

Abstract
How bacteria grow and divide while retaining a defined shape is a fundamental question in microbiology, but technological advances are now driving a new understanding of how the shape-maintaining bacterial peptidoglycan sacculus grows. In this Review, we highlight the relationship between peptidoglycan synthesis complexes and cytoskeletal elements, as well as recent evidence that peptidoglycan growth is regulated from outside the sacculus in Gram-negative bacteria. We also discuss how growth of the sacculus is sensitive to mechanical force and nutritional status, and describe the roles of peptidoglycan hydrolases in generating cell shape and of D-amino acids in sacculus remodelling.
Bacteria come in a range of shapes (such as cocci and rods), and their internal volume ranges from ~10−2 to ~106 μm3 (REF 1). Importantly, however, cells of any given species are rather uniform in shape and size during vegetative growth. Therefore, growing bacteria must have robust mechanisms to maintain their shape and pass it on to their progeny. How bacteria achieve this remains a fundamental question in microbiology.
The peptidoglycan sacculus maintains cell shape and provides mechanical strength to resist osmotic challenges2. The mesh-like sacculus surrounds the cytoplasmic or inner membrane and is composed of glycan chains crosslinked by short peptides. Growth of the sacculus is a dynamic process requiring synthases to make peptidoglycan and attach it to the existing sacculus, and presumably hydrolases to cleave the sacculus to allow insertion of the newly synthesized material3. Because these enzymes are several orders of magnitude smaller than the sacculus itself, their activity must be spatiotemporally controlled so that insertion of new material maintains the cell shape. This Review focuses on our current understanding of sacculus growth, primarily in Gram-negative bacteria, and its relationship to bacterial growth and morphogenesis.
Peptidoglycan synthesis and hydrolysis
Synthesis of peptidoglycan occurs in three overall stages (FIG. 1). First, soluble, activated nucleotide precursors (UDP-N-acetylglucosamine and UDP-N-acetylmuramyl pentapeptide) are synthesized in the cytoplasm4. Second, at the inner leaflet of the inner membrane, the nucleotide precursors are assembled with undecaprenyl phosphate to form the lipid-anchored disaccharide-pentapeptide monomer subunit (lipid II), and are flipped across the membrane5,6. Third, lipid II is polymerized, releasing undecaprenyl pyrophosphate, and the resulting glycan chains are inserted into the sacculus. Peptidoglycan synthesis and insertion are guided by distinct elements of the bacterial cytoskeleton at different phases in the cell cycle. With the cooperation of actin-like rod-shapedetermining protein MreB, newly divided cells elongate by inserting peptidoglycan into multiple sites in the lateral wall of the cell (‘dispersed’ elongation). Later, the tubulin-like cell division protein FtsZ localizes to the midcell to guide a ‘preseptal’ phase of cell elongation, followed by ‘constrictive’ septum synthesis, which enables cell division and daughter cell separation. TABLE 1 summarizes the established peptidoglycan enzymes and cell morphogenesis proteins of Escherichia coli.
Figure 1. Peptidoglycan synthesis and cleavage.
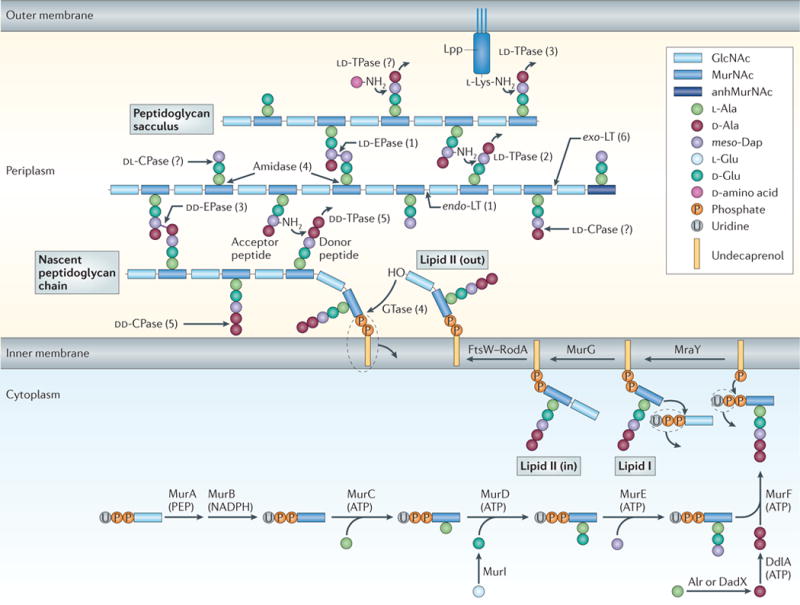
The synthesis and attachment of a new peptidoglycan strand to the existing sacculus, with particular emphasis on the different synthetic and degrading enzymes. Precursors are synthesized in the cytoplasm, linked to the transport lipid (undecaprenyl phosphate) and flipped accross the inner membrane by FtsW–RodA. A glycosyltransferase (GTase) catalyses polymerization of a nascent peptidoglycan chain from lipid II precursor at the inner membrane, followed by attachment of the new chain to the sacculus by a DD-transpeptidase (DD-TPase). Peptides are trimmed by DD-, LD-and DL-carboxypeptidases (CPases), and crosslinks are cleaved by the DD-and LD-endopeptidases (EPases). Amidases remove peptides from glycan chains, and exo-or endo-specific lytic transglycosylases (LTs) cleave in the glycan chain to form 1,6-anhydro-N-acetylmuramic acid (anhMurNAc) residues, which are the hallmark of glycan chain ends. LD-TPases are responsible for the formation of LD-crosslinks, the attachment of the major outer-membrane lipoprotein (Lpp), which is anchored in the outer membrane, and the binding of unusual D-amino acids. The number of known Escherichia coli enzymes for each group is shown in brackets, but this is probably an underestimate, as even in E. coli not all players are known and/or characterized. Alr, Ala racemase, biosynthetic; DadX, Ala racemase, catabolic; DdlA, D-Ala–D-Ala ligase A; GlcNAc, N-acetylglucosamine; meso-Dap, meso-diaminopimelic acid; MraY, UDP-MurNAc-pentapeptide phosphotransferase; MurA, UDP-GlcNAc enolpyruvyl transferase; MurB, UDP-MurNAc dehydrogenase; MurC, UDP-MurNAc–L-Ala ligase; MurD, UDP-MurNAc-L-Ala–D-Glu ligase; MurE, UDP-MurNAc-L-Ala-D-Glu-meso-Dap ligase; MurF, UDP-MurNAc-tripeptide–D-alanyl-D-Ala ligase; MurG, UDP-GlcNAc-undecaprenoyl-pyrophosphoryl-MurNAc-pentapeptide transferase; Murl, Glu racemase; PEP, phosphoenolpyruvate.
Table 1.
Peptidoglycan synthesis enzymes and cell morphogenesis proteins in Escherichia coli
| Function | Activity or category | Proteins* | Relevant features and remarks‡ | |
|---|---|---|---|---|
| Precursor synthesis | Transferase and dehydrogenase, respectively | MurA, MurB | • Synthesis of UDP-MurNAc from UDP-GlcNAc | |
| Amino acid ligases | MurC, MurD, MurE, MurF, Ddl | • Cytoplasmic steps leading to the UDP-MurNAc pentapeptide | ||
| Racemases | Alr, DadX, MurI | • Synthesis of D-Ala or D-Glu from L-Ala or L-Glu, respectively | ||
| GTases | MraY, MurG | • Inner membrane-localized steps of lipid II synthesis from the UDP-MurNAc pentapeptide | ||
| Peptidoglycan synthesis | GTases and DD-TPases (class A PBPs) | PBP1A | • Major peptidoglycan synthase, mainly involved in cell elongation • Anchored in the inner membrane • Interacts with LpoA |
|
| PBP1B | • Major peptidoglycan synthase, mainly involved in cell division • Anchored in the inner membrane • Dimerizes and interacts with PBP3, FtsN, MipA and LpoB • Crystal structure available |
|||
| PBP1C | • Cellular role unknown • Anchored in the inner membrane |
|||
| DD-TPases (class B PBPs) | PBP2 | • Essential for cell elongation • Dependent on MreB filament for localization • Anchored in the inner membrane |
||
| PBP3 | • Essential for cell division • Part of the divisome • Anchored in the inner membrane • Interacts with PBP1B, MtgA, FtsQLB, FtsW and FtsN |
|||
| GTase | MtgA | • Localizes to the division site • Interacts with PBP3, FtsW and FtsN • Anchored in the inner membrane |
||
| Regulation of peptidoglycan synthesis | Activators of peptidoglycan synthase | LpoA, LpoB | • Regulate PBP1A (LpoA) and PBP1B (LpoB) TPase activity • Outer-membrane lipoproteins |
|
| Formation of 3–3 crosslinks | LD-TPases | YnhG,YcbB | • Form the minor type of β-lactam-insensitive peptide crosslinks, the function of which is unknown | |
| Cell envelope stability and the creation of a firm connection between peptidoglycan and the outer membrane | Structural protein | Lpp (Braun’s lipoprotein) | • Outer-membrane lipoprotein • The bound form is covalently attached to peptidoglycan • The free form forms trimers and is embedded in the outer membrane |
|
| LD-TPases | ErfK, YbiS, YcfS | • Attachment of Lpp to peptidoglycan | ||
| Regulation of peptidoglycan structure | DD-CPases (class C PBPs) | PBP5, PBP4B, PBP6, PBP6B | • Proposed regulatory role in peptidoglycan synthesis by removal of excess pentapeptide donors in newly made peptidoglycan | |
| Peptidoglycan hydrolysis (autolysis) | DD-EPases | PBP4, PBP7 | • Septum cleavage (PBP7) • Biofilm formation (PBP7) • Also has DD-CPase activity (PBP4) |
|
| DD-and LD-EPase | MepA | • LAS family metallopeptidase | ||
| LTs | Slt70, MltA, MltB, MltC, MltD, MltE (also known as EmtA), MltF | • Major autolysins • Interact with PBP7 (Slt70) or PBP1B via MipA (MltA) • Septum cleavage (Slt70, MltA, MltB, MltC, MltD) • Outer membrane-anchored lipoproteins (Mlt proteins) |
||
| Amidases | AmiA, AmiB, AmiC, AmiD | • Septum cleavage (AmiA, AmiB and AmiC) | ||
| Regulation of peptidoglycan hydrolysis | Activators of amidases | EnvC, NlpD, YgeR, YebA | • Have a LytM peptidoglycan-binding domain • Activators of AmiA and AmiB (EnvC) or AmiC (NlpD) • The roles of YgeR and YebA are unknown |
|
| Inhibitor of LTs | Ivy | • Inhibitor of MltB | ||
| Cell elongation | Cytoskeletal structure, ATPase, GTPase | MreB | • Actin structural homologue • Forms a cytoplasmic, membrane-attached helix or patches |
|
| MreB-associated proteins | MreC, MreD, RodZ, RodA, PBP2 | • MreB-associated and inner membrane-associated proteins (MreC, MreD and RodZ) • Lipid II flippase (RodA) |
||
| Cell division | Cytoskeletal structure, GTPase | FtsZ | • Tubulin structural homologue • Forms a dynamic cytoplasmic ring structure at midcell |
|
| ‘Early’ association with the Z ring | FtsA, ZipA, ZapA, ZapB, ZapC, FtsE, FtsX, FtsK | • Stabilization and membrane-attachment of FtsZ polymers (FtsA, ZipA, ZapA, ZapB, ZapC) • Recruitment of proteins and DNA transport (FtsK) |
||
| ‘Late’ association with the Z ring | FtsQ, FtsL, FtsB, FtsW, FtsN, PBP3, DamX, DedD, RlpA | • Interactions with peptidoglycan synthases PBP3 (FtsQLB, FtsW and FtsN) and PBP1B (PBP3 and FtsN) • Lipid II flippase (FtsW) • Peptidoglycan binding (FtsN, DamX, DedD and RlpA) |
||
| Outer-membrane invagination | TolQ, TolR, TolA, TolB, Pal | • Form an envelope-spanning complex for outer-membrane invagination during septation • Peptidoglycan binding (Pal) |
||
Alr, Ala racemase, biosynthetic; CPase, carboxypeptidase; DadX, Ala racemase, catabolic; Ddl, D-Ala–D-Ala ligase; EPase, endopeptidase; GlcNAc, N-acetlyglucosamine; GTase, glycosyltransferase; Ivy, inhibitor of vertebrate lysozyme; LT, lytic transglycosylase; Mlt, membrane-bound lytic murein transglycosylase; MraY, UDP-MurNAc-pentapeptide phosphotransferase; MurA, UDP-GlcNAc enolpyruvyl transferase; MurB, UDP-MurNAc dehydrogenase; MurC, UDP-MurNAc–L-Ala ligase; MurD, UDP-MurNAc-L-Ala–D-Glu ligase; MurE, UDP-MurNAc-L-Ala-D-Glu–meso-diaminopimelic acid ligase; MurF, UDP-MurNAc-tripeptide–D-alanyl-D-Ala ligase; MurG, UDP-GlcNAc-undecaprenoyl-pyrophosphoryl-MurNAc-pentapeptide transferase; MurI, Glu racemase; MurNAc, N-acetylmuramic acid; PBP, penicillin-binding protein; TPase, transpeptidase. *Proteins were assigned to one category, although many of them would fit into more than one category. The DD-TPases (class B PBPs) were added to peptidoglycan synthesis and to cell elongation (PBP2) or cell division (PBP3) to illustrate their specific functions in the cell cycle. ‡References are given in the main text.
Peptidoglycan synthases
Peptidoglycan synthesis requires glycosyltransferases (GTases) to polymerize the glycan chains and DD-transpeptidases (DD-TPases) to crosslink the peptides7 (FIG. 1). TPases, also called penicillin-binding proteins (PBPs), were initially identified because of their ability to covalently bind penicillin8. There are three types of these peptidoglycan synthases: bifunctional GTase–TPases (the class A PBPs), monofunctional TPases (the class B PBPs) and monofunctional GTases7. E. coli has three bifunctional synthases (PBP1A, PBP1B and PBP1C), a GTase (MgtA) and two TPases (which are essential either for cell elongation (PBP2) or for cell division (PBP3; also known as FtsI)). PBP1A and PBP1B are partially redundant; the cell requires one of them for viability9. PBP1C may be used in host cells10. PBP1A, which is thought to have a role in cell elongation, interacts with the elongation TPase PBP2 (M.B. and WV., unpublished observations). PBP1B interacts with two essential division proteins (FtsN and the division TPase PBP3) and is enriched at the septum11.
The TPase and GTase activities of bifunctional synthases have been reconstituted in vitro with their lipid II substrate12,13. Under conditions that favour PBP1B dimerization, this synthase produces glycan chains of ~28 disaccharide units on average, crosslinking about 40–50% of the peptides. PBP1A produces shorter glycan chains (~20 disaccharide units) and crosslinks ~22% of the peptides. Importantly, although glycan chain polymerization occurs in the absence of transpeptidation (for example, in the presence of penicillin), efficient transpeptidation requires ongoing GTase reactions12,13. It was suggested, based on the crystal structure of PBP1B, that the growing glycan chain produced by the GTase domain moves towards the TPase domain, enabling the pentapeptide of the glycan chain to enter the active site of the TPase domain and serve as a donor for transpeptidation14. Surprisingly, crystal structures of peptidoglycan TPases do not show any obvious binding site for the acceptor peptide, indicating that transpeptidation, the target of β- lactam antibiotics, is not fully understood at the molecular level15.
Peptidoglycan hydrolases
Simply attaching new material to the sacculus by transpeptidation thickens the sacculus without elongating it, suggesting that peptidoglycan growth requires the cleavage of covalent bonds in the sacculus to allow the newly attached material to insert into the layer without increasing its thickness. Peptidoglycan cleavage is also required for reductive cell division and cell separation. E. coli has at least 13 periplasmic peptidoglycan hydrolases ( autolysins), which can collectively cleave almost any glycoside and amide bond16 (FIG. 1). Studies in a range of bacteria have indicated that hydrolases sculpt the shape, size and thickness of peptidoglycan and are essential for separation of daughter cells during cell division or after it is completed (see below). However, possibly owing to high redundancy, no single hydrolase gene knockout prevents growth of E. coli, and multiple hydrolase genes have to be deleted for chains of non-separated cells to form. Amidases have a prominent role in septum cleavage17, but lytic transglycosylases and endopeptidases also contribute to cell separation18,19, and their role is probably understated owing to their greater redundancy. In E. coli, as much as 40–50% of the total peptidoglycan material is removed every generation by the action of peptidoglycan hydrolases, and lytic transglycosylases are major contributors to this process20. The soluble fragments that are removed from the sacculus are reused via an efficient peptidoglycan-recycling pathway21.
Sacculus growth
Biophysical and electron cryotomography (ECT) data on isolated E. coli sacculi support a singlelayer model of peptidoglycan architecture, with somewhat disordered glycan chains running perpendicular to the long axis of the cell, on average22. Enlargement of the peptidoglycan requires new glycan chains to attach and insert while peptidoglycan thickness and cell shape are maintained. Labelling experiments have confirmed that peptidoglycan grows by the insertion of newly synthesized glycan chains, or patches of them, into the existing sacculus23,24. The absence of unattached oligomeric intermediates of nascent glycan chains in the cell suggests that the polymerization of new glycan chains and their attachment to the sacculus by transpeptidation occur simultaneously25, which fits with the existence of bifunctional enzymes or enzyme complexes that perform both reactions. The attachment of newly made peptidoglycan to sacculi by transpeptidation has been reconstituted in vitro with the bifunctional synthase PBP1A13. The newly synthesized peptidoglycan is rich in pentapeptides and has glycan chains with an average length of ~50–60 disaccharides. Shortly after synthesis, the pentapeptides are trimmed to tetrapeptides by DD-carboxypeptidases; some tetrapeptides are further shortened to tripeptides by LD-carboxypeptidases. In addition, lytic transglycosylases reduce the average length of glycan chains to ~35–40 disaccharides, and LD-TPases covalently attach major outer-membrane lipoprotein Lpp (also known as Braun’s lipoprotein) to the sacculus24 (FIG. 1).
The mechanism for insertion of new glycan chains remains unsettled. During cell division, when peptide crosslinks are formed exclusively between newly synthesized glycan chains, about one-third of the new septal peptidoglycan is removed26, consistent with the model that peptidoglycan grows by simultaneously inserting three new chains and removing one old one (the ‘3 for 1’ model3). It is less clear whether this model applies to lateral cell wall growth during cell elongation, when peptide crosslinks seem to form predominantly between new and old chains and peptidoglycan turnover is low21,23,26,27.
The cytoskeleton and peptidoglycan growth
Our understanding of the interplay between bacterial cytoskeletal elements and peptidoglycan growth is evolving (FIG. 2). For instance, there is an increasing appreciation of the diversity of bacterial cytoskeletal elements and the parts that they play in force generation and sensing cellular states. For more information on bacterial cytoskeletal elements, the reader is referred to an excellent recent review28.
Figure 2. Different peptidoglycan synthesis complexes are active at different stages of the Escherichia coli cell cycle.
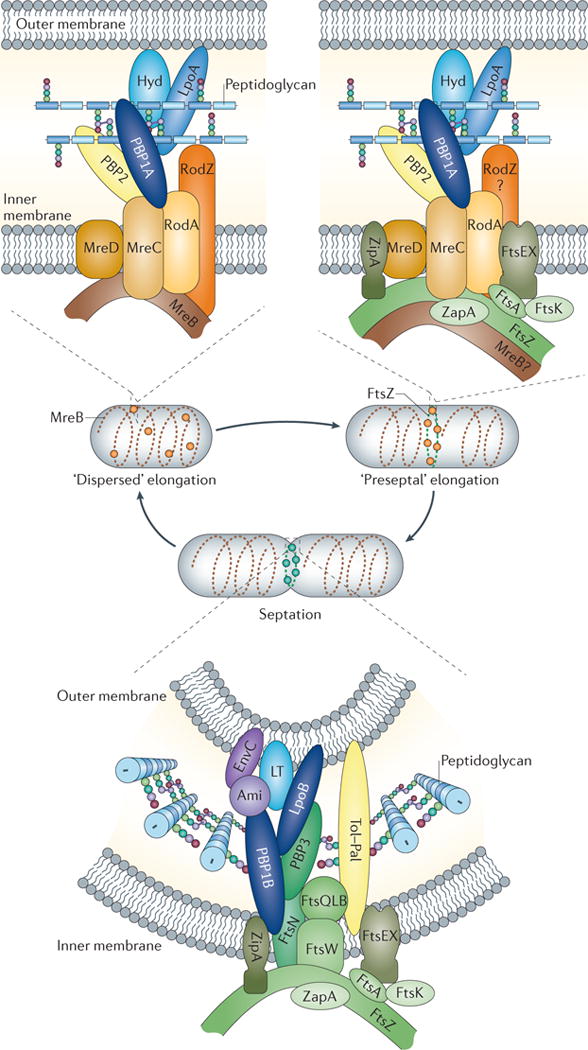
As shown in the upper left panel, MreB and associated membrane proteins control or position the peptidoglycan synthases penicillin-binding protein 1A (PBP1A) and PBP2, as well as still-unknown hydrolases (Hyd), during the ‘dispersed’ mode of elongation. As illustrated in the upper right panel, FtsZ and other early cell division proteins control the elongation-specific peptidoglycan synthesis complex during a ‘preseptal’ mode of elongation. It is not known whether MreB and associated proteins participate in preseptal elongation. Finally, as depicted in the lower panel, the cell division complex contains essential, inner membrane-localized cell division proteins, the peptidoglycan synthases PBP1B and PBP3, and amidase enzymes (Ami) with their activators, as well as proteins of the Tol–Pal complex for constriction of the outer membrane. Activity of the PBPs is regulated in part by outer membrane-anchored lipoproteins such as LpoA and LpoB. LT, lytic transglycosylase.
MreB and peptidoglycan synthesis during cell elongation
The actin-like protein MreB is used by many rod-shaped bacteria in their elongation mode of peptidoglycan synthesis. MreB forms filaments29–31 and interacts with the conserved inner-membrane proteins MreC, MreD and RodZ32–36, as well as with the lipid II synthesis enzymes MraY and MurG37. The MreB filament is tethered to the inner membrane via an interaction with the cytoplasmic domain of RodZ38. E. coli MreB also binds directly to the inner membrane through an N-terminal amphipathic helix39. When MreB is depleted, or filament formation is specifically inhibited (for example, with A22 (S-(3,4-dichlorobenzyl)isothiourea)), cells rapidly stop elongating, increase their diameter and grow with spherical morphology, indicating that MreB is required for rod-shaped growth40–42.
MreB was initially thought to form dynamic helical structures spanning the length of the cell30,43. Preliminary evidence of treadmilling-type movement, with polymerization at one end of the helix and depolymerization at the other end, led to the suggestion that polymerization dynamics guide filament movement44–46. However, recent studies have challenged this view. High-resolution imaging indicates that MreB filaments do not span the cell length either in E. coli or in the Gram-positive bacterium Bacillus subtilis. Instead, MreB forms small filament patches that move around the cell circumference perpendicularly to its long axis47–49. Perturbing the ATP cycle of MreB, either by adding the inhibitor A22 (REF. 49) or by mutation of mreB48, does not affect MreB motion, indicating that polymerization might not be the driving force for its movement. By contrast, antibiotic-mediated inhibition of cell wall synthesis rapidly stops filament movement47–49. Interestingly, inhibition or depletion of the specific elongation class B TPases (PBP2 in E. coli, and PBP2A and PBPH in B. subtilis) eliminates filament movement in both organisms, suggesting that peptidoglycan synthesis itself is the motor that drives filament movement47–49. Consistent with this idea, both MreB and the peptidoglycan synthesis complexes move with comparable velocities in roughly similar circumferential directions47–48, mirroring the position of glycan chains in the sacculus22. Thus, MreB is integral to shape determination, as it directly or indirectly recruits and/or positions peptidoglycan biosynthesis machineries31,50,51, but it cannot move without ongoing peptidoglycan synthesis. Instead, both MreB filaments and peptidoglycan complexes move as a coordinated unit in the direction of the glycan chains, at least in a short range, and are functionally interdependent. It remains to be determined whether such short-range coordinated movements contribute to a coherent rod shape. Interestingly, ovococci, such as Streptococcus pneumoniae, lack MreB but retain MreC and MreD, which are essential; recent genetic evidence suggests that MreC and MreD control peripheral peptidoglycan synthesis by affecting the activity or localization of PBP1A52.
FtsZ and peptidoglycan synthesis
FtsZ, a tubulin-like protein, is the master regulator of bacterial cell division. Prior to cell division, FtsZ forms a dynamic ringlike structure at midcell, called the Z ring. The Z ring consists of arches of bundles of FtsZ filaments that rapidly polymerize and depolymerize on binding and hydrolysis of GTP, a process that is regulated by numerous effectors53. More than ten essential cell division proteins localize to the Z ring to form the divisome54. Cells lacking functional FtsZ are unable to divide and instead grow as filaments.
In E. coli, the first step of divisome assembly begins before constriction, when early cell division proteins, including FtsZ, FtsA and ZipA, localize to the future division site55 (FIG. 2), and ZipA and the actin-like protein FtsA interact with and stabilize the Z ring at the inner membrane. Enzymes for lipid II synthesis, and presumably peptidoglycan synthesis, are recruited to the Z ring to carry out preseptal elongation7,56. Notably, this stage of synthesis is independent of the TPase PBP3, an essential late divisome protein required for constrictive peptidoglycan synthesis57. Preseptal peptidoglycan synthesis contributes substantially to cell elongation in Caulobacter crescentus and less so in E. coli56,57.
When assembly is complete, the divisome synthesizes the new cell poles, including the peptidoglycan layer of the daughter cells (FIG. 2). PBP3, a core member of the divisome, is essential for septal peptidoglycan synthesis. It is recruited to the septum by the lipid II flippase FtsW6,58 and interacts with the FtsQLB complex59,60, PBP1B11 and FtsN61. Although FtsN has a central role in triggering septation and stabilizing the divisome ring62, a hyperactive FtsA can bypass some of its effects63. The carboxy-terminal peptidoglycan-binding SPOR domain of FtsN may contribute to its midcell localization via specific recognition of septal peptidoglycan64,65. The essential function of FtsN is mediated by a short (35 amino acid) periplasmic region, which does not include its SPOR domain. It is tempting to speculate that the essentiality of FtsN is related to its interactions with the peptidoglycan synthases PBP1B and/or PBP3 (REF 66).
Although most core divisome components are widely conserved, their order of assembly may differ across species. For example, in C. crescentus, FtsA arrives at midcell well after FtsZ and the FtsEX complex, and shortly before FtsN, FtsQ, PBP3, FtsK (the DNA translocase) and FtsL. Septation occurs only after FtsW and FtsB arrive last at midcell67.
The role of peptidoglycan hydrolases in septation is discussed below. How the cell controls the switch from dispersed to preseptal peptidoglycan elongation and from preseptal to septal peptidoglycan synthesis is unknown, but it probably involves changes in the localization and dynamics of the cytoskeletal-like elements.
Additional classes of cytoskeleton-like elements
Cytoskeletal elements with similarity to eukaryotic intermediate filaments (IFs) have been recently identified via ECT68,69 and seminal C. crescentus studies70,71. Crescentin (CreS) is anchored in the inner membrane through MreB70 and has a role in generating curvature in C. cresentus72. The IF-like protein, FilP, is required for hyphal mechanical strength in the Gram-positive bacterium Streptomyces coelicolor73.
Bactofilins are a new class of cytoskeletal element. In C. crescentus, they form a sheet-like polymer that lines the inner membrane of the stalked cell pole and recruits the synthase PBPC to nucleate stalk biogenesis. Bactofilins are widely conserved and associate spontaneously in ribbon-or rod-like filaments in vitro74. Bacterofilins participate in diverse processes: cell division in Shewanella oneidensis74, social motility and colony morphology in Myxococcus xanthus74,75, and helical shape formation in Helicobacter pylori (specifically, the protein curved cell morphology A (CcmA; encoded by the locus HPG27_1480)76). The extent to which these cytoskeletal elements are scaffolds for coordinating peptidoglycan synthesis and hydrolysis remains to be addressed.
Cytoskeletal elements and mechanical control of growth
When transient mechanical forces bend a rod-shaped bacterial cell, the bent cell straightens after the force is removed77. However, the outcome is quite different when cells experience constant mechanical force from one side during growth. Filamentous E. coli cells growing in a curved cell shape along microchamber walls retain their bent cell shape when removed from the microchamber. Thus, a long-term mechanical force on one side of the cell makes the cells produce a curved peptidoglycan sacculus during growth78. This experiment demonstrates that mechanical force can affect the topology of peptidoglycan synthesis and bacterial morphogenesis, just as mechanical force can affect growth and shape of eukaryotic organisms79. Here, we consider how forces exerted by cytoskeletal elements alter the mechanical properties of the cell and affect peptidoglycan growth (FIG. 3).
Figure 3. Force generation by cytoskeletal elements.
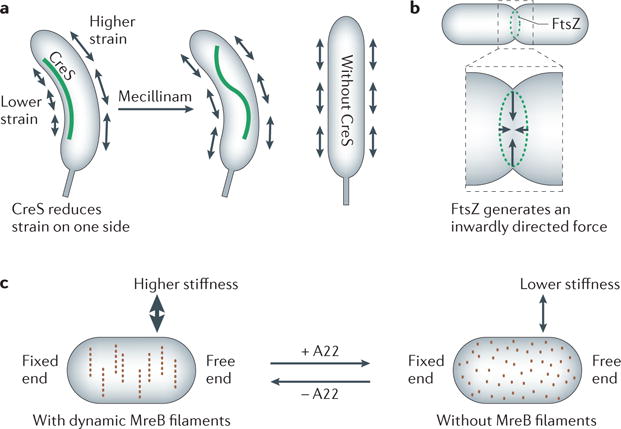
a | Crescentin (CreS) reduces the strain at one side of the cell, causing Caulobacter crescentusto grow in a bent shape. Detachment of the CreS filament from the membrane (on addition of mecillinam) results in rapid loss of the filament’s stretched form but does not cause an instant change in cell shape. Cells lacking CreS grow with a straight shape. b | FtsZ generates an inwardly directed constriction force in vesicle tubes and presumably also in the cell. c | Depolymerization of MreB filaments by addition of the drug A22 (S-(3,4-dichlorobenzyl)isothiourea) reduces the stiffness of Escherichia coli cells.
CreS localizes exclusively to the inner cell curvature by an unknown mechanism involving MreB70,71 (FIG. 3a). By reducing the physical strain on the inner side of the cell wall, CreS reduces the peptidoglycan growth rate on the inner relative to outer side of the sacculus, inducing curvature. CreS is necessary and sufficient for curvature72. A mechanical role for CreS is supported by the fact that the straight rod phenotype of the creS mutant is suppressed by external mechanical force when cells elongate along the walls of a microchamber, and by the observed relaxation of the CreS filament on antibiotic-induced membrane detachment in vivo72 (FIG. 3a). Heterologously expressed CreS also bends E. coli cells72. The rate of straightening of C. crescentus cells following disruption of the CreS filament is related to the processivity of peptidoglycan synthesis enzymes80. Without mechanical force, processive peptidoglycan synthesis in the direction of the glycan chains perpendicular to the long axis may provide a robust mechanism for straight growth of rod-shaped cells80. A biophysical model simulating cellular growth dynamics indicates that robust cell wall synthesis, which is insensitive to local peptidoglycan density, is a key additional contributing feature to rod-shape maintenance81.
Other cytoskeletal elements also exert forces. The cell division protein FtsZ generates a constriction force at the division site of the inner membrane, possibly through bending of the FtsZ protofilaments (FIG. 3b). A membrane-attached version of FtsZ generates constrictions in tubular liposomes in the presence of GTP. Thus, when FtsZ becomes membrane attached, other cellular components are not required to generate force82. Helical MreB filaments also affect the mechanical properties of E. coli. When MreB polymers are disassembled with the antibiotic A22, the bending stiffness of cells, as measured by optical traps, decreases (FIG. 3c). Reformation of the MreB cytoskeleton restores the stiffness77, indicating that force generation by MreB may contribute to cellular mechanical properties. It is possible that the forces generated by the FtsZ-and MreB-containing cytoskeletal structures cause local stretching of the sacculus, promoting peptidoglycan synthesis during cell division and elongation.
Cytoskeleton-like elements provide metabolic inputs
Recent reports suggest that information on the metabolic status of the cell can be conveyed through cytoskeletal elements to the peptidoglycan synthesis machinery. UgtP, a B. subtilis processive diacylglycerol glucosyltransferase, prevents FtsZ assembly when UDP-glucose levels are high. This allows cells in carbon-rich media to grow bigger before they divide83. YvcK, a novel putative B. subtilis metabolic enzyme, builds helical structures that are required for the normal rod-shaped cell morphology and for localization of PBP1 (REF. 84). PBP1 localization is also dependent on MreB, and mreB mutants swell and lyse unless they are supplemented with Mg2+ (REF 50). Interestingly, YvcK and MreB helices (or, more accurately, dynamic patches) build independently of each other and do not overlap, but slight overexpression of either YvcK or MreB compensates for the loss of the other protein, restoring PBP1 localization and cell shape. The physiological role of YvcK is connected with gluconeogenic growth, when it becomes essential for PBP1 localization and the rod shape84. YvcK is also conserved in spherical Gram-positive bacteria and is essential in Staphylococcus aureus85, begging the question of its role in such organisms. Finally, CTP synthase (CtpS; also known as PyrG) forms filaments both in E. coli and C. crescentus. These filaments interact with CreS and inhibit curvature in C. crescentus. Although the CTP synthase activity of CtpS is not required for filament formation, its glutamine amidotransferase activity is absolutely essential86, suggesting that filament formation conveys information on the cellular metabolic status to influence sacculus growth.
Peptidoglycan hydrolases sculpt the cell
Peptidoglycan hydrolases have intimate roles in peptidoglycan growth, cell division and bacterial shape. Although these hydrolases are usually highly redundant, recent studies in a wide range of bacteria are beginning to uncover their diverse roles in the cell.
Role of DD-carboxypeptidases
E. coli has at least six DD-carboxypeptidases, but PBP5 (also known as DacA) is the most active. Cells lacking PBP5 have slightly altered morphology; when additional PBPs are removed, the cells branch, kink and bend87,88. Guided by its membrane anchor and the availability of its pentapeptide substrate, PBP5 localizes to sites of peptidoglycan synthesis89, where it removes terminal D-Ala residues from pentapeptides and reduces the number of donors for transpeptidation (FIG. 1). DD-carboxypeptidases are important shape regulators in E. coli but have only a minor role in other species, such as C. crescentus90, which must possess peptidoglycan growth mechanisms that permit the enlargement of a pentapeptide-rich sacculus.
Roles of the amidases and M23–LytM endopeptidases
The three N-acetylmuramyl-L-Ala amidases in E. coli, AmiA, AmiB and AmiC, are important for septation and cell separation. Double and triple amidase mutants form cell chains in which individual cells constrict their inner membrane and peptidoglycan but not their outer membrane, resulting in abnormally large periplasmic spaces17. These cells also exhibit unusually high outer-membrane permeability19. Although AmiB and AmiC are septally localized, AmiA is not91. Interestingly, cells with only AmiA can still partially separate17.
The endopeptidases of the M23–LytM family are also implicated in septation in E. coli, as a quadruple knockout of all E. coli LytM paralogues (EnvC, NlpD, YgeR and YebA) is defective in cell separation92. The two characterized members, EnvC and NlpD, are catalytically inactive but, instead, are the first proteins shown to regulate amidase activity. NlpD activates AmiC, and EnvC activates AmiA and AmiB, both by as-yet-unknown mechanisms93. Transient interaction of AmiA with EnvC, coupled with its activation, is likely to be sufficient for septal cleavage, thereby explaining why non-septally localized AmiA can function as the sole amidase. Interestingly, EnvC and NlpD localize to the septum earlier than their cognate amidases, which require active PBP3 for localization, thereby ensuring that peptidoglycan synthesis initiates before the amidases concentrate at the septum94.
In contrast to the regulatory role of the characterized E. coli LytM family peptidases, in C. crescentus the LytM peptidase, DipM, is essential for cell constriction95–97. DipM locates to midcell through an interaction with FtsN and is then likely to bind septal peptidoglycan via its four LysM peptidoglycan-binding domains96. The LytM peptidase domain is required for septum cleavage96,97 and may have peptidoglycan hydrolase activity itself96. Remarkably, compared with wild-type cells, dipM mutants have a peptidoglycan layer of 2–3-fold greater average thickness and exhibit outer-membrane blebbing95, indicating that the hydrolase activity of DipM is involved in a mechanism to maintain constant peptidoglycan thickness in C. crescentus.
The H. pylori LytM paralogues, cell shape determinant 1 (Csd1), Csd2 and Csd3, are all required for the helical shape of these cells, as mutants grow as slightly (in the case of csd1 or csd2 mutants) or highly (in the case of csd3 mutants) curved rods, rather than helically76. The increased crosslinkages in the sacculi of these mutant strains supports the proposal that Csd proteins have an endopeptidase activity. Csd3 exhibits both DD-endopeptidase and DD-carboxypeptidase activities, consistent with the higher number of crosslinks and pentapeptides in csd3-mutant cells76,98. Hence, the Csd proteins use their endopeptidase activity to achieve controlled local relaxation of peptidoglycan crosslinks, thereby introducing curvature and twist that leads to the helical cell shape of H. pylori.
Regulation of hydrolase activity
On inhibition of peptidoglycan synthesis, uncontrolled hydrolase activity results in rupture of the sacculus and cell lysis, making it imperative to control hydrolase activity. It is likely, as proposed by Höltje3, that hydrolase activity is controlled by incorporation into multi-enzyme complexes that span the periplasm, extending from inner membrane-anchored synthases to the hydrolases, many of which are localized to the outer membrane. As part of a complex, hydrolases would be localized only to sites of peptidoglycan synthesis, thereby preventing them from hydrolysing peptidoglycan elsewhere. Intact complexes have not yet been isolated, possibly because they are dynamic, held together by weak interactions and/or tend to dissociate on breakage of the sacculus during cell lysis. However, protein interaction data7,99,100 and the fact that each major bifunctional synthase has an outer-membrane protein regulator101,102 support the existence of such complexes.
Hydrolase regulation is likely to be widespread. Above, we describe one situation in which hydrolases (or non-catalytic hydrolase variants) bind to and activate other hydrolases93, and other examples of regulation have been reported recently100. The Pseudomonas aeruginosa protein inhibitor of vertebrate lysozyme (Ivy) illustrates a different regulatory paradigm. Originally thought to be a defence mechanism against only host lysozyme, Ivy also works against bacterial hydrolases, inhibiting membrane-bound lytic murein transglycosylase B (MltB)103. Such studies imply that bacteria have specific inhibitors to control their hydrolases. Indeed, some type VI secretion systems (T6SSs) involved in bacterial interspecies interactions use peptidoglycan hydrolases as effectors104. Interestingly, periplasmic immunity proteins protect the producer cell by binding to and inhibiting hydrolases delivered by neighbouring cells104.
Control of sacculus growth from the outside
Recently, outer membrane-anchored lipoproteins that reach through the sacculus were found to control peptidoglycan synthases, complementing synthase dependence on cytoskeletal elements located in the sacculus. PBP1A and PBP1B each require a dedicated lipoprotein regulator for function in vivo101,102. Indeed, deleting the outer-membrane PBP activator lipoproteins, LpoA and LpoB, recapitulates the in vivo phenotypes of deleting PBP1A and PBP1B, respectively101,102. Each Lpo protein stimulates the TPase activity of its cognate PBP in vitro by an unknown mechanism102. LpoB has an additional small effect on the GTase rate of PBP1B in vitro, possibly at the initiation step101. Further studies are necessary to elucidate the basis for Lpo essentiality in vivo.
The Lpo proteins are limited to gammaproteobacteria (LpoA) or even to enterobacteria (LpoB), although they control universally conserved enzymes. LpoA and LpoB arose independently and have no sequence homology. Interestingly, LpoA and LpoB bind to portions of their cognate PBPs that are present in bacterial lineages roughly coincident with those harbouring the Lpo proteins themselves, suggesting that the Lpo proteins and their interacting domains co-evolved102. The additional level of regulation provided by Lpo proteins may enable niche-specific adaptation.
Other bacterial groups may have proteins with regulatory roles similar to those identified in Lpo proteins. In favour of this idea, class A PBPs in many bacterial clades exhibit diversified clade-specific non-catalytic regions near their termini or sandwiched between the GTase and TPase domains (FIG. 4). Importantly, in some cases, these regions or domains are functionally linked by genomic context or co-occurrence to outer-membrane lipoproteins or a peptidoglycan-related enzyme. For example, P. aeruginosa PBP1B has two non-catalytic domains that are highly conserved in members of the order Pseudomonadales (FIG. 4) and are functionally linked to an adjacent Pseudomonas spp.-specific outer-membrane lipoprotein containing an SPR domain, PA4699 (REF. 105). Similarly, Agrobacterium tumefaciens PBPC (encoded by the locus Atu0103) has unique domains that are not found outside the order Rhizobiales (FIG. 4) but are functionally linked to three M23–LytM family endopeptidases: Atu1832, Atu4178 and the outer-membrane lipoprotein Atu1700 (REF 105). Synthase diversity is also evident in Gram-positive organisms. B. subtilis PBP1 has a C-terminal region of ~220 amino acids that is present only in this organism (FIG. 4); closely related Bacillus spp. retain at least part of this domain. The recent crystal structure of Acinetobacter baumannii PBP1A shows an additional oligonucleotide–oligosaccharide-binding (OB)-fold domain inserted into the TPase domain106.
Figure 4. Species-specific non-catalytic regions in penicillin-binding proteins.
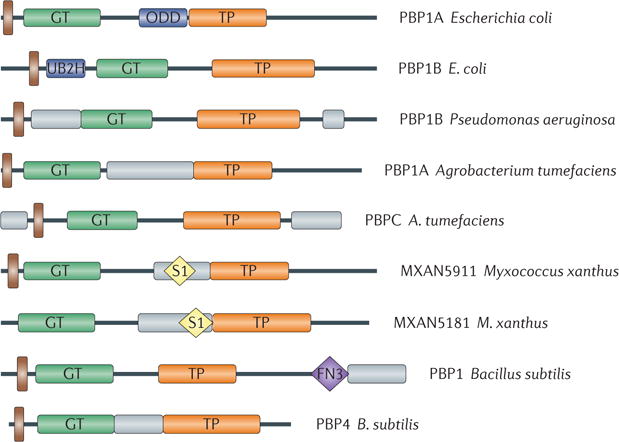
Different class A penicillin-binding proteins (PBPs) in comparison with Escherichia coli PBP1A and PBP1B. Predicted or known transmembrane domains are shown in brown, newly evolved domains in E. coli PBP1A and PBP1B in dark blue and other species-specific regions with no function prediction in grey. Glycosyltransferase (GT) and transpeptidase (TP) domains are labelled, along with the fibronectin type 3 (FN3) domain and the ribosomal protein S1-like RNA-binding (S1) domains. The species-specific regions with no function prediction in the two Myxococcus xanthus proteins contain an S1 domain and are only conserved in Stigmatella and Myxococcus spp., whereas the analogous regions in the two Bacillus subtilis proteins consist of one that is unique in B. subtilis (the carboxy-terminal region in PBP1) and one that is more conserved among the bacilli (the domain in PBP4); the FN3 domain found in PBP1 is also conserved only in bacilli. ODD, outer-membrane PBP1A docking domain; UB2H, UvrB domain 2 homologue.
Cell division in Gram-negative organisms involves simultaneous invagination of the outer membrane and constriction of the peptidoglycan and inner-membrane layers, an issue not yet discussed. The Tol–Pal complex participates in outer-membrane constriction107 and is essential in the alphaproteobacterium C. crescentus108. However, the Tol–Pal complex is not essential in E. coli and other gammaproteobacteria, raising the possibility that other proteins facilitate outer-membrane constriction in gammaproteobacteria. Removing PBP1B or LpoB from E. coli cells lacking the Tol–Pal complex results in synthetic sickness or lethality and severe division defects, suggesting that the PBP1B–LpoB complex participates in the process102. This unique role for PBP1B–LpoB in outer-membrane invagination is consistent with its preferential localization at the divisome102. How the two systems coordinate their actions remains to be determined.
Why regulate peptidoglycan synthesis from outside the sacculus?
Using outer-membrane regulators of peptidoglycan synthases ensures that these multiprotein synthetic complexes stretch from the inner membrane to the outer membrane. If the outer-membrane proteins also anchored the hydrolases, this would provide the kind of complex envisioned by Höltje to coordinate peptidoglycan synthesis and turnover, providing a rationale for the existence of outer-membrane regulators. Alternatively, or in addition, requiring the Lpo proteins to traverse the peptidoglycan sacculus might enable the physical properties of the sacculus to directly regulate peptidoglycan synthesis. Outer membrane-anchored LpoB must reach through pores in the peptidoglycan layer to interact with its docking domain in PBP1B102 (FIG. 5a). Several interesting regulatory scenarios can be imagined. First, the maximal molecular length of the Lpo activators might provide a molecular ruler to limit peptidoglycan thickness to a single layer; a thick, multilayered peptidoglycan might prevent access of the outer-membrane regulator to its synthases. Second, the method of activating peptidoglycan synthesis through the pores could be a robust, autoregulatory mechanism to adjust the peptidoglycan growth rate to the overall cellular growth rate based on peptidoglycan stretching (FIG. 5b). Unlike the much stiffer walls of algae and higher plants, which are only slightly strained under normal turgor109, the E. coli peptidoglycan net is elastic and can be stretched by turgor in vivo110–112 or by mechanical force in vitro113,114, resulting in larger pore size. As turgor is high in fast-growing cells or those in low-osmolality medium110, such cells might have a stretched peptidoglycan with bigger pores, thereby facilitating Lpo access to its synthases and activation of peptidoglycan synthesis. This homeostatic strategy would mechanically couple the peptidoglycan synthesis rate to the growth rate via peptidoglycan pore size (FIG. 5c). Moreover, the periplasmic volume and dimensions change during steady-state growth in different osmolality conditions and/or osmotic shifts110,115, and this could influence Lpo-mediated PBP activation by changing the distance between Lpo proteins and PBPs. Finally, it is important to note other mechanisms that might couple peptidoglycan synthesis and stretching. At high turgor, the net is likely to be more ‘ordered’, possibly resulting in a less flexible or less variable orientation of glycan chains and peptides. Optimally positioning the synthases and their sacculus substrates could increase peptidoglycan synthesis116. Thus, internal turgor can have a mechanical impact on peptidoglycan growth and thereby determine cell growth and morphology117.
Figure 5. Regulation of peptidoglycan synthesis by outer-membrane proteins.
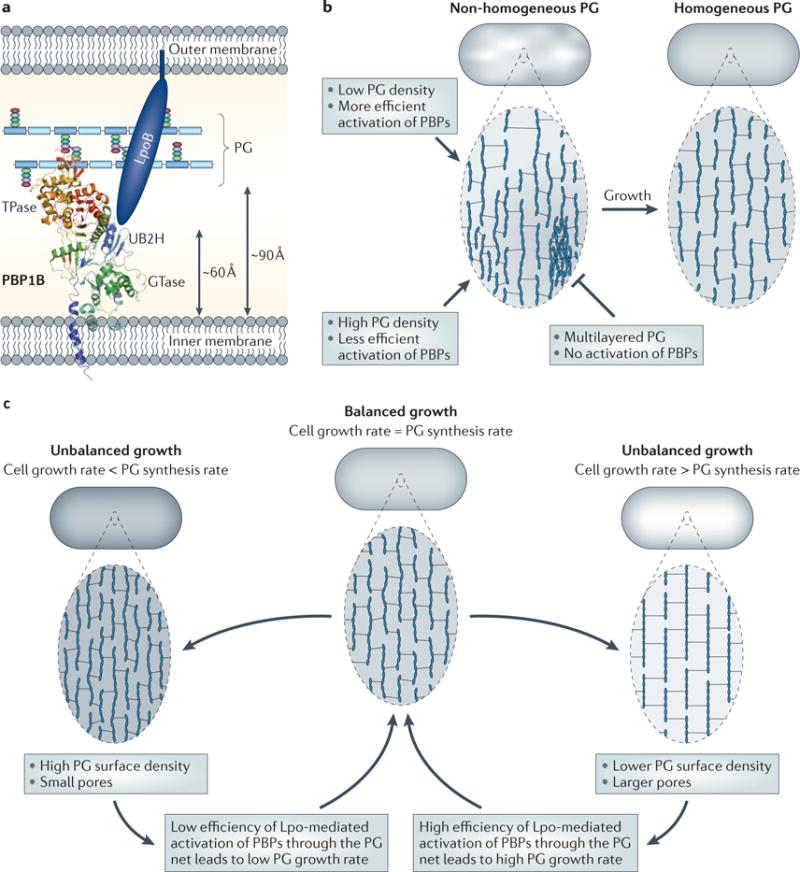
a | Side view of the Escherichia coli cell envelope with the crystal structure of penicillin-binding protein 1B (PBP1B; Protein Data Bank accession 3FWM)14 and the distances between the inner membrane, peptidoglycan (PG) and outer membrane drawn to scale. The glycosyltransferase (GTase) and transpeptidase (TPase) domains are shown. The structure of the activator protein for PBP1B, LpoB, is unknown. LpoB is anchored in the outer membrane and interacts with the PBP1B UB2H (UvrB domain 2 homologue) domain, which is situated between the inner membrane and the PG layer, not more than ~60 Å away from the inner membrane14. The distance from the inner membrane to the PG is ~90 Å138. b | A hypothetical self-repair mechanism to maintain a homogeneous peptidoglycan layer. The cell on the left has a non-homogeneous peptidoglycan layer consisting of large and small pores. Pore size-responsive activation of peptidoglycan synthase activity results in a more homogeneous peptidoglycan layer (on the right). c | A hypothetical homeostatic mechanism to balance the peptidoglycan growth rate with the overall cellular growth rate. When the peptidoglycan growth rate falls behind or exceeds that of overall cell growth, the peptidoglycan net stretches or relaxes, respectively. The resulting change in pore size alters the efficiency with which Lpo proteins can activate peptidoglycan synthases and therefore re-aligns the peptidoglycan growth rate with the overall cellular growth rate.
Peptidoglycan remodelling
The peptidoglycan composition of stationary-phase E. coli cells differs from that of exponentially growing cells. The average glycan chain length decreases by ~30%, whereas crosslinkage (especially 3–3 crosslinks) and covalent attachment of Lpp increase118. These changes may reflect peptidoglycan maturation, as the rate of insertion of new material into the sacculus slows down in stationary phase. Remodelling of peptidoglycan occurs more frequently and to a larger extent in Gram-positive bacteria. The glycan chains are O-acetylated and/or N-deacetylated for lysozyme resistance; the peptides are amidated and have cell wall proteins and anionic surface polymers, such as teichoic acids or capsular polysaccharides, covalently attached2,119.
E. coli can covalently link certain D-amino acids (for example D-Cys, D-Met or D-Trp) to its peptidoglycan. Indeed, in vivo labelling of peptidoglycan with exogenous D-Cys is a valuable tool for the study of peptidoglycan growth and segregation in various species56,57. Although D-amino acids are produced by some eukaryotes and bacteria for non-ribosomal peptide synthesis, they were not thought to have extensive roles in bacteria and their attachment to peptidoglycan was considered to be a side reaction of peptidoglycan enzymes. This view has changed with the discovery of distinct amino acid racemases for the synthesis of D-Met, D-Leu, D-Tyr and D-Phe in Vibrio cholerae and B. subtilis and the release of these D-amino acids into the growth medium, where they accumulate at millimolar concentrations120. The D-amino acids are incorporated into peptidoglycan and, in the case of V. cholerae, cause a rod-to-sphere transition in a strain lacking PBP1A, even in vegetatively growing cells. Subsequent experiments showed that D-amino acids are produced by many bacteria, accumulate mainly during stationary phase and are incorporated into peptidoglycan in several ways120,121. LD-TPases can add D-amino acids to position 4 of peptidoglycan tetrapeptides in E. coli and V. cholerae, in addition to forming 3–3 crosslinks or attaching Lpp (FIG. 1). Moreover, in V. cholerae, two enzymes involved in synthesis of the activated nucleotide precursors, D-Ala–D-Ala ligase (Ddl) and UDP-N-acetylmuramyl tripeptide–D-alanyl-D-Ala ligase (MurF), can use D-amino acids to give rise to modified peptidoglycan pentapeptides121. It is interesting that the E. coli TPase PBP1A can incorporate D-amino acids into pentapeptides of peptidoglycan fragments in vitro122, although D-amino acid-modified pentapeptides have not been detected yet in vivo.
What is the physiological role of modification with D-amino acids? These amino acids could have regulatory roles in cell wall remodelling during stationary phase and/or act as extracellular regulatory signals in a similar way to muropeptides, which have been shown to trigger developmental decisions in bacteria123. Astonishingly, a mixture of four D-amino acids at nanomolar concentrations inhibits biofilm formation in B. subtilis and causes disassembly of already formed biofilms. D-amino acids are also able to disperse biofilms of S. aureus or P. aeruginosa, but the molecular mechanisms underlying these processes have yet to be elucidated124.
Future directions
Despite the substantial progress made in recent decades, we are far from fully understanding peptidoglycan synthesis and its control, because of the complexity of this process, the functional redundancy of players involved and the multiple interconnections with other central processes. New technologies are gradually revealing the contours of this process (BOX 1). Fuelled by improved imaging techniques, the increasing diversity of bacteria investigated, more sophisticated genetics, and the entrance of biophysicists, modellers and engineers to the field, peptidoglycan research is currently experiencing a renaissance period. Perhaps for the first time, we have the ability to address fundamental questions about the entire process.
Box 1. New technologies will help to address long-standing questions.
High-throughput genetic screens
Genetic screens have become extremely powerful for dissecting the physiological roles of partially redundant gene functions. Unbiased, high-throughput reverse-genetics screens probe and quantify high numbers of genetic interactions in Escherichia coli125,126, and chemical genetic screens quantify the fitness of a large number of mutants under numerous environmental and chemical stresses127. These two methods, along with targeted genetics and proteomics, led to the identification of the Lpo proteins101,102,127 and provided additional leads about other peptidoglycan-related functions (C.G., A.T. and W.V., unpublished observations).
New imaging technology
New, high-resolution imaging techniques are being increasingly used to understand sacculus growth and organization. Visualization of cell machineries and the ultrastructure by electron cryotomography (ECT) has revealed the orientation of glycan chains and facilitated the discovery of new cytoskeletal elements22,71,86. Atomic force microscopy (AFM), which provides nanoscale images of native cell surfaces of living bacteria128,129, revealed the unexpected complexity of the peptidoglycan architecture in Bacillus subtilis and Staphylococcus aureus130,131 and is used to study assembly dynamics of cytoskeletal elements in vitro132. Total internal reflection fluorescence microscopy (TIRF microscopy) and photoactivated localization microscopy (PALM) methodologies45 are also increasingly employed. Indeed, TIRF measurements underlie the recent proposal that peptidoglycan synthesis drives MreB filament movement47,48.
Modelling and biophysical techniques
Computational models are being used to explain data and guide future experimentation49,80,81,133, and a suite of high-resolution biophysical analyses is also proving transformative. Optical traps are being used to assess the forces that are important in peptidoglycan maintenance77. The structural dynamics of entire sacculi, as well as of sacculus–protein interactions, are being identified using solid-state NMR spectroscopy134,135, and Förster resonance energy transfer (FRET) is being applied to identify protein interactions in vivo136.
In vitro reconstruction
In vitro reconstitution studies are increasingly providing insights into molecular mechanisms, demonstrating that FtsZ has the intrinsic ability to mediate constriction when present in lipsomes137 and that the transpeptidase and glycosyltransferase activities of the bifunctional synthases are interdependent12,13.
The recent identification of key molecules in peptidoglycan synthesis illustrates that this process is not completely described. The new technologies now available will be able to identify the missing players, determine their physiological roles and reconstitute the relevant complexes in vitro to study both mechanism and structure.
How peptidoglycan synthesis is tied to overall cellular growth and coordinated with other processes is under intense investigation. Equally important is to understand how peptidoglycan synthesis is coordinated with the synthesis and transport of other envelope macromolecules. These studies must address how the growing cell changes size while maintaining its integrity and shape. Some of the species-specific non-catalytic domains found in peptidoglycan synthesis enzymes (FIG. 4) may be used as docking domains to other processes or pathways that facilitate coordination between peptidoglycan growth and remodelling and envelope biogenesis.
The recent spate of articles seeking to understand the driving force behind peptidoglycan growth clearly indicates the diversity of cutting-edge technologies addressing this issue. These technologies will undoubtedly be employed to explore related general issues, such as the role of mechanical force and cell wall stresses in cell wall growth, and to identify the minimal physical parameters required for shape determination in bacteria.
The overarching principles coupling cell growth to cell shape and cell shape to peptidoglycan synthesis are likely to be broadly conserved. However, the diversity of cell shapes, developmental programmes and ecological niches that exists suggests that there are species-specific ways of tailoring these processes and/or that completely new mechanisms have arisen to facilitate coordination. Studying morphogenesis in differently shaped bacteria with more complicated cell cycle programmes will not only broaden our understanding of the mechanisms underlying cell growth and morphology, but also pinpoint strategies used to make such mechanisms robust.
Acknowledgments
This work was supported by grants from the UK Biotechnology and Biological Sciences Research Council (BB/G015902/1 and BBI020012/1 to W.V.), the European Commission (DIVINOCELL HEALTH-F3-2009-223,431 to W.V.), the Royal Society (to W.V.) and the US National Institutes of Health (R01 GM085697, ARRA GM085697-01S1 and R01 GM036278 to C.A.G., and K99GM092984 to A.T.).
Glossary
- Sacculus
A bag-like macromolecule that is made of peptidoglycan chains crosslinked by short peptides. The sacculus completely encases the cytoplasmic membrane in most bacteria, and isolated sacculi retain the shape of the bacterial cell
- Bacterial cytoskeleton
A filamentous and often dynamic cytoplasmic structure that includes bacterial structural homologues of actin, tubulin or intermediate filaments and is essential for bacterial growth, motility, cell division, morphology and DNA segregation
- Actin
A eukaryotic cytoskeletal protein with ATPase activity. MreB and ParM, two bacterial proteins involved in cell elongation and plasmid partitioning, respectively, are distant actin homologues
- Tubulin
A cytoskeletal protein that forms microtubules in eukaryotes;the bacterial tubulin-like protein FtsZ, is a GTPase and forms dynamic filaments to drive cell division.
- Penicillin-binding proteins (PBPs)
A protein family involved in the synthesis (the class A and class B PBPs) or hydrolysis (the class C PBPs) of D-amino acid–D-amino acid peptide bonds. They contain an active-site Ser residue that participates in the transfer of an acyl compound to an amino group or water. PBPs are the targets of β-lactam antibiotics (such as penicillin). Pathogen resistance to β-lactams can be caused by low-affinity PBPs
- β-lactam antibiotics
An important class of antibiotics, members of which contain a β-lactam ring and inhibit peptidoglycan synthesis by covalent binding to the active-site Ser of penicillin-binding proteins
- Autolysins
Proteins that are located in the periplasm of Gram-negative bacteria or in the cell wall of Gram-positive bacteria and can lyse the cell using their peptidoglycan-hydrolysing activity. Autolysins can have muramidase, glucosaminidase, amidase and/or endopeptidase activity
- Electron cryotomography (ECT)
An electron microscopy technique that provides high-resolution pictures of an object from different angles, permitting its three-dimensional reconstitution; plunge-freezing of the samples prevents staining and fixation artefacts. In the case of the bacterial sacculus, ECT has yielded a nanometre-scale three-dimensional representation of the fine structure.
- Intermediate filaments
Filaments formed by coiled-coil-rich cytoskeletal proteins, such as keratin. Crescentin is a bacterial version of an intermediate filament and is required for the bent cell shape of Caulobacter crescentus.
- Blebbing
The release of vesicles from the outer membrane of Gram-negative bacteria. Blebbing occurs during normal growth and is enhanced in certain mutants that are impaired in cell division
- Lysozyme
An antibacterial enzyme that is produced in animals, plants, fungi and even bacteria and is capable of lysing sensitive bacteria by hydrolysing the N-acetylmuramic acid–N-acetylglucosamine bonds in peptidoglycan chains
- Type VI secretion systems (T6SSs)
A recently discovered secretion apparatus that is widely distributed in Gram-negative bacteria. Some of its components are similar to phage injection systems.The T6SS punctures both eukaryotic and bacterial cells, often injecting toxic effector proteins into them
- Turgor
The osmotic pressure of a compartment (here, the bacterial cytoplasm) that is due to the lower activity of water
- D-amino acids
Rare chiral forms (mirror structures) of the abundant L-amino acids that build proteins. D-amino acids are present in peptidoglycan and in some non-ribosomally synthesized antibiotics
- Atomic force microscopy (AFM)
A microscopy technique that uses a cantilever tip to scan the surface of a probe, either in direct contact or in oscillation mode, to produce topography images with nanometre-scale resolution
- Total internal reflection fluorescence microscopy (TIRF microscopy)
A fluorescence microscopy technique that uses an evanescent wave to selectively excite a fluorophore in a small area of a specimen adjacent to a glass–water interface to reduce background fluorescence. This technique provides a superior axial resolution
- Photoactivated localization microscopy (PALM)
A super-resolution fluorescence microscopy technique based on the controlled activation and sampling of subsets of photoconvertible fluorescent molecules in the sample. This technique can achieve 10–20 nm resolution
- Solid-state NMR spectroscopy
NMR spectroscopy of insoluble polymers. The technique requires rapid spinning of the sample at a certain ‘magic’ angle. It provides information on the structural flexibility of a polymer and the interactions of chemical entities within it (for example, amino acids or sugars in peptidoglycan sacculi)
- Förster resonance energy transfer (FRET)
A technique that detects and characterizes the interaction between two molecules coupled to two fluorophores, by measuring the excitation of one fluorophore by the light emitted from the other. A positive FRET signal indicates a distance of less than 10 nm between the fluorophores
Footnotes
Competing interests statement
The authors declare no competing financial interests.
References
- 1.Young KD. The selective value of bacterial shape. Microbiol Mol Biol Rev. 2006;70:660–703. doi: 10.1128/MMBR.00001-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Vollmer W, Blanot D, de Pedro MA. Peptidoglycan structure and architecture. FEMS Microbiol Rev. 2008;32:149–167. doi: 10.1111/j.1574-6976.2007.00094.x. [DOI] [PubMed] [Google Scholar]
- 3.Höltje J-V. Growth of the stress-bearing and shape-maintaining murein sacculus of Escherichia coli Microbiol. Mol Biol Rev. 1998;62:181–203. doi: 10.1128/mmbr.62.1.181-203.1998. A landmark review on peptidoglycan synthesis in E. coli, with details of the ‘3 for 1’ growth model. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Barreteau H, et al. Cytoplasmic steps of peptidoglycan biosynthesis. FEMS Microbiol Rev. 2008;32:168–207. doi: 10.1111/j.1574-6976.2008.00104.x. [DOI] [PubMed] [Google Scholar]
- 5.Bouhss A, Trunkfield AE, Bugg TD, Mengin-Lecreulx D. The biosynthesis of peptidoglycan lipid-linked intermediates. FEMS Microbiol Rev. 2008;32:208–233. doi: 10.1111/j.1574-6976.2007.00089.x. [DOI] [PubMed] [Google Scholar]
- 6.Mohammadi T, et al. Identification of FtsW as a transporter of lipid-linked cell wall precursors across the membrane. EMBO J. 2011;30:1425–1432. doi: 10.1038/emboj.2011.61. An article that identifies a member of the conserved SEDS (shape, elongation, division and sporulation) family of integral membrane proteins as the elusive lipid II flippase. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Vollmer W, Bertsche U. Murein (peptidoglycan) structure, architecture and biosynthesis in Escherichia coli. Biochim Biophys Acta. 2008;1778:1714–1734. doi: 10.1016/j.bbamem.2007.06.007. [DOI] [PubMed] [Google Scholar]
- 8.Suginaka H, Blumberg PM, Strominger JL. Multiple penicillin-binding components in Bacillus subtilis, Bacillus cereus, Staphylococcus aureus, and Escherichia coli. J Biol Chem. 1972;247:5279–5288. [PubMed] [Google Scholar]
- 9.Yousif SY, Broome-Smith JK, Spratt BG. Lysis of Escherichia coli by β-lactam antibiotics: deletion analysis of the role of penicillin-binding proteins 1A and 1B. J Gen Microbiol. 1985;131:2839–2845. doi: 10.1099/00221287-131-10-2839. [DOI] [PubMed] [Google Scholar]
- 10.Budd A, Blandin S, Levashina EA, Gibson TJ. Bacterial α2-macroglobulins: colonization factors acquired by horizontal gene transfer from the metazoan genome? Genome Biol. 2004;5:R38. doi: 10.1186/gb-2004-5-6-r38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Bertsche U, et al. Interaction between two murein (peptidoglycan) synthases, BP3 and BP1B in Escherichia coli. Mol Microbiol. 2006;61:675–690. doi: 10.1111/j.1365-2958.2006.05280.x. The first study to provide evidence for a direct interaction between peptidoglycan synthases. [DOI] [PubMed] [Google Scholar]
- 12.Bertsche U, Breukink E, Kast T, Vollmer W. In vitro murein peptidoglycan synthesis by dimers of the bifunctional transglycosylase-transpeptidase PBP1B from Escherichia coli. J Biol Chem. 2005;280:38096–38101. doi: 10.1074/jbc.M508646200. [DOI] [PubMed] [Google Scholar]
- 13.Born P, Breukink E, Vollmer W. In vitro synthesis of cross-linked murein and its attachment to sacculi by BP1A from Escherichia coli. J Biol Chem. 2006;281:26985–26993. doi: 10.1074/jbc.M604083200. Together with reference12, this work establishes a novel in vitro peptidoglycan assay for simultaneous detection of GTase and TPase products, demonstrating that these activities are coupled, and reconstitutes for the first time the naturally occurring reaction of TPase-mediated attachment of newly made peptidoglycan to the sacculus. [DOI] [PubMed] [Google Scholar]
- 14.Sung MT, et al. Crystal structure of the membrane-bound bifunctional transglycosylase BP1b from Escherichia coli. Proc Natl Acad Sci USA. 2009;106:8824–8829. doi: 10.1073/pnas.0904030106. This article describes the crystal structure of BP1B which includes the transmembrane region, and suggests a model for the coupling of GTase and TPase reactions. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Macheboeuf P, Contreras-Martel C, Job V, Dideberg O, Dessen A. Penicillin binding proteins: key players in bacterial cell cycle and drug resistance processes. FEMS Microbiol Rev. 2006;30:673–691. doi: 10.1111/j.1574-6976.2006.00024.x. [DOI] [PubMed] [Google Scholar]
- 16.Vollmer W, Joris B, Charlier P, Foster S. Bacterial peptidoglycan (murein) hydrolases. FEMS Microbiol Rev. 2008;32:259–286. doi: 10.1111/j.1574-6976.2007.00099.x. [DOI] [PubMed] [Google Scholar]
- 17.Heidrich C, et al. Involvement of N-acetylmuramyl-L-alanine amidases in cell separation and antibiotic-induced autolysis of Escherichia coli. Mol Microbiol. 2001;41:167–178. doi: 10.1046/j.1365-2958.2001.02499.x. [DOI] [PubMed] [Google Scholar]
- 18.Priyadarshini R, Popham DL, Young KD. Daughter cell separation by penicillin-binding proteins and peptidoglycan amidases in Escherichia coli. J Bacteriol. 2006;188:5345–5355. doi: 10.1128/JB.00476-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Heidrich C, Ursinus A, Berger J, Schwarz H, Höltje JV. Effects of multiple deletions of murein hydrolases on viability, septum cleavage, and sensitivity to large toxic molecules in Escherichia coli. J Bacteriol. 2002;184:6093–6099. doi: 10.1128/JB.184.22.6093-6099.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kraft AR, Prabhu J, Ursinus A, Höltje JV. Interference with murein turnover has no effect on growth but reduces β -lactamase induction in Escherichia coli. J Bacteriol. 1999;181:7192–7198. doi: 10.1128/jb.181.23.7192-7198.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Park JT, Uehara T. How bacteria consume their own exoskeletons (turnover and recycling of cell wall peptidoglycan) Microbiol Mol Biol Rev. 2008;72:211–227. doi: 10.1128/MMBR.00027-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Gan L, Chen S, Jensen GJ. Molecular organization of Gram-negative peptidoglycan. Proc Natl Acad Sci USA. 2008;105:18953–18957. doi: 10.1073/pnas.0808035105. In this study, ECT solves a long-standing dispute about the orientation of the glycan chains in the single-layered peptidoglycan in Gram-negative bacteria. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Burman LG, Park JT. Molecular model for elongation of the murein sacculus of Escherichia coli. Proc Natl Acad Sci USA. 1984;81:1844–1848. doi: 10.1073/pnas.81.6.1844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Glauner B, Höltje J-V. Growth pattern of the murein sacculus of Escherichia coli. J Biol Chem. 1990;265:18988–18996. A paper that illustrates the alterations in peptidoglycan structure that occur during maturation. [PubMed] [Google Scholar]
- 25.Goodell EW, Markiewicz Z, Schwarz U. Absence of oligomeric murein intermediates in Escherichia coli. J Bacteriol. 1983;156:130–135. doi: 10.1128/jb.156.1.130-135.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Uehara T, Park JT. Growth of Escherichia coli: significance of peptidoglycan degradation during elongation and septation. J Bacteriol. 2008;190:3914–3922. doi: 10.1128/JB.00207-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.de Jonge BL, et al. Peptidoglycan synthesis during the cell cycle of Escherichia coli: composition and mode of insertion. J Bacteriol. 1989;171:5783–5794. doi: 10.1128/jb.171.11.5783-5794.1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cabeen MT, Jacobs-Wagner C. The bacterial cytoskeleton. Annu Rev Genet. 2010;44:365–392. doi: 10.1146/annurev-genet-102108-134845. [DOI] [PubMed] [Google Scholar]
- 29.Daniel RA, Errington J. Control of cell morphogenesis in bacteria: two distinct ways to make a rod-shaped cell. Cell. 2003;113:767–776. doi: 10.1016/s0092-8674(03)00421-5. This work uses labelling of peptidoglycan synthesis sites to determine the topography of peptidoglycan growth in rod-shaped bacteria with or without MreB. [DOI] [PubMed] [Google Scholar]
- 30.Jones LJ, Carballido-Lopez R, Errington J. Control of cell shape in bacteria: helical, actin-like filaments in Bacillus subtilis. Cell. 2001;104:913–922. doi: 10.1016/s0092-8674(01)00287-2. A seminal paper demonstrating that MreB filaments control cell elongation. [DOI] [PubMed] [Google Scholar]
- 31.Vats P, Shih YL, Rothfield L. Assembly of the MreB-associated cytoskeletal ring of Escherichia coli. Mol Microbiol. 2009;72:170–182. doi: 10.1111/j.1365-2958.2009.06632.x. [DOI] [PubMed] [Google Scholar]
- 32.Alyahya SA, et al. RodZ, a component of the bacterial core morphogenic apparatus. Proc Natl Acad Sci USA. 2009;106:1239–1244. doi: 10.1073/pnas.0810794106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bendezu FO, Hale CA, Bernhardt TG, de Boer PA. RodZ (YfgA) is required for proper assembly of the MreB actin cytoskeleton and cell shape in E. coli EMBO J. 2009;28:193–204. doi: 10.1038/emboj.2008.264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Kruse T, Bork-Jensen J, Gerdes K. The morphogenetic MreBCD proteins of Escherichia coli form an essential membrane-bound complex. Mol Microbiol. 2005;55:78–89. doi: 10.1111/j.1365-2958.2004.04367.x. [DOI] [PubMed] [Google Scholar]
- 35.Shiomi D, Sakai M, Niki H. Determination of bacterial rod shape by a novel cytoskeletal membrane protein. EmBo J. 2008;27:3081–3091. doi: 10.1038/emboj.2008.234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.van den Ent F, et al. Dimeric structure of the cell shape protein MreC and its functional implications. Mol Microbiol. 2006;62:1631–1642. doi: 10.1111/j.1365-2958.2006.05485.x. [DOI] [PubMed] [Google Scholar]
- 37.Mohammadi T, et al. The essential peptidoglycan glycosyltransferase MurG forms a complex with proteins involved in lateral envelope growth as well as with proteins involved in cell division in Escherichia coli. Mol Microbiol. 2007;65:1106–1121. doi: 10.1111/j.1365-2958.2007.05851.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.van den Ent F, Johnson CM, Persons L, de Boer P, Löwe J. Bacterial actin MreB assembles in complex with cell shape protein RodZ. EMBO J. 2010;29:1081–1090. doi: 10.1038/emboj.2010.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Salje J, van den Ent F, de Boer P, Löwe J. Direct membrane binding by bacterial actin MreB. Mol Cell. 2011;43:478–487. doi: 10.1016/j.molcel.2011.07.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gitai Z, Dye NA, Reisenauer A, Wachi M, Shapiro L. MreB actin-mediated segregation of a specific region of a bacterial chromosome. Cell. 2005;120:329–341. doi: 10.1016/j.cell.2005.01.007. [DOI] [PubMed] [Google Scholar]
- 41.Karczmarek A, et al. DNA and origin region segregation are not affected by the transition from rod to sphere after inhibition of Escherichia coli MreB by A22. Mol Microbiol. 2007;65:51–63. doi: 10.1111/j.1365-2958.2007.05777.x. [DOI] [PubMed] [Google Scholar]
- 42.Takacs CN, et al. MreB drives de novo rod morphogenesis in Caulobacter crescentus via remodeling of the cell wall. J Bacteriol. 2010;192:1671–1684. doi: 10.1128/JB.01311-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gitai Z, Dye N, Shapiro L. An actin-like gene can determine cell polarity in bacteria. Proc Natl Acad Sci USA. 2004;101:8643–8648. doi: 10.1073/pnas.0402638101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Popp D, et al. Filament structure, organization, and dynamics in MreB sheets. J Biol Chem. 2010;285:15858–15865. doi: 10.1074/jbc.M109.095901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Biteen JS, Moerner WE. Single-molecule and superresolution imaging in live bacteria cells. Cold Spring Harb Perspect Biol. 2010;2:a000448. doi: 10.1101/cshperspect.a000448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kim SY, Gitai Z, Kinkhabwala A, Shapiro L, Moerner WE. Single molecules of the bacterial actin MreB undergo directed treadmilling motion in Caulobacter crescentus. Proc Natl Acad Sci USA. 2006;103:10929–10934. doi: 10.1073/pnas.0604503103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dominguez-Escobar J, et al. Processive movement of MreB-associated cell wall biosynthetic complexes in bacteria. Science. 2011;333:225–228. doi: 10.1126/science.1203466. [DOI] [PubMed] [Google Scholar]
- 48.Garner EC, et al. Circumferential motions of the cell wall synthesis machinery drive cytoskeletal dynamics in B. subtilis Science. 2011;333:222–225. doi: 10.1126/science.1203285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.van Teeffelen S, et al. The bacterial actin MreB rotates, and rotation depends on cell-wall assembly. Proc Natl Acad Sci USA. 2011;108:15822–15827. doi: 10.1073/pnas.1108999108. References47–49describe high-resolution fluorescence microscopy showing peptidoglycan synthesis-dependent movement of MreB perpendicular to the long axis. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kawai Y, Daniel RA, Errington J. Regulation of cell wall morphogenesis in Bacillus subtilis by recruitment of PBP1 to the MreB helix. Mol Microbiol. 2009;71:1131–1144. doi: 10.1111/j.1365-2958.2009.06601.x. [DOI] [PubMed] [Google Scholar]
- 51.White CL, Kitich A, Gober JW. Positioning cell wall synthetic complexes by the bacterial morphogenetic proteins MreB and MreD. Mol Microbiol. 2010;76:616–633. doi: 10.1111/j.1365-2958.2010.07108.x. [DOI] [PubMed] [Google Scholar]
- 52.Land AD, Winkler ME. Requirement for pneumococcal MreC and MreD is relieved by inactivation of the gene encoding PBP1a. J Bacteriol. 2011;193:4166–4179. doi: 10.1128/JB.05245-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Adams DW, Errington J. Bacterial cell division: assembly, maintenance and disassembly of the Z ring. Nature Rev Microbiol. 2009;7:642–653. doi: 10.1038/nrmicro2198. [DOI] [PubMed] [Google Scholar]
- 54.Erickson HP, Anderson DE, Osawa M. FtsZ in bacterial cytokinesis: cytoskeleton and force generator all in one. Microbiol Mol Biol Rev. 2010;74:504–528. doi: 10.1128/MMBR.00021-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Aarsman ME, et al. Maturation of the Escherichia coli divisome occurs in two steps. Mol Microbiol. 2005;55:1631–1645. doi: 10.1111/j.1365-2958.2005.04502.x. [DOI] [PubMed] [Google Scholar]
- 56.Aaron M, et al. The tubulin homologue FtsZ contributes to cell elongation by guiding cell wall precursor synthesis in Caulobacter crescentus. Mol Microbiol. 2007;64:938–952. doi: 10.1111/j.1365-2958.2007.05720.x. This investigation demonstrates the FtsZ-dependent preseptal phase of cell elongation in C. crescentus. [DOI] [PubMed] [Google Scholar]
- 57.de Pedro MA, Quintela JC, Höltje JV, Schwarz H. Murein segregation in Escherichia coli. J Bacteriol. 1997;179:2823–2834. doi: 10.1128/jb.179.9.2823-2834.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Fraipont C, et al. The integral membrane FtsW protein and peptidoglycan synthase PBP3 form a subcomplex in Escherichia coli. Microbiology. 2011;157:251–259. doi: 10.1099/mic.0.040071-0. [DOI] [PubMed] [Google Scholar]
- 59.Buddelmeijer N, Beckwith J. A complex of the Escherichia coli cell division proteins FtsL, FtsB and FtsQ forms independently of its localization to the septal region. Mol Microbiol. 2004;52:1315–1327. doi: 10.1111/j.1365-2958.2004.04044.x. [DOI] [PubMed] [Google Scholar]
- 60.Masson S, et al. Central domain of DivIB caps the C-terminal regions of the FtsL/DivIC coiled-coil rod. J Biol Chem. 2009;284:27687–27700. doi: 10.1074/jbc.M109.019471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wissel MC, Weiss DS. Genetic analysis of the cell division protein FtsI (PBP3): amino acid substitutions that impair septal localization of FtsI and recruitment of FtsN. J Bacteriol. 2004;186:490–502. doi: 10.1128/JB.186.2.490-502.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rico AI, Garcia-Ovalle M, Palacios P, Casanova M, Vicente M. Role of Escherichia coli FtsN protein in the assembly and stability of the cell division ring. Mol Microbiol. 2010;76:760–771. doi: 10.1111/j.1365-2958.2010.07134.x. [DOI] [PubMed] [Google Scholar]
- 63.Bernard CS, Sadasivam M, Shiomi D, Margolin W. An altered FtsA can compensate for the loss of essential cell division protein FtsN in Escherichia coli. Mol Microbiol. 2007;64:1289–1305. doi: 10.1111/j.1365-2958.2007.05738.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Gerding MA, et al. Self-enhanced accumulation of FtsN at division sites and roles for other proteins with a SPOR domain (DamX, DedD, and RlpA) in Escherichia coli cell constriction. J Bacteriol. 2009;191:7383–7401. doi: 10.1128/JB.00811-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ursinus A, et al. Murein (peptidoglycan) binding property of the essential cell division protein FtsN from Escherichia coli. J Bacteriol. 2004;186:6728–6737. doi: 10.1128/JB.186.20.6728-6737.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Müller P, et al. The essential cell division protein FtsN interacts with the murein (peptidoglycan) synthase PBP1B in Escherichia coli. J Biol Chem. 2007;282:36394–36402. doi: 10.1074/jbc.M706390200. [DOI] [PubMed] [Google Scholar]
- 67.Goley ED, et al. Assembly of the Caulobacter cell division machine. Mol Microbiol. 2011;80:1680–1698. doi: 10.1111/j.1365-2958.2011.07677.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Briegel A, et al. Multiple large filament bundles observed in Caulobacter crescentus by electron cryotomography. Mol Microbiol. 2006;62:5–14. doi: 10.1111/j.1365-2958.2006.05355.x. [DOI] [PubMed] [Google Scholar]
- 69.Li Z, Jensen GJ. Electron cryotomography: a new view into microbial ultrastructure. Curr Opin Microbiol. 2009;12:333–340. doi: 10.1016/j.mib.2009.03.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Charbon G, Cabeen MT, Jacobs-Wagner C. Bacterial intermediate filaments: in vivo assembly, organization, and dynamics of crescentin. Genes Dev. 2009;23:1131–1144. doi: 10.1101/gad.1795509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ausmees N, Kuhn JR, Jacobs-Wagner C. The bacterial cytoskeleton: an intermediate filament-like function in cell shape. Cell. 2003;115:705–713. doi: 10.1016/s0092-8674(03)00935-8. This work identifies the first bacterial IF protein, CreS, which is required for the bent cell shape of C. crescentus. [DOI] [PubMed] [Google Scholar]
- 72.Cabeen MT, et al. Bacterial cell curvature through mechanical control of cell growth. EMBO J. 2009;28:1208–1219. doi: 10.1038/emboj.2009.61. A study which shows that mechanical force on the cell envelope, generated by CreS, causes C. crescentus and E. coli cells to grow with a bent shape. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Bagchi S, Tomenius H, Belova LM, Ausmees N. Intermediate filament-like proteins in bacteria and a cytoskeletal function in Streptomyces. Mol Microbiol. 2008;70:1037–1050. doi: 10.1111/j.1365-2958.2008.06473.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kühn J, et al. Bactofilins, a ubiquitous class of cytoskeletal proteins mediating polar localization of a cell wall synthase in Caulobacter crescentus. EMBO J. 2010;29:327–339. doi: 10.1038/emboj.2009.358. An article describing the identification of a new class of bacterial cytoskeleton proteins: the bactofilins. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Koch MK, McHugh CA, Hoiczyk E. BacM, an N-terminally processed bactofilin of Myxococcus xanthus, is crucial for proper cell shape. Mol Microbiol. 2011;80:1031–1051. doi: 10.1111/j.1365-2958.2011.07629.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Sycuro LK, et al. Peptidoglycan crosslinking relaxation promotes Helicobacter pylori’s helical shape and stomach colonization. Cell. 2010;141:822–833. doi: 10.1016/j.cell.2010.03.046. This investigation demonstrates that peptidoglycan endopeptidases and a bactofilin participate in generating the helical cell shape in H. pylori. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wang S, Arellano-Santoyo H, Combs PA, Shaevitz JW. Actin-like cytoskeleton filaments contribute to cell mechanics in bacteria. Proc Natl Acad Sci USA. 2010;107:9182–9185. doi: 10.1073/pnas.0911517107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Takeuchi S, DiLuzio WR, Weibel DB, Whitesides GM. Controlling the shape of filamentous cells of Escherichia coli. Nano Lett. 2005;5:1819–1823. doi: 10.1021/nl0507360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Hamant O, Traas J. The mechanics behind plant development. New Phytol. 2010;185:369–385. doi: 10.1111/j.1469-8137.2009.03100.x. [DOI] [PubMed] [Google Scholar]
- 80.Sliusarenko O, Cabeen MT, Wolgemuth CW, Jacobs-Wagner C, Emonet T. Processivity of peptidoglycan synthesis provides a built-in mechanism for the robustness of straight-rod cell morphology. Proc Natl Acad Sci USA. 2010;107:10086–10091. doi: 10.1073/pnas.1000737107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Furchtgott L, Wingreen NS, Huang KC. Mechanisms for maintaining cell shape in rod-shaped Gram-negative bacteria. Mol Microbiol. 2011;81:340–353. doi: 10.1111/j.1365-2958.2011.07616.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Osawa M, Anderson DE, Erickson HP. Curved FtsZ protofilaments generate bending forces on liposome membranes. EMBO J. 2009;28:3476–3484. doi: 10.1038/emboj.2009.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Weart RB, et al. A metabolic sensor governing cell size in bacteria. Cell. 2007;130:335–347. doi: 10.1016/j.cell.2007.05.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Foulquier E, Pompeo F, Bernadac A, Espinosa L, Galinier A. The YvcK protein is required for morphogenesis via localization of PBP1 under gluconeogenic growth conditions in Bacillus subtilis. Mol Microbiol. 2011;80:309–318. doi: 10.1111/j.1365-2958.2011.07587.x. [DOI] [PubMed] [Google Scholar]
- 85.Chaudhuri RR, et al. Comprehensive identification of essential Staphylococcus aureus genes using Transposon-Mediated Differential Hybridisation (TMDH) BMC Genomics. 2009;10:291. doi: 10.1186/1471-2164-10-291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Ingerson-Mahar M, Briegel A, Werner JN, Jensen GJ, Gitai Z. The metabolic enzyme CTP synthase forms cytoskeletal filaments. Nature Cell Biol. 2010;12:739–746. doi: 10.1038/ncb2087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.de Pedro MA, Young KD, Höltje JV, Schwarz H. Branching of Escherichia coli cells arises from multiple sites of inert peptidoglycan. J Bacteriol. 2003;185:1147–1152. doi: 10.1128/JB.185.4.1147-1152.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Nelson DE, Young KD. Penicillin binding protein 5 affects cell diameter, contour, and morphology of Escherichia coli. J Bacteriol. 2000;182:1714–1721. doi: 10.1128/jb.182.6.1714-1721.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Potluri L, et al. Septal and lateral wall localization of PBP5, the major D,D-carboxypeptidase of Escherichia coli, requires substrate recognition and membrane attachment. Mol Microbiol. 2010;77:300–323. doi: 10.1111/j.1365-2958.2010.07205.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Markiewicz Z, Glauner B, Schwarz U. Murein structure and lack of DD-and LD-carboxypeptidase activities in Caulobacter crescentus. J Bacteriol. 1983;156:649–655. doi: 10.1128/jb.156.2.649-655.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bernhardt TG, de Boer PA. The Escherichia coli amidase AmiC is a periplasmic septal ring component exported via the twin-arginine transport pathway. Mol Microbiol. 2003;48:1171–1182. doi: 10.1046/j.1365-2958.2003.03511.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Uehara T, Dinh T, Bernhardt TG. LytM-domain factors are required for daughter cell separation and rapid ampicillin-induced lysis in Escherichia coli. J Bacteriol. 2009;191:5094–5107. doi: 10.1128/JB.00505-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Uehara T, Parzych KR, Dinh T, Bernhardt TG. Daughter cell separation is controlled by cytokinetic ring-activated cell wall hydrolysis. EMBO J. 2010;29:1412–1422. doi: 10.1038/emboj.2010.36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Peters NT, Dinh T, Bernhardt TG. A fail-safe mechanism in the septal ring assembly pathway generated by the sequential recruitment of cell separation amidases and their activators. J Bacteriol. 2011;193:4973–4983. doi: 10.1128/JB.00316-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Goley ED, Comolli LR, Fero KE, Downing KH, Shapiro L. DipM links peptidoglycan remodelling to outer membrane organization in Caulobacter. Mol Microbiol. 2010;77:56–73. doi: 10.1111/j.1365-2958.2010.07222.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Moll A, Schlimpert S, Briegel A, Jensen GJ, Thanbichler M. DipM, a new factor required for peptidoglycan remodelling during cell division in Caulobacter crescentus. Mol Microbiol. 2010;77:90–107. doi: 10.1111/j.1365-2958.2010.07224.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Poggio S, Takacs CN, Vollmer W, Jacobs-Wagner C. A protein critical for cell constriction in the Gram-negative bacterium Caulobacter crescentus localizes at the division site through its peptidoglycan-binding LysM domains. Mol Microbiol. 2010;77:74–89. doi: 10.1111/j.1365-2958.2010.07223.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Bonis M, Ecobichon C, Guadagnini S, Prevost MC, Boneca IG. A M23B family metallopeptidase of Helicobacter pylori required for cell shape, pole formation and virulence. Mol Microbiol. 2010;78:809–819. doi: 10.1111/j.1365-2958.2010.07383.x. [DOI] [PubMed] [Google Scholar]
- 99.Legaree BA, Clarke AJ. Interaction of penicillinbinding protein 2 with soluble lytic transglycosylase B1 in Pseudomonas aeruginosa. J Bacteriol. 2008;190:6922–6926. doi: 10.1128/JB.00934-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Morlot C, Uehara T, Marquis KA, Bernhardt TG, Rudner DZ. A highly coordinated cell wall degradation machine governs spore morphogenesis in Bacillus subtilis. Genes Dev. 2010;24:411–422. doi: 10.1101/gad.1878110. This and reference93show for the first time that septum-splitting peptidoglycan hydrolases require activation by other proteins. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Paradis-Bleau C, et al. Lipoprotein cofactors located in the outer membrane activate bacterial cell wall polymerases. Cell. 2010;143:1110–1120. doi: 10.1016/j.cell.2010.11.037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Typas A, et al. Regulation of peptidoglycan synthesis by outer membrane proteins. Cell. 2010;143:1097–1109. doi: 10.1016/j.cell.2010.11.038. Together with reference101, this work demonstrates that peptidoglycan synthesis is controlled from outside the sacculus by newly identified outer-membrane lipoproteins. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Clarke CA, Scheurwater EM, Clarke AJ. The vertebrate lysozyme inhibitor Ivy functions to inhibit the activity of lytic transglycosylase. J Biol Chem. 2010;285:14843–14847. doi: 10.1074/jbc.C110.120931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Russell AB, et al. Type VI secretion delivers bacteriolytic effectors to target cells. Nature. 2011;475:343–347. doi: 10.1038/nature10244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Jensen LJ, et al. STRING 8—a global view on proteins and their functional interactions in 630 organisms. Nucleic Acids Res. 2009;37:D412–D416. doi: 10.1093/nar/gkn760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Han S, et al. Distinctive attributes of (β-lactam target proteins in Acinetobacter baumannii relevant to development of new antibiotics. J Am Chem Soc. 2011;133:20536–20545. doi: 10.1021/ja208835z. [DOI] [PubMed] [Google Scholar]
- 107.Gerding MA, Ogata Y, Pecora ND, Niki H, de Boer PA. The trans-envelope Tol–Pal complex is part of the cell division machinery and required for proper outer-membrane invagination during cell constriction in E. coli Mol Microbiol. 2007;63:1008–1025. doi: 10.1111/j.1365-2958.2006.05571.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Yeh YC, Comolli LR, Downing KH, Shapiro L, McAdams HH. The Caulobacter Tol-Pal complex is essential for outer membrane integrity and the positioning of a polar localization factor. J Bacteriol. 2010;192:4847–4858. doi: 10.1128/JB.00607-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Zimmermann U. Physics of turgor-and osmoregulation. Annu Rev Plant Physiol. 1978;29:121–148. [Google Scholar]
- 110.Cayley DS, Guttman HJ, Record MT., Jr Biophysical characterization of changes in amounts and activity of Escherichia coli cell and compartment water and turgor pressure in response to osmotic stress. Biophys J. 2000;78:1748–1764. doi: 10.1016/s0006-3495(00)76726-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Doyle RJ, Marquis RE. Elastic, flexible peptidoglycan and bacterial cell wall properties. Trends Microbiol. 1994;2:57–60. doi: 10.1016/0966-842x(94)90127-9. [DOI] [PubMed] [Google Scholar]
- 112.Koch AL. Shrinkage of growing Escherichia coli cells by osmotic challenge. J Bacteriol. 1984;159:919–924. doi: 10.1128/jb.159.3.919-924.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Koch AL, Woeste S. Elasticity of the sacculus of Escherichia coli. J Bacteriol. 1992;174:4811–4819. doi: 10.1128/jb.174.14.4811-4819.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Yao X, Jericho M, Pink D, Beveridge T. Thickness and elasticity of gram-negative murein sacculi measured by atomic force microscopy. J Bacteriol. 1999;181:6865–6875. doi: 10.1128/jb.181.22.6865-6875.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Sochacki KA, Shkel IA, Record MT, Weisshaar JC. Protein diffusion in the periplasm of E. coli under osmotic stress. Biophys J. 2011;100:22–31. doi: 10.1016/j.bpj.2010.11.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Vollmer W, Seligman SJ. Architecture of peptidoglycan: more data and more models. Trends Microbiol. 2010;18:59–66. doi: 10.1016/j.tim.2009.12.004. [DOI] [PubMed] [Google Scholar]
- 117.Jiang H, Sun SX. Morphology, growth, and size limit of bacterial cells. Phys Rev Lett. 2010;105:028101. doi: 10.1103/PhysRevLett.105.028101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Glauner B, Höltje JV, Schwarz U. The composition of the murein of Escherichia coli. J Biol Chem. 1988;263:10088–10095. [PubMed] [Google Scholar]
- 119.Vollmer W. Structural variation in the glycan strands of bacterial peptidoglycan. FEMS Microbiol Rev. 2008;32:287–306. doi: 10.1111/j.1574-6976.2007.00088.x. [DOI] [PubMed] [Google Scholar]
- 120.Lam H, et al. D-amino acids govern stationary phase cell wall remodeling in bacteria. Science. 2009;325:1552–1555. doi: 10.1126/science.1178123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Cava F, de Pedro MA, Lam H, Davis BM, Waldor MK. Distinct pathways for modification of the bacterial cell wall by non-canonical D-amino acids. EMBO J. 2011;30:3442–3453. doi: 10.1038/emboj.2011.246. Together with reference120, this paper shows that unusual D-amino acids are secreted and linked to peptidoglycan in many bacteria. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Lupoli TJ, et al. Transpeptidase-mediated incorporation of D-amino acids into bacterial peptidoglycan. J Am Chem Soc. 2011;133:10748–10751. doi: 10.1021/ja2040656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Shah IM, Laaberki MH, Popham DL, Dworkin J. A eukaryotic-like Ser/Thr kinase signals bacteria to exit dormancy in response to peptidoglycan fragments. Cell. 2008;135:486–496. doi: 10.1016/j.cell.2008.08.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Kolodkin-Gal I, et al. D -amino acids trigger biofilm disassembly. Science. 2010;328:627–629. doi: 10.1126/science.1188628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Butland G, et al. eSGA: E. coli synthetic genetic array analysis. Nature Methods. 2008;5:789–795. doi: 10.1038/nmeth.1239. [DOI] [PubMed] [Google Scholar]
- 126.Typas A, et al. High-throughput, quantitative analyses of genetic interactions in E coli. Nature Methods. 2008;5:781–787. doi: 10.1038/nmeth.1240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Nichols RJ, et al. Phenotypic landscape of a bacterial cell. Cell. 2011;144:143–156. doi: 10.1016/j.cell.2010.11.052. This article describes a high-throughput chemical genomic screen that provides links for the function of numerous orphan proteins in E. coli. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Andre G, et al. Imaging the nanoscale organization of peptidoglycan in living Lactococcus lactis cells. Nature Commun. 2010;1:27. doi: 10.1038/ncomms1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Scheuring S, Dufrene YF. Atomic force microscopy: probing the spatial organization, interactions and elasticity of microbial cell envelopes at molecular resolution. Mol Microbiol. 2010;75:1327–1336. doi: 10.1111/j.1365-2958.2010.07064.x. [DOI] [PubMed] [Google Scholar]
- 130.Hayhurst EJ, Kailas L, Hobbs JK, Foster SJ. Cell wall peptidoglycan architecture in Bacillus subtilis. Proc Natl Acad Sci USA. 2008;105:14603–14608. doi: 10.1073/pnas.0804138105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Turner RD, et al. Peptidoglycan architecture can specify division planes in Staphylococcus aureus. Nature Commun. 2010;1:26. doi: 10.1038/ncomms1025. [DOI] [PubMed] [Google Scholar]
- 132.Mingorance J, et al. Visualization of single Escherichia coli FtsZ filament dynamics with atomic force microscopy. J Biol Chem. 2005;280:20909–20914. doi: 10.1074/jbc.M503059200. [DOI] [PubMed] [Google Scholar]
- 133.Huang KC, Mukhopadhyay R, Wen B, Gitai Z, Wingreen NS. Cell shape and cell-wall organization in Gram-negative bacteria. Proc Natl Acad Sci USA. 2008;105:19282–19287. doi: 10.1073/pnas.0805309105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Kern T, et al. Dynamics characterization of fully hydrated bacterial cell walls by solid-state NMR: evidence for cooperative binding of metal ions. J Am Chem Soc. 2010;132:10911–10919. doi: 10.1021/ja104533w. [DOI] [PubMed] [Google Scholar]
- 135.Kern T, et al. Toward the characterization of peptidoglycan structure and protein–peptidoglycan interactions by solid-state NMR spectroscopy. J Am Chem Soc. 2008;130:5618–5619. doi: 10.1021/ja7108135. [DOI] [PubMed] [Google Scholar]
- 136.Alexeeva S, Gadella TW, Jr, Verheul J, Verhoeven GS, den Blaauwen T. Direct interactions of early and late assembling division proteins in Escherichia coli cells resolved by FRET. Mol Microbiol. 2010;77:384–398. doi: 10.1111/j.1365-2958.2010.07211.x. [DOI] [PubMed] [Google Scholar]
- 137.Osawa M, Anderson DE, Erickson HP. Reconstitution of contractile FtsZ rings in liposomes. Science. 2008;320:792–794. doi: 10.1126/science.1154520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Matias VR, Al-Amoudi A, Dubochet J, Beveridge TJ. Cryo-transmission electron microscopy of frozen-hydrated sections of Escherichia coli and Pseudomonas aeruginosa. J Bacteriol. 2003;185:6112–6118. doi: 10.1128/JB.185.20.6112-6118.2003. Together with reference82, this study demonstrates that membrane-attached FtsZ is sufficient to produce a constrictive force in vitro. [DOI] [PMC free article] [PubMed] [Google Scholar]