [36]. After contamination of the birds at the farm, bacteria colonize their intestines and can infect their carcasses at slaughter [37].
Elgroud et al. [29] found that 73.3% of the poultry slaughterhouses in Constantine were Salmonella-positive [29]. Among the avian serotypes isolated in our study, Kentucky was the most predominant. Currently, this serotype is distributed worldwide, especially its ST198 [3]. Salmonella ser. Heidelberg has been isolated from broiler carcasses collected at four provinces in the center of Algeria [38]. Despite the fact that we have isolated Salmonella ser. Enteritidis in only 9% of the samples, it is worth noting that this serotype is the most common in animal products, especially poultry [31]. Our Salmonella strains exhibited a high resistance rate to fluoroquinolones, and interestingly, a Kentucky ciprofloxacin-resistant serotype was isolated in France from a patient who had previously stayed in Algeria [39]. In the present study, we report the presence of ESBLs in Salmonella ser. Heidelberg, Senftenberg, Infantis and Newport of avian and human origins, with the CTX-M groups as the most prevalent. To our knowledge, this is the first report of bla CTX-M-1 genes in avian Salmonella strains in Algeria. In fact, this group is the principal ESBL type in human Salmonella encountered in Europe [9]. CTX-M-15 was identified in all human Salmonella strains. This finding corroborates well with several studies performed in Algeria showing that different Salmonella serotypes (Heidelberg, Kedougou, Infantis and Enteritidis) isolated from humans harbored this gene [40–43].
The present study had demonstrated the presence of bla TEM genes in one avian Salmonella ser. Newport, one avian Salmonella ser. Heidelberg strain and all human ESBL-positive strains. Olesen et al. (2004) reported that in Denmark that the major recorded ESBL was the TEM group [44].
In Algeria, cephalosporin use is uncommon in poultry production [22], and the fact that ESBL-positive Salmonella strains of avian origin were isolated suggest that these resistant strains may have been introduced into the poultry production chain from other sources, or resulted from the acquisition by avian Salmonella strains of ESBL resistance determinants that are generally carried on mobile genetic elements (such as plasmids) [45].
The fact that seven avian and human Salmonella ser. Heidelberg strains were of the same ST and clustered together suggests that this clone is circulating in the poultry production chain as well as in the human community. Chicken-to-human transmission of Salmonella during farming has been widely demonstrated. It may take place through the food chain [46], and also through occupational exposure from direct contact with live animals and their environment in the broiler chicken industry [47].
Conclusion
The results of this study demonstrate that Salmonella contamination is highly prevalent in broiler poultry farms and slaughterhouses in the region of Skikda (northeastern Algeria), with an increasing resistance to medically important antibiotics. To the best of our knowledge, our results present for the first time, the emergence of ESBL-producing S. enterica isolates in poultry in this region. In addition, despite the different sampling times of avian and human Salmonella strains, their relatedness has been clearly demonstrated. The clonal relationship between human and avian strains indicates that the poultry industry may act as an important reservoir for ESBL-producing Salmonella that are transmitted to humans by direct contact or essentially through the food chain, but more discriminatory typing methods may be able to add more information as to the epidemiology of ESBL-producing Salmonella strains in Algeria. Infections caused by multidrug-resistant Salmonella species and therapeutic failures increase the risk of death. This is why surveillance programs, rational use of antibiotics and strict biosecurity measures have to be implemented in order to identify the sources, the exact routes of bacterial transmission and to limit the spread of these health-threatening bacteria in the local and national poultry industries.
Additional files
Main characteristics of patients with the corresponding serotype and Salmonella strain ID n°. (DOC 77 kb)
Results of antibiotics sensitivity testing of poultry and human strains. (XLS 39 kb)
Acknowledgments
The authors would like to thank the poultry farmers and slaughterhouses managers for their precious cooperation in collecting samples at both sites and C. Belkader and the entire staff of the bacteriology laboratory at the Pasteur Institute of Algeria for their technical assistance. Prof. C. Bentchouala (Microbiology Laboratory- University Teaching Hospital of Constantine) is highly respectful for providing the human Salmonella strains used in this study. The authors also thank TradOnline for corrections to the English language manuscript.
Funding
This work was partly funded by the CNRS and IHU Méditerranée Infection.
Availability of data and materials
The dataset supporting the conclusions of this article is included in the [zenodo database] repository [https://zenodo.org/record/50346], DOI 10–5281/zenodo 50,346, Datasets Open access, License (for files):Creative Commons CCZero.
Authors’ contributions
SD has actively worked on the isolation of avian Salmonella strains, identification of strains by mass spectrometry and their characterization (antibiotic-susceptibility testing, molecular typing of ESBL genes, sequencing, clonality assessment and Multilocus sequence typing MLST), data interpretation, drafting the paper and revising it. SB performed the data analysis, including sequence checking, Dendrogram of ESBL-positive Salmonella strains and revision of the article. BM, RE, and OB conceived and designed the study. AA participated actively in data interpretation, drafting the paper and critically revising it. SH contributed actively to the draft writing and revision of the article. SC participated in strains analysis (Identification by mass spectrometry and PCR characterization of human ESBL-positive Salmonella strains). KR contributed in part to the study design and data analysis (Serotyping and antibiotic- susceptibility testing of avian strains). JMR conceived and designed the study of ESBL-producing isolates, the clonality testing, antibiotic-susceptibility testing of the strains and contributed to the draft writing and the revision of the article. All the authors read and approved the article.
Competing of interest
The authors declare that they have no conflicts of interest.
Consent for publication
“Not applicable” in this section.
Ethics approval and consent to participate
Prior to sampling, poultry farmers and poultry slaughterhouse owners were contacted and informed about the purpose and the methods of the study to obtain their consent. All of them agreed to participate to this survey. Live bird swabbing methods were approved by thescientific council of the Institute of Veterinary Sciences (University Frère Mentouri, Constantine, Algeria) and comply with the guidelines for animal care and use in research andteaching.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- DDST
Double-disc synergy test
- ESBLs
Extended spectrum β-lactamases
- MALDI-TOF
Matrix-assisted laser desorption/ionization time-of-flight.
- MLST
Multilocus sequence typing
- PCR
Polymerase chain reaction
- PWB
Peptone water broth
- ST
Sequence type
- UTH
University Teaching Hospital
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s12917-017-1050-3) contains supplementary material, which is available to authorized users.
Contributor Information
Samia Djeffal, Email: sam9veto@gmail.com.
Sofiane Bakour, Email: b.sofiane-microbio@hotmail.fr.
Bakir Mamache, Email: mamachebakir@yahoo.fr.
Rachid Elgroud, Email: elgrouddz@yahoo.fr.
Amir Agabou, Email: amirveto@gmail.com.
Selma Chabou, Email: chabouselma@gmail.com.
Sana Hireche, Email: sana_veto@hotmail.fr.
Omar Bouaziz, Email: gspalabo@yahoo.fr.
Kheira Rahal, Email: atb.diag@yahoo.fr.
Jean-Marc Rolain, Phone: (33) 4 91 32 43 75, Email: jean-marc.rolain@univ-amu.fr.
References
- 1.Sanchez S, Hofacre CL, Lee MD, Maurer JJ, Doyle MP. Animal sources of salmonellosis in humans. J Am Vet Med Assoc. 2002;4:492–497. doi: 10.2460/javma.2002.221.492. [DOI] [PubMed] [Google Scholar]
- 2.Majowicz SE, Musto J, Scallan E, Angulo FJ, Kirk M, O'Brien SJ, Jones TF, Fazil A, Hoekstra RM. The global burden of nontyphoidal Salmonella gastroenteritis. Clin Infect Dis. 2010;50:882–889. doi: 10.1086/650733. [DOI] [PubMed] [Google Scholar]
- 3.Le Hello S, Hendriksen RS, Doublet B, Fisher I, Nilsen EM, Whichard JM. International spread of an epidemic population of Salmonella enterica serotype Kentucky ST198 resistant to ciprofloxacin. J Infect Dis. 2011;204:675–684. doi: 10.1093/infdis/jir409. [DOI] [PubMed] [Google Scholar]
- 4.Mekademi K, Saidani K. Contamination of broilers by Salmonella non Typhi in Mitidja. J Curr Res Sci. 2013;4:213–217. [Google Scholar]
- 5.Mouffok F. Situation en matière TIA en Algérie de 2010 à 2011. In: Deuxième congrés magrébin sur les TIA Tunis; 2011. http://mail.pasteur.dz/pasteur-old-2048/presentation.php?page=72. Accessed 26 June 2015.
- 6.Refregier-Petton J, Kemp GK, Nebout JM, Allo JC, Salvat G. Post treatment effects of a SANOVA immersion treatment on turkey carcases and subsequent influence on recontamination and cross contamination of breast fillet meat during turkey processing. Br Poult Sci. 2003;44:790–791. doi: 10.1080/00071660410001666808. [DOI] [PubMed] [Google Scholar]
- 7.Journal officiel de la république Algérienne N°36. Arrêté interministériel de 17 Dhou El Kaada 1423 correspondant au 20 Janvier 2003 définissant les mesures de prévention et de luttes spécifiques au salmonellose aviaires à Salmonella Entéritidis, Typhimurium, Typhi, arizona, dublin, paratyphi et pullorum gallinarum. 2003. http://www.joradp.dz/FTP/jo-francais/2003/F2003036.pdf.Accessed 20 November 2016.
- 8.Murgia M, Bouchrif B, Timinouni M, Al-Qahtani A, Al-Ahdal MN, Cappuccinelli P, Rubino S, Paglietti B. Antibiotic resistance determinants and genetic analysis of Salmonella enterica isolated from food in Morocco. Int J Food Microbiol. 2015;215:31–39. doi: 10.1016/j.ijfoodmicro.2015.08.003. [DOI] [PubMed] [Google Scholar]
- 9.Livermore DM, Canton R, Gniadkowski M, Nordmann P, Rossolini GM, Arlet G, Ayala J, Coque TM, Kern-Zdanowicz I, Luzzaro F, Poirel L, Woodford N. CTX-M: changing the face of ESBLs in Europe. J Antimicrob Chemother. 2007;59:165–174. doi: 10.1093/jac/dkl483. [DOI] [PubMed] [Google Scholar]
- 10.Bradford PA. Extended-spectrum beta-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat. Clin Microbiol Rev. 2001;14:933–951. doi: 10.1128/CMR.14.4.933-951.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bush K, Jacoby GA, Medeiros AA. A functional classification scheme for beta-lactamases and its correlation with molecular structure. Antimicrob Agents Chemother. 1995;39:1211–1233. doi: 10.1128/AAC.39.6.1211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Livermore DM. beta-Lactamases in laboratory and clinical resistance. Clin Microbiol Rev. 1995;8:557–584. doi: 10.1128/cmr.8.4.557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cantón R, Coque TM. The CTX-M beta-lactamase pandemic. Curr Opin Microbiol. 2006;9:466–475. doi: 10.1016/j.mib.2006.08.011. [DOI] [PubMed] [Google Scholar]
- 14.Paterson DL, Bonomo RA. Extended-spectrum beta-lactamases: a clinical update. Clin Microbiol Rev. 2005;18:657–686. doi: 10.1128/CMR.18.4.657-686.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rahal K, Reghal A. A nosocomial epidemic of Salmonella Mbandaka which produces various broad spectrum beta-lactamases: preliminary results. Med Trop. 1994;54:227–230. [PubMed] [Google Scholar]
- 16.Bulletin Sanitaire vétérinaire, Ministère de l’agriculture et du développement rural, Algérie. Avril 2011. http://www.fao.org/fileadmin/user_upload/remesa/library/Bulletin%20Sanitaire%20Vétérinaire%20Avril%202011.pdf. Accessed 20 November 2016.
- 17.Norme française NF U47-101 . Isolement et identification de tout sérovar ou de sérovar(s) spécifié(s) de salmonelles chez les oiseaux. France: Editions AFNOR; 2005. p. 34. [Google Scholar]
- 18.ISO.(Comité international de normalisation AW/9 ISO6579) Microbiology General Guidance on methods for the detection of Salmonella −2002/Amendment 1, Annex D: Detection of Salmonella spp. in animal feces and in environmental samples from the primary production stage. 2007.
- 19.Seng P, Drancourt M, Gouriet F, La SB, Fournier PE, Rolain JM, Raoult D. Ongoing revolution in bacteriology: routine identification of bacteria by matrix-assisted laser desorption ionization time-of-flight mass spectrometry. Clin Infect Dis. 2009;49:543–551. doi: 10.1086/600885. [DOI] [PubMed] [Google Scholar]
- 20.Guibourdenche M, Roggentin P, Mikoleit M, Fields PI, Bockemuhl J, Grimont PA, Weill FX. Suppl 47 2003-2007 to the White-Kauffmann-Le Minor scheme. Res Microbiol. 2010;161:26–29. doi: 10.1016/j.resmic.2009.10.002. [DOI] [PubMed] [Google Scholar]
- 21.EUCAST. (European Comitee on Antimicrobial Suceptibility Testing) Breakpoint tables for interpretation of MICs and zone diameters. Version 5.0, 2015. http://www.eucast.org. Accessed 10 Mar 2016.
- 22.SAEN (ED). Standardisation de l'antibiogramme à l'échelle nationale (médecine vétérinaire et humaine), Algérie. 2011.
- 23.Roschanski N, Fischer J, Guerra B, Roesler U. Development of a Multiplex Real-Time PCR for the Rapid Detection of the Predominant Beta-Lactamase Genes CTX-M, SHV,TEM and CIT-Type AmpCs in Enterobacteriaceae. PLoS One. 2014;9:e100956. doi: 10.1371/journal.pone.0100956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Edelstein M, Pimkin M, Palagin I, Edelstein I, Stratchounski L. Prevalence and molecular epidemiology of CTX-M extended-spectrum beta-lactamase producing Escherichia coli and Klebsiella pneumoniae in Russian hospitals. Antimicrob Agents Chemother. 2003;47:3724–3732. doi: 10.1128/AAC.47.12.3724-3732.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kruger T, Szabo D, Keddy KH, Deeley K, Marsh JW, Hujer AM, Bonomo RA, Paterson DL. Infections with non typhoidal Salmonella species producing TEM-63 or a novel TEM enzyme, TEM-131, in South Africa. Antimicrob Agents Chemother. 2004;48:4263–42670. doi: 10.1128/AAC.48.11.4263-4270.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Yagi T, Kurokawa H, Shibata N, Shibayama K, Arakawa Y. A preliminary survey of extended-spectrum-lactamase (ESBLs) in clinical isolates of Klebsiella pneumoniae and Escherichia coli in Japan. FEMS Microbiol Lett. 2015;184:53–56. doi: 10.1111/j.1574-6968.2000.tb08989.x. [DOI] [PubMed] [Google Scholar]
- 27.Gupta SK, Padmanabhan BR, Diene SM, Lopez-Rojas R, Kempf M, Landraud L, Rolain JM. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob Agents Chemother. 2014;58:212–220. doi: 10.1128/AAC.01310-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kidgell C, Reichard U, Wain J, Linz B, Torpdahl M, Dougan G, Achman M. Salmonella Typhi, the causative agent of typhoid fever, is approximately 50,000 years old. Infect Genet Evol. 2002;2:39–45. doi: 10.1016/S1567-1348(02)00089-8. [DOI] [PubMed] [Google Scholar]
- 29.Elgroud R, Zerdoumi F, Benazzouz M, Bouzitouna-Bentchouala C, Granier SA, Fremy S, Brisabois A, Dufour B, Millemann Y. Characteristics of Salmonella contamination of broilers and slaughterhouses in the region of Constantine (Algeria) Zoonoses Public Health. 2009;56:84–93. doi: 10.1111/j.1863-2378.2008.01164.x. [DOI] [PubMed] [Google Scholar]
- 30.Ayachi A, Alloui N, Bennoune O, Kassah-Laouar A. Survey of Salmonella serovars in broilers and laying breeding reproducers in East of Algeria. J Infect Dev Ctries. 2010;4:103–106. doi: 10.3855/jidc.562. [DOI] [PubMed] [Google Scholar]
- 31.European Food Safety Authority (EFSA) Scientific Report of EFSA and ECDC the European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2001. EFSA J. 2013;11:3129. doi: 10.2903/j.efsa.2013.3129. [DOI] [Google Scholar]
- 32.Lamas A, Fernandez-No IC, Miranda JM, Vazquez B, Cepeda A, Franco CM. Prevalence, molecular characterization and antimicrobial resistance of Salmonella serovars isolated from northwestern Spanish broiler flocks (2011-2015). Poult Sci 2016;0:1-9. [DOI] [PubMed]
- 33.Chaiba A, Rhazi Filali F. Prévalence de la contamination par Salmonella des élevage. de poulet de chair au Maroc. Cah Agric. 2016;doi:10.1051/cagri/2016017. https://www.cahiersagricultures.fr/articles/cagri/pdf/2016/03/cagri160054.pdf. Accessed 13 July 2016.
- 34.Lahellec C, Colin P. Relationship between serotypes of Salmonellae from hatcheries and rearing farms and those from processed poultry carcases. Br Poult Sci. 1985;26:179–186. doi: 10.1080/00071668508416802. [DOI] [PubMed] [Google Scholar]
- 35.Gradel KO, Rattenborg E. A questionnaire-based, retrospective field study of persistence of Salmonella Enteritidis and Salmonella Typhimurium in Danish broiler houses. Prev Vet Med. 2003;56:267–284. doi: 10.1016/S0167-5877(02)00211-8. [DOI] [PubMed] [Google Scholar]
- 36.Salvat G. Prévention des problèmes de santé publique liés aux produits issus de la filière avicole. Acad Vet France. 1997;70:43–48. [Google Scholar]
- 37.Van Scothorst M, Notermans S. Food-borne diseases associated with poultry in: MEAD, G.C and Freeman, B.M (Editors) Meat quality in poultry and game birds. Edinburgh Brit Poultry Sci Ltd. 1980;90:79–90. [Google Scholar]
- 38.Bounar-Kechih S, Hamdi TM, Mezali L, Assaous F, Rahal K. Antimicrobial resistance of 100 Salmonella strains isolated from Gallus gallus in 4 wilayas of Algeria. Poult Sci. 2012;91:1179–1185. doi: 10.3382/ps.2011-01620. [DOI] [PubMed] [Google Scholar]
- 39.Weill FX, LE Hello S. Rapport d'activité annuel Centre National de Référence de Salmonella CNR. 2011. [Google Scholar]
- 40.Kermas R, Touati A, Brasme L, Le Magrex-Debar E, Mehrane S, Weill FX. De champs C. Characterization of extended-spectrum beta-lactamase-producing Salmonella enterica serotype Brunei and Heidelberg at the Hussein Dey hospital in Algiers (Algeria) Foodborne Pathog Dis. 2012;9:803–808. doi: 10.1089/fpd.2012.1159. [DOI] [PubMed] [Google Scholar]
- 41.Touati A, Benallaoua S, Gharout A, Amar AA, Le Magrex DE, Brasme L, Madoux J. De champs C, Weill FX. First report of CTX-M-15 in Salmonella enterica serotype Kedougou recovered from an Algerian hospital. Pediatr Infect Dis J. 2008;27:479–480. doi: 10.1097/INF.0b013e31816a35bd. [DOI] [PubMed] [Google Scholar]
- 42.Naas T, Bentchouala C, Cuzon G, Yaou S, Lezzar A, Smati F, Nordmann P. Outbreak of Salmonella enterica serotype Infantis producing ArmA 16S RNA methylase and CTX-M-15 extended-spectrum beta-lactamase in a neonatology ward in Constantine. Algeria Int J Antimicrob Agents. 2011;38:135–139. doi: 10.1016/j.ijantimicag.2011.04.012. [DOI] [PubMed] [Google Scholar]
- 43.Bouzidi N, Aoun L, Dekhil M, Granier SA, Poirel L, Brisabois A, Nordmann P, Millemann Y. Co-occurrence of aminoglycoside resistance gene armA in non-Typhi Salmonella isolates producing CTX-M-15 in Algeria. J Antimicrob Chemother. 2011;66:2180–2181. doi: 10.1093/jac/dkr237. [DOI] [PubMed] [Google Scholar]
- 44.Olesen I, Hasman H, Aarestrup FM. Prevalence of beta-lactamases among ampicillin-resistant Escherichia coli and Salmonella isolated from food animals in Denmark. Microb Drug Resist. 2004;10:334–340. doi: 10.1089/mdr.2004.10.334. [DOI] [PubMed] [Google Scholar]
- 45.Bae D, Cheng CM, Khan AA. Characterization of extended-spectrum beta-lactamase (ESBL) producing non-typhoidal Salmonella (NTS) from imported food products. Int J Food Microbiol. 2015;214:12–17. doi: 10.1016/j.ijfoodmicro.2015.07.017. [DOI] [PubMed] [Google Scholar]
- 46.Currie A, MacDougall L, Aramini J, Gaulin C, Ahmed R, Isaacs S. Frozen chicken nuggets and strips and eggs are leading risk factors for Salmonella Heidelberg infections in Canada. Epidemiol Infect. 2005;133:809–816. doi: 10.1017/S0950268805004383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kim A, Lee YJ, Kang MS, Kwag SI, Cho JK. Dissemination and tracking of Salmonella spp. in integrated broiler operation. J Vet Sci. 2007;8:155–161. doi: 10.4142/jvs.2007.8.2.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Main characteristics of patients with the corresponding serotype and Salmonella strain ID n°. (DOC 77 kb)
Results of antibiotics sensitivity testing of poultry and human strains. (XLS 39 kb)
Data Availability Statement
The dataset supporting the conclusions of this article is included in the [zenodo database] repository [https://zenodo.org/record/50346], DOI 10–5281/zenodo 50,346, Datasets Open access, License (for files):Creative Commons CCZero.