

Immature wild orangutans cyclically vary mothers’ milk consumption over 8 or more years, weaning later than other mammals.
Keywords: non-human primates, nursing, weaning, orangutan, Seasonality, life history, barium
Abstract
Nursing behavior is notoriously difficult to study in arboreal primates, particularly when offspring suckle inconspicuously in nests. Orangutans have the most prolonged nursing period of any mammal, with the cessation of suckling (weaning) estimated to occur at 6 to 8 years of age in the wild. Milk consumption is hypothesized to be relatively constant over this period, but direct evidence is limited. We previously demonstrated that trace element analysis of bioavailable elements from milk, such as barium, provides accurate estimates of early-life diet transitions and developmental stress when coupled with growth lines in the teeth of humans and nonhuman primates. We provide the first detailed nursing histories of wild, unprovisioned orangutans (Pongo abelii and Pongo pygmaeus) using chemical and histological analyses. Laser ablation inductively coupled plasma mass spectrometry was used to determine barium distributions across the teeth of four wild-shot individuals aged from postnatal biological rhythms. Barium levels rose during the first year of life in all individuals and began to decline shortly after, consistent with behavioral observations of intensive nursing followed by solid food supplementation. Subsequent barium levels show large sustained fluctuations on an approximately annual basis. These patterns appear to be due to cycles of varying milk consumption, continuing until death in an 8.8-year-old Sumatran individual. A female Bornean orangutan ceased suckling at 8.1 years of age. These individuals exceed the maximum weaning age reported for any nonhuman primate. Orangutan nursing may reflect cycles of infant demand that relate to fluctuating resource availability.
INTRODUCTION
Life history, or the way in which organisms allocate energy to growth, reproduction, and maintenance, is fundamental to a species’ biology and behavior. Lactation is a significant energetic investment for mammalian mothers, whose reproductive success is also affected by environmental conditions (1). Orangutans live in highly variable and unpredictable environments and have lower daily energy expenditures when compared with humans and other mammals (2, 3). This strategy of energy minimization has been hypothesized to facilitate the avoidance of starvation during episodes of fruit scarcity in these large-bodied primates, resulting in the slowest life histories of any nonhuman primate (2, 4, 5). However, little is known about weaning age in wild orangutans, nor other aspects of life history, such as life span. Experienced field biologists have concluded that observational approaches do not permit comprehensive quantification of suckling duration or milk intake in wild orangutans and other arboreal primates (6). Infants and older immature individuals may suckle inconspicuously, as well as while nesting with mothers at night, both difficult to detect in an arboreal environment (7). Moreover, continuous long-term field studies encounter obstacles while documenting a process that may last for seven or more years, and they typically rely on age estimations because exact birth dates are rarely known (4, 8, 9).
During lactation, mammals produce calcium-rich milk by drawing on skeletal reserves. Approximately 99% of the calcium in the body is found in bone, and the remaining amount in blood and organ tissues is continuously adjusted to maintain homeostasis. Certain low-level nonessential elements, such as barium and lead, often follow the movement of calcium in the body because they share certain transport pathways. These elements are preferentially concentrated in mothers’ milk, although barium passes through a partial filtering system (biopurification), making it proportionately less common than calcium. We have recently detailed an approach to document nursing in primates that integrates elemental mapping of barium concentrations and developmental analysis of teeth (10). Barium is an accurate indicator of milk provisioning because of the shared chemical properties with calcium, and it is more readily absorbed from maternal milk than from other dietary items (10, 11). For example, barium sulfate is used as a contrast agent for x-ray imaging because very little is absorbed by the gastrointestinal tract (12).
Nursing infants rapidly incorporate barium from milk into their skeletal tissues, including bones and teeth (10), as is the case for calcium and lead (13, 14). Thus, barium passes from one generation to the next, because stores originally incorporated into skeletal tissues during the mother’s development are mobilized for her infant. Minor contributions may also come from dietary sources, air, and water; however, most of the barium that is not incorporated into skeletal tissues is excreted within 1 to 2 weeks (12). Macaque mothers on a uniform diet show differences in barium levels in their milk (10), representing additional evidence that barium concentrations are primarily a function of somatic stores rather than dietary sources of barium. Therefore, elevated barium values in the teeth of nursing infants and immature individuals reflect the ingestion of maternal milk rather than nonmilk foods consumed directly or by their mothers while nursing.
Our method complements dietary studies of wild primates that produce temporally restricted measures of isotopes or trace elements from samples of hair or feces [reviewed by Tsutaya and Yoneda (15)]. Trace element analyses of teeth can be anchored to incremental growth lines to assign ages to specific values, yielding a precise longitudinal perspective that begins before birth and may continue until death (10, 16). Here, we show that accurate nursing histories and weaning ages can be determined from primate dentitions in zoological museums that were collected many decades ago. This approach is especially timely for critically endangered taxa, such as Sumatran and Bornean orangutans, Pongo abelii and Pongo pygmaeus, whose slow rate of growth and reproduction is a significant liability in a region that is being rapidly deforested.
RESULTS
Histological and elemental maps of the molars of four immature orangutans reveal an increase in barium values throughout the first year of life (Figs. 1 and 2). After the first year, the four immature orangutans in our study show episodic decreases in barium concentrations on an approximately yearly basis (Figs. 1, 3, and 4 and fig. S1). Cyclical patterns of barium levels persist into the eighth year of life in a Bornean female and into the ninth year of life in a Sumatran male and a second Sumatran individual of unknown sex. One Bornean female shows a sustained decrease in barium concentrations below baseline levels around 8.1 years of age, suggesting complete cessation of suckling before her death at 8.4 years of age (Fig. 3). Our oldest individual, an 8.8-year-old Sumatran individual of unknown sex, shows a broad elevated barium band for several months before its death (Fig. 4). Thus, at this age, it does not appear to have ceased suckling.
Fig. 1. Temporal map and barium distribution in the first molar of an immature wild Bornean orangutan.
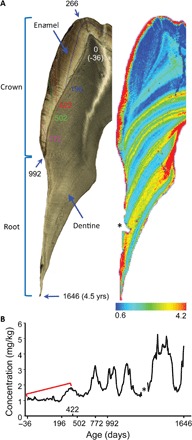
(A) Light microscopy image (left) of the mesiolingual cusp of the developing upper first molar (MCZ 5290) contrasted with an elemental map of the same section (right) showing the distribution of barium in the crown and root. Numbers on the microscopic image represent the age in days. High barium values in the outer enamel (red fringe) represent postdepositional modification of the subsurface enamel. Note that a small section of the subsurface root dentine was lost during preparation for elemental mapping [indicated by “*” here and in (B)]. (B) Calcium-normalized barium concentrations quantified in the dentine just below the enamel/root surface from the beginning of calcification (dentine horn tip below the enamel cusp) until death at 4.5 years of age (root tip). The approximate age of barium incorporation during dentine mineralization was determined from concurrently forming accentuated lines mapped with light microscopy (A). These ages are presented on a nonlinear scale that relates to the changing rate of extension during molar crown formation. The red bracket denotes a pattern of increasing barium incorporation due to milk consumption during the first year of life. Approximately annual marked decreases in barium/calcium begin around 400 days and continue until death, likely reflecting cycles of increased solid food consumption and reduced milk intake.
Fig. 2. Dentine barium patterns across the first molars of three immature wild orangutans.
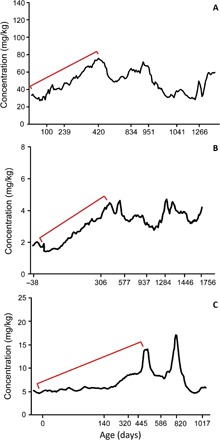
Calcium-normalized barium concentrations quantified from the beginning of first molar calcification (dentine horn tip) until the end of crown formation or the end of available root dentine in three immature wild orangutans. Ages are presented on a nonlinear scale that relates to the changing rate of extension during molar crown formation. Red brackets denote an increasing pattern of barium incorporation due to milk consumption during the first 1 to 1.5 years of life. (A) Bornean individual (ZSM 1981/48), (B) Sumatran individual (ZSM 1981/246), and (C) Sumatran individual (ZMB 83508). Although barium concentrations were markedly different between animals, values were consistent among teeth from the same individual (see Figs. 3 and 4 and fig. S1). Interindividual variation is consistent with variation in barium breast milk from six modern human subjects (39).
Fig. 3. Barium distribution across three molars from an 8.4-year-old female Bornean orangutan (ZSM 1981/48).
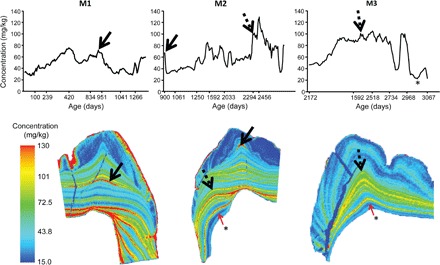
Calcium-normalized barium concentrations quantified from the beginning of calcification (near the dentine horn tip) in the first molar (M1) until the cessation of third molar (M3) crown formation at death. Ages are presented on a nonlinear scale that relates to the changing rate of extension during molar crown formation. Absolute barium values found here are greater than for the other individuals in this study, as is the case for strontium, although calcium and other trace element values are comparable to other individuals. Solid arrows show the barium pattern used to register the concurrently forming M1 and M2, and dashed arrows show the M2 to M3 registry, which allowed the age at death to be determined from counts and measurements of incremental features. Complete cessation of suckling appears to have occurred just after a prolonged enrichment of barium concentration that ended at 8.1 years, which is apparent as a deep blue band in the last-formed dentine of the M2 and M3 (indicated by “*”). The M3 concentration plot shows this event, whereas the M2 plot does not because of the missing root tip.
Fig. 4. Barium distribution across three molars from an 8.8-year-old Sumatran orangutan (ZMB 83508).
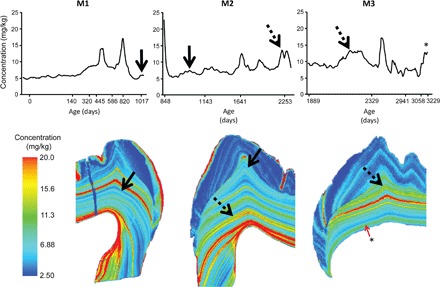
Calcium-normalized barium concentrations quantified from the beginning of calcification in the first molar (M1) until the cessation of third molar (M3) crown formation at death. Ages are presented on a nonlinear scale that relates to the changing rate of extension during molar crown formation. Solid arrows show the barium pattern used to register the concurrently forming M1 and M2, and dashed arrows show the M2 to M3 registry, which allowed the age at death to be determined from counts and measurements of incremental features. A prolonged period of barium enrichment occurred just before death at 8.8 years, which is apparent as a green band in the last-formed dentine of the M3 and as elevated values in the M3 barium concentration plot (indicated by “*”). The histological map of this individual is given in fig. S2.
DISCUSSION
Nursing patterns during the first year of life
All four orangutans show a gradual increase in barium values from birth through their first year of life. Consistent with our findings, behavioral observations of wild Bornean orangutans report that they rely almost exclusively on mothers’ milk in their first year; solid food supplementation only becomes an appreciable component of their diet between 12 and 18 months of age (6, 9). A similar pattern of increasing barium concentration was found in the first molars of rhesus macaques in their first 3 months of life (10). The initially low milk volume in the neonatal period increases during exclusive milk feeding as a function of infant growth, increased gastric capacity, and expanded behavioral repertoire (17). After 3 months, the barium levels in macaque teeth decreased, coincident with the consumption of solid foods. Thus, the initial pattern of barium concentration slowly increasing, peaking, and declining in wild orangutans and captive macaques reflects the inverted U-shaped curve of primate lactation (18).
Nursing patterns after the first year of life
We found that barium values decreased between 12 and 18 months of age in the four orangutans and then began to cycle on an approximately annual basis. This decreasing barium signature in teeth corresponds with the transition from a predominant milk diet to one characterized by increasing intake of solid foods. In contrast to seasonally breeding primates, such as rhesus macaques, orangutan females nurse their offspring over multiple years, enduring extreme and unpredictable inter- and intra-annual fluctuations in fruit availability (Fig. 5) (5, 9, 19–22). Cyclical episodes of decreased barium concentrations in immature orangutans likely reflect elevated solid food consumption during periods of high fruit availability. Confirmation of this hypothesis will require behavioral data from individuals whose barium concentrations may be quantified concurrently. This is a particular challenge because most museum specimens, including those in the current study, were collected long before primatological field sites were established in Indonesia and Malaysia (23). Periods of increased fruit availability not only alleviate infant demand of maternal resources but also afford females the opportunity to replenish energetic reserves to sustain lactation and/or transition to their subsequent reproduction.
Fig. 5. Fruit availability (above) and average energetic intake (below) of four lactating wild Bornean orangutan females over 7 years.
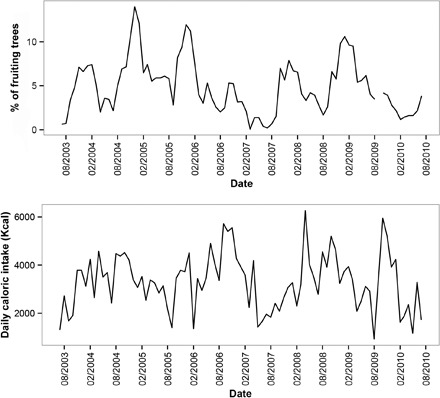
Data are from the Tuanan Orangutan Research Area, Central Kalimantan, Indonesia [detailed by Vogel et al. (22, 51)]. Top: The percentage of fruiting trees derives from 1868 marked trees in monthly monitored phenology plots, 98% of which are species consumed by orangutans. Bottom: Mean daily caloric intake for four adult mother orangutans with nursing offspring (ranging from 0 to 6 years in age) collected from 2003 to 2010. Data are based on 12,669 hours from 1100 full-day focal animal follows.
We interpret sustained cyclical increases in barium after the first year of life as periods of volumetric increases in milk consumption by infants when fruit or other preferred foods were limited. Immature orangutans may struggle to recognize or to process certain food items and will beg or attempt to scrounge maternal foods while nursing and possibly beyond the weaning process (6). Physiological studies of captive orangutans indicate that they experience an extreme form of low-energy throughput (2), and wild adult Bornean individuals are known to catabolize fat reserves and muscle tissue during periods of nutrient insufficiency (5, 24, 25). These physiological responses are hypothesized to be due to the extreme ecological unpredictability of Indonesian forests, which may be a particular challenge for immature individuals with limited fat reserves, muscle tissue, and foraging ability. Thus, the prolonged cyclical increases in barium concentration in our individuals’ dentitions are likely due to intensified reliance on their mothers’ milk—a rich source of bioavailable barium. This demand likely relaxes during periods of high fruit availability.
More abbreviated increases in barium concentrations (discrete spikes in barium graphs and thin elevated bands in elemental maps) may denote periods of physiological stress where trace elements in the infant’s own skeletal stores are released into the bloodstream and incorporated into mineralizing teeth. Moderate to severe illnesses and periods of significant weight loss are correlated with rapid increases in barium concentrations in the teeth of recently weaned captive macaques (26). These short-lived increases in barium are distinct from the more gradual pattern seen during nursing. Indications of physiological stress are consistent with developmental disruptions in the dentitions of several of our individuals (27). Comparative studies of external defects (enamel hypoplasias) report higher frequencies in orangutans than in other great apes (28). Wild orangutans appear to experience early life stress routinely, at least during the decades when most zoological museums acquired specimens from Indonesian expeditions. The presence of developmental defects alone is insufficient to distinguish among causes, such as nutritional deficiencies, immune challenges, or social stress (29, 30). Although data on captive primates and human children with associated medical records have provided insight into the etiology of these defects [for example, (26, 29, 31)], these conclusions are more difficult for wild primates (30). Characterizations of structural defects and elemental incorporation coupled with behavioral and ecological data are needed to determine why immature orangutans show high proportions of developmental defects in their teeth.
Barium and primate diets
Available evidence on barium and other nonessential elements that are typically found at low physiological levels indicates limited absorption by humans and other primates from nonmilk dietary sources. The primary naturally occurring forms of barium are not water-soluble; thus, barium levels in drinking water are negligible unless contaminated from industrial waste sites (12). Few studies have measured barium in food, and none could be found specific to plant-based foods or insects in the diets of Indonesian orangutans. Evidence of barium levels across different parts of plants (for example, leaf, root, fruit, and flower) is mixed; there is some evidence that mature leaves contain more barium than young leaves (32–34). Even in cases where barium levels are high in certain plants or plant parts, barium is not absorbed in amounts that would be toxic to consumers (12, 32, 35).
Barium absorption has been linked to the efficiency of calcium absorption rather than concentration in the food item (36). Our previous study of macaques revealed that the transition to nonmilk foods was associated with a sharp decline in tooth barium levels (10), likely due to the low bioavailability of barium in most plant-based foods. Therefore, the marked cyclical increases in barium concentration in the teeth of orangutans are unlikely to result from their consumption of nonmilk foods. The idea that these increases are due to barium in maternal diets is even less likely because gastrointestinal absorption of barium is age-dependent. For example, human adults absorb one-third to one-half of the barium absorbed by infants and immature individuals (37). Barium is discriminated against in favor of calcium (38), causing the barium/calcium ratio to decrease during the mother’s low-level absorption from her diet, then during excretion into maternal milk, and finally during absorption from maternal milk by the offspring. Thus, changes in maternal dietary barium intake have less influence on the offspring’s tooth barium than on the mother’s serum levels.
The magnitude and cyclicity of fluctuations in tooth barium concentrations are unlikely to be caused by elemental variation in maternal milk during lactation. Following the initial phase of milk production, barium remains relatively constant throughout the rest of lactation, as is the case for lead (13). Barium values in milk from six human mothers collected over 5 months did not differ significantly within mothers (39). Samples of milk from two macaque mothers show only fairly small changes over 4 months (10). The infants of these two mothers do not show perfectly parallel trends in milk barium and tooth barium; the latter is highest during the period when they are exclusively suckling, peaking before breast milk barium values. We interpret this as a volumetric effect. It is also unlikely that the barium content of maternal milk changes appreciably during periods when orangutan mothers rely on reserves, releasing energy stored in body fat and protein (24). Muscles, fat, and connective tissue contain only 1 to 2% of the somatic stores of barium found in humans (12). An appreciable release of barium from skeletal reserves has only been documented during acute illness or starvation in mammals, and only for a short duration in these cases (26). Orangutan mothers would seriously compromise their health by mobilizing skeletal reserves in this extreme way during recurring periods of decreased energy intake. Although the amount of variation in dietary barium intake by orangutan mothers is unknown, the available evidence suggests that it is the quantity of ingested milk that varies throughout nursing, leading to fluctuations in the barium incorporated in teeth.
Orangutan lactation and weaning
Female primates that lactate over multiple seasons or years are hypothesized to provide a relatively stable amount of milk over this period (9); thus, dependent offspring must supplement their increasing energetic needs by increasing their solid food intake. Our results suggest that there may be intra-annual fluctuations in milk intake after the first 1 to 1.5 years of an individual’s life. Sustained cyclical increases in barium in the teeth of dependent offspring are likely due to increased milk consumption, as is the case during the first year of life. We hypothesize that this occurs during periods of fruit scarcity, which coincides with periods of limited energy intake for lactating females (Fig. 5) and their dependent offspring. Immature individuals may be forced to consume a greater volume of milk to meet their energetic needs during cyclical periods of fruit scarcity.
Our results are consistent with behavioral evidence that wild orangutans continue nursing for a remarkably long period when compared to other mammals, perhaps only ceasing when offspring are able to achieve and sustain complete nutritional independence. Although primate infants may continue to suckle during the early stages of their mother’s subsequent pregnancy (reviewed by Smith et al. 40), orangutan infants do not appear to show the same degree of weaning conflict seen in other primates, such as rhesus macaques (9, 41). The cessation of milk transfer in Bornean orangutans has been reported to occur at 5.75 and 7.5 years in two individuals of known birthdates (9). The cessation of suckling documented here for a Bornean female at 8.1 years of age is the oldest weaning age reported for a nonhuman primate. Another individual who died at 8.8 years of age was on track to have weaned at an even older age; it appears to have been nursing for the last few months before its death. Although it is possible that this individual experienced skeletal remineralization resulting in elevated values in the last few months of life, these stress events tend to show a more discrete and narrow “spike” in elemental intensity (26), which is unlike the pattern observed here. This leads us to conclude that this individual continued to suckle until it was shot. Unfortunately, there are no reliable observational data on the cessation of suckling in known-aged wild Sumatran orangutans for comparison. One of the main difficulties in documenting this in wild arboreal primates, such as orangutans, is the fact that suckling is often inconspicuous, and it is particularly difficult to assess when individuals are nesting with their mothers at night (7).
Biologists have long endeavored to understand reproductive physiology in large-bodied and long-lived mammalian species. Some have suggested that the limited and unpredictable resource availability of Southeast Asian forests may have contributed to orangutans’ prolonged life history, including their late ages at weaning, reproductive maturation, and female growth cessation (8, 42). Others have elaborated that physiological and behavioral adaptations to subsist on low levels of energy may be the proximate cause of life history prolongation, which is ultimately driven by limited and/or inconsistent resource availability (2). These theories are consistent with evidence from baboon infants in marginal mountainous environments, who show increased time on the nipple, less time independently foraging, and later weaning ages than those in East African savannas (43). Increasingly marked seasons and unstable habitats have been invoked as a significant selective pressure that drove the evolution of early humans and great apes (44, 45), yet precise methods that document environmental variation during the lifetime of fossil primates have been lacking. Future applications may compare fine-scaled barium concentrations in primates from environments with varied degrees of seasonality and cyclical resource availability, which may then be extended to interpret patterns in the fossil record (10). Although further research is needed to test hypotheses for the evolution of primate life histories, the approach documented here provides a novel view into the nursing behavior and weaning process of one of the most poorly understood and highly endangered great apes.
MATERIALS AND METHODS
Study design
The objective of the study was to measure barium distributions in teeth from four immature orangutans to determine their nursing behavior in the wild, which is poorly understood. Wild orangutan dental samples are rare; thus, the sample size was based on material from a previous study of orangutan dental development (27).
Experimental design
Histological sections of the molars of four immature, wild-shot orangutans were prepared according to standard protocols, and accentuated lines in the enamel were spatiotemporally mapped from incremental features formed after the neonatal (birth) line (27). In brief, counts and measurements of daily cross-striations were made along a prism track from the neonatal line to each successive accentuated line. Ages were assigned to accentuated lines in the lateral enamel by adding time represented by previous long-period growth lines to the age at cusp formation (Fig. 1, fig. S2, and table S1).
Histological analyses of immature dentitions can yield ages of death within ~2% of the actual age of the individual (46–48). In some instances, these estimates can be exact or within 1 day of the individual’s age at death (31, 49). It is likely that the estimate of the youngest individual in the current study (MCZ 5290) is the most accurate because the age was determined from matching two teeth, whereas the older individuals required registry of accentuated lines combined with the barium banding pattern across several teeth. The timing of accentuated lines and the age at death of these older individuals were estimated to have an error of plus or minus 1 month (27).
We used a New Wave Research NWR193 laser ablation system connected to an Agilent Technologies 8800s inductively coupled plasma mass spectrometer (ICP-MS) by Tygon tubing. Details of our analytical methods have been previously published (10, 26). A 35-μm-diameter laser beam was rastered along the sample surface in a straight line at a speed of 70 μm s−1, producing data points that correspond to a pixel size of approximately 35 × 35 μm. Barium concentrations were calculated from concentrations determined using the certified standard National Institute of Standards and Technology 612 glass and 43Ca and 138Ba isotopes. There are no matrix-matched standards for laser ablation ICP-MS analysis of teeth. We used certified glass standards and calcium levels as an internal standard, which limited absolute quantification of metals. Elemental maps were processed using the interactive R Shiny application “shinyImaging.” Details of the image processing methods have been previously published (50), and the application can be accessed at http://labs.icahn.mssm.edu/lautenberglab/. Color scales were applied using the linear blue-red Lookup Table. Image backgrounds were converted to white (absent from the color intensity scale) to clarify sample boundaries from the substrate. Elemental maps were overlaid on light microscopy images using the Georeferencer tool in QGIS (version 2.8.2-Wien). Reference points were placed along the outer edge and along the enamel-dentine junction of the tooth, and the thin-plate spline transformation was applied.
Supplementary Material
Acknowledgments
C. Borries, L. Cook, Z. Machanda, and M. van Noordwijk provided helpful comments on this research. M. Ade, J. Chupasko, and O. Roehrer-Ertl provided access to living great ape material from the Humboldt Museum (Berlin), State Anthropological Collection (Munich), and the Harvard University Museum of Comparative Zoology. We thank two anonymous reviewers and the editor for comments that significantly improved the manuscript. Funding: This research was funded by NSF BCS 0213994 (T.M.S.); NSF BCS 0643122, U.S. Agency for International Development APS-497-11-000001, LSB Leakey Foundation, and Rutgers University, Center for Human Evolutionary Studies (E.R.V.); NIEHS DP2ES025453 and R00ES019597 (M.A.); Stony Brook University; the Max Planck Society; and Harvard University. Author contributions: T.M.S. and M.A. designed the study; T.M.S., C.A., E.R.V., and M.A. collected the data and analyzed the results; T.M.S., C.A., and E.R.V. prepared the figures; and all authors contributed to the writing of the paper. Competing interests: The authors declare that they have no competing interests. Data and materials availability: All data needed to evaluate the conclusions in the paper are present in the paper and/or the Supplementary Materials. Additional data related to this paper may be requested from the authors.
SUPPLEMENTARY MATERIALS
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/3/5/e1601517/DC1
table S1. Immature wild orangutan individuals examined in the current study.
fig. S1. Barium distribution across two molars from an 8.5-year-old male Sumatran orangutan (ZSM 1981/246).
fig. S2. Accentuated line ages in the first to third molars of an 8.8-year-old Sumatran orangutan (ZMB 83508).
REFERENCES AND NOTES
- 1.Lee P. C., Majluf P., Gordon I. J., Growth, weaning and maternal investment from a comparative perspective. J. Zool. 225, 99–114 (1991). [Google Scholar]
- 2.Pontzer H., Raichlen D. A., Shumaker R. W., Ocobock C., Wich S. A., Metabolic adaptation for low energy throughput in orangutans. Proc. Natl. Acad. Sci. U.S.A. 107, 14048–14052 (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pontzer H., Brown M. H., Raichlen D. A., Dunsworth H., Hare B., Walker K., Luke A., Dugas L. R., Durazo-Arvizu R., Schoeller D., Plange-Rhule J., Bovet P., Forrester T. E., Lambert E. V., Thompson M. E., Shumaker R. W., Ross S. R., Metabolic acceleration and the evolution of human brain size and life history. Nature 533, 390–392 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wich S. A., Utami-Atmoko S. S., Setia T. M., Rijksen H. D., Schürmann C., van Hooff J. A. R. A. M., van Schaik C. P., Life history of wild Sumatran orangutans (Pongo abelii). J. Hum. Evol. 47, 385–398 (2004). [DOI] [PubMed] [Google Scholar]
- 5.C. D. Knott, M. E. Thompson, S. A. Wich, in Orangutans: Geographic Variation in Behavioral Ecology and Conservation, S. A. Wich, S. S. Utami Atmoko, T. M. Setia, C. P. van Schaik, Eds. (Oxford Univ. Press, 2009), pp. 171–188. [Google Scholar]
- 6.M. A. van Noordwijk, S. E. B. Sauren, N. A. Ahbam, H. C. Morrogh-Bernard, S. S. Utami Atmoko, C. P. van Schaik, in Orangutans: Geographic Variation in Behavioral Ecology and Conservation, S. A. Wich, S. S. Utami Atmoko, T. M. Setia, C. P. van Schaik, Eds. (Oxford Univ. Press, 2009), pp. 189–203. [Google Scholar]
- 7.Borries C., Lu A., Ossi-Lupo K., Larney E., Koenig A., The meaning of weaning in wild Phayre’s leaf monkeys: Last nipple contact, survival, and independence. Am. J. Phys. Anthropol. 154, 291–301 (2014). [DOI] [PubMed] [Google Scholar]
- 8.van Noordwijk M. A., van Schaik C. P., Development of ecological competence in Sumatran orangutans. Am. J. Phys. Anthropol. 127, 79–94 (2005). [DOI] [PubMed] [Google Scholar]
- 9.van Noordwijk M. A., Willems E. P., Utami Atmoko S. S., Kuzawa C. W., van Schaik C. P., Multi-year lactation and its consequences in Bornean orangutans (Pongo pygmaeus wurmbii). Behav. Ecol. Sociobiol. 67, 805–814 (2013). [Google Scholar]
- 10.Austin C., Smith T. M., Bradman A., Hinde K., Joannes-Boyau R., Bishop D., Hare D. J., Doble P., Eskenazi B., Arora M., Barium distributions in teeth reveal early-life dietary transitions in primates. Nature 498, 216–219 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Uenishi K., Ezawa I., Kajimoto M., Tsuchiya F., Calcium absorption from milk, fish (pond smelt, sardine) and vegetables (komatsuna-green, Jew’s marrow, saltwort) in Japanese young women. Nippon Eiyo Shokuryo Gakkaishi 51, 259–266 (1998). [Google Scholar]
- 12.D. Moffett, C. Smith, Y.-W. Stevens, L. Ingerman, S. Swarts, L. Chappell, A Toxicological Profile for Barium and Barium Compound (U.S. Department of Health and Human Services, Agency for Toxic Substances and Disease Registry, 2007). [Google Scholar]
- 13.Gulson B. L., Mahaffey K. R., Jameson C. W., Mizon K. J., Korsch M. J., Cameron M. A., Eisman J. A., Mobilization of lead from the skeleton during the postnatal period is larger than during pregnancy. J. Lab. Clin. Med. 131, 324–329 (1998). [DOI] [PubMed] [Google Scholar]
- 14.Kovacs C. S., Calcium and bone metabolism during pregnancy and lactation. J. Mammary Gland Biol. Neoplasia 10, 105–118 (2005). [DOI] [PubMed] [Google Scholar]
- 15.Tsutaya T., Yoneda M., Reconstruction of breastfeeding and weaning practices using stable isotope and trace element analyses: A review. Am. J. Phys. Anthropol. 156, 2–21 (2015). [DOI] [PubMed] [Google Scholar]
- 16.Humphrey L. T., Dean M. C., Jeffries T. E., Penn M., Unlocking evidence of early diet from tooth enamel. Proc. Natl. Acad. Sci. U.S.A. 105, 6834–6839 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.K. Hinde, in Building Babies, K. B. H. Clancy, K. Hinde, J. N. Rutherford, Eds. (Springer, 2013), pp. 187–207. [Google Scholar]
- 18.Neville M. C., Allen J. C., Archer P. C., Casey C. E., Seacat J., Keller R. P., Lutes V., Rasbach J., Neifert M., Studies in human lactation: Milk volume and nutrient composition during weaning and lactogenesis. Am. J. Clin. Nutr. 54, 81–92 (1991). [DOI] [PubMed] [Google Scholar]
- 19.Harrison M. E., Zweifel N., Husson S. J., Cheyne S. M., D’Arcy L. J., Harsanto F. A., Morrogh-Bernard H. C., Purwanto, Rahmatd, Santiano P., Vogel E. R., Wich S. A., van Noordwijk M. A., Disparity in onset timing and frequency of flowering and fruiting events in two bornean peat-swamp forests. Biotropica 48, 188–197 (2016). [Google Scholar]
- 20.Knott C. D., Changes in orangutan caloric intake, energy balance, and ketones in response to fluctuating fruit availability. Int J. Primatol. 19, 1061–1079 (1998). [Google Scholar]
- 21.Curran L. M., Leighton M., Vertebrate responses to spatiotemporal variation in seed production of mast-fruiting Dipterocarpaceae. Ecol. Monogr. 70, 101–128 (2000). [Google Scholar]
- 22.Vogel E. R., Alavi S. E., Utami-Atmoko S. S., van Noordwijk M. A., Bransford T. D., Erb W., Zulfa A., Sulistyo F., Farida W. R., Rothman J. M., Nutritional ecology of wild Bornean orangutans (Pongo pygmaeus wurmbii) in a peat swamp habitat: Effects of age, sex, and season. Am. J. Primatol. 79, 1–20 (2017). [DOI] [PubMed] [Google Scholar]
- 23.Schaller G. B., The orang-utan in Sarawak. Zoologica 46, 73–82 (1961). [Google Scholar]
- 24.Vogel E. R., Knott C. D., Crowley B. E., Blakely M. D., Larsen M. D., Dominy N. J., Bornean orangutans on the brink of protein bankruptcy. Biol. Lett. 8, 333–336 (2012). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Harrison M. E., Morrogh-Bernard H. C., Chivers D. J., Orangutan energetics and the influence of fruit availability in the nonmasting peat-swamp forest of Sabangau, Indonesian Borneo. Int. J. Primatol. 31, 585–607 (2010). [Google Scholar]
- 26.Austin C., Smith T. M., Farahani R. M. Z., Hinde K., Carter E. A., Lee J., Lay P. A., Kennedy B. J., Sarrafpour B., Wright R. J., Wright R. O., Arora M., Uncovering system-specific stress signatures in primate teeth with multimodal imaging. Sci. Rep. 6, 18802 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Smith T. M., Dental development in living and fossil orangutans. J. Hum. Evol. 94, 92–105 (2016). [DOI] [PubMed] [Google Scholar]
- 28.Guatelli-Steinberg D., Ferrell R. J., Spence J., Linear enamel hypoplasia as an indicator of physiological stress in great apes: Reviewing the evidence in light of enamel growth variation. Am. J. Phys. Anthropol. 148, 191–204 (2012). [DOI] [PubMed] [Google Scholar]
- 29.S. Hillson, Tooth Development in Human Evolution and Bioarchaeology (Cambridge Univ. Press, 2014). [Google Scholar]
- 30.Smith T. M., Boesch C., Developmental defects in the teeth of three wild chimpanzees from the Taï forest. Am. J. Phys. Anthropol. 157, 556–570 (2015). [DOI] [PubMed] [Google Scholar]
- 31.Schwartz G. T., Reid D. J., Dean M. C., Zihlman A. L., A faithful record of stressful life events recorded in the dental developmental record of a juvenile gorilla. Int. J. Primatol. 27, 1201–1219 (2006). [Google Scholar]
- 32.W. O. Robinson, R. R. Whetstone, G. Edgington, in The occurrence of barium in soils and plants (Technical Bulletin no. 1013, United States Department of Agriculture, 1950).
- 33.Bowen H. J. M., Dymond J. A., Strontium and barium in plants and soils. Proc. R. Soc. B 144, 355–368 (1955). [Google Scholar]
- 34.McBride M. B., Shayler H. A., Spliethoff H. M., Mitchell R. G., Marquez-Bravo L. G., Ferenz G. S., Russell-Anelli J. M., Casey L., Bachman S., Concentrations of lead, cadmium and barium in urban garden-grown vegetables: The impact of soil variables. Environ. Pollut. 194, 254–261 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Goncalves A. M., Fernandes K. G., Ramos L. A., Cavalheiro É. T. G., Nóbrega J. A., Determination and fractionation of barium in Brazil nuts. J. Braz. Chem. Soc. 20, 760–769 (2009). [Google Scholar]
- 36.Elias R. W., Hirao Y., Patterson C. C., The circumvention of the natural biopurification of calcium along nutrient pathways by atmospheric inputs of industrial lead. Geochim. Cosmochim. Acta 46, 2561–2580 (1982). [Google Scholar]
- 37.ICRP, Age-dependent doses to members of the public from intake of radionuclides: Part 2, Ingestion dose coefficients, ICRP publication 67, Annals of ICRP 23 (3/4) (Pergamon Press, 1993). [PubMed] [Google Scholar]
- 38.Burton J. H., Price T. D., Middleton W. D., Correlation of bone Ba/Ca and Sr/Ca due to biological purification of calcium. J. Archaeol. Sci. 26, 609–616 (1999). [Google Scholar]
- 39.Anderson R. R., Longitudinal changes of trace elements in human milk during the first 5 months of lactation. Nutr. Res. 13, 499–510 (1993). [Google Scholar]
- 40.Smith T. M., Machanda Z., Bernard A. B., Donovan R. M., Papakyrikos A. M., Muller M. N., Wrangham R., First molar eruption, weaning, and life history in living wild chimpanzees. Proc. Natl. Acad. Sci. U.S.A. 110, 2787–2791 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Berman C. M., Rasmussen K. L. R., Suomi S. J., Reproductive consequences of maternal care patterns during estrus among free-ranging rhesus monkeys. Behav. Ecol. Sociobiol. 32, 391–399 (1993). [Google Scholar]
- 42.Leigh S. R., Shea B. T., Ontogeny and the evolution of adult body size dimorphism in apes. Am. J. Primatol. 36, 37–60 (1995). [DOI] [PubMed] [Google Scholar]
- 43.Lycett J. E., Henzi S. P., Barrett L., Maternal investment in mountain baboons and the hypothesis of reduced care. Behav. Ecol. Sociobiol. 42, 49–56 (1998). [Google Scholar]
- 44.Potts R., Environmental hypotheses of hominin evolution. Am. J. Phys. Anthropol. 41, 93–136 (1998). [DOI] [PubMed] [Google Scholar]
- 45.Potts R., Paleoenvironmental basis of cognitive evolution in great apes. Am. J. Primatol. 62, 209–228 (2004). [DOI] [PubMed] [Google Scholar]
- 46.Smith T. M., Reid D. J., Sirianni J. E., The accuracy of histological assessments of dental development and age at death. J. Anat. 208, 125–138 (2006). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Antoine D., Hillson S., Dean M. C., The developmental clock of dental enamel: A test for the periodicity of prism cross-striations in modern humans and an evaluation of the most likely sources of error in histological studies of this kind. J. Anat. 214, 45–55 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Smith T. M., Smith B. H., Reid D. J., Siedel H., Vigilant L., Hublin J.J., Boesch C., Dental development of the Taï Forest chimpanzees revisited. J. Hum. Evol. 58, 363–373 (2010). [DOI] [PubMed] [Google Scholar]
- 49.Smith T. M., Teeth and human life-history evolution. Annu. Rev. Anthropol. 42, 191–208 (2013). [Google Scholar]
- 50.Niedzwiecki M., Austin C., Remark R., Merad M., Gnjatic S., Estrada-Gutierrez G., Espejel-Nunez A., Borboa-Olivares H., Guzman-Huerta M., Wright R.J., Wright R.O., Arora M., A multimodal imaging workflow to visualize metal mixtures in the human placenta and explore colocalization with biological response markers. Metallomics 8, 444–452 (2016). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Vogel E. R., Harrison M. E., Zulfa A., Bransford T. D., Alavi S. E., Husson S., Morrogh-Bernard H., Santiano, Firtsman T., Utami-Atmoko S. S., van Noordwijk M. A., Farida W. R., Nutritional differences between two orangutan habitats: Implications for population density. PLOS ONE 10, e0138612 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material for this article is available at http://advances.sciencemag.org/cgi/content/full/3/5/e1601517/DC1
table S1. Immature wild orangutan individuals examined in the current study.
fig. S1. Barium distribution across two molars from an 8.5-year-old male Sumatran orangutan (ZSM 1981/246).
fig. S2. Accentuated line ages in the first to third molars of an 8.8-year-old Sumatran orangutan (ZMB 83508).