

Abstract
Febrile seizures are childhood convulsions resulting from an infection that leads to an inflammatory response and subsequent convulsions. Prenatal stress has been shown to heighten the progression and intensity of febrile seizures. Current medications are costly and have adverse effects associated with prolonged use. Quercetin flavonoid exhibits anti-inflammatory, anti-convulsant, and anti-stress effects. This study was aimed to investigate the therapeutic effect of quercetin in a prenatally stressed rat model of febrile seizures. We hypothesized that quercetin will alleviate the effects of prenatal stress in a febrile seizure rat model. On gestational day 13, Sprague-Dawley rat dams were subjected to restraint stress for 1 hour/d for 7 days. Febrile seizures were induced on postnatal day 14 on rat pups by intraperitoneally injecting lipopolysaccharide followed by kainic acid and quercetin on seizure onset. Hippocampal tissue was harvested to profile cytokine concentrations. Our results show that quercetin suppresses prenatal stress–induced pro-inflammatory marker (interleukin 1 beta) levels, subsequently attenuating febrile seizures. This shows that quercetin can be therapeutic for febrile seizures in prenatally stressed individuals.
Keywords: Febrile seizure, prenatal stress, inflammation, quercetin
Introduction
Febrile seizures are childhood convulsions triggered by fever that occur following common childhood infections, such as chickenpox and otitis media, resulting in pro-inflammatory cytokine release by peripheral immune cells.1 These cytokines include interleukin 1 beta (IL-1β), tumour necrosis factor α (TNF-α), and interleukin 6 (IL-6).2 Several hours after initiation of the inflammatory response, anti-inflammatory cytokines are released; these include the following: interleukin 1 receptor antagonist (IL-1ra) and interleukin 10 (IL-10).2 Pro-inflammatory cytokines then cross the blood-brain barrier (BBB) into the central nervous system (CNS).3 Entrance of pro-inflammatory cytokines into the brain activates glial cells, triggering an immune response in the CNS, thus more release of cytokines.2 The cytokines in the CNS elicit a cascade that results in fever and also leads to an imbalance in IL-1β and IL-1ra, one of the major role players in the pathogenesis of febrile seizures.4 The cytokines IL-1β and IL-1ra compete for binding at the interleukin 1 receptor type I (IL1RI) and IL1RI binds IL-β with higher affinity than IL-1ra, so a few IL-1β-occupied receptor complexes are required for cellular signaling.4 Therefore, high molar excess (100-fold to 1000-fold) of IL-1ra is essential to counteract IL-1β effects.4 Imbalance in the IL-1β/IL-1ra ratio alters the balance of the excitatory glutamate and inhibitory γ-aminobutyric acid neurotransmitters in the brain.4 The imbalance favours IL-1β in febrile seizures resulting in glutamate excitation, leading to neuronal hyperexcitability and thus seizures.4,5 Studies show that IL-10 has neuroprotective effects against pro-inflammatory cytokine-induced hyperexcitation of neurons seen in febrile seizures.6 Interleukin 10 exerts its anti-inflammatory activity and neuroprotection through suppressing release of pro-inflammatory cytokines.7 This suggests that IL-10 could possibly play a role in the attenuation of febrile seizures.6 Furthermore, early-life exposure to stress such as prenatal stress has been shown to enhance inflammatory cytokine release in the periphery and CNS and to promote febrile seizure progression.8,9 Antipyretics and anti-epileptic drugs are currently used to manage febrile seizures, but with prolonged use, these drugs can have adverse side effects including respiratory and CNS depression and gastric ulcers, among others.10,11 In addition, these drugs are also expensive making them inaccessible in low socioeconomic communities, so there is a need for an alternative treatment with fewer adverse effects and that is affordable. Quercetin is a flavonoid of great interest in research for its anti-inflammatory, anti-oxidant, anti-convulsant properties, among others.12 Therefore, the aim of this study was to investigate the potential therapeutic effect of quercetin in a prenatally stressed rat model of febrile seizures.
Methods and Materials
Prenatal handling
Animals
In total, 10 female and 5 male Sprague-Dawley rats were used for mating; these rats were acquired from the Biomedical Resource Centre of the University of KwaZulu-Natal where they were accommodated under standard laboratory conditions: room temperature of ±22°C, humidity of 70%, and a 12-hour light/dark cycle. Animals had free access to food and water ad libitum, and all experimental procedures performed were approved by the Animal Ethics Research Committee of the University of KwaZulu-Natal (ethics number AREC/057/015) in accordance with guidelines of the National Institutes of Health, USA.
Mating procedure
To prepare the female rats for mating, their oestrus cycle was synchronized 4 days before mating by putting 2 females in 1 cage. Studies show that ovarian cycles of female rats that live together become synchronized and this is mediated by pheromones.13,14 The oestrus cycle is a reproductive cycle of female rats lasting for 4 to 5 days and is observed by morphological modifications in the uterus, ovaries, and vagina.15 It is divided into 4 phases as follows: pro-oestrus, oestrus, metoestrus, and dioestrus.16 These phases are distinguished based on cell types observed in vaginal smears.16 The pro-oestrus and oestrus phases each last for 12 hours, whereas metoestrus takes 21 hours and dioestrus persists for 57 hours. The oestrus cycle was assessed daily by examination of a vaginal smear, and when a female was at pro-oestrus phase, a male was placed into the cage and a vaginal smear was obtained the following morning. Vaginal smears were collected as described by Marcondes et al,15 and the presence of sperm in the vaginal smear indicated that mating had taken place and that day was marked as gestational day 0 (GND0). The male rat was then taken out of the cage, and the pregnant dams were left undisturbed until gestational day 14 (GND14).
Prenatal stress protocol
On GND14, the pregnant dams were divided into 2 main groups: stressed group and non-stressed. The stress protocol was conducted from GND14 to GND20 as described by Cassim et al.17 Stress was introduced on GND14 because this is when gross neural structures of the CNS start specializing and also because the placental barrier becomes less active, thus the foetus is exposed to maternal stress hormones.18,19
Postnatal handling
Febrile seizure induction
Following birth, the pups remained with their dams until postnatal day (PND) 14. A total number of 24 pups (n = 3) were used in the study. Each group was divided into the following subgroups: non-stressed saline (NS-S), who received saline only; non-stressed febrile seizure (NS-FS), who received lipopolysaccharide (LPS) + kainic acid (KA); non-stressed quercetin, who received saline + quercetin (Q); and NS-FS treated with quercetin (NS-FSQ), who received LPS + KA + Q. Stressed saline (S-S), who received saline only; stressed febrile seizure (S-FS), who received LPS + KA; stressed quercetin, who received saline + quercetin; and S-FS treated with quercetin (S-FSQ), who received LPS + KA + Q. On PND 14, pups were separated from their dams and moved to the experimental room 1 hour before inducing febrile seizures for acclimatization. The control animals were injected intraperitoneally with 1-mL saline solution (0.9% NaCl; Adcock Ingram, Mid-rand, South Africa). The febrile seizure group was injected with a gram-negative bacterial endotoxin LPS (200 µg/kg, 0.2 mL, intraperitoneal; Sigma-Aldrich, St. Louis, MO, USA) and 2.5 hours later injected with a sublethal dose of the glutamate analogue KA (1.75 mg/kg, 0.2 mL, intraperitoneal; Sigma-Aldrich) to trigger convulsions.17 The treatment group was also injected with quercetin (10 mg/kg, 0.2 mL, intraperitoneal; Sigma-Aldrich) once there were noticeable convulsions in all the rats injected with LPS and KA.20 All the drugs were dissolved in 0.9% normal saline.
Seizure assessment
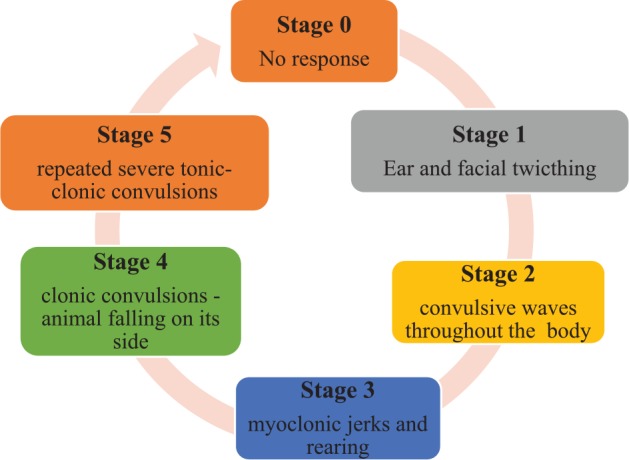
The assessment of seizure intensity is schematically represented in Figure 1.
Figure 1.
Decapitation
All the animals were moved to the autopsy room 1 hour post-seizure to be sacrificed. The animals were decapitated using a guillotine; the brains were harvested and dissected to collect hippocampal tissue. The tissue was then placed in 2-mL Eppendorf tubes, snap frozen in liquid nitrogen, and stored at −80°C bio-freezer for later analysis of cytokine levels.
Neurochemical analysis
The total protein content of the hippocampal samples was determined using an assay that was a reducing agent compatible as well as a detergent compatible (Bio-Rad Laboratories, Hercules, CA, USA). The samples were prepared for analysis by adding ice-cold phosphate-buffered saline (0.01 M, pH = 7.4) and volume added based on the sample weight. The samples were then placed in ice and homogenized to disrupt the cell wall and release the cell contents using an ultrasonic sonicator (Qsonica, Newtown, CT, USA). The homogenates were then centrifuged for 5 minutes at 5000g to obtain the supernate. The supernate was transferred to new Eppendorf tubes. The concentration of IL-1ra was measured using a commercially available Rat IL-1ra enzyme-linked immunosorbent assay kit (Elabscience Biotechnology, Wuhan, China). The concentration of IL-1β, TNF-α, IL-6, and IL-10 was measured using a commercially available Bio-Plex Multiplex Rat Cytokine immunoassay (Bio-Rad Laboratories).
Statistical analysis
The data were analysed with GraphPad Software Inc. La Jolla, CA, USA (version 5) software. Analysis of variance was performed on the rat hippocampal concentration of the following cytokines: IL-1β, TNF-α, IL-6, IL-1ra, and IL-10 with stress and febrile seizure as factors between rats. Significant main effects were followed by Bonferroni post hoc test. Differences were considered significant when P ⩽ .05.
Results
IL-1β concentration in hippocampal tissue
Interleukin 1 beta concentration was measured in the rat hippocampal tissue in the following groups: NS-S, NS-FS, NS-FSQ, S-S, S-FS, and S-FSQ. Exposure to febrile seizures alone in non-stressed rats significantly increased IL-1β concentration in comparison with their control (NS-S vs NS-FS; F3.16 = 18.40, P < .01). The treatment with quercetin resulted in increased IL-1β concentration in non-stressed rats subjected to febrile seizures (NS-FS vs NS-FSQ; F3.16 = 18.40, P < .001). Exposure to both prenatal stress and febrile seizures significantly increased IL-1β concentration in comparison with the non-stressed rats subjected to febrile seizures (NS-FS vs S-FS; F3.16 = 84.91, P < .05). Exposure to both prenatal stress and febrile seizures resulted in increased IL-1β concentration in comparison with rats subjected prenatal stress alone (S-S vs S-FS; F3.16 = 84.91, P < .0001). However, treatment with quercetin decreased IL-1β concentration in rats exposed to both prenatal stress and febrile seizures in comparison with untreated rats exposed to both prenatal stress and febrile seizures (S-FS vs S-FSQ; F3.16 = 18.40, P < .05) and also in comparison with treated rats exposed to febrile seizures alone (NS-FSQ vs S-FSQ; F3.16 = 18.40, P < .0001) (Figure 2).
Figure 2.

Graph showing the hippocampal concentration of IL-1β in the following groups: **(NS-S vs NS-FS; P < .01), ***(NS-FS vs NS-FSQ; P < .001), *(NS-FS vs S-FS; P < .05), ###(S-S vs S-FS; P < .001), #(S-FS vs S-FSQ; P < .05), and **(NS-FSQ vs S-FSQ; P < .01). IL-1β indicates interleukin 1 beta.
IL-1ra concentration in hippocampal tissue
Interleukin 1 receptor antagonist concentration was measured in the rat hippocampal tissue in the following groups: NS-S, NS-FS, NS-FSQ, S-S, S-FS, and S-FSQ. Exposure to febrile seizure resulted in increased IL-1ra concentration in the hippocampus of non-stressed rats subjected to febrile seizure compared with previously stressed rats subjected to febrile seizure (NS-FS vs S-FS; F3.16 = 19.74, P < .05). Treatment with quercetin resulted in increased IL-1ra concentration in the hippocampus of rats subjected to febrile seizure alone when compared with rats exposed to both prenatal stress and febrile seizures (NS-FSQ vs S-FSQ; F3.16 = 5.31, P < .001) (Figure 3).
Figure 3.

Graph showing the concentration of IL-1ra in the rat hippocampus in the following groups: *(NS-FS vs S-FS; P < .05) and ***(NS-FSQ vs S-FSQ; P < .001). IL-1ra indicates interleukin 1 receptor antagonist.
IL-1ra/IL-1β ratio in hippocampal tissue
We further assessed the IL-1ra/IL-1β ratio in the hippocampus in the following groups: NS-S, NS-FS, NS-FSQ, S-S, S-FS, and S-FSQ. Our findings show that the IL-1ra/IL-1β ratio was significantly lower in non-stressed rats with febrile seizure compared with the control (NS-S vs NS-FS; F3.16 = 8.62, P < .005) (Figure 4).
Figure 4.

Graph showing IL-1β/IL-1ra ratio in the rat hippocampus: *(NS-S vs N-FS; P < .05). IL-1β indicates interleukin 1 beta; IL-1ra, interleukin 1 receptor antagonist.
TNF-α concentration in hippocampal tissue
Tumour necrosis factor α concentration was measured in the rat hippocampal tissue in the following groups: NS-S, NS-FS, NS-FSQ, S-S, S-FS, and S-FSQ. Exposure to both prenatal stress and febrile seizures increased TNF-α concentration in the hippocampus compared with non-stressed rats with febrile seizures (NS-FS vs S-FS; F3.16 = 14.56, P < .001). Exposure to both febrile seizures and prenatal stress resulted in elevated levels of TNF-α concentration in the rat hippocampus when compared with the rats exposed to prenatal stress alone (S-S vs S-FS; F3.16 = 35.99, P < .001) (Figure 5).
Figure 5.

Graph showing concentration of TNF-α in the rat hippocampus in the following groups: ***(NS-FS vs S-FS; P < .001) and ###(S-S vs S-FS; P < .001). TNF-α indicates tumour necrosis factor α.
IL-6 concentration in hippocampal tissue
Interleukin 6 concentration was measured in the rat hippocampal tissue in the following groups: NS-S, NS-FS, NS-FSQ, S-S, S-FS, and S-FSQ. Exposure to both prenatal stress and febrile seizures increased IL-6 concentration in the hippocampus of prenatally stressed rats subjected to febrile seizure compared with non-stressed rats subjected to febrile seizure (NS-FS vs S-FS; F3.16 = 7.99, P < .001). Exposure to febrile seizure resulted in increased IL-6 concentration in the hippocampus of rats exposed to prenatal stress rats when compared with their saline control group (S-S vs S-FS; F3.16 = 30.58, P < .0001) (Figure 6).
Figure 6.

Graph showing the concentration of IL-6 in the rat hippocampus in the following groups: ***(NS-FS vs S-FS; P < .001) and ###(S-S vs S-FS; P < .0001). IL-6 indicated interleukin 6.
IL-10 concentration in hippocampal tissue
Interleukin 10 concentration was measured in the rat hippocampal tissue in the following groups: NS-S, NS-FS, NS-FSQ, S-S, S-FS, and S-FSQ. Exposure to febrile seizure decreased hippocampal concentration of IL-10 in non-stressed rats subjected to febrile seizure compared with the control (NS-S vs NS-FSQ; F3.16 = 9.28, P < .005). Exposure to febrile seizures in non-stressed rats resulted in decreased IL-10 concentration in rat hippocampal tissue when compared with rats exposed to both prenatal stress and febrile seizures (NS-FS vs S-FS; F3.16 = 9.28, P < .01). Treatment with quercetin resulted in increased IL-10 concentration in the hippocampus of non-stressed rats subjected to febrile seizure compared with untreated non-stressed rats subjected to febrile seizures (NS-FS vs NS-FSQ; F3.16 = 6.475, P < .005) (Figure 7).
Figure 7.

Graph showing IL-10 concentration in the rat hippocampus in the following groups: *(NS-S vs NS-FS; P < .005), **(NS-FS vs S-FS; P < .01), and *(NS-FS vs NS-FSQ; P < .05). IL-10 indicates interleukin 10.
Discussion
In this study, we investigated the effects of quercetin as a potential therapeutic agent for febrile seizures in animals that were previously exposed to prenatal stress by measuring concentrations of pro- and anti-inflammatory cytokines in the hippocampus of both stressed and non-stressed rats with febrile seizures.
Our findings show that exposure to febrile seizures alone resulted in increased levels of IL-1β concentration when compared with their saline control rats. These findings are in agreement with previous studies showing that there is a direct correlation between development of febrile seizures and high levels of IL-1β in the hippocampus, indicating that IL-1β is the principal initiator in the febrile response and neuronal hyperexcitability that leads to the onset of seizure activity.2,4,23 Therefore, this confirms that administration of LPS followed by KA successfully mimicked an infection, thus triggering an inflammatory response shown by high IL-1β levels observed. In addition, treatment with quercetin resulted in a further increase in IL-1β levels in rats exposed to febrile seizures alone compared with the untreated rats. Our findings contradict with previous studies which reported the anti-inflammatory properties of quercetin in various acute and chronic models of inflammation.24,25 Our findings show that quercetin failed to decrease the LPS-induced increase in IL-1β levels. The possible mechanism is that a Jarisch-Herxheimer reaction (JHR)–like response occurred after quercetin treatment; this JHR is triggered by endotoxins released into the systemic circulation by injured or dead bacteria.26–28 The rate that endotoxins are released is faster than the body can control it, so this triggers a sudden and exaggerated inflammatory response, which is characterized by an elevation of systemic pro-inflammatory cytokines; this phenomenon is usually seen in patients during treatment of infections such as syphilis, tuberculosis, and relapsing fever and it lasts for several hours, and after it has subsided, the patients start to feel better.27,28 Therefore, we postulate that post-treatment with quercetin, there was an exaggerated systemic release of IL-1β and other pro-inflammatory cytokines which then crossed the BBB to the CNS and activated glial cells to release more IL-1β. This exaggerated response would have probably subsided after several hours, but we were not able to observe that because we collected the hippocampal tissue an hour post-seizure.
Furthermore, our findings show that exposure to both prenatal stress and febrile seizures resulted in increased IL-1β in comparison with their non-stressed counterparts. This result is in agreement with studies showing that consequences of early exposure to maternal stress during gestation result in a maladaptive hypothalamic-pituitary-adrenal (HPA) axis in the offspring.29 It has been shown that the malfunction of the HPA axis due to prenatal stress results in an increase in corticotrophin-releasing factor (CRF) in the paraventricular nucleus (PVN) of the hypothalamus, thus eventually leading to higher than normal basal circulating corticosterone (cortisol in humans).30 This then leads to a higher than normal basal expression of pro-inflammatory cytokines in the CNS on an encounter with an infection.31
Literature also shows that the CNS regulates the immune system via the neuroendocrine pathways such as the HPA axis.32 Therefore, we postulate that the high IL-1β levels we observed in prenatally stressed rats with febrile seizures were due to activation of an already dysregulated HPA axis by IL-1β in the CNS which led to a greater release of corticosterone, which elicited more release of systemic pro-inflammatory cytokines and consequently those in hippocampus as well. However, treatment with quercetin resulted in decreased IL-1β levels in stressed rats compared with their non-stressed counterparts. This finding correlates with other findings showing that quercetin attenuates activation of the HPA axis due to stress by antagonizing CRF.25,33 These results suggest that quercetin may have decreased IL-1β concentration in the hippocampus by restoring the HPA axis, thus enabling it to properly regulate the secretion of CRF by the PVN; this then could have led to a decreased release of corticosterone and consequently decreasing IL-1β levels.
In addition, we assessed the levels of an IL-1β antagonist, IL-1ra, in rat hippocampal tissue. Our findings show that induction of febrile seizures significantly increased IL-1ra levels in non-stressed rats compared with the stressed rats. Interleukin 1 receptor antagonist is released in response to seizures to terminate IL-1β–mediated pro-inflammatory activity, but IL-1ra reaches its peak concentration several hours later than IL-1β.34,35 Therefore, because the inflammatory response was exaggerated in non-stressed rats compared with the stressed ones, this indicates that IL-1ra release had to increase as well to attenuate IL-1β activity. Our findings have also shown that the IL-1ra/IL-1β ratio was significantly lower in non-stressed rats with febrile seizures compared with their control, suggesting a high pro-inflammatory cytokine profile in the hippocampus. Therefore, our findings confirm that IL-1β binding is indeed favoured over IL-1ra during febrile seizures, showing the competitive binding for IL1RI between IL-1ra and IL-1β. Treatment with quercetin increased IL-1ra concentration in non-stressed rats with febrile seizures compared with stressed rats with febrile seizures. The possible mechanism is that quercetin possibly enhanced IL-1ra release, so this then suggests that quercetin not only antagonizes pro-inflammatory cytokines but also stimulates anti-inflammatory cytokines.
Moreover, our findings also show that TNF-α and IL-6 follow the same trend as IL-1β of being heightened in rats exposed to prenatal stress and febrile seizures compared with the non-stressed rats with febrile seizure. The findings of this study are in accordance with literature showing that stress induces hyperproduction of TNF-α and IL-6.31,36 These findings also show the supporting role played by TNF-α and IL-6 alongside IL-1β in the development of febrile seizures. We postulate that an increase in IL-1β levels triggered production and release of TNF-α and IL-6 via both the HPA axis pathway as well as the febrile seizure pathway. Treatment with quercetin did not show any significant effects on both TNF-α and IL-6; this is possibly due to that these pro-inflammatory cytokines only serve a supporting role during development of febrile seizures and the effect was most prominent in IL-1β because it is the key initiator during febrile seizures.
Finally, we assessed the levels of IL-10 an anti-inflammatory cytokine, and our findings show that febrile seizures decreased IL-10 levels in non-stressed rats with febrile seizures compared with their control and also compared with stressed rats with febrile seizures. Interleukin 10 is well known for relaying negative feedback signals that mitigate the activated immune system after an inflammatory trigger.34 Findings by Swarup et al37 show that following an infection, IL-10 levels are low, whereas pro-inflammatory cytokines are high. Literature also shows that a surge in IL-10 levels is observed 24 to 72 hours after the onset of seizures.34 Our findings are in agreement with the previous studies. The possible mechanism for these findings then would be that IL-10 had not yet surged to an efficacious concentration by the time tissue was collected, which was an hour post-seizure. However, treatment with quercetin increased IL-10 levels almost restoring them back to basal levels in non-stressed rats with febrile seizures. Quercetin has anti-inflammatory properties and acts by suppressing pro-inflammatory cytokines.12 The mechanism by which quercetin increased IL-10 levels is not quite clear, but we suspect that it possibly triggered an early release of IL-10 to limit the activity of pro-inflammatory cytokines; this therefore suggests that it had neuroprotective effects.
Conclusions
This study reports that exposure to prenatal stress increases the pro-inflammatory cytokine profile during febrile seizures. Treatment with quercetin decreases the pro-inflammatory cytokine profile in the presence of both prenatal stress and febrile seizures but increases the profile in the presence of febrile seizures alone, whereas it increases the anti-inflammatory cytokine profile in non-stressed groups with febrile seizure and offers neuroprotection. Therefore, from these results, we can conclude that quercetin offers therapeutic effect against febrile seizures only in the presence of prenatal stress. Based on these findings, we would conclude that quercetin is not efficacious in individuals with febrile seizure alone. Future studies could focus on applying techniques such as immunohistochemistry to gain more insights on the activity of quercetin and also assessing its activity at different time points.
Acknowledgments
The authors would like to thank the technical staff of the Biomedical Resource Centre of the University of KwaZulu-Natal for training and technical support. They also thank Dr K Moodley and Mr P Pillay from the Human Physiology Department, University Of KwaZulu-Natal for technical assistance as well as the University of KwaZulu-Natal Neuroscience group for support and constructive advice.
Footnotes
PEER REVIEW: Five peer reviewers contributed to the peer review report. Reviewers’ reports totalled 1989 words, excluding any confidential comments to the academic editor.
FUNDING: The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The author(s) received financial support from the University of KwaZulu-Natal for the publication of this article.
DECLARATION OF CONFLICTING INTERESTS: The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
NVPM and LQ conceived and designed the experiments, contributed to the writing of the manuscript, and jointly developed the structure and arguments for the paper. NVPM analysed the data and wrote the first draft of the manuscript. NVPM, LQ, and MVM agree with manuscript results and conclusions and made critical revisions and approved final version. All authors reviewed and approved the final manuscript.
Disclosures and Ethics
As a requirement of publication, author(s) have provided to the publisher signed confirmation of compliance with legal and ethical obligations including but not limited to the following: authorship and contributorship, conflicts of interest, privacy and confidentiality, and (where applicable) protection of human and animal research subjects. The authors have read and confirmed their agreement with the ICMJE authorship and conflict of interest criteria. The authors have also confirmed that this article is unique and not under consideration or published in any other publication, and that they have permission from rights holders to reproduce any copyrighted material. Any disclosures are made in this section. The external blind peer reviewers report no conflicts of interest.
REFERENCES
- 1.Shrestha D, Dhakal A, Shakya H, Shakya A, Shah S, Mehata S. Clinical characteristics of children with febrile seizure. J Nepal Health Res Counc. 2014;12:162–166. [PubMed] [Google Scholar]
- 2.Heida J, Moshe S, Pittman Q. The role of interleukin-1beta in febrile seizures. Brain Dev. 2009;31:388–393. doi: 10.1016/j.braindev.2008.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Salkeni M, Lynch J, Otamis-Price T, Banks W. Lipopolysaccharide impairs blood-brain barrier P-glycoprotein function in mice through prostaglandin- and nitric oxide-independent pathways. J Neuroimmune Pharmacol. 2009;4:276–282. doi: 10.1007/s11481-008-9138-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Rijkers K, Majoie H, Hoogland G, Kenis G, Baets MD, Vles J. The role of interleukin-1 in seizures and epilepsy: a critical review. Exp Neurol. 2009;216:258–271. doi: 10.1016/j.expneurol.2008.12.014. [DOI] [PubMed] [Google Scholar]
- 5.Balosso S, Maroso M, Sanchez-Alavez M, et al. A novel non-transcriptional pathway mediates the proconvulsive effects of interleukin-1β. Brain. 2008;131:3256–3265. doi: 10.1093/brain/awn271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ishizaki Y, Kira R, Fukuda M, Torisu H, Sakai Y, Sanfuji M. Interleukin-10 is associated with resistance to febrile seizures: genetic association and experimental animal studies. Epilepsia. 2009;50:761–767. doi: 10.1111/j.1528-1167.2008.01861.x. [DOI] [PubMed] [Google Scholar]
- 7.Sultani M, Stringer A, Bowen J, Gibson R. Anti-inflammatory cytokines: important immunoregulatory factors contributing to chemotherapy-induced gastrointestinal mucositis. Chemother Res Pract. 2012;2012:490804. doi: 10.1155/2012/490804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vanbesien-Mailliot C, Wolowczuk I, Mairesse J, et al. Prenatal stress has pro-inflammatory consequences on the immune system in adult rats. Psychoneuroendocrinology. 2007;32:114–124. doi: 10.1016/j.psyneuen.2006.11.005. [DOI] [PubMed] [Google Scholar]
- 9.Qulu L, Daniels W, Mabandla M. Exposure to prenatal stress enhances the development of seizures in young rats. Metab Brain Dis. 2012;27:399–404. doi: 10.1007/s11011-012-9300-3. [DOI] [PubMed] [Google Scholar]
- 10.Loddenkemper T, Goodkin H. Treatment of pediatric status epilepticus. Curr Treat Options Neurol. 2011;13:560–573. doi: 10.1007/s11940-011-0148-3. [DOI] [PubMed] [Google Scholar]
- 11.Hino H, Takahashi H, Suzuki Y, Tanaka J, Ishii E, Fukuda M. Anticonvulsive effect of paeoniflorin on experimental febrile seizures in immature rats: possible application for febrile seizures in children. PLoS ONE. 2012;7:e42920. doi: 10.1371/journal.pone.0042920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Comalada M, Camuesco D, Sierra S, et al. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through downregulation of the NF-kappaB pathway. Eur J Immunol. 2005;35:584–592. doi: 10.1002/eji.200425778. [DOI] [PubMed] [Google Scholar]
- 13.Schank J, McClintock M. A coupled-oscillator model of ovarian-cycle synchrony among female rats. J Theor Biol. 1992;157:317–362. doi: 10.1016/s0022-5193(05)80614-9. [DOI] [PubMed] [Google Scholar]
- 14.McClintock MK. Synchronizing ovarian and birth cycles by female pheromones. In: Müller-Schwarze D, Silverstein RM, editors. Chemical Signals in Vertebrates 3. Boston, MA: Springer; 1983. pp. 159–178. [Google Scholar]
- 15.Marcondes F, Bianchi F, Tanno A. Determination of the oestrus cycle phases of rats: some helpful considerations. Braz J Biol. 2002;62:609–614. doi: 10.1590/s1519-69842002000400008. [DOI] [PubMed] [Google Scholar]
- 16.Paccola C, Resende C, Stumpp T, Miraglia S, Cipriano I. The rat estrous cycle revisited: a quantitative and qualitative analysis. Anim Reprod. 2013;10:677–683. [Google Scholar]
- 17.Cassim S, Qulu L, Mabandla MV. Prenatal stress and early life febrile convulsions compromise hippocampal genes MeCP2/REST function in mid-adolescent life of Sprague-Dawley rats. Neurobiol Learn Mem. 2015;125:195–201. doi: 10.1016/j.nlm.2015.09.002. [DOI] [PubMed] [Google Scholar]
- 18.Patin V, Vincent A, Lordi B, Caston J. Does prenatal stress affect the motoric development of the rat pups? Brain Res Dev Brain Res. 2004;149:85–92. doi: 10.1016/j.devbrainres.2003.12.008. [DOI] [PubMed] [Google Scholar]
- 19.Mabandla M, Kellaway L, Daniels W, Russell V. Effect of exercise on dopamine neuron survival in prenatally stressed rats. Metab Brain Dis. 2009;24:525–539. doi: 10.1007/s11011-009-9161-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hu K, Li S, Xiao B, Bi F, Lu X, Wu X. Protective effects of quercetin against status epilepticus induced hippocampal neuronal injury in rats: involvement of X-linked inhibitor of apoptosis protein. Acta Neurol Belg. 2011;111:205–212. [PubMed] [Google Scholar]
- 21.Qulu L, Daniels W, Russell V, Mabandla M. Searsia chirindensis reverses the potentiating effect of prenatal stress on the development of febrile seizures and decreased plasma interleukin-1β levels. Neurosci Res. 2016;103:54–58. doi: 10.1016/j.neures.2015.08.004. [DOI] [PubMed] [Google Scholar]
- 22.Ojewole J. Anticonvulsant effect of Rhus chirindensis (Baker F.) (Anacardiaceae) stem-bark aqueous extract in mice. J Ethnopharmacol. 2008;117:130–135. doi: 10.1016/j.jep.2008.01.026. [DOI] [PubMed] [Google Scholar]
- 23.Bartfai T, Sanchez-Alavez M, Andell-Jonsson S, et al. Interleukin-1 system in CNS stress: seizures, fever, and neurotrauma. Ann N Y Acad Sci. 2007;1113:173–177. doi: 10.1196/annals.1391.022. [DOI] [PubMed] [Google Scholar]
- 24.Kanashiro A, Machado RR, Malvar DC, Aguiar FA, Souza GEP. Quercetin does not alter lipopolysaccharide-induced fever in rats. J Pharm Pharmacol. 2008;60:357–362. doi: 10.1211/jpp.60.3.0011. [DOI] [PubMed] [Google Scholar]
- 25.Li Y, Yao J, Han C, et al. Quercetin, inflammation and immunity. Nutrients. 2016;8:167. doi: 10.3390/nu8030167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Pound MW, May DB. Proposed mechanisms and preventative options of Jarisch-Herxheimer reactions. J Clin Pharm Ther. 2005;30:291–295. doi: 10.1111/j.1365-2710.2005.00631.x. [DOI] [PubMed] [Google Scholar]
- 27.Belum GR, Belum VR, Chaitanya Arudra SK, Reddy BSN. The Jarisch-Herxheimer reaction: revisited. Travel Med Infect Dis. 2013;11:231–237. doi: 10.1016/j.tmaid.2013.04.001. [DOI] [PubMed] [Google Scholar]
- 28.Rosen T, Rubin H, Ellner K, Tschen J, Cochran R. Vesicular Jarisch-Herxheimer reaction. Arch Dermatol. 1989;125:77–81. [PubMed] [Google Scholar]
- 29.Charil A, Laplante D, Vaillancourt C, King S. Prenatal stress and brain development. Brain Res Rev. 2010;65:56–79. doi: 10.1016/j.brainresrev.2010.06.002. [DOI] [PubMed] [Google Scholar]
- 30.Bosch OJ, Müsch W, Bredewold R, Slattery DA, Neumann ID. Prenatal stress increases HPA axis activity and impairs maternal care in lactating female off-spring: implications for postpartum mood disorder. Psychoneuroendocrinology. 2007;32:267–278. doi: 10.1016/j.psyneuen.2006.12.012. [DOI] [PubMed] [Google Scholar]
- 31.Sorrells S, Caso J, Munhoz C, Sapolsky R. The stressed CNS: when glucocorticoids aggravate inflammation. Neuron. 2009;64:33–39. doi: 10.1016/j.neuron.2009.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Otmishi P, Gordon J, El-Oshar S, et al. Neuroimmune interaction in inflammatory diseases. Clin Med Circ Respirat Pulm Med. 2008;2:35–44. doi: 10.4137/ccrpm.s547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kawabata K, Kawai Y, Terao J. Suppressive effect of quercetin on acute stress-induced hypothalamic-pituitary-adrenal axis response in Wistar rats. J Nutr Biochem. 2010;21:374–380. doi: 10.1016/j.jnutbio.2009.01.008. [DOI] [PubMed] [Google Scholar]
- 34.Youn Y, Sung IK, Lee IG. The role of cytokines in seizures: interleukin (IL)-1β, IL-1Ra, IL-8, and IL-10. Korean J Pediatr. 2013;56:271–274. doi: 10.3345/kjp.2013.56.7.271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Vezzani A, Balosso S, Ravizza T. The role of cytokines in the pathophysiology of epilepsy. Brain Behav Immun. 2008;22:797–803. doi: 10.1016/j.bbi.2008.03.009. [DOI] [PubMed] [Google Scholar]
- 36.Himmerich H, Fischer J, Bauer K, Kirkby KC, Sack U, Krugel U. Stress-induced cytokine changes in rats. Eur Cytokine Netw. 2013;24:97–103. doi: 10.1684/ecn.2013.0338. [DOI] [PubMed] [Google Scholar]
- 37.Swarup V, Ghosh J, Duseja R, Ghosh S, Basu A. Japanese encephalitis virus infection decrease endogenous IL-10 production: correlation with microglial activation and neuronal death. Neurosci Lett. 2007;420:144–149. doi: 10.1016/j.neulet.2007.04.071. [DOI] [PubMed] [Google Scholar]