

Abstract
Cancer metastasis remains the primary cause of pain, suffering, and death in cancer patients, and even the most current therapeutic strategies have not been highly successful in preventing or inhibiting metastasis. In most patients with scirrhous gastric cancer (one of the most aggressive of diffuse‐type gastric cancer), recurrence occurs even after potentially curative resection, most frequently in the form of peritoneal metastasis. Given that the occurrence of diffuse‐type gastric cancers has been increasing, the development of new strategies to combat metastasis of this disease is critically important. Plasminogen activator inhibitor‐1 (PAI‐1) is a critical factor in cancer progression; thus, PAI‐1 RNAi may be an effective therapy against cancer metastasis. In the present study, we used an RNAi technique to reduce PAI‐1 expression in an in vivo model system for gastric cancer metastasis. Ex vivo plasmid transfection and adenovirus infection were tested as mechanisms to incorporate specific PAI‐1 RNAi vectors into human gastric carcinoma cells. Both approaches significantly decreased peritoneal tumor growth and the formation of bloody ascites in the mouse model, suggesting that this approach may provide a new, effective strategy for inhibiting cancer metastasis. (Cancer Sci 2012; 103: 228–232)
There are approximately 450 000 deaths from cancer each year in the US alone and estimates indicate that 90% of these deaths are the result of cancer metastatic disease.( 1 ) However, current treatments are not very successful in inducing remission. The incidence and mortality of overall gastric carcinoma in the US has been declining over the past 50 years, but the incidence of diffuse‐type gastric cancer has been increasing.( 2 ) Scirrhous gastric cancer is a diffuse‐type gastric cancer that is characterized by diffusely infiltrating carcinoma with frequent peritoneal dissemination. Patients with scirrhous gastric cancer have a poorer prognosis than patients with other types of gastric cancer.( 3 , 4 ) In most patients with scirrhous gastric cancer, the recurrence of cancer occurs even after potentially curative resection, most frequently in the form of peritoneal metastasis.( 4 ) Thus, with an increasing incidence of aggressive tumors, better therapeutic approaches are greatly needed.
Plasminogen activator inhibitor‐1 (PAI‐1) has been long considered as a cancer inhibitor.( 5 ) However, PAI‐1 is associated with a poor prognosis in a variety of cancers, including breast, skin, lung, colon, brain, ovarian, and stomach.( 6 ) A recent study has also reported that PAI‐1 is an integrator of cell signaling and migration.( 7 ) In animal models, high levels of PAI‐1 have been shown to be correlated with the ability of human melanoma cells to metastasize in the nude mouse,( 8 ) and a monoclonal antibody to PAI‐1 has been shown to suppress the metastatic potential of human HT1080‐P4 fibrosarcoma cells.( 9 ) However, overexpression of PAI‐1 in breast carcinoma MDA‐MB‐435 cells reduced migration and invasion, with an antibody that blocks PAI‐1 activity restoring migration and invasion.( 10 ) Antisense oligonucleotides to PAI‐1 have been reported to increase the invasion of HLE hepatocellular carcinoma cells.( 11 ) Recently, physiological concentrations of PAI‐1 were shown to promote tumor invasion and angiogenesis, whereas supraphysiological concentrations inhibited tumor vascularization.( 12 ) These reasons underlying these apparently conflicting results are not fully understood. Thus, the first aim of the present study was to elucidate whether cancer cell‐derived PAI‐1 is critical for tumor progression in vivo.
Using RNAi induces a sequence‐specific degradation of homologous mRNA, leading to post‐transcriptional gene silencing( 13 ) that can be at least a hundred‐fold more efficient than antisense RNA in mouse oocytes.( 14 ) This approach has been applied successfully to inhibit tumorigenicity( 15 ) and tumor progression.( 16 ) Knock‐out methods are very useful for research, but cannot be applied as yet for the therapy of solid cancers. Thus, we used knocked down host‐ and tumor cell‐derived PAI‐1 expression to investigate therapeutic potential. The second goal of the present study was to evaluate the potential of PAI‐1 RNAi as a treatment for cancer metastasis.
Materials and Methods
Cell culture. The human scirrhous gastric cancer cell line OCUM‐2M was established in our department from a resected primary tumor.( 17 ) An extensively peritoneal‐seeding cell line, namely OCUM‐2MD3, was established from the OCUM‐2M cell line using orthotopic tissue implantation in nude mice.( 18 ) The cell line was maintained in DMEM (Invitrogen, Frederick, MD, USA), supplemented with 10% heat‐inactivated bovine serum, 100 IU/mL penicillin, and 0.5 mM sodium pyruvate, at 37°C in a humidified atmosphere containing 5% carbon dioxide.
Plasmid RNAi construction and ex vivo transient transfection. We have described the plasmid vector‐mediated PAI‐1 RNAi construct and transfection conditions previously.( 19 ) Briefly, Hairpin siRNA template oligonucleotides for PAI‐1 were designed using an algorithm from Ambion (Austin, TX, USA) and synthesized by Invitrogen. The sense loop sequence is 5′‐GATCCCGTGACCGACATGTTCAGACATTCAAGAGATGTCTGAACATGTCGGTCATTTTTTGGAAA‐3′ and the antisense sequence is 5′‐AGCTTTTCCAAAAAATGACCGACATGTTCAGACATCTATTGAATGTCTGAACATGTCGGTCACGG‐3′. The annealed hairpin siRNA oligonucleotides were inserted into pSilencer™4.1‐CMV hygro vector (Ambion) according to the manufacturer’s instructions. The negative control consisted of an siRNA template sequence that lacks significant homology to the mouse, human, and rat genome databases. The vectors were cloned into Douglas Hanaham 5 (DH5)α‐competent cells. The inserted sequences were verified by DNA sequencing. The OCUM‐2MD3 cells were seeded in 100‐mm dishes at a density of 6 × 106 cells/dish and grown overnight to approximately 60% confluence prior to transfection. Cells were transfected with siPORT™XP‐1 transfection reagent (Ambion) according to the manufacturer’s instructions. Appropriate amounts of the siRNA expression plasmids were used (10 μg/100 mm dish).
Adenovirus RNAi construction. The siRNA template oligonucleotides for PAI‐1 were designed using an algorithm from Ambion and synthesized by Invitrogen. The sense strand was 5′‐TCGAGTGACCGACATGTTCAGACATTCAAGAGATGTCTGAACATGTCGGTCATTA‐3′ and the antisense strand is 5′‐CTAGTAATGACCGACATGTTCAGACATCTCTTGAATGTCTGAACATGTCGGTCAC‐3′. The annealed siRNA oligonucleotides were inserted into pSilencer adeno 1.0‐CMV Shuttle vector (Ambion). The linearized shuttle vector and the linearized Adenoviral LacZ Backbone plasmid (Ambion) were mixed and transfected in HEK‐293 cells using a calcium phosphate method according to the manufacturer’s instructions. All the viruses were propagated in a package containing 293 cells, purified twice via ultracentrifugation in a cesium chloride gradient and subjected to dialysis. The titer of the viruses (i.e. viral particles [vp]) was determined according to the absorbance at 260 nm (= optical density at 260 nm [OD260nm]).
Western blotting for in vitro and in vivo knock‐down efficiency. To evaluate in vitro knock‐down efficiency, OCUM‐2MD3 cells were plated in 24‐well culture plates at a density of 1 × 105 cells/well 1 day before infection and were subsequently infected at an MOI of 1000 (vp). Cells were collected, washed with PBS, and lysed in RIPA buffer 2 days after infection. Lysates were centrifuged at 13 500g for 15 min at 4°C. The protein concentration of each lysate was determined using the BCA protein assay (Pierce Biotechnology, Rockford, IL, USA). To evaluate in vivo knock‐down efficiency, 4‐week‐old female athymic mice were killed at neuropsy, blood was collected and the serum was separated. A 6.5‐μL aliquot of each serum sample was used for loading. Samples were resolved by SDS‐PAGE and transferred to nitrocellulose membrane. Membranes were blocked with 5% milk in Tris‐buffered saline Tween‐20 (TBST) at room temperature for 1 h. Membranes were incubated with antibodies against PAI‐1, GAPDH (Santa Cruz Biotechnology, Santa Cruz, CA, USA), or albumin (Bethyl Laboratory, Montgomery, TX, USA) at room temperature for 2 h or at 4°C overnight, after which they were incubated with an HRP‐conjugated secondary antibody for 1 h. Immunoreactivity was detected using chemiluminescent substrate (SuperSignal West Pico Chemiluminescent Substrate; Pierce Biotechnology). Subsequently, PAI‐1 expression levels were determined by densitometry analysis. Images were analyzed using ImageJ 1.44p (developed by Wayne Rasband, National Institutes of Health [NIH], Bethesda, MD, USA).
Animal experiments. Mice were maintained in microisolator cages in a specific pathogen‐free facility and experiments were conducted in accordance with NIH guidelines for the care and use of laboratory animals. To determine the effects of PAI‐1 knockdown ex vivo, OCUM‐2MD3 cells expressing PAI‐1 siRNA or control siRNA were inoculated i.p. into 4‐week‐old female athymic nude mice obtained from Charles River Laboratories (Raleigh, NC, USA). The PAI‐1 siRNA or control siRNA plasmids were transfected 1 day before inoculation. Briefly, cells were harvested from subconfluent culture and collected by centrifugation at 1500g for 5 min at room temperature, washed once and resuspended in serum‐free DMEM at a density of 1 × 107 cells/mL. Mice (n = 5 per group) were injected i.p. with OCUM‐2MD3 cells (2 × 106 cells in 200 μL) and observed for an additional 4 weeks. To examine the effects of PAI‐1 knockdown in vivo, mice (n = 10 per group) were injected i.p. with OCUM‐2MD3 cells (2 × 106 cells in 200 μL serum‐free DMEM). Adenovirus (2 × 1011 vp) with either PAI‐1 RNAi or control RNAi in 200 μL of 3% sucrose/PBS were injected i.p. twice a week. Mice were killed by CO2 4 weeks after the final injection. At necropsy, the extent of peritoneal metastasis was assessed macroscopically. Tumor volume was calculated using the equation a × b 2 × 0.5, where a and b are the largest and smallest tumor diameters, respectively. Each experiment was performed twice.
Immunohistochemistry and microvessel density scoring for angiogenesis. Paraffin‐embedded tumor sections (4 μm) were deparaffinated and rehydrated according standard procedures and washed with PBS solution. To block endogenous peroxidase activity, sections were put in 3% hydrogen peroxide in methanol for 20 min and then rinsed in PBS. Sections were blocked with an avidin/biotin block and rabbit serum. Immunostaining was performed using a goat Vectas stain ABC elite kit (Vector Laboratories, Burlingame, CA, USA) according to the manufacturer’s instructions, with the following modifications. Sections were incubated for 2 × 30 min with anti‐CD31 antibodies (Santa Cruz Biotechnology) diluted 1:750 in Dako Antibody Diluent (DakoCytomation, Carpinteria, CA, USA). Slides were counterstained with Mayer’s hematoxylin for 1 min and washed with water. Histologic slides were blind‐coded during assessment. Tissue sections were viewed at a magnification of ×100 (0.565 mm2 per field). Four fields per section were randomly chosen for analysis. Microvessel density (MVD) was calculated as the number of CD31‐positive objects/0.565 mm2 and the mean values of MVD in each group were calculated from seven tumor samples.
Statistical analysis. Unless indicated otherwise, data are presented as the mean ± SEM. Data were analyzed using Student’s t‐test and the chi‐square test. P < 0.05 was considered significant.
Results
Ex vivo plasmid vector‐mediated PAI‐1 RNAi effectively suppress peritoneal metastasis of OCUM‐2MD3 cells in nude mice. Initially, we examined the effects of PAI‐1 knockdown on tumor progression in vivo by using ex vivo transfection. Control and PAI‐1 RNAi expression vector‐transfected OCUM‐2MD3 cells were injected i.p. into mice. The peritoneal cavities were examined both macro‐ and microscopically 4 weeks after injection. Typical images of abdominal cavities in each group are shown in Figure 1(a). Peritoneal metastases were recognized as white nodules in the peritoneal cavity and confirmed by microscopic examination. The number of metastatic nodules in the PAI‐1‐knockdown group was less than that in the control group, but the difference failed to reach statistical significance (Fig. 1b). There was no difference between the two groups in the number of mice that developed metastatic nodules (Fig. 1e). However, the total volume of the metastatic nodules and the incidence of bloody ascites were significantly less in the PAI‐1‐knockdown group compared with the control group (2970.5 ± 713.1 vs 294.6 ± 214.2 mm3, respectively; 4/5 vs 0/5, respectively; Fig. 1c,d). These data are representative of two independent experiments.
Figure 1.
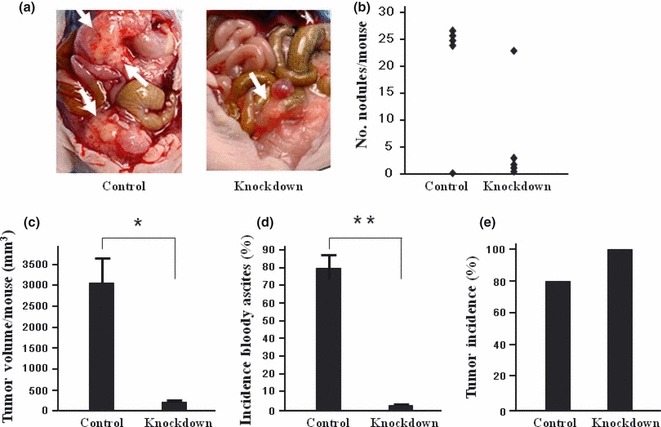
(a) Representative images of mice injected with either control (Control) or plasminogen activator inhibitor‐1 (PAI‐1) knockdown (Knockdown) siRNA expression plasmid vector‐transfected OCUM‐2MD3 cells. Arrows indicate tumor nodules. Bloody ascites was observed in mice carrying control vector‐transfected OCUM‐2MD3 cells. (b) There was a tendency for the number of metastatic tumors in mice (n = 5) injected with PAI‐1‐knockdown cells to be less that in mice injected with control cells, but the difference failed to reach statistical significance (P = 0.06). (c,d) There were significant decreases total metastatic tumor volume (c) and the formation of bloody ascites in mice injected with PAI‐1 knockdown cells compared with mice injected with control cells. (e) There was no significant difference in tumor incidence between the two groups. Data are shown as the mean ± SEM from experiments performed in duplicate. *P < 0.05, **P < 0.01.
In vivo suppression of angiogenesis by PAI‐1 RNAi. It has been reported that one of the crucial roles of PAI‐1 in cancer progression is in the development of new blood vessels that are critical for tumor nourishment and spread (i.e. angiogenesis). The MVD has been frequently used as a measure of angiogenesis. Therefore, in the present study we evaluated MVD in the tumor samples in vivo. Typical images of CD31 staining in each group are shown in Figure 2(a). The MVD was decreased in the PAI‐1‐knockdown group compared with the control group (11.6/0.565 vs 15.4/0.565 mm2, respectively; P < 0.05; Fig. 2b).
Figure 2.
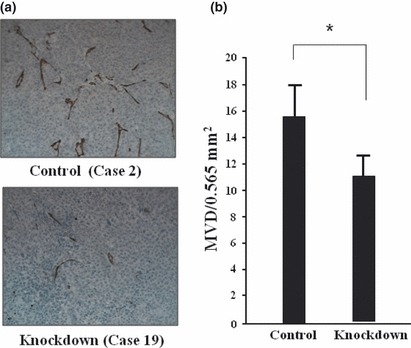
Microvessel density (MVD) of tumors in mice injected with either control (Control) or plasminogen activator inhibitor‐1 (PAI‐1) knockdown (Knockdown) siRNA expression plasmid vector‐transfected OCUM‐2MD3 cells. Tumors were stained with anti‐CD31 antibodies. (a) Representative images from each group. CD‐31‐positive objects were seen in tumor tissue (brown). (b) Mean ± SEM MVD values in each group calculated from seven tumor samples. *P < 0.05. The MVD was inhibited by PAI‐1 RNAi.
Adenovirus‐mediated PAI‐1 RNAi effectively suppresses peritoneal metastasis. Next we examined the potential clinical application of this by investigating the effects of the in vivo infection of RNAi. Three targets were tested and the most effective target was selected (data not shown). The PAI‐1‐knockdown efficiency was confirmed in vitro by using western blotting (Fig. 3a). Infection in vivo was confirmed by X‐gal staining of the peritoneal tumor (Fig. 3b). Typical images of abdominal cavities in each group are shown in Figure 4(a). The knockdown efficiency of PAI‐1 in vivo was confirmed by serum western blotting. Densitometry revealed a significant reduction in knockdown group, compared with control group (P < 0.05; Fig. 3d). The results are summarized in Figure 4. Although there was a tendency for the number of tumors in the knockdown group to be lower than that in the control group (7.81 ± 2.9 vs 2.41 ± 0.51, respectively), the difference failed to reach statistical significance (P = 0.09; Fig. 4b). Similarly, although there was a tendency for adenoviral knockdown to reduce tumor volume (mean [±SD] 1674 ± 1489 vs 616 ± 514 mm3 in the control and knockdown groups, respectively), the difference failed to reach statistical significance (P = 0.07; Fig. 4c). However, adenoviral knockdown significantly reduced the formation of bloody ascites formation (4/19 vs 0/20) and tumor incidence (12/19 vs 6/20; P < 0.05 for both; Fig. 4d,e). We did not see any effect on the hosts or the hosts’ tissues when we used adenovirus. Figures 1 and 4 show aggregate data of two independent experiments.
Figure 3.
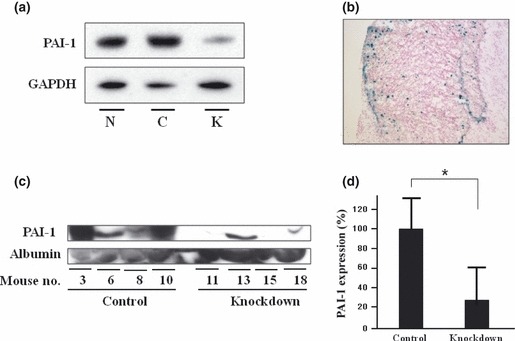
Adenovirus efficiency of knockdown and infection. (a) Plasminogen activator inhibitor‐1 (PAI‐1) protein expression was decreased by adenovirus‐mediated PAI‐1 siRNA in vitro. N, no adenovirus infection; C, control targeted knockdown; K, PAI‐1‐targeted knockdown. (b) Adenovirus infection was confirmed by X‐gal staining of peritoneal tumors in vivo. (Original magnification ×20.) (c) Serum was obtained 4 weeks after cancer cell injection and serum PAI‐1 and albumin were measured by western blotting. Adenovirus‐mediated PAI‐1 RNAi decreased PAI‐1 expression. (d) Densitomtric analysis of serum PAI‐1 expression. Images were analyzed using ImageJ 1.44p (developed by Wayne Rasband, National Institutes of Health). Densitometry revealed a significant reduction in PAI‐1 expression. Data are the mean ± SEM. *P < 0.05.
Figure 4.
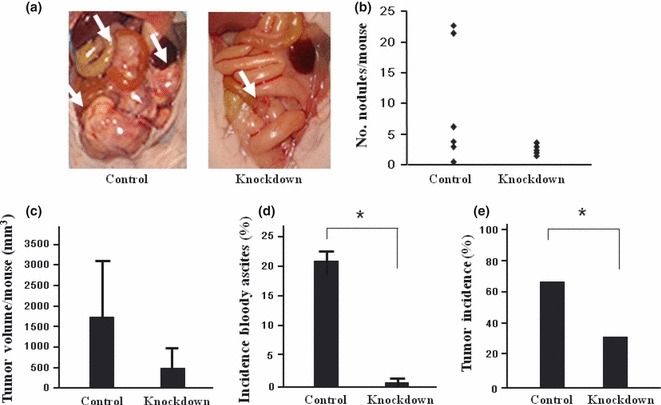
(a) Representative images of mice injected with adenovirus‐mediated siRNA. Control, mice injected with control siRNA; Knockdown, mice injected with plasminogen activator inhibitor‐1 (PAI‐1) knockdown siRNA. Arrows indicate peritoneal metastatic tumors. (b,c) There was a tendency for the number of metastatic tumors (b) and total metastatic tumor volume in mice (n = 10) injected with PAI‐1 RNAi adenovirus to be less than that in the control group, but the differences failed to reach statistical significance (P = 0.09 and 0.07, respectively). (d,e) There were significant reductions in the formation of bloody ascites (d) and tumor incidence (e) in the knockdown group compared with the control group. Data are the mean ± SEM. *P < 0.05.
Discussion
Plasminogen activator inhibitor‐1 has been implicated as a mediator of invasion and metastasis( 19 ) and presents an attractive potential target because 80% of patients with scirrhous gastric cancer show recurrence of cancer after tumor resection. This recurrence most frequently occurs in the form of peritoneal metastasis.( 4 ) However, there has been no report about the role/s of PAI‐1 in scirrhous gastric cancer and the therapeutic potential of PAI‐1 knockdown for peritoneal metastasis. In the present study, we demonstrated that PAI‐1 RNAi has potential as a therapy for peritoneal metastasis in as much as two different methods of PAI‐1 RNAi inhibited in vivo metastasis by human gastric cancer cells in a mouse model.
Host‐derived PAI‐1 is critical for in vivo angiogenesis and growth, and PAI‐1 produced by tumor cells did not overcome the absence of PAI‐1 in the host.( 12 ) Nevertheless our ex vivo model showed that PAI‐1 knockdown of cancer cells reduced tumor growth and angiogenesis in vivo, indicating that tumor cell‐derived PAI‐1 is also critical for tumor angiogenesis and growth. Previously, we reported that plasmid vector‐mediated PAI‐1 RNAi significantly decreased invasion ability and apoptosis.( 19 ) In the present study, PAI‐1 knockdown did not affect the adhesion of OCUM‐2MD3 cells (data not shown). There was an occasional mouse with a high number of nodules, as shown in Figure 1(b), perhaps due to loss of knockdown because there was no selection pressure in vivo. We also noticed that tumor progression in the control group in the adenovirus experiments seemed to be less than that in the ex vivo transfect experiments. We think this could be due to the cytotoxic effect of the adenovirus.
Systemic chemotherapy is often used for gastric cancer metastasis therapy. Although gastric cancer is a relatively chemosensitive disease with response rates of 30–40%, results of most classical chemotherapy have been unsatisfactory in terms of survival rate.( 21 ) When the combination of cisplatin and 5‐fluorouracil was used in Phase II studies, the response rate was 40%, but median survival was only 9 months.( 22 , 23 ) Moreover most chemotherapy does not achieve a better quality of life for patients. Thus, new approaches are needed. Many targets for cancer metastasis therapy have been identified and some have been used in the development of drugs that are used in both preclinical research and as clinical therapies.( 1 ) Although clinical studies of the MMP inhibitor marimastat,( 24 ) a monoclonal antibody against the integrin receptor,( 25 ) a monoclonal antibody against vascular endothelial growth factor‐A,( 26 ) and a Bcr‐Abl kinase inhibitor( 27 ) have been useful, there are still some concerns about improvements in survival for patients. The interest in PAI‐1 as a potential target for anticancer therapy stems from the diverse biological effects of PAI‐1 in different cell types.( 28 ) In patients with complete PAI‐1 deficiency, the clinical manifestations appear to be restricted to abnormal bleeding after trauma.( 29 ) Folkman( 30 ) reported that one of the critical roles of PAI‐1 in tumor progression is angiogenesis mediated through angiostatin, a circulating antiangiogenesis factor. Treatment of metastatic cancer with angiostatin has been reported to be feasible in a clinical study.( 31 ) These results and those of the present study suggest that it may be possible to use PAI‐1 RNAi for specific cases of anticancer therapy.
The present study shows for the first time that PAI‐1 RNAi in vivo effectively reduces gastric peritoneal metastasis. The RNAi of PAI‐1 effectively decreased tumor progression. Although a more effective vector or carrier for the delivery of RNAi remains to be discovered, these finding have important implications for the clinical use of PAI‐1 siRNA.
Disclosure Statement
The authors have no potential conflicts of interest.
Acknowledgments
This work was supported by the Intramural Research Program of the National Institute of Health (NIH) and National Institute of Environmental Health Sciences (NIEHS). The authors thank Dr Steven K. Akiyama (Laboratory of Molecular Carcinogenesis, NIEHS, Piedmont, NC, USA) for helpful discussions.
References
- 1. Sawyer TK. Cancer metastasis therapeutic targets and drug discovery: emerging small‐molecule protein kinase inhibitors. Expert Opin Investig Drugs 2004; 13: 1–19. [DOI] [PubMed] [Google Scholar]
- 2. Henson DE, Dittus C, Younes M, Nguyen H, Albores‐Saavedra J. Differential trends in the intestinal and diffuse types of gastric carcinoma in the United States, 1973–2000: increase in the signet ring cell type. Arch Pathol Lab Med 2004; 128: 765–70. [DOI] [PubMed] [Google Scholar]
- 3. Yashiro M, Chung YS, Nishimura S, Inoue T, Sowa M. Fibrosis in the peritoneum induced by scirrhous gastric cancer cells may act as “soil” for peritoneal dissemination. Cancer 1996; 77: 1668–75. [DOI] [PubMed] [Google Scholar]
- 4. Kaibara N, Maeta M, Hamazoe R, Inoue Y, Koga S. Hyperthermia adjunct to surgery in the treatment of scirrhous carcinoma of the stomach. Gan To Kagaku Ryoho 1988; 15: 1269–72 (in Japanese). [PubMed] [Google Scholar]
- 5. Soff GA, Sanderowitz J, Gately S et al. Expression of plasminogen activator inhibitor type 1 by human prostate carcinoma cells inhibits primary tumor growth, tumor‐associated angiogenesis, and metastasis to lung and liver in an athymic mouse model. J Clin Invest 1995; 96: 2593–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Andreasen PA, Egelund R, Petersen HH. The plasminogen activation system in tumor growth, invasion, and metastasis. Cell Mol Life Sci 2000; 57: 25–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Czekay RP, Wilkins‐Port CE, Higgins SP et al. PAI‐1: an integrator of cell signaling and migration. Int J Cell Biol 2011; 2011: 562481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Quax PH, van Muijen GN, Weening‐Verhoeff EJ et al. Metastatic behavior of human melanoma cell lines in nude mice correlates with urokinase‐type plasminogen activator, its type‐1 inhibitor, and urokinase‐mediated matrix degradation. J Cell Biol 1991; 115: 191–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Tsuchiya H, Sunayama C, Okada G, Matsuda E, Tomita K, Binder BR. Plasminogen activator inhibitor‐1 accelerates lung metastasis formation of human fibrosarcoma cells. Anticancer Res 1997; 17: 313–6. [PubMed] [Google Scholar]
- 10. Whitley BR, Palmieri D, Twerdi CD, Church FC. Expression of active plasminogen activator inhibitor‐1 reduces cell migration and invasion in breast and gynecological cancer cells. Exp Cell Res 2004; 296: 151–62. [DOI] [PubMed] [Google Scholar]
- 11. Morita Y, Hayashi Y, Kanamaru T et al. Inhibitory role of plasminogen activator inhibitor‐1 in invasion and proliferation of HLE hepatocellular carcinoma cells. Jpn J Cancer Res 1999; 90: 747–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Bajou K, Maillard C, Jost M et al. Host‐derived plasminogen activator inhibitor‐1 (PAI‐1) concentration is critical for in vivo tumoral angiogenesis and growth. Oncogene 2004; 23: 6986–90. [DOI] [PubMed] [Google Scholar]
- 13. Hammond SM, Bernstein E, Beach D, Hannon GJ. An RNA‐directed nuclease mediates post‐transcriptional gene silencing in Drosophila cells. Nature 2000; 404: 293–6. [DOI] [PubMed] [Google Scholar]
- 14. Stein P, Svoboda P, Schultz RM. Transgenic RNAi in mouse oocytes: a simple and fast approach to study gene function. Dev Biol 2003; 256: 187–93. [DOI] [PubMed] [Google Scholar]
- 15. Brummelkamp TR, Bernards R, Agami R. Stable suppression of tumorigenicity by virus‐mediated RNA interference. Cancer Cell 2002; 2: 243–7. [DOI] [PubMed] [Google Scholar]
- 16. Sumimoto H, Miyagishi M, Miyoshi H et al. Inhibition of growth and invasive ability of melanoma by inactivation of mutated BRAF with lentivirus‐mediated RNA interference. Oncogene 2004; 23: 6031–9. [DOI] [PubMed] [Google Scholar]
- 17. Yashiro M, Chung YS, Nishimura S, Inoue T, Sowa M. Establishment of two new scirrhous gastric cancer cell lines: analysis of factors associated with disseminated metastasis. Br J Cancer 1995; 72: 1200–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Yashiro M, Chung YS, Nishimura S, Inoue T, Sowa M. Peritoneal metastatic model for human scirrhous gastric carcinoma in nude mice. Clin Exp Metastasis 1996; 14: 43–54. [DOI] [PubMed] [Google Scholar]
- 19. Nishioka N, Matsuoka T, Yashiro M, Hirakawa K, Olden K, Roberts JD. Linoleic acid enhances angiogenesis through suppression of angiostatin induced by plasminogen activator inhibitor 1. Br J Cancer 2011; 105: 1750–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Bell WR. The fibrinolytic system in neoplasia. Semin Thromb Hemost 1996; 22: 459–78. [DOI] [PubMed] [Google Scholar]
- 21. Wohrer SS, Raderer M, Hejna M. Palliative chemotherapy for advanced gastric cancer. Ann Oncol 2004; 15: 1585–95. [DOI] [PubMed] [Google Scholar]
- 22. Lacave AJ, Baron FJ, Anton LM et al. Combination chemotherapy with cisplatin and 5‐fluorouracil 5‐day infusion in the therapy of advanced gastric cancer: a phase II trial. Ann Oncol 1991; 2: 751–4. [DOI] [PubMed] [Google Scholar]
- 23. Rougier P, Ducreux M, Mahjoubi M et al. Efficacy of combined 5‐fluorouracil and cisplatinum in advanced gastric carcinomas. A phase II trial with prognostic factor analysis. Eur J Cancer 1994; 30A: 1263–9. [DOI] [PubMed] [Google Scholar]
- 24. Bramhall SR, Rosemurgy A, Brown PD, Bowry C, Buckels JA. Marimastat as first‐line therapy for patients with unresectable pancreatic cancer: a randomized trial. J Clin Oncol 2001; 19: 3447–55. [DOI] [PubMed] [Google Scholar]
- 25. Gutheil JC, Campbell TN, Pierce PR et al. Targeted antiangiogenic therapy for cancer using vitaxin: a humanized monoclonal antibody to the integrin alphavbeta3. Clin Cancer Res 2000; 6: 3056–61. [PubMed] [Google Scholar]
- 26. Mulcahy MF, Benson AB 3rd. Bevacizumab in the treatment of colorectal cancer. Expert Opin Biol Ther 2005; 5: 997–1005. [DOI] [PubMed] [Google Scholar]
- 27. Ugurel S, Hildenbrand R, Zimpfer A et al. Lack of clinical efficacy of imatinib in metastatic melanoma. Br J Cancer 2005; 92: 1398–405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Durand MK, Bodker JS, Christensen A et al. Plasminogen activator inhibitor‐I and tumour growth, invasion, and metastasis. Thromb Haemost 2004; 91: 438–49. [DOI] [PubMed] [Google Scholar]
- 29. Fay WP, Parker AC, Condrey LR, Shapiro AD. Human plasminogen activator inhibitor‐1 (PAI‐1) deficiency: characterization of a large kindred with a null mutation in the PAI‐1 gene. Blood 1997; 90: 204–8. [PubMed] [Google Scholar]
- 30. Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med 1995; 1: 27–31. [DOI] [PubMed] [Google Scholar]
- 31. Beerepoot LV, Witteveen EO, Groenewegen G et al. Recombinant human angiostatin by twice‐daily subcutaneous injection in advanced cancer: a pharmacokinetic and long‐term safety study. Clin Cancer Res 2003; 9: 4025–33. [PubMed] [Google Scholar]