

Abstract
The first step of the base excision repair (BER) pathway responsible for removing oxidative DNA damage utilizes DNA glycosylases to find and remove the damaged DNA base. How glycosylases find the damaged base amidst a sea of undamaged bases has long been a question in the BER field. Single molecule total internal reflection fluorescence microscopy (SM TIRFM) experiments have allowed for an exciting look into this search mechanism and have found that DNA glycosylases scan along the DNA backbone in a bidirectional and random fashion. By comparing the search behavior of bacterial glycosylases from different structural families and with varying substrate specificities, it was found that glycosylases search for damage by periodically inserting a wedge residue into the DNA stack as they redundantly search tracks of DNA that are 450–600 bp in length. These studies open up a wealth of possibilities for further study in real time of the interactions of DNA glycosylases and other BER enzymes with various DNA substrates.
Keywords: Base excision repair, DNA glycosylases, Single molecule fluorescence, Search for radiation damage, Glycosylase diffusion, Wedge residue
Graphical abstract
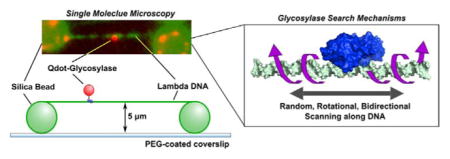
1. How are oxidized bases formed in cellular DNA?
Purine and pyrimidine bases in DNA are oxidized by reactive oxygen species (ROS) that are produced endogenously, by ionizing radiation and by a number of chemical agents (for reviews see [1,2]). ROS-induced oxidation produces a broad spectrum of DNA damages including damaged bases, sites of base loss and single strand breaks. There are a plethora of oxidized purine and pyrimidine bases produced by ROS (for reviews see [3,4]); the structures of these lesions were determined in the last century by radiation chemists who analyzed the damaged bases resulting from in vitro irradiation of nucleosides, nucleotides and DNA [5]. A similar base lesion spectrum results from endogenous ROS. The most common source of ROS in eukaryotic cells occurs in the mitochondria where about 1–1.5% of the oxygen intake results in the production of the superoxide anion radical, which can form hydrogen peroxide by dismutation, spontaneously or catalyzed by superoxide dismutase. In the presence of transition metals, hydroxyl radicals are produced from hydrogen peroxide in the so-called Fenton reaction. In addition, the superoxide anion radical can react with nitric oxide to produce peroxynitrite and other reactive species (for reviews see [6,7]). When the highly reactive hydroxyl radical interacts with DNA purines and pyrimidines, a wide variety of structurally diverse lesions are formed (see Fig. 1) (and for a review see [8]), about 5000–10,000 per human cell per day [9]. These oxidized DNA bases need to be removed from cellular DNA before they are encountered by DNA polymerases and result in mutation or cell lethality (for a review see [10]).
Fig. 1.

Structures of DNA bases and commonly occurring oxidative damages.
Of the four DNA bases, guanine has the lowest redox potential and hence is the most readily oxidized giving rise to 8-oxo-7,8-dihydro-guanine (8-oxoG) under oxidizing conditions, and 2,6-diamino-4-hydroxy-5-formamidopyrimidine (FapyG) under reducing conditions (for reviews see [11–14]). 8-OxoG in DNA also has a lower redox potential than the four normal bases and can be further oxidized to two major products, spiroiminodihydantoin (Sp) and guanidinohydantoin (Gh) (for a review see [15]). Attack on adenine by ROS gives rise to 8-oxo-7,8-dihydroadenine (8-oxoA) and 4,6-diamino-5-formamidopyri-midine (FapyA); 2-hydroxyadenine can also be formed [16]. 8-oxopur-ines and formamidopyrimidines are produced in DNA in both the presence and absence of oxygen, although the relative concentrations of oxygen and reducing agents affect the yields of each.
Thymine glycol and cytosine glycol are the most common products of oxidation of thymine and cytosine, respectively. Cytosine glycol is unstable and deaminates to uracil glycol, which can dehydrate to 5-hydroxyuracil; the direct dehydration product of cytosine glycol is 5-hydroxycytosine [11,12,17–19].
2. Which enzymes remove oxidized purines and pyrimidines from DNA?
Oxidized DNA bases are removed and repaired by the Base Excision Repair (BER) pathway (for reviews see [10,20–24]). DNA glycosylases are the first enzymes in this pathway and are responsible for locating and removing DNA base lesions. Because oxidized DNA bases comprise a diverse set of structures (Fig. 1), many of which are mutagenic, organisms had to develop a way to ensure genome stability. One could imagine a paradigm where each damaged base would have to have a specific glycosylase that recognized it, requiring an enormous number of functionally distinct enzymes. This has not proven to be the case. Instead, it turns out that each glycosylase has broad substrate specificity (Table 1), and a relatively small number of homologous enzymes are able to protect genomes that range in length from virus to human. The first glycosylase that was shown to recognize oxidized bases is endonuclease III, or Nth. This glycosylase was discovered in Escherichia coli based on its ability to recognize radiation-damaged DNA bases [25]. The substrates for Nth and its NTHL1 homologs include a broad spectrum of oxidized pyrimidines as well as FapyA (for reviews see [10,26]). Formamidopyrimidine DNA glycosylase, Fpg, was the next glycosylase identified in E. coli [27] and was isolated based on its ability to remove formamidopyrimidines from DNA. Fpg also removes 8-oxoG and the further oxidation products of 8-oxoG, Sp and Gh when they are positioned opposite C (for a review see [22]) (Table 1). Fpg is the only bacterial enzyme responsible for removing 8-oxoG and is also found in plants and some fungi [28]. However, the mammalian lineage uses a glycosylase that is structurally distinct from Fpg (see below), called OGG1, to remove 8-oxoG. OGG1 also removes formamidopyrimidines but not Sp or Gh [29–31]. Endonuclease VIII (Nei), which has significant substrate overlap with Nth and some substrate overlap with Fpg, was identified much later [32]. Nei [32,33] and its homologs, the NEIL [34,35] glycosylases, recognize a broad spectrum of pyrimidines, formamidopyrimidines [35–38] and Sp and Gh [39–41] (Table 1). Clearly there is a substantial redundancy in the substrate specificities of the glycosylases that recognize and remove oxidized DNA bases.
Table 1.
Substrates for Escherichia coli Nth, Nei and Fpg [26]. Substrates are color coded as follows: black text for substrates common to all three enzymes,
 text for substrates common to E. coli Nth and Nei,
text for substrates common to E. coli Nth and Nei,
 text for substrates common to Nth and Fpg,
text for substrates common to Nth and Fpg,
 lettering for substrates unique to the given enzyme. It should be noted that Table 1 only lists the substrates tested for each particular E. coli enzyme and does not reflect relative activities, whether the substrate was tested and found to be negative, or whether the substrate was tested with other enzymes.
lettering for substrates unique to the given enzyme. It should be noted that Table 1 only lists the substrates tested for each particular E. coli enzyme and does not reflect relative activities, whether the substrate was tested and found to be negative, or whether the substrate was tested with other enzymes.
| Nth | 2,6-diamino-4-oxo-5-formamidopyrimidine (FapyG) |
| guanidinohydantoin | |
| spiroiminodihydantoin | |
| 4,6-diamino-5-formamidopyrimidine (FapyA) | |
| abasic (AP) sites | |
| 5,6-dihydroxy-5,6-dihydrothymine (thymine glycol) | |
| 5,6-dihydrothymine | |
| 5-hydroxy-6-hydrothymine | |
| 5-hydroxy-5-methylhydatoin | |
| urea | |
| 5-hydroxycytosine | |
| 5,6-dihydroxycytosine | |
| 5,6-dihydroxy-5,6-dihydrouracil (uracil glycol) | |
| 5-hydroxyuracil | |
| 5,6 dihydrouracil | |
| 5,6-dihydroxyuracil | |
| 2,6-diamino-4-oxo-5-(N-methyl)formamidopyrimidine | |
| 6-hydroxy-5,6-dihydrothymine | |
| N-(2-deoxy-β-D-erythropentofuranosyl)-N-3-[2R]-hydroxyisobutyric acid) urea | |
| methyltartronylurea | |
| 5,6-dihydroxy-5,6 dihydrocytosine (cytosine glycol) | |
| 5-formyluracil | |
| 5,5-dihydroxypyrimidine-2,4,6(lH,3H,5H)-trione (alloxan) | |
| 5-hydroxyhydantoin | |
| trans-l-carbamoyl-2-oxo-4,5-dihydroxyimidazolidine | |
| l,N6ethenoadenine degradation product | |
|
| |
| Nei | 2,6-diamino-4-oxo-5-formamidopyrimidine |
| guanidinohydantoin | |
| spiroiminodihydantoin | |
| 4,6-diamino-5-formamidopyrimidine (FapyA) | |
| abasic (AP) sites | |
| 5,6-dihydroxy-5,6-dihydrothymine | |
| 5,6-dihydrothymine | |
| 5-hydroxy-6-hydrothymine | |
| 5-hydroxy-5-methylhydatoin | |
| urea | |
| 5-hydroxycytosine | |
| 5,6-dihydroxycytosine | |
| 5,6-dihydroxy-5,6-dihydrouracil (uracil glycol) | |
| 5-hydroxyuracil | |
| 5,6 dihydrouracil | |
| 5,6-dihydroxyuracil | |
| β-ureidoisobutyric acid | |
| 5-hydroxy-6-hydrouracil | |
| oxanine | |
| xanthine (3,7-dihydropurine-2,6-dione) | |
|
| |
| Fpg | 2,6-diamino-4-oxo-5-formamidopyrimidine (FapyG) |
| guanidinohydantoin | |
| spiroiminodihydantoin | |
| 4,6-diamino-5-formamidopyrimidine (FapyA) | |
| abasic (AP) sites | |
| 2,6-diamino-4-oxo-5-(N-methyl)formamidopyrimidine | |
| 8-oxo-7,8-dihydroadenine | |
| 8-oxo-7,8-dihydroguanine | |
The DNA glycosylases specific for oxidized purines and pyrimidines fall into two structural families, the HhH superfamily, which includes the Nth orthologs and OGG1 (for a review see [42]), and the Fpg/Nei family [43,44]. Both these glycosylase families also contain lyase activities which, after removal of the damaged base, cleave the DNA backbone leaving two possible nicked products. The HhH superfamily leaves a 3′ α,β-unsaturated aldehyde and a 5′ phosphate, while the Fpg/Nei family leaves a 3′ and a 5′ phosphate (Fig. 2). Crystal structures of glycosylases with target DNA characteristically show that the enzyme binds to DNA in the minor groove, bends the DNA backbone, and extrudes the lesion from the major groove into the enzyme’s active pocket (for reviews see [42,45–47]). It had been assumed that lesion identification and lesion specificity were determined by interactions between the damaged base and glycosylase amino acids in the active site pocket. This proposal was supported by the structures of uracil DNA glycosylase [48–50] and OGG1 [51] with their respective lesions. However, both of these enzymes have fairly narrow substrate specificities. Structures of Fpg show that the specificity of Fpg for 8-oxoG relies on a structural loop that covers the active site pocket and forms specific interactions with 8-oxoG thus stabilizing it in this pocket [52]. Members of the Fpg/Nei family lacking this structural loop do not remove 8-oxoG [53]. Obtaining structures of the glycosylases that remove other oxidized bases and have broad substrate specificities has proven to be decidedly more difficult. There are currently no structures of Nth or its homologs with a lesion in the active site pocket. Only recently has a structure of a NEIL1 ortholog bound to DNA containing either thymine glycol (Tg) or 5-hydroxyuracil (5OHU) been solved [54]. Interestingly, these structures revealed that there were no specific interactions between the enzyme’s amino acids and the lesions. A recent crystal structure of NEIL1 together with Tg-containing DNA [55] did show that NEIL1 promoted a tautomeric shift in Tg that promoted its stable binding in the active site pocket and facilitated excision. However, taken together, these observations beg the question of how the glycosylases locate an oxidized DNA base lesion in the sea of undamaged bases?
Fig. 2.
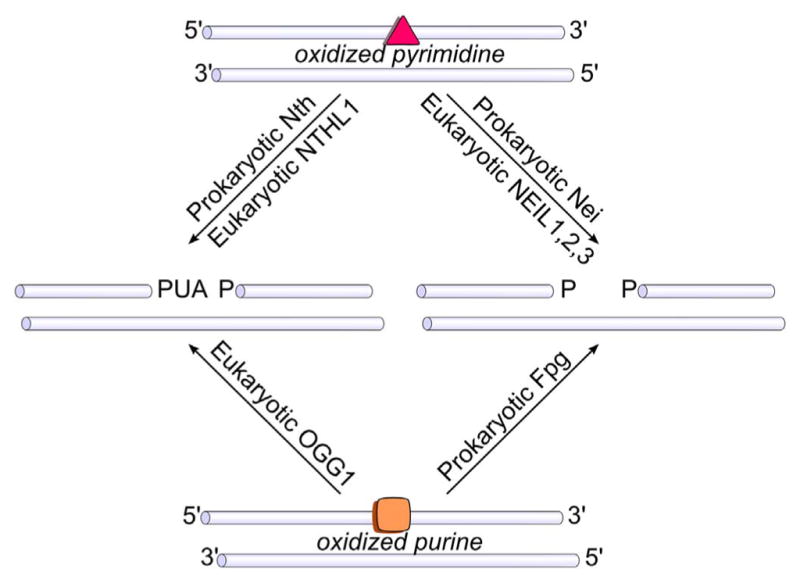
Products of the associated lyase activities of the DNA glycosylases. Abbreviations: PUA, α, β-unsaturated aldehyde; P, phosphate.
3. How do the DNA glycosylases search for oxidatively damaged DNA bases?
The glycosylase search for oxidatively damaged DNA bases is the very first step of the BER pathway, and yet there are still many things that are unknown about how glycosylases identify the slight physical changes of the damaged DNA base amidst an extensive array of undamaged base pairs. One of the amazing things about DNA glycosylases is that they are able to locate damages using only thermal energy as a driving force. The copy number of glycosylases in the cell is low when compared with the sheer magnitude of DNA bases, which makes it physically impossible for the enzymes to efficiently encounter damages through random three-dimensional collisions. There have been several suggestions as to how a glycosylase might capitalize on the polymer structure of DNA to help find damage sites. Sliding along the DNA backbone without release is often referred to as “facilitated diffusion,” but glycosylases may also make small “hops” between close sites on the DNA or traverse longer distances through intrastrand and/ or interstrand transfer events (for reviews see [56–58]). Ensemble methods measuring correlated cleavage of closely spaced damage sites in long oligodeoxyribonucleotides provided evidence that the glycosylases were capable of remaining in continuous contact with the DNA molecule after removing a lesion [59–66]. However, direct visualization of the movement of glycosylases along longer tracts of undamaged DNA remained a challenge until a breakthrough occurred using single molecule (SM) fluorescence microscopy imaging.
SM microscopy experiments of glycosylases use total internal reflection fluorescence microscopy (TIRFM) for the observation in real time of a single fluorescently-tagged glycosylase enzyme as it moves along an individual DNA molecule. Since glycosylases encounter long regions of undamaged DNA in vivo, special long DNA substrates and sample conditions were developed to allow for observation of these interactions. The first experiments measuring glycosylase diffusion used lambda DNA that was tethered to a polyethylene surface through a single biotin linkage, and the DNA was stretched under continuous buffer flow during the experiment [67] providing 48 kilobases of extended undamaged DNA. The first glycosylase studied was human OGG1, which was labeled at a cysteine residue with a fluorescent dye for visualization of scanning behavior. The observed movement of OGG1 was consistent with one-dimensional facilitated diffusion, showing random, bidirectional sliding along tracts of DNA that were several thousand bases in length. OGG hopping on the DNA molecule was not observed, and the binding lifetimes were found to be quite short. The mean squared displacement (MSD) versus time was plotted for each displacement trajectory, and a diffusion constant was then determined for each individual OGG1 molecule. Further studies using these techniques showed that the diffusive properties of OGG1 were consistent with rotational diffusion along the DNA backbone [68].
In order to characterize possible differences in scanning behavior among glycosylases with different structures and substrate specificities, our laboratory went on to examine the diffusive properties of E. coli Fpg, Nei and Nth [69,70]. In these experiments, a different methodology was utilized to immobilize and stretch lambda DNA. Silica beads were used to elevate the DNA above the surface to create DNA tightropes that remain extended in the absence of flow (Fig. 3). Glycosylases were labeled with quantum dots, allowing for very long observation times without photobleaching. The three E. coli glycosylases all scanned along the DNA in a manner similar to that of OGG1. All moved randomly in both directions and scanned repeatedly over a single area of DNA with high redundancy. One difference observed between OGG1 and the E. coli enzymes was that all of the E. coli enzymes showed longer binding lifetimes than reported for OGG1, but it is unclear whether flow conditions might have affected these values. Initially, the single molecule trajectories were analyzed using the methods described for OGG1 wherein a single diffusion constant is fit to each trajectory, and like OGG1, the diffusion constants of Fpg, Nei and Nth were consistent with rotational diffusion along the DNA backbone (Fig. 4). A further examination of the displacement versus time data revealed that rates of diffusion were highly variable within a given displacement trajectory, suggesting that an individual glycosylase switches between fast and slow movement. For this reason, a sliding window analysis was developed to allow for characterization of diffusive heterogeneity within a single trajectory (Fig. 5). What is fascinating is that all three E. coli enzymes exhibited a very broad distribution of diffusion constants that ranged from 0.001 to 0.1 μm2/ second.
Fig. 3.
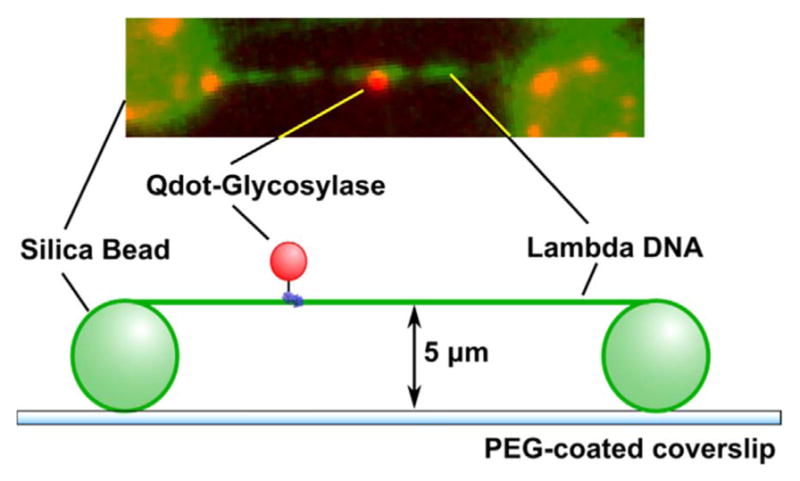
Sample configuration for TIRFM experiments with DNA tightropes. Lambda DNA is extended across polylysine-coated silica beads which are nonspecifically adhered to a PEG-coated coverslip. The six-histidine tagged glycosylase is labeled through a biotinylated anti-his antibody and a streptavidin coated Qdot (not to scale).
Fig. 4.

Diffusive behavior of WT and wedge variant DNA glycosylases. A, B, and C show the mean diffusion constant for Fpg (A), Nei (B), and Nth (C) wild-type and wedge variants scanning along undamaged lambda DNA and lambda DNA containing specific damages. The damaged purines for Fpg (A) were generated using methylene blue plus visible light, while damaged pyrimidines for Nei (B) and Nth (C) were generated using osmium tetroxide. The details for these experiments can be found in Ref. [70] (***P < 0.001).
Fig. 5.

Diffusive behavior of WT (blue) and wedge variant (green) DNA glycosylases on undamaged lambda DNA. Columns A, B, and C show the time-weighted distribution of diffusive behavior for Fpg (A), Nei (B), and Nth (C). Details for these experiments can be found in Ref. [70].
Our lab then went on to examine the structural features of the DNA glycosylases that may play a role in the search process in order to answer the question of how the enzymes actually probe for oxidatively damaged DNA bases. A search mechanism in which every base in the DNA stack is tested by capturing an extruded form is thermodynamically impossible; thus, Blainey and coworkers [67] proposed that glycosylases search a small region of DNA with high redundancy, and the lesion is selected because it is extruded at a higher rate than other bases. Although the high search redundancy observed in the SM experiments was consistent with this proposal, we suspected the mechanism might be more complicated. Our studies were guided by two observations: First, while scanning DNA, the enzymes exhibited a broad range of diffusion constants, suggesting that they were undergoing a combination of slow and fast diffusion processes that differed from one another and might correspond to two search modes (Fig. 5). Secondly, the Verdine laboratory had crystallized Geobacter stereothermophilus Fpg covalently bound to undamaged DNA and observed an amino acid inserted into the DNA molecule suggesting that Fpg might be utilizing this amino acid residue to actively probe the base stack while scanning [71]. It had been known from many structural studies with glycosylases bound to damage-containing DNA that the DNA damage is extruded into the active side pocket of the enzyme and three amino acids, insertion residues, are inserted into the DNA molecule to stabilize both the DNA helix and the extruded base in the active site pocket of the glycosylase (Fig. 6) (see for examples [43,44,72]). However, the Verdine lab observed that one of these three amino acids was inserted into undamaged DNA in the absence of base eversion, and this same residue actually stacked between two bases opposite the extruded lesion in damaged DNA. Thus, it seemed propitious to ask whether this amino acid, this “wedge residue,” might affect how the glycosylase travels along undamaged DNA.
Fig. 6.
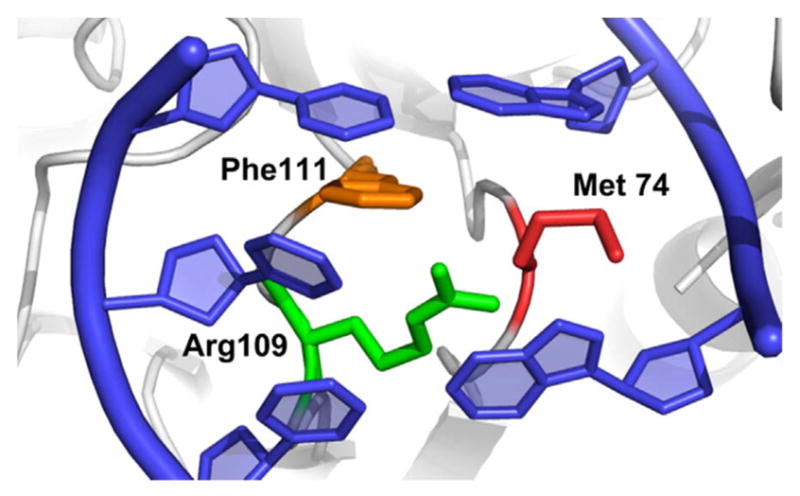
Crystal structure of the DNA helix following eversion of the damaged base into the active site of Fpg. The wedge residue (Phe111) is orange, while the other two residues in the intercalation loop Arg109 and Met 74 are green and red, respectively. (PDB 1K82).
To investigate the role of the wedge residue in glycosylase scanning on undamaged DNA, the wedge residues of all three bacterial DNA glycosylases were substituted with alanine, and the diffusion of these variants was observed on lambda DNA tightropes using TIRFM microscopy. In all cases, the diffusion constants of the wedge variants were much faster than those of the corresponding wild type enzymes (Figs. 4 and 5). These changes appeared to be due to the loss of the slower portion of the distribution of diffusion constants, which is consistent with the variants lacking a slow diffusion search mode linked to the insertion of the wedge residue into the undamaged DNA base stack. We conclude that the function of the wedge amino acid is to sense damaged DNA bases before eversion of the damaged base into the enzyme’s active site. In fact, more recent structural studies with the G. stearothermophilus Fpg and undamaged DNA show that its wedge residue, phenylalanine 114, can insert into sequential, neighboring DNA bases, and that these target bases remain intrahelical and stacked [73]. In addition, the Fedorova laboratory was able to use stopped flow kinetics to study early stages in the process of lesion recognition, and they have shown that the Fpg wedge residue disrupts structural stability around the lesion to promote eversion [74]. Once the lesion has been everted, the wedge residue, together with the remaining two insertion residues, occupy the space left by the everted base thus stabilizing the DNA duplex together with the everted base in the enzyme’s active site pocket. These workers used similar techniques to also show that the early steps of lesion recognition by Nth involve insertion of Leu81, its wedge residue, into the base stack [75].
4. How do glycosylases behave when they locate a damaged base?
Although our studies on undamaged DNA provided insight into how the glycosylases probe DNA for damages, these data did not tell us what happens when the enzyme encounters the damaged base. To address this question, we randomly introduced damages into lambda DNA. Oxidized purines were generated using methylene blue plus visible light, and oxidized pyrimidines, primarily Tg, were produced by osmium tetroxide treatment [70]. DNA tightropes were prepared under two conditions: low levels (~110 damages/lambda DNA molecule) and high levels (~300 damages/lambda DNA molecule) of the random damage. The diffusive behavior of Fpg on its substrate, oxidized purine-containing DNA (Fig. 7A), and the diffusive behavior of Nei and Nth on their substrate, DNA containing Tg (Fig. 7B and C, respectively) were then examined. For all three glycosylases, the binding lifetimes increased and the diffusion constants decreased in the presence of the damage that they recognized [70]. These changes were even more pronounced at higher levels of damage, confirming that the slower scanning and increased residence time of a glycosylase on DNA is due to its interaction with the damage. Consistent with these findings, when the diffusive behavior of Fpg was examined on osmium tetroxide-treated DNA, which is not a substrate for Fpg, Fpg behavior was similar to what was observed in the absence of damage [70]. Further analysis of the scanning trajectories provided additional information about what happens after processing of a lesion. For example, it was found that approximately a third of Fpg glycosylases continue to scan without dissociating from the DNA after stopping at a damage [70]. From these studies it may be concluded, that regardless of structure or substrate specificity, the diffusive behavior of these E. coli glycosylases is greatly modulated by the presence of specifically recognized damage sites. Moreover, the longer binding lifetimes likely reflect the process of damage recognition, stopping, and in all probability, removing the damaged base.
Fig. 7.

Diffusive behavior of WT and wedge variant DNA glycosylases on undamaged lambda DNA and lambda DNA that contains randomly distributed damages. Columns A, B, and C show the time-weighted distribution of diffusive behavior for Fpg (A), Nei (B), and Nth (C). The light blue distributions in the top row are WT enzyme on undamaged lambda DNA, the dark blue distributions in the second row are WT enzyme in the presence of randomly distributed specific damage in lambda DNA, and the dark green distributions in the third row show the wedge variant enzyme in the presence of randomly distributed specific damage in lambda DNA. Damaged purines for Fpg (A) were generated using methylene blue plus visible light, while damaged pyrimidines for Nei (B) and Nth (C) were generated using osmium tetroxide. Details for these experiments can be found in Ref. [70].
Using the diffusive behavior of Fpg, Nei and Nth, together with kinetics data from the Fedorova lab describing individual steps of DNA glycosylase activity [76–80], a simple simulation was developed to interpret the mechanistic implications of the single molecule data [70]. During a scanning event in which the glycosylase tracks along the DNA backbone at a rate of 0.05 μm2/second, the wedge is inserted into the DNA at a rate of 2000 s−1 and removed from the DNA at a slower rate of 500 s−1. If, while the wedge is inserted into the DNA, a damage is detected, the enzyme stops and remains bound to the lesion. The lesion-bound wild-type enzyme most often moves forward through catalysis, but rarely, may also be released from the damage before it is repaired. One interesting finding from our SM studies is that the wedge variant enzymes still locate the damage, as shown by slower scanning in the presence of damaged DNA. However, according to our model, when a variant enzyme lacking the wedge residue is bound to a damage, it is much more likely than wild type to release before catalysis can take place. These findings explain how mutating the wedge residue greatly decreases enzyme activity observed in gel-based ensemble assays. One unexpected prediction of our model is that checking with the wedge is periodic, not constant. Thus, the high redundancy of glycosylase scanning along a region of DNA is necessary for adequate coverage and discovery of the damaged sites. Consistent with what has been measured in ensemble correlated cleavage experiments, our diffusion constants and binding lifetimes predict a contact distance of 450–600 base pairs. However, these bases are scanned repeatedly, allowing for wedge interrogation of the majority of them. Given our predicted scanning rates, the size of the E. coli genome, and the copy numbers of these glycosylases, we predict that a given site within the E. coli genome will see each of these glycosylases, Fpg, Nei, and Nth approximately once every 10 min, resulting in continual removal of oxidized purines and pyrimidines.
5. Future directions
Although SM studies have led to several exciting discoveries about the glycosylase search for oxidative DNA damages, many questions remain. So far, the bulk of the reported studies have focused on E. coli DNA glycosylases, leaving the basic question of whether eukaryotic/ human enzymes scan along undamaged DNA in a way that is similar to what has been observed. Does scanning behavior differ for human glycosylases such as NEIL1, which are suspected to function as part of a larger repair complex? One of the properties that we might expect to vary greatly between bacterial and eukaryotic DNA glycosylases is the number of base pairs scanned in each encounter. These numbers are affected by both the diffusion constant and the binding lifetime. Also, it would be interesting to know if the wedge residue plays a critical role in the search process of eukaryotic glycosylases, and if wedge insertion is a general paradigm in the glycosylase search for damage. Continuing along these lines, what is the role of other residues in the intercalation loop, and what does scanning look like for a glycosylase such as MvNei2/3 [81] and NEIL3 [82], that essentially lack an intercalation loop?
It has been shown that eukaryotic DNA glycosylases often show very high affinity and long binding lifetimes to their abasic site products, which serve to protect the abasic site until it can be handed off to the next enzyme in the BER cascade (for reviews see [83,84]). However, these handoff events have never been directly observed for any member of the BER pathway. SM studies present an exciting opportunity to watch these handoff events in real time. Furthermore, the motion of other BER enzymes on DNA has not yet been observed. Of course, these experiments require monitoring a specific damage site in the DNA, and techniques have recently been developed that allow for the construction of long substrates that contain damage sites in predetermined locations throughout the DNA [85]. Through these methods, it will be possible to create DNA containing almost any damaged base that can be synthesized in an oligodeoxyribonucleotide, as well as a variety of structural variations in the DNA duplex, such as hairpins and bubble regions. These extended DNA substrates will yield rich information about glycosylase scanning in the presence of specific substrates. However, eukaryotic DNA exists wrapped around histones as tightly packed chromatin fibers. Recent single molecule tracking experiments in the presence of histone-wrapped DNA [86–88] demonstrate that it will also be feasible to monitor DNA glycosylase interactions with these substrates. Thus, it may be possible to observe whether glycosylases and other BER enzymes are able to disrupt wrapping of the nucleosome during the process of repair. SM TIRFM experiments hold great promise for exciting new insights into the glycosylase search for oxidatively-induced DNA damage.
Acknowledgments
The authors would like to thank the National Institutes of Health Grant P01 CA098993, awarded by the National Cancer Institute, for supporting the work in our laboratory. The authors are also grateful to our collaborator, Dr. David Warshaw, for his expertise, to Dr. Shane Nelson for computational simulation of the data and to Dr. Andrew Dunn for initiating the SM studies in our laboratory.
References
- 1.Halliwell B, Gutteridge JMC. Free Radicals in Biology and Medicine. Oxford University Press; New York, NY: 2007. [Google Scholar]
- 2.Jezek P, Hlavata L. Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism. Int J Biochem Cell Biol. 2005;37(12):2478–2503. doi: 10.1016/j.biocel.2005.05.013. [DOI] [PubMed] [Google Scholar]
- 3.Dizdaroglu M. Oxidatively induced DNA damage and its repair in cancer. Mutat Res Rev Mutat Res. 2015;763:212–245. doi: 10.1016/j.mrrev.2014.11.002. [DOI] [PubMed] [Google Scholar]
- 4.Cadet J, Wagner JR. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb Perspect Biol. 2013;5(2) doi: 10.1101/cshperspect.a012559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.von Sonntag C. The Chemical Basis of Radiation Biology. Taylor & Francis; London and New York, NY: 1987. [Google Scholar]
- 6.Balazy M, Nigam S. Aging, lipid modifications and phospholipases – new concepts. Ageing Res Rev. 2003;2(2):191–209. doi: 10.1016/s1568-1637(02)00065-x. [DOI] [PubMed] [Google Scholar]
- 7.Boveris A, Cadenas E. Mitochondrial production of hydrogen peroxide regulation by nitric oxide and the role of ubisemiquinone. IUBMB Life. 2000;50(4–5):245–250. doi: 10.1080/713803732. [DOI] [PubMed] [Google Scholar]
- 8.Cadet J, Wagner JR. Oxidatively generated base damage to cellular DNA by hydroxyl radical and one-electron oxidants: similarities and differences. Arch Biochem Biophys. 2014;557:47–54. doi: 10.1016/j.abb.2014.05.001. [DOI] [PubMed] [Google Scholar]
- 9.Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, Ellenberger T. DNA Repair and Mutagenesis. 2. ASM Press; Washington, DC: 2005. [Google Scholar]
- 10.Duclos S, Doublié S, Wallace SS. Consequences and repair of oxidative DNA damage. In: Greim H, Albertini RJ, editors. The Cellular Response to the Genotoxic Insult: The Question of Threshold for Genotoxic Carcinogens. The Royal Society of Chemistry; Cambridge, United Kingdom: 2012. pp. 109–153. [Google Scholar]
- 11.Breen AP, Murphy JA. Reactions of oxyl radicals with DNA. Free Radic Biol Med. 1995;18(6):1033–1077. doi: 10.1016/0891-5849(94)00209-3. [DOI] [PubMed] [Google Scholar]
- 12.Dizdaroglu M. Oxidative damage to DNA in mammalian chromatin. Mutat Res. 1992;275(3–6):331–342. doi: 10.1016/0921-8734(92)90036-o. [DOI] [PubMed] [Google Scholar]
- 13.Dizdaroglu M. Mechanisms of free radical damage to DNA. In: Aruoma OI, Halliwell B, editors. DNA & Free Radicals: Techniques, Mechanisms & Applications. OICA International (UK) Ltd; St. Lucia: 1998. pp. 3–26. [Google Scholar]
- 14.Steenken S. Purine bases, nucleosides, and nucleotides: aqueous solution redox chemistry and transformation reactions of their radical cations and e- and OH adducts. Chem Rev. 1989;89(3):503–520. [Google Scholar]
- 15.Neeley WL, Essigmann JM. Mechanisms of formation, genotoxicity, and mutation of guanine oxidation products. Chem Res Toxicol. 2006;19(4):491–505. doi: 10.1021/tx0600043. [DOI] [PubMed] [Google Scholar]
- 16.Lesiak KB, Wheeler KT. Formation of alpha-deoxyadenosine in polydeoxynucleotides exposed to ionizing radiation under anoxic conditions. Radiat Res. 1990;121(3):328–337. [PubMed] [Google Scholar]
- 17.Dizdaroglu M, Holwitt E, Hagan MP, Blakely WF. Formation of cytosine glycol and 5,6-dihydroxycytosine in deoxyribonucleic acid on treatment with osmium tetroxide. Biochem J. 1986;235(2):531–536. doi: 10.1042/bj2350531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Dizdaroglu M, Laval J, Boiteux S. Substrate specificity of the Escherichia coli endonuclease III: excision of thymine- and cytosine-derived lesions in DNA produced by radiation-generated free radicals. Biochemistry. 1993;32(45):12105–12111. doi: 10.1021/bi00096a022. [DOI] [PubMed] [Google Scholar]
- 19.Wagner JR, Cadet J. Oxidation reactions of cytosine DNA components by hydroxyl radical and one-electron oxidants in aerated aqueous solutions. Acc Chem Res. 2010;43(4):564–571. doi: 10.1021/ar9002637. [DOI] [PubMed] [Google Scholar]
- 20.David SS, O’Shea VL, Kundu S. Base-excision repair of oxidative DNA damage. Nature. 2007;447(7147):941–950. doi: 10.1038/nature05978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kim YJ, Wilson DM., 3rd Overview of base excision repair biochemistry. Curr Mol Pharmacol. 2012;5(1):3–13. doi: 10.2174/1874467211205010003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Prakash A, Doublie S, Wallace SS. The Fpg/Nei family of DNA glycosylases: substrates, structures, and search for damage. Prog Mol Biol Transl Sci. 2012;110:71–91. doi: 10.1016/B978-0-12-387665-2.00004-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wallace SS. DNA glycosylases search for and remove oxidized DNA bases. Environ Mol Mutagen. 2013;54(9):691–704. doi: 10.1002/em.21820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wilson DM, 3rd, Sofinowski TM, McNeill DR. Repair mechanisms for oxidative DNA damage. Front Biosci. 2003;8:d963–d981. doi: 10.2741/1109. [DOI] [PubMed] [Google Scholar]
- 25.Strniste GF, Wallace SS. Endonucleolytic incision of x-irradiated deoxyribonucleic acid by extracts of Escherichia coli. Proc Natl Acad Sci USA. 1975;72(6):1997–2001. doi: 10.1073/pnas.72.6.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lee AJ, Wallace SS. The DNA glycosylases that recognize and remove free radical-damaged pyrimidines. In: Wilson DM III, editor. DNA Damage and Repair: BER from Molecular Biology to Disease Detection and Therapy. World Scientific Publishing; 2016. [Google Scholar]
- 27.Chetsanga CJ, Lindahl T. Release of 7-methylguanine residues whose imidazole rings have been opened from damaged DNA by a DNA glycosylase from Escherichia coli. Nucleic Acids Res. 1979;6(11):3673–3684. doi: 10.1093/nar/6.11.3673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kathe SD, Barrantes-Reynolds R, Jaruga P, Newton MR, Burrows CJ, Bandaru V, Dizdaroglu M, Bond JP, Wallace SS. Plant and fungal Fpg homologs are formamidopyrimidine DNA glycosylases but not 8-oxoguanine DNA glycosylases. DNA Repair (Amst) 2009;8(5):643–653. doi: 10.1016/j.dnarep.2008.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Asagoshi K, Yamada T, Terato H, Ohyama Y, Monden Y, Arai T, Nishimura S, Aburatani H, Lindahl T, Ide H. Distinct repair activities of human 7,8-dihydro-8-oxoguanine DNA glycosylase and formamidopyrimidine DNA glycosylase for formamidopyrimidine and 7,8-dihydro-8-oxoguanine. J Biol Chem. 2000;275(7):4956–4964. doi: 10.1074/jbc.275.7.4956. [DOI] [PubMed] [Google Scholar]
- 30.Bjoras M, Luna L, Johnsen B, Hoff E, Haug T, Rognes T, Seeberg E. Opposite base-dependent reactions of a human base excision repair enzyme on DNA containing 7,8-dihydro-8-oxoguanine and abasic sites. EMBO J. 1997;16(20):6314–6322. doi: 10.1093/emboj/16.20.6314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zharkov DO. Base excision DNA repair. Cell Mol Life Sci. 2008;65(10):1544–1565. doi: 10.1007/s00018-008-7543-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Melamede RJ, Hatahet Z, Kow YW, Ide H, Wallace SS. Isolation and characterization of endonuclease VIII from Escherichia coli. Biochemistry. 1994;33(5):1255–1264. doi: 10.1021/bi00171a028. [DOI] [PubMed] [Google Scholar]
- 33.Jiang D, Hatahet Z, Melamede RJ, Kow YW, Wallace SS. Characterization of Escherichia coli endonuclease VIII. J Biol Chem. 1997;272(51):32230–32239. doi: 10.1074/jbc.272.51.32230. [DOI] [PubMed] [Google Scholar]
- 34.Bandaru V, Sunkara S, Wallace SS, Bond JP. A novel human DNA glycosylase that removes oxidative DNA damage and is homologous to Escherichia coli endonuclease VIII. DNA Repair. 2002;1(7):517–529. doi: 10.1016/s1568-7864(02)00036-8. [DOI] [PubMed] [Google Scholar]
- 35.Hazra TK, Izumi T, Boldogh I, Imhoff B, Kow YW, Jaruga P, Dizdaroglu M, Mitra S. Identification and characterization of a human DNA glycosylase for repair of modified bases in oxidatively damaged DNA. Proc Natl Acad Sci USA. 2002;99(6):3523–3528. doi: 10.1073/pnas.062053799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Hu J, de Souza-Pinto NC, Haraguchi K, Hogue BA, Jaruga P, Greenberg MM, Dizdaroglu M, Bohr VA. Repair of formamidopyrimidines in DNA involves different glycosylases: role of the OGG1, NTH1, and NEIL1 enzymes. J Biol Chem. 2005;280(49):40544–40551. doi: 10.1074/jbc.M508772200. [DOI] [PubMed] [Google Scholar]
- 37.Jaruga P, Birincioglu M, Rosenquist TA, Dizdaroglu M. Mouse NEIL1 protein is specific for excision of 2,6-diamino-4-hydroxy-5-formamidopyrimidine and 4,6-diamino-5-formamidopyrimidine from oxidatively damaged DNA. Biochemistry. 2004;43(50):15909–15914. doi: 10.1021/bi048162l. [DOI] [PubMed] [Google Scholar]
- 38.Wiederholt CJ, Patro JN, Jiang YL, Haraguchi K, Greenberg MM. Excision of formamidopyrimidine lesions by endonucleases III and VIII is not a major DNA repair pathway in Escherichia coli. Nucleic Acids Res. 2005;33(10):3331–3338. doi: 10.1093/nar/gki655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hailer MK, Slade PG, Martin BD, Rosenquist TA, Sugden KD. Recognition of the oxidized lesions spiroiminodihydantoin and guanidinohydantoin in DNA by the mammalian base excision repair glycosylases NEIL1 and NEIL2. DNA Repair. 2005;4(1):41–50. doi: 10.1016/j.dnarep.2004.07.006. [DOI] [PubMed] [Google Scholar]
- 40.Hazra TK, Muller JG, Manuel RC, Burrows CJ, Lloyd RS, Mitra S. Repair of hydantoins, one electron oxidation product of 8-oxoguanine, by DNA glycosylases of Escherichia coli. Nucleic Acids Res. 2001;29(9):1967–1974. doi: 10.1093/nar/29.9.1967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Krishnamurthy N, Zhao X, Burrows CJ, David SS. Superior removal of hydantoin lesions relative to other oxidized bases by the human DNA glycosylase hNEIL1. Biochemistry. 2008;47(27):7137–7146. doi: 10.1021/bi800160s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Fromme JC, Banerjee A, Verdine GL. DNA glycosylase recognition and catalysis. Curr Opin Struct Biol. 2004;14(1):43–49. doi: 10.1016/j.sbi.2004.01.003. [DOI] [PubMed] [Google Scholar]
- 43.Zharkov DO, Golan G, Gilboa R, Fernandes AS, Gerchman SE, Kycia JH, Rieger RA, Grollman AP, Shoham G. Structural analysis of an Escherichia coli endonuclease VIII covalent reaction intermediate. EMBO J. 2002;21(4):789–800. doi: 10.1093/emboj/21.4.789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gilboa R, Zharkov DO, Golan G, Fernandes AS, Gerchman SE, Matz E, Kycia JH, Grollman AP, Shoham G. Structure of formamidopyrimidine-DNA glycosylase covalently complexed to DNA. J Biol Chem. 2002;277(22):19811–19816. doi: 10.1074/jbc.M202058200. [DOI] [PubMed] [Google Scholar]
- 45.Hitomi K, Iwai S, Tainer JA. The intricate structural chemistry of base excision repair machinery: implications for DNA damage recognition, removal, and repair. DNA Repair. 2007;6(4):410–428. doi: 10.1016/j.dnarep.2006.10.004. [DOI] [PubMed] [Google Scholar]
- 46.Huffman JL, Sundheim O, Tainer JA. DNA base damage recognition and removal: new twists and grooves. Mutat Res. 2005;577(1–2):55–76. doi: 10.1016/j.mrfmmm.2005.03.012. [DOI] [PubMed] [Google Scholar]
- 47.Yang W. Structure and mechanism for DNA lesion recognition. Cell Res. 2008;18(1):184–197. doi: 10.1038/cr.2007.116. [DOI] [PubMed] [Google Scholar]
- 48.Kavli B, Slupphaug G, Mol CD, Arvai AS, Peterson SB, Tainer JA, Krokan HE. Excision of cytosine and thymine from DNA by mutants of human uracil-DNA glycosylase. EMBO J. 1996;15(13):3442–3447. [PMC free article] [PubMed] [Google Scholar]
- 49.Savva R, McAuley-Hecht K, Brown T, Pearl L. The structural basis of specific base-excision repair by uracil-DNA glycosylase. Nature. 1995;373(6514):487–493. doi: 10.1038/373487a0. [DOI] [PubMed] [Google Scholar]
- 50.Slupphaug G, Mol CD, Kavli B, Arvai AS, Krokan HE, Tainer JA. A nucleotide-flipping mechanism from the structure of human uracil-DNA glycosylase bound to DNA. Nature. 1996;384(6604):87–92. doi: 10.1038/384087a0. [DOI] [PubMed] [Google Scholar]
- 51.Bruner SD, Norman DP, Verdine GL. Structural basis for recognition and repair of the endogenous mutagen 8-oxoguanine in DNA. Nature. 2000;403(6772):859–866. doi: 10.1038/35002510. [DOI] [PubMed] [Google Scholar]
- 52.Fromme JC, Verdine GL. DNA lesion recognition by the bacterial repair enzyme MutM. J Biol Chem. 2003;278(51):51543–51548. doi: 10.1074/jbc.M307768200. [DOI] [PubMed] [Google Scholar]
- 53.Duclos S, Aller P, Jaruga P, Dizdaroglu M, Wallace SS, Doublie S. Structural and biochemical studies of a plant formamidopyrimidine-DNA glycosylase reveal why eukaryotic Fpg glycosylases do not excise 8-oxoguanine. DNA Repair. 2012;11(9):714–725. doi: 10.1016/j.dnarep.2012.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Imamura K, Averill A, Wallace SS, Doublie S. Structural characterization of viral ortholog of human DNA glycosylase NEIL1 bound to thymine glycol or 5-hydroxyuracil-containing DNA. J Biol Chem. 2012;287(6):4288–4298. doi: 10.1074/jbc.M111.315309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zhu C, Lu L, Zhang J, Yue Z, Song J, Zong S, Liu M, Stovicek O, Gao YQ, Yi C. Tautomerization-dependent recognition and excision of oxidation damage in base-excision DNA repair. Proc Natl Acad Sci USA. 2016;113(28):7792–7797. doi: 10.1073/pnas.1604591113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lee AJ, Warshaw DM, Wallace SS. Insights into the glycosylase search for damage from single-molecule fluorescence microscopy. DNA Repair. 2014;20:23–31. doi: 10.1016/j.dnarep.2014.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Zharkov DO, Grollman AP. The DNA trackwalkers: principles of lesion search and recognition by DNA glycosylases. Mutat Res. 2005;577(1–2):24–54. doi: 10.1016/j.mrfmmm.2005.03.011. [DOI] [PubMed] [Google Scholar]
- 58.Lee AJ, Wallace SS. Visualizing the Search for Radiation-damaged DNA Bases in Real Time. Radiat Phys Chem Oxf Engl. 2016;1993(128):126–133. doi: 10.1016/j.radphyschem.2016.05.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bennett SE, Sanderson RJ, Mosbaugh DW. Processivity of Escherichia coli and rat liver mitochondrial uracil-DNA glycosylase is affected by NaCl concentration. Biochemistry. 1995;34(18):6109–6119. doi: 10.1021/bi00018a014. [DOI] [PubMed] [Google Scholar]
- 60.Dowd DR, Lloyd RS. Biological significance of facilitated diffusion in protein-DNA interactions. Applications to T4 endonuclease V-initiated DNA repair. J Biol Chem. 1990;265(6):3424–3431. [PubMed] [Google Scholar]
- 61.Gruskin EA, Lloyd RS. Molecular analysis of plasmid DNA repair within ultraviolet-irradiated Escherichia coli. I. T4 endonuclease V-initiated excision repair. J Biol Chem. 1988;263(25):12728–12737. [PubMed] [Google Scholar]
- 62.Higley M, Lloyd RS. Processivity of uracil DNA glycosylase. Mutat Res. 1993;294(2):109–116. doi: 10.1016/0921-8777(93)90019-d. [DOI] [PubMed] [Google Scholar]
- 63.Mechetin GV, Zharkov DO. Mechanism of translocation of uracil-DNA glycosylase from Escherichia coli between distributed lesions. Biochem Biophys Res Commun. 2011;414(2):425–430. doi: 10.1016/j.bbrc.2011.09.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Porecha RH, Stivers JT. Uracil DNA glycosylase uses DNA hopping and short-range sliding to trap extrahelical uracils. Proc Natl Acad Sci USA. 2008;105(31):10791–10796. doi: 10.1073/pnas.0801612105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Purmal AA, Lampman GW, Pourmal EI, Melamede RJ, Wallace SS, Kow YW. Uracil DNA N-glycosylase distributively interacts with duplex polynucleotides containing repeating units of either TGGCCAAGCU or TGGCCAAGCTTGGCCAAGCU. J Biol Chem. 1994;269(35):22046–22053. [PubMed] [Google Scholar]
- 66.Sidorenko VS, Mechetin GV, Nevinsky GA, Zharkov DO. Correlated cleavage of single- and double-stranded substrates by uracil-DNA glycosylase. FEBS Lett. 2008;582(3):410–414. doi: 10.1016/j.febslet.2008.01.002. [DOI] [PubMed] [Google Scholar]
- 67.Blainey PC, van Oijen AM, Banerjee A, Verdine GL, Xie XS. A base-excision DNA-repair protein finds intrahelical lesion bases by fast sliding in contact with DNA. Proc Natl Acad Sci USA. 2006;103(15):5752–5757. doi: 10.1073/pnas.0509723103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Blainey PC, Luo G, Kou SC, Mangel WF, Verdine GL, Bagchi B, Xie XS. Nonspecifically bound proteins spin while diffusing along DNA. Nat Struct Mol Biol. 2009;16(12):1224–1229. doi: 10.1038/nsmb.1716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Dunn AR, Kad NM, Nelson SR, Warshaw DM, Wallace SS. Single Qdot-labeled glycosylase molecules use a wedge amino acid to probe for lesions while scanning along DNA. Nucleic Acids Res. 2011;39(17):7487–7498. doi: 10.1093/nar/gkr459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Nelson SR, Dunn AR, Kathe SD, Warshaw DM, Wallace SS. Two glycosylase families diffusively scan DNA using a wedge residue to probe for and identify oxidatively damaged bases. Proc Natl Acad Sci USA. 2014;111(20):E2091–E2099. doi: 10.1073/pnas.1400386111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Banerjee A, Santos WL, Verdine GL. Structure of a DNA glycosylase searching for lesions. Science. 2006;311(5764):1153–1157. doi: 10.1126/science.1120288. [DOI] [PubMed] [Google Scholar]
- 72.Fromme JC, Verdine GL. Structure of a trapped endonuclease III-DNA covalent intermediate. EMBO J. 2003;22(13):3461–3471. doi: 10.1093/emboj/cdg311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Qi Y, Nam K, Spong MC, Banerjee A, Sung RJ, Zhang M, Karplus M, Verdine GL. Strandwise translocation of a DNA glycosylase on undamaged DNA. Proc Natl Acad Sci USA. 2012;109(4):1086–1091. doi: 10.1073/pnas.1111237108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kuznetsov NA, Bergonzo C, Campbell AJ, Li H, Mechetin GV, de los Santos C, Grollman AP, Fedorova OS, Zharkov DO, Simmerling C. Active destabilization of base pairs by a DNA glycosylase wedge initiates damage recognition. Nucleic Acids Res. 2015;43(1):272–281. doi: 10.1093/nar/gku1300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Kuznetsov NA, Kladova OA, Kuznetsova AA, Ishchenko AA, Saparbaev MK, Zharkov DO, Fedorova OS. Conformational Dynamics of DNA Repair by Escherichia coli Endonuclease III. J Biol Chem. 2015;290(23):14338–14349. doi: 10.1074/jbc.M114.621128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Koval VV, Kuznetsov NA, Ishchenko AA, Saparbaev MK, Fedorova OS. Real-time studies of conformational dynamics of the repair enzyme E. coli formamidopyrimidine-DNA glycosylase and its DNA complexes during catalytic cycle. Mutat Res. 2010;685(1–2):3–10. doi: 10.1016/j.mrfmmm.2009.08.018. [DOI] [PubMed] [Google Scholar]
- 77.Koval VV, Kuznetsov NA, Zharkov DO, Ishchenko AA, Douglas KT, Nevinsky GA, Fedorova OS. Pre-steady-state kinetics shows differences in processing of various DNA lesions by Escherichia coli formamidopyrimidine-DNA glycosylase. Nucleic Acids Res. 2004;32(3):926–935. doi: 10.1093/nar/gkh237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kuznetsov NA, Koval VV, Zharkov DO, Vorobjev YN, Nevinsky GA, Douglas KT, Fedorova OS. Pre-steady-state kinetic study of substrate specificity of Escherichia coli formamidopyrimidine – DNA glycosylase. Biochemistry. 2007;46(2):424–435. doi: 10.1021/bi060787r. [DOI] [PubMed] [Google Scholar]
- 79.Kuznetsov NA, Vorobjev YN, Krasnoperov LN, Fedorova OS. Thermodynamics of the multi-stage DNA lesion recognition and repair by formamidopyrimidine-DNA glycosylase using pyrrolocytosine fluorescence – stopped-flow pre-steady-state kinetics. Nucleic Acids Res. 2012;40(15):7384–7392. doi: 10.1093/nar/gks423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kuznetsov NA, Zharkov DO, Koval VV, Buckle M, Fedorova OS. Reversible chemical step and rate-limiting enzyme regeneration in the reaction catalyzed by formamidopyrimidine-DNA glycosylase. Biochemistry. 2009;48(48):11335–11343. doi: 10.1021/bi901100b. [DOI] [PubMed] [Google Scholar]
- 81.Prakash A, Eckenroth BE, Averill AM, Imamura K, Wallace SS, Doublie S. Structural investigation of a viral ortholog of human NEIL2/3 DNA glycosylases. DNA Repair. 2013;12(12):1062–1071. doi: 10.1016/j.dnarep.2013.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Liu M, Imamura K, Averill AM, Wallace SS, Doublie S. Structural characterization of a mouse ortholog of human NEIL3 with a marked preference for single-stranded DNA. Structure. 2013;21(2):247–256. doi: 10.1016/j.str.2012.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Prasad R, Beard WA, Batra VK, Liu Y, Shock DD, Wilson SH. A review of recent experiments on step-to-step “hand-off” of the DNA intermediates in mammalian base excision repair pathways. Mol Biol. 2011;45(4):586–600. [PMC free article] [PubMed] [Google Scholar]
- 84.Wilson SH, Kunkel TA. Passing the baton in base excision repair. Nat Struct Biol. 2000;7(3):176–178. doi: 10.1038/73260. [DOI] [PubMed] [Google Scholar]
- 85.Ghodke H, Wang H, Hsieh CL, Woldemeskel S, Watkins SC, Rapic-Otrin V, Van Houten B. Single-molecule analysis reveals human UV-damaged DNA-binding protein (UV-DDB) dimerizes on DNA via multiple kinetic intermediates. Proc Natl Acad Sci USA. 2014;111(18):E1862–E1871. doi: 10.1073/pnas.1323856111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Finkelstein IJ, Visnapuu ML, Greene EC. Single-molecule imaging reveals mechanisms of protein disruption by a DNA translocase. Nature. 2010;468(7326):983–987. doi: 10.1038/nature09561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Gorman J, Plys AJ, Visnapuu ML, Alani E, Greene EC. Visualizing one-dimensional diffusion of eukaryotic DNA repair factors along a chromatin lattice. Nat Struct Mol Biol. 2010;17(8):932–938. doi: 10.1038/nsmb.1858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Visnapuu ML, Greene EC. Single-molecule imaging of DNA curtains reveals intrinsic energy landscapes for nucleosome deposition. Nat Struct Mol Biol. 2009;16(10):1056–1062. doi: 10.1038/nsmb.1655. [DOI] [PMC free article] [PubMed] [Google Scholar]