

Abstract
Hedgehog signaling is required for many aspects of development in vertebrates and invertebrates. Misregulation of the Hedgehog pathway causes developmental abnormalities and has been implicated in certain types of cancer. Large-scale genetic screens in zebrafish have identified a group of mutations, termed you-class mutations, that share common defects in somite shape and in most cases disrupt Hedgehog signaling. These mutant embryos exhibit U-shaped somites characteristic of defects in slow muscle development. In addition, Hedgehog pathway mutations disrupt spinal cord patterning. We report the positional cloning of you, one of the original you-class mutations, and show that it is required for Hedgehog signaling in the development of slow muscle and in the specification of ventral fates in the spinal cord. The you gene encodes a novel protein with conserved EGF and CUB domains and a secretory pathway signal sequence. Epistasis experiments support an extracellular role for You upstream of the Hedgehog response mechanism. Analysis of chimeras indicates that you mutant cells can appropriately respond to Hedgehog signaling in a wild-type environment. Additional chimera analysis indicates that wild-type you gene function is not required in axial Hedgehog-producing cells, suggesting that You is essential for transport or stability of Hedgehog signals in the extracellular environment. Our positional cloning and functional studies demonstrate that You is a novel extracellular component of the Hedgehog pathway in vertebrates.
Genetic studies in zebrafish have identified a new protein involved in Hedgehog signaling - a key pathway required for development in vertebrates and invertebrates
Introduction
The coordination of growth, proliferation, and differentiation during development requires transmission of information in the form of extracellular signals. Hedgehog signaling is of fundamental importance in the development of a wide variety of tissues and organ systems. Much of the initial functional analysis of Hedgehog signaling focused on the patterning of Drosophila larval segments and imaginal discs, dorsoventral patterning of the vertebrate neural tube, and anterior–posterior patterning of vertebrate limbs; in addition, many recent studies have illuminated the widespread and conserved role of Hedgehog signaling in development (reviewed in [1]). Misregulation of Hedgehog signaling has been implicated in several diseases and developmental abnormalities, including basal cell carcinoma [2,3,4], medulloblastoma [5,6,7], pancreatic cancer [8], and holoprosencephaly [9,10].
After release from signaling cells, the activity and distribution of Hedgehog proteins are modulated by a variety of factors in the extracellular environment. In Drosophila, diffusion of lipid-modified Hedgehog proteins is dependent on the action of tout velu, a gene involved in the synthesis of heparan sulfate proteoglycans [11,12]. The diffusion of Hedgehog is also attenuated via sequestration by its receptor, Patched [13]. In vertebrates, Hedgehog proteins may be further regulated by binding to the growth-arrest specific gene product Gas1 [14], and Hedgehog-interacting protein Hip1, which is itself induced by Hedgehog signaling [15]. Moreover, the ability of Hedgehog proteins to diffuse over significant distances in the developing vertebrate limb bud appears to depend on the cholesterol modification of the Hedgehog protein; this modification may facilitate the assembly of Hedgehog proteins into a multimeric structure, perhaps conferring increased stability or mobility [16,17]. Genetic and biochemical evidence suggests that the low-density receptor-related protein Megalin may also play a role in Hedgehog signaling in vertebrates, perhaps by binding to Hedgehog proteins and facilitating their endocytosis [18,19].
Hedgehog pathway function in zebrafish has been analyzed primarily in the context of skeletal muscle development and differentiation [20,21,22,23,24,25,26,27,28]. In zebrafish embryos at 24 h post fertilization (hpf), skeletal muscle can be subdivided into two distinct classes based on morphological characteristics and gene expression. Slow muscle fibers are mononucleate, express characteristic slow muscle forms of myosin heavy chain, and show strong nuclear expression of the transcription factor prox1. In contrast, fast muscle fibers can be identified via their multinucleate morphology and lack of prox1 expression [28]. Cell labeling experiments have demonstrated that slow muscle fibers derive from the adaxial cells that lie immediately adjacent to the notochord [21]. As development progresses, a subset of these developing slow muscle cells migrates laterally through the myotome to form the superficial slow fibers [21,27,28]. Slow muscle fibers that remain near the midline—the muscle pioneers—express high levels of Engrailed and organize the somites into their distinctive chevron shape [22,28,29,30]. The remaining muscle cells in the interior of the myotome form multinucleate fast muscle fibers [21,28].
Many lines of evidence indicate that Hedgehog signals from axial tissues specify slow muscle in zebrafish. Slow muscle fibers are reduced or absent in embryos with Hedgehog pathway mutations [25,26,31,32,33]. Conversely, slow muscle is expanded at the expense of fast muscle in embryos with increased Hedgehog pathway activity [20,23,24,34]. Moreover, addition of Hedgehog protein to cultured zebrafish myoblasts induces expression of slow-muscle-specific forms of myosin heavy chain [35].
Genetic screens have identified a number of mutations disrupting the Hedgehog pathway in zebrafish [32,33,36,37,38,39,40,41]. Many of these Hedgehog pathway mutants share characteristic defects, the most obvious of which is abnormal somite morphology resulting from disrupted slow muscle specification and the lack of horizontal myoseptum [22]. These mutants are thus termed “you-class” mutants because of their U-shaped somites. Five of the seven you-class mutations have been cloned, and four of these genes, syu/shh, yot/gli2, smu/smoh, and con/disp1, encode members of the Hedgehog signaling pathway [32,33,36,37,39]. The exception is ubo/prdm1, which encodes a transcriptional switch that acts downstream of Hedgehog signaling in the development of slow muscle [27,42]. Careful analysis reveals differences between the ubo and Hedgehog pathway mutant phenotypes. For example, Hedgehog pathway mutants have defects in the lateral floor plate of the neural tube and the dorsal aorta, which are apparently normal in ubo mutants [22,43]. Examination of Hedgehog-induced gene expression also reveals a clear distinction between Hedgehog pathway mutations and ubo: Hedgehog pathway mutations reduce or abolish expression of the Hedgehog target ptc1, whereas ptc1 expression is normal in ubo mutants, indicating that they can receive Hedgehog signals [22,25,26,32,33,36,39].
Previous phenotypic characterization of mutants for the eponymous you-class gene, you, has revealed delayed development of the dorsal aorta and the absence of lateral floor plate marker expression in addition to slow muscle defects [22,43]. Moreover, expression of Hedgehog target genes, including ptc1 and adaxial myod, is reduced in you mutants [25]. These results suggest that the you gene acts within the Hedgehog pathway itself rather than downstream of Hedgehog signaling in processes specific to slow muscle development. Prior to this study, the molecular identity of the you gene has remained unknown. We report the positional cloning of the you gene and show that it encodes a novel extracellular EGF-CUB (epidermal growth factor–complement Uegf Bmp1) protein required for Hedgehog signaling. Functional studies provide evidence that you is essential for the transport or stability of Hedgehog signals in the extracellular environment.
Results
you Function Is Required for Hedgehog Signaling
In wild-type zebrafish embryos, the dorsal and ventral portions of each myotome converge at a point where the horizontal myoseptum forms, giving the somites their characteristic chevron shape (Figure 1A). In contrast, you mutants lacked the horizontal myoseptum and exhibit the U-shaped morphology that defines mutants of the you class (Figure 1B; [22]). The formation of the horizontal myoseptum in zebrafish depends on the proper development of slow muscle, a process that is defective in you-class mutants [22,31]. Hedgehog activity in the context of slow muscle cell specification can be assayed by analyzing Engrailed expression in the muscle pioneers and adaxial myod expression during somitogenesis. Wild-type embryos at 24 hpf exhibit strong Engrailed staining in characteristic elongate nuclei of 2–6 muscle pioneers—slow muscle cells that develop along the prospective horizontal myoseptum—per myotomal segment (Figure 1C). In addition, weaker Engrailed expression can be observed in the typically more rounded nuclei of mulitnucleate fast muscle fibers, which are situated farther from the horizontal myoseptum (Figure 1C; [28,30]). you mutant embryos completely lacked the strong Engrailed expression in the muscle pioneers (Figure 1D; [22]), but very weak labeling was sometimes observed in a small number of cells. When weakly expressing cells were present, they were confined to the nine most anterior somites. In wild-type embryos, myod is expressed during somitogenesis in the adaxial cells of both somitic and presomitic mesoderm, and laterally along the posterior borders of developing somites (Figure 1E). you mutant embryos retained the lateral expression of myod, but the adaxial expression of myod was absent in the trunk and reduced in the tail bud (Figure 1F; [25]). In addition, expression of ptc1—both a member of the Hedgehog pathway and a sensitive transcriptional readout of Hedgehog signaling—was reduced in the adaxial cells of you mutants at 22 somites and other stages (Figure 1G and 1H; data not shown; [25]).
Figure 1. you Mutants Exhibit Hedgehog-Associated Defects in Slow Muscle and Ventral Spinal Cord.

(A and B) Lateral views of live zebrafish at 22 hpf. Wild-type embryos (A) have chevron-shaped somites and a clearly visible floor plate (arrowhead), while you mutants (B) exhibit U-shaped somites and an indistinct floor plate (arrowhead).
(C and D) Lateral views of somites 8–13 in whole-mount embryos at 24 hpf. Wild-type embryos (C) show strong Engrailed expression in muscle pioneers (arrow), and weaker expression in multinucleate medial fast fibers (arrowheads). Engrailed expression in you mutants (D) is mostly absent, though very weak expression can occasionally be observed (arrowhead).
(E and F) Dorsal views of posterior trunk and tail bud in 12-somite embryos. Wild-type embryos (E) exhibit adaxial myod expression throughout the somitic (arrowhead) and presomitic (arrow) mesoderm, while you mutants (F) lack expression in the somitic (arrowhead) and in parts of the presomitic (arrow) mesoderm.
(G and H) Lateral view of somites 9–15 in whole-mount embryos at 22 hpf. Wild-type embryos (G) exhibit strong expression of ptc1, while you mutants (H) show weaker levels of ptc1 expression.
(I and J) Lateral view of spinal cord in the posterior trunk of whole-mount embryos at 24 hpf. Wild-type embryos (I) show expression of nkx2.2 in the ventral spinal cord, while in you mutants (J) this expression is strongly reduced.
(K–N) Dorsal views of whole-mount embryos at bud stage (10 hpf). Expression in you mutant embryos of both ehh (L) and shh (N) is similar to that observed wild-type embryos (K and M).
Anterior is to the left in all images. Genotypes of all embryos were determined by PCR after photography.
In addition to these disruptions in developing somites, you mutant embryos showed defects in patterning of the central nervous system. nkx2.2, a Hedgehog-induced marker of ventral cell types in the spinal cord [44], was absent in the trunk and tail of you mutants (Figure 1I and 1J). Moreover, our analysis and prior work has revealed that you embryos show delayed and weakened blood circulation in the dorsal aorta (data not shown; [45]). These results and previous phenotypic analyses support the conclusion that you gene function is required for Hedgehog signaling in development of slow muscle, ventral spinal cord, and the dorsal aorta.
In zebrafish, three hedgehog genes—ehh, shh, and twhh—are expressed at the midline in early embryonic stages. To determine whether you is required for hedgehog gene transcription, we analyzed the expression of hedgehog genes in wild-type and you mutant embryos. Expression of all three hedgehog genes appeared normal in you embryos at bud stage (10 hpf; Figure 1K–1N; data not shown).
Positional Cloning of you
As the first step toward identifying the you gene, we mapped the you mutation to a 1-cM (12 recombinants among 1,156 meioses) region of LG 7, between simple sequence length polymorphism (SSLP) markers Z11119 and Z15270 (Figure 2A). By comparing the position of the you mutation to zebrafish genetic maps ([46,47]; unpublished data), we excluded as candidate genes more than 60 zebrafish orthologs of Hedgehog pathway genes and genes known to interact with the Hedgehog pathway. We therefore adopted a positional cloning strategy to identify the gene disrupted by the you mutation. Using a marker linked with Z15270 on a contig from the Sanger Institute whole-genome shotgun assembly, we screened a pooled bacterial artificial chromosome (BAC) library and initiated a chromosome walk beginning with BAC zC172H20. After identifying polymorphisms in BAC end sequences, testing these polymorphisms on our mapping panel, and iteratively rescreening the pooled BAC library, we identified a contiguous stretch of genomic sequence spanning portions of five BACs with ends that flanked you (Figure 2A).
Figure 2. Positional Cloning of the you Locus.

(A) Genetic and physical map of the you region on LG 7, showing the initial flanking SSLPs and the BACs used in the chromosome walk. The number of recombinants in 1,156 meioses is shown for the SSLP markers and for mapped BAC end sequences.
(B) Diagram of BAC zC93A15, with expressed sequence tag markers BM184987 and AI722938 shown flanking the you locus. Both BM184987 and AI722938 showed one recombinant out of 6,514 meioses, and were genetically localized on opposite sides of you.
(C) The genomic region of the you locus is depicted by horizontal black bars. Gaps in these bars represent fragments of genomic sequence that were not obtained in the sequencing analysis. Exons are depicted by blue rectangles, and the number of recombinants in 6,514 meioses is shown below each mapped exon. Single nucleotide polymorphisms in four exons always segregated with the you locus, and one of these exons harbored a single nucleotide lesion predicted to change a glutamine codon to a stop codon and truncate the open reading frame.
To reduce the critical interval that contained you, we improved the resolution of our map by increasing our mapping panel to 6,514 meioses. By scoring sequences identified from BAC zC93A15 in key recombinants from the mapping panel, we localized the you gene to a portion of this BAC between polymorphisms identified in expressed sequence tag markers BM184987 and AI722938 (Figure 2B). Further sequence analysis and mapping identified other exons of the same gene as AI722938, some of which were on the opposite side of the mutation from the original AI722938 marker (Figure 2C). We isolated a full-length cDNA clone for this gene and used this new sequence information to identify polymorphisms in other exons. In all, high-resolution mapping identified four exons that failed to recombine with you and confirmed that the two ends of the gene flanked the youty97 mutation. Sequence analysis of the four non-recombining exons in wild-type and youty97 mutant genomic DNA identified a nonsense mutation that truncates the predicted protein approximately two-thirds of the way through the open reading frame. These findings, together with others described below, indicate that this gene is disrupted by the you mutation.
you Is Orthologous to Scube2
The protein encoded by you comprises 1,010 amino acids, and comparison of the predicted you amino acid sequence against the protein database indicated that the you protein is highly similar to a family of proteins founded by mouse SCUBE1 (Signal sequence, CUB domain, EGF-related; [48]). Pairwise sequence comparisons between you and SCUBE family members revealed that the You protein most closely matched SCUBE2 in mouse (65% identity) and SCUBE2 in human (66% identity). The orthology of these genes was further supported by comparative mapping: the human genes SCUBE2, LMO1, STK33, and ST5 exhibited conserved synteny with orthologous genes in both mouse and zebrafish ([49]; unpublished data).
SCUBE proteins are characterized by a signal peptide and by two types of conserved extracellular domains: EGF and CUB [50]. In all identified SCUBE family members, the N-terminal signal sequence is followed by nine EGF repeats, a spacer region, and a single C-terminal CUB domain [48,51,52,53]. Figure 3A shows an alignment comparing the predicted you amino acid sequence with selected SCUBE proteins in mouse and human. Similarity between the You protein and mouse SCUBE2 was particularly high in the CUB domain (89% identity), the C-terminal sequence following the CUB domain (90% identity), and the EGF repeats (74% identity). A spacer region in the center of the amino acid sequence showed lower conservation (47% identity). Examination of this spacer region in the vicinity of the CUB domain revealed a repeated motif of six cysteine residues with characteristic and regular spacing, shown in yellow in Figure 3A. Conservation in amino acid sequence was notably higher in this region (66% identical) than in the remainder of the spacer domain (33% identical). This six-cysteine repeat motif does not match the general structure of EGF repeats. The functional significance of this motif is not presently known, but it does lie within a region of SCUBE1 that is required in cell culture for secretion and cell surface expression [52].
Figure 3. SCUBE Protein Alignment and Truncation of the You Protein.

In (A) and (C), the signal peptide is labeled in blue, the nine EGF domains are labeled in red, and the CUB domain is labeled in green.
(A) Alignment of the predicted You amino acid sequence with SCUBE proteins in mouse and human. Identical amino acids are shaded, and similar amino acids are boxed. Conserved cysteines in these domains and elsewhere in the alignment are indicated by filled circles. A conserved six-cysteine repeat motif N-terminal to the CUB domain is labeled in yellow. The location of the glutamine residue at amino acid 644 in the zebrafish protein, which is changed to a stop codon in youty97 mutants, is boxed in bold.
(B) Sequence traces from homozygous wild-type and youty97 embryos. In youty97 mutants, a C to T transition is predicted to change a glutamine codon (CAA) to a stop codon (TAA) and truncate the open reading frame.
(C) Model of You protein domain structure. The You protein in youty97 mutants is predicted to be truncated prior to the six-cysteine repeat motifs, the CUB domain, and the conserved C-terminus.
In youty97 mutants, a C to T transition alters the coding sequence at residue 644, changing a glutamine codon to a stop codon (Figure 3A and 3B). The predicted mutant protein is truncated immediately prior to the six-cysteine repeat motif, so that these repeats, the CUB domain, and the highly conserved C-terminus are lacking (Figure 3C).
Morpholino Phenocopy and RNA Rescue of you
To confirm that this zebrafish EGF-CUB gene is disrupted in you mutants, we performed morpholino oligonucleotide (MO) injection experiments to phenocopy defects seen in you, and mRNA injection experiments to rescue the you phenotype in mutants (Figure 4). All wild-type embryos injected with a MO targeting the translational start site showed reduced expression of myod (n = 112) in the adaxial cells (Figure 4B), and an absence of strong Engrailed expression (n = 70) in the muscle pioneers (Figure 4D). Embryos injected with a mismatch control MO did not exhibit these defects in either myod (n = 65) or Engrailed (n = 48) expression (Figure 4A and 4C).
Figure 4. MO-Induced Phenocopy of you Defects and Rescue of the you Phenotype with mRNA Injection.

(A, B, E, and F) Dorsal view of the posterior trunk and tail bud of 12-somite embryos.
(C, D, G, and H) Lateral views of somites 8–13 in whole-mount embryos at 24 hpf.
Anterior is to the left in all images. Injection at the 1–4-cell stage of 420 pg of a MO targeting the translational start site of the you mRNA (ATG MO) resulted in decreased adaxial expression of myod in the somitic (arrowhead) and presomitic (arrow) mesoderm of wild-type embryos (B). Injection of an equivalent amount of a mismatch control (mismatch MO) did not produce these defects (A). Similarly, wild-type embryos injected with 420 pg of the mismatch MO (C) exhibited strong Engrailed expression in muscle pioneers (arrow) and weaker expression in medial fast fibers (arrowheads). In contrast, Engrailed expression was strongly reduced in wild-type embryos injected with 420 pg of the ATG MO (D), though very weak expression was still observed (arrowhead). Genotypically you mutant embryos (E) showed rescued expression of adaxial myod in somitic (arrowhead) and presomitic (arrow) mesoderm when injected with 50 pg of synthetic you mRNA at the 1–4-cell stage, while mutants injected with 50 pg of a frameshift mutant form of you mRNA (F) did not exhibit rescue of adaxial myod expression. At 24 hpf, genotypically you mutant embryos injected at the 1–4-cell stage with 50 pg of you mRNA (G) showed rescue of strong Engrailed expression in the muscle pioneers (arrow) and weaker expression in the medial fast fibers (arrowheads). Mutant embryos injected with 50 pg of the mutant mRNA (H) did not show rescued Engrailed expression, though very weak Engrailed expression (arrowhead) was observed in some cases. Engrailed expression at the MHB was normal in all analyzed embryos (data not shown). Genotypes of embryos shown in (E–H) were determined by PCR after photography.
When injected with 50 pg of synthetic wild-type you mRNA, 98.7% (n = 665) of embryos from you/+ intercrosses showed expression of myod in adaxial cells at 12 somites (Figure 4E). Genotyping of 571 embryos with wild-type myod expression from these intercrosses showed that 137 (24%) were homozygous mutant for you. In contrast, 23.6% (n = 127) of embryos from you/+ intercrosses injected with a mutant form of you mRNA lacked expression of myod in adaxial cells (Figure 4F); 20 of the mutants were genotyped and confirmed to be you mutant homozygotes. Similarly, all embryos (n = 62) from a you/+ intercross injected with wild-type mRNA showed strong Engrailed labeling in the muscle pioneers (Figure 4G; genotyping of 32 phenotypically wild-type embryos showed that seven were homozygous youty97), whereas 24% (n = 33) of embryos injected with control mRNA lacked Engrailed expression (Figure 4H; eight phenotypic mutants were confirmed as homozygous for the you mutation). In addition, injection of you MOs resulted in loss of nkx2.2 expression in the trunk and tail of wild-type embryos at 24 hpf, and injection of 50 pg of you mRNA was sufficient to rescue nkx2.2 expression in you mutants (data not shown).
you Expression
you transcripts appear to be maternally deposited in zebrafish embryos (Figure 5A) and are distributed widely in the embryo through early gastrulation stages (Figure 5B). During late gastrulation, the distribution of you transcripts in the embryo began to be restricted, and at bud stage (10 hpf) you was expressed in the eye field, in distinct bilateral domains within the developing brain, and in the developing trunk of the embryo in broad paraxial stripes (Figure 5C). During somitogenesis, you expression continued to be refined, such that six-somite embryos exhibited expression in the eye field, in stripes in the midbrain and the midbrain–hindbrain boundary (MHB), in a complex pattern in the hindbrain, and in paraxial stripes along the anterior–posterior axis (Figure 5D). At 24 hpf (Figure 5E and 5F), you transcripts were localized to the border of the ventral telencephalon and the dorsal diencephalon and to the ventral tectum, and were strongly expressed in the MHB, the hindbrain, and along the length of the embryo in the dorsal spinal cord. In addition, you expression was observed in the ventral tail and posterior to the yolk extension at the developing urogenital opening (Figure 5E). In the following 24 h of development, dorsal spinal cord expression continued, and you transcripts persisted in a complex and dynamic pattern in the brain. At 48 hpf (Figure 5G and 5H), you expression was particularly strong in the cerebellum and in the hindbrain along the rhombic lip.
Figure 5. Expression of you Examined by In Situ Hybridization.

(A and B) Maternal you transcripts were evident in cleavage-stage embryos (A) (16-cell; 1.5 hpf), and you mRNA was widely expressed into the gastrula period (B) (shield stage; 6 hpf). In addition, you mRNA was detectable by RT-PCR at 2 hpf, prior to the zebrafish midblastula transition.
(C) Toward the end of gastrulation, you transcripts began to be restricted, so that at the bud stage (10 hpf), you expression was evident in the eye field (white arrowhead), in the developing midbrain and hindbrain (black arrowheads), and in posterior paraxial stripes (arrow).
(D) During early somitogenesis (12 hpf), you expression was observed in the eye field (white arrowhead), in stripes in the midbrain and the MHB (black arrowheads), in a complex pattern in the hindbrain (white arrow), and in paraxial stripes along the developing trunk and tail bud (black arrow).
(E and F) At 24 hpf, you transcripts were observed dorsal to the hypothalamus (black arrow), at the boundary between the telencephalon and the diencephalon (white arrow), in the ventral tectum (white arrowhead), in the region of the presumptive cerebellum (asterisk), and dorsally along the length of the spinal cord. Additional expression of you at this stage and later was observed in the ventral tail and at the urogenital opening (arrowheads; data not shown).
(G and H) At 48 hpf, you transcripts were highly expressed in the cerebellum (black arrow), and were also present in the rhombic lip (white arrowhead), and continuing along the length of the anterior–posterior axis in the dorsal spinal cord (black arrowhead; data not shown).
Orientation of images: (A) lateral view; (B) lateral view, dorsal to the right; (C, D, F, and G) dorsal views, anterior to the left; (E and H) lateral views, anterior to the left.
Permissive Role of you Upstream of the Hedgehog Cellular Response
To explore the possibility that the you gene may induce a gain-of-function phenotype when overexpressed, we injected wild-type embryos with an amount (50 pg) of synthetic you mRNA that was sufficient to rescue the phenotypic defects observed in you mutants (Figure 6). When compared to embryos injected with a mutant form of you mRNA (myod, n = 81, Figure 6A; Engrailed, n = 25, Figure 6E; nkx2.2, n = 17, Figure 6I), overexpression of you in wild-type embryos did not result in obvious ectopic expression of myod (n = 394; Figure 6B), Engrailed (n = 25; Figure 6F), or nkx2.2 (n = 17; Figure 6J). This result suggests that you functions as a permissive factor in Hedgehog signaling, rather than as a potent activator of the Hedgehog pathway. When 50 pg of synthetic mRNA encoding a potent Hedgehog pathway activator (shh) was injected into embryos from a you/+ intercross, ectopic expression of myod (n = 53; Figure 6C and 6D), Engrailed (n = 78; Figure 6G and 6H), and nkx2.2 (n = 78; Figure 6K and 6L) was induced in all embryos. Genotyping a subset of these embryos indicated that both genotypically wild-type and you mutant embryos showed ectopic expression of each of these markers (myod: 41 wild-type, 12 you; Engrailed: 34 wild-type, 8 you; nkx2.2: 63 wild-type, 15 you). Because downstream targets of the Hedgehog pathway were rescued or upregulated in shh-injected you mutants, components of the Hedgehog pathway downstream of shh are most likely functional in you embryos. These results are consistent with you acting upstream of or parallel with shh in the Hedgehog pathway.
Figure 6. Early Overexpression of you in Wild-type Embryos and Rescue of you Defects by shh mRNA Injection.

(A–D) Dorsal views of the posterior trunk and tail bud of whole-mount embryos at 12 somites (15 hpf).
(E–F) Lateral views of somites 2–7 in 24-hpf embryos.
(G–H) Lateral views of somites 8–13 in 24-hpf embryos.
(I–L) Lateral views of 24-hpf embryos. Anterior is to the left in all images.
When 50 pg of you mRNA was injected into wild-type embryos at the 1–4-cell stage, no obvious expansion of myod (B), Engrailed (F), or nkx2.2 (J) expression was observed when compared either with wild-type embryos injected with equivalent amounts of mutant mRNA (A, E, and I) or with uninjected embryos (see Figure 1). Muscle pioneers were counted in a subset of the embryos; there were 4.0 ± 0.8 Engrailed-expressing muscle pioneers per somite in embryos injected with the control mRNA (n = 3 embryos, 33 somites) and 4.6 ± 1.1 muscle pioneers per somite in embryos injected with synthetic you mRNA (n = 8 embryos, 88 somites). Injection of 50 pg of shh mRNA into embryos at the 1–4-cell stage resulted in expansion of myod, Engrailed, and nkx2.2 expression in both wild-type (C, G, and K) and you mutant (D, H, and L) embryos. shh injection rescued adaxial expression of myod (D), muscle pioneer expression of Engrailed (H), and ventral spinal cord expression of nkx2.2 (L) in genotypically mutant you embryos (compare with Figure 1). Genotypes of all embryos were determined by PCR after photography.
Additional evidence that You acts upstream of the Hedgehog response derived from a loss-of-function approach, in which we activated the Hedgehog pathway by knocking down patched activity with MOs (Figure 7). Expression of myod in adaxial cells was rescued or expanded in all you mutant embryos that were injected with MOs targeting ptc1 (Figure 7E and 7G; n = 8 mutants) or a combination of MOs targeting both ptc1 and ptc2 (Figure 7I and 7K; n = 13 mutants). you mutant embryos injected with a ptc1 mismatch control MO did not exhibit rescued myod expression (Figure 7A and 7C; n = 8 mutants). In similar experiments, injection of patched MOs was sufficient to rescue or expand Engrailed expression in muscle pioneers of you mutant embryos (Figure 7B, 7D, 7F, 7H, 7J, and 7L; ptc1 MO, n = 11 mutants; ptc1 + ptc2 MO, n = 11 mutants).
Figure 7. Knockdown of patched Function Rescues Slow Muscle Defects in you .

After injection of 420 pg of a mismatch control ptc1 MO, adaxial expression of myod (A) and Engrailed (B) was normal in wild-type embryos, but absent in you mutant embryos (C and D). When injected with 420 pg of a MO targeting ptc1, however, myod expression in mutants (E) was rescued to levels comparable to wild-type embryos (G). Engrailed expression was slightly expanded in both wild-type (F) and mutant (H) embryos injected with 420 pg of ptc1 MOs. Both adaxial myod expression and Engrailed expression was slightly expanded in wild-type (I and J) and you mutant embryos (K and L) injected with MOs targeting both ptc1 and ptc2 (420 pg each). Embryos assayed for myod expression are shown in flat mount at the 12-somite stage, and somites 5–9 of Engrailed-expressing embryos are shown in lateral view at 24 hpf. Anterior is to the left in all panels. Genotypes of all embryos were determined by PCR after photography.
you Acts Non-Autonomously in Muscle Pioneer Differentiation
To determine whether a cell must be wild-type for you function to respond to Hedgehog signaling, we created genetic chimeras by transplanting cells from mutant embryos into wild-type hosts (Figure 8). Cells derived from you mutant embryos were able to differentiate as muscle pioneers, as defined by characteristic strong Engrailed expression in elongate nuclei of mononucleate cells at the proper position in the somite (Figure 8C–8E; n = 13 embryos). Similarly, cells from embryos in which you function had been reduced with MOs were able to differentiate as Engrailed-expressing muscle pioneers when introduced into embryos treated with mismatch control MOs (data not shown).
Figure 8. you Acts Non-Autonomously in Muscle Pioneers and Is Not Required in Cells Producing Hedgehog Signals.

(A) Donor embryos were labeled at the 1–4-cell stage with Oregon Green dextran.
(B) Cells from donor embryos were transplanted into unlabeled hosts during late blastula and early gastrula stages.
(C–E) Images from a chimera made by transplanting cells from labeled mutant donors (green in C and E) into unlabeled wild-type hosts. At 24 hpf, muscle pioneer cells in chimeric embryos were labeled with anti-Engrailed antibody (red in D and E). When transplanted into wild-type embryos, mutant cells were able to differentiate as muscle pioneers, as shown by co-labeling with the anti-Engrailed antibody (E, yellow arrows).
(F–K) Images from chimeras made by transplanting cells from labeled wild-type donors (green in F, H, I, and K) into unlabeled mutant hosts. Expression of Engrailed (red in G, H, J, and K) in some mutant muscle pioneers (one marked by red arrows in G, H, J, and K) was rescued in a subset of embryos (see also Table 1). Donor cells in the embryo shown in (F–H) contributed solely to muscle and to non-floor-plate identities within the neural tube. Moreover, in a subset of chimeras, cells derived from wild-type donors differentiated as muscle pioneer cells (yellow arrows in J and K), simultaneously showing both the characteristic strong nuclear Engrailed expression and the typical flattened and mononucleate morphology of this cell type. The somite labeled with the arrows in J and K contains two muscle pioneers, one derived from the wild-type donor (yellow arrow) and another derived from the mutant host (red arrow). Donor cells in the embryo shown in (I–K) contributed primarily to muscle and to non-floor-plate identities within the neural tube; in addition, a group of seven floor plate cells derived from the wild-type donor was present in the tail of this embryo (not shown).
Muscle Pioneer Differentiation Does Not Require you Function in Axial Hedgehog-Producing Cells
To determine which cell types must be wild-type for you function in order for target cells to appropriately respond to Hedgehog signals, we transplanted cells derived from wild-type donors into you mutant hosts. Of 91 chimeric mutant hosts, ten embryos exhibited rescue of Engrailed expression in genotypically mutant muscle pioneers, as defined by characteristic strong Engrailed expression in elongate nuclei at the proper position of posterior somites, where Engrailed is not normally expressed in you mutants (Table 1; Figure 8F–8K). In addition, wild-type cells differentiated as muscle pioneers in two of the ten chimeras with rescued mutant muscle pioneers. In these cases (Figure 8I–8K), the muscle pioneer identity of cells strongly expressing Engrailed was further confirmed by presence of the lineage tracer dye, which showed that these cells had the characteristic flattened and mononucleate morphology of muscle pioneers. In all ten embryos with rescued Engrailed expression, wild-type cells were present in the muscle and in non-floor-plate regions within the neural tube. In five of the chimeric embryos with rescued Engrailed expression, the floor plate and notochord were derived entirely from you mutant cells, indicating that you function is not required in axial Hedgehog-producing cells. Collectively, the transplantation experiments indicate that you function is not required in either the signaling or responding cells, and instead suggest that you is essential for the transport or stability of Hedgehog signals.
Table 1. Chimeras with Wild-Type Donor Cells in you Mutant Hosts.
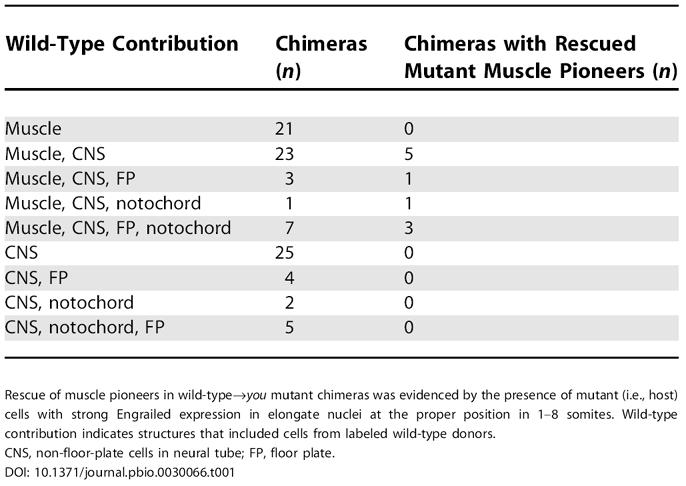
Rescue of muscle pioneers in wild-type→you mutant chimeras was evidenced by the presence of mutant (i.e., host) cells with strong Engrailed expression in elongate nuclei at the proper position in 1–8 somites. Wild-type contribution indicates structures that included cells from labeled wild-type donors
CNS, non-floor-plate cells in neural tube; FP, floor plate
Discussion
Using a positional cloning approach, we have shown that the you gene encodes a novel EGF-CUB protein essential for Hedgehog signaling in zebrafish. High-resolution mapping indicates that the you locus is tightly linked to a zebrafish homolog of mouse Scube2. youty97 mutants harbor a nonsense lesion that truncates the open reading frame upstream of the CUB domain and other highly conserved sequences. MO-mediated knockdown of the gene phenocopies defects observed in you mutants, and injection of wild-type mRNA into mutant embryos rescues the you phenotype. Taken together, these experiments provide compelling evidence that the you mutation disrupts this EGF-CUB gene.
Biochemical studies of SCUBE family members have shown that they are extracellular, membrane-associated glycoproteins [52,53], but no previous work implicates these proteins in the Hedgehog signaling pathway. Although the Hedgehog pathway has been extensively studied in flies and mammals, several factors have likely obscured the connection between this Scube gene and the Hedgehog pathway. Prior to this study, no loss of function analysis had been performed on any Scube family gene; there is no known Scube gene in the fly, and mouse mutants have not been reported. Also, because overexpression of synthetic you mRNA does not significantly hyperactivate Hedgehog signaling, Scube gene function in the Hedgehog pathway would not be apparent in gain-of-function screens to identify pathway components.
you mutant embryos exhibit phenotypic defects characteristic of reduced Hedgehog signaling in zebrafish, indicating that the you gene is a positively acting component of the Hedgehog pathway. Development of slow muscle is disrupted, as shown by lack of adaxial myod expression during somitogenesis and the absence of Engrailed-expressing muscle pioneer cells. Moreover, you mutants lack nkx2.2 expression in the ventral spinal cord, showing that specification of ventral neural fates is disrupted in this region of you embryos. Additionally, expression of the Hedgehog target gene ptc1 is reduced in you mutants. Analysis of the mutant phenotype, therefore, demonstrates that the you gene is essential for Hedgehog signaling in development of slow muscle and ventral spinal cord fates.
Current evidence indicates that EGF-CUB proteins, including You, have extracellular functions. The you gene product and other SCUBE proteins contain a signal peptide sequence targeting the protein for secretion, as well as EGF and CUB domains characteristic of extracellular proteins [48,50,51,52,53]. The You homolog SCUBE1 is a glycosylated peripheral membrane protein when expressed in 293T cells, and is also present at low levels in the culture medium [52]. EGF and CUB domains are found together in a small but diverse group of extracellularly acting proteins, including the complement subunits C1s and C1r, the metalloproteinase Tolloid, the sea urchin extracellular matrix protein Fibropellin, the serum glycoprotein Attractin, and the scavenger receptor Cubilin. CUB domains have been implicated in mediating protein–protein interactions and may confer specificity to ligand binding; for example, specific CUB domains in Cubilin have been implicated in facilitating the binding and subsequent endocytosis of specific ligands (reviewed in [54]).
Although you is essential for Hedgehog signaling, our analysis indicates that you mutant cells are able to produce and respond to Hedgehog signals. hedgehog gene expression in the embryonic midline is normal in you mutants, indicating that You acts downstream of hedgehog gene transcription (see Figure 1). Further evidence that you function is not required in cells generating Hedgehog signals derives from the analysis of chimeric embryos: muscle pioneers can differentiate in chimeras in which the notochord and floor plate are formed entirely from mutant cells (see Figure 8; Table 1). Conversely, you mutants can respond to Hedgehog pathway activation, mediated by either shh overexpression or disruption of patched, demonstrating that the defect in the mutants lies upstream of cellular response mechanism (see Figures 6 and 7). In addition, you mutant cells can respond to Hedgehog and differentiate as muscle pioneers when transplanted into a wild-type host (see Figure 8), indicating that you gene function is not required in cells responding to Hedgehog signals. The analysis of chimeras also demonstrates that the presence of wild-type cells in the paraxial mesoderm and neural plate is sufficient to allow you mutant cells to respond to Hedgehog signals produced from you mutant notochord and floor plate. Thus, our epistasis and transplantation experiments provide evidence that You functions in the extracellular environment in the field of responding cells, likely acting to transport or stabilize Hedgehog signals. It is possible that You interacts with components of the extracellular matrix, some of which are known to regulate the action of Hedgehog signals in other systems [11,12,55,56].
Another possible model is that You activates the Hedgehog pathway indirectly, by inhibiting an extracellular pathway antagonist such as Hip1 or Gas1. Our results, however, do not support such a model. Overexpression of high levels of synthetic you mRNA (up to ten times the amount required to rescue you mutants) in wild-type embryos did not result in obvious phenotypes when assayed by gross morphology or by expression of Hedgehog target genes, including adaxial myod, Engrailed in muscle pioneers, or nkx2.2 in the ventral spinal cord (see Figure 6; data not shown). The finding that overexpression of you does not significantly hyperactivate Hedgehog targets argues against simple models in which you functions to counteract an endogenous repressor of the Hedgehog pathway.
An interesting aspect of the you mutant phenotype is that it encompasses only a subset of defects seen in other zebrafish Hedgehog pathway mutants. Whereas mutants for syu/shh, yot/gli2, smu/smoh, and con/disp1 have prominent midline abnormalities in the head, such as ipsilateral retinotectal projections, reduction of anterior pituitary, and defects in medial neurocranial cartilage, these phenotypes are not evident in you mutants [26,32,33,37,43,57,58,59]. Also, development of pectoral fins, which is disrupted in syu, con, and smu [26,60], appears normal in you embryos. you mutants, therefore, show characteristic Hedgehog signaling defects in slow muscle specification, patterning of ventral spinal cord, and the development of the dorsal aorta, but you is apparently not required for Hedgehog signaling in some other regions of the zebrafish embryo. Because the primary cell types disrupted in you mutants all develop in close proximity to the notochord, it is possible that you gene function may be required for the transport or stability of Hedgehog signals in the vicinity of the developing notochord but not some other regions. The notochord is a defining feature of chordates, and a notochord-associated function would explain why no you counterpart is required for Hedgehog signaling in the fly. It is not clear, however, why Hedgehog signaling near the notochord would require a special extracellular mediator. Another possibility is that maternal you function could mask earlier requirements in zygotic you mutants; future work with maternal-zygotic you mutants is needed to address this possibility. A third explanation of the requirement for you in only a subset of Hedgehog-regulated processes is that additional factors with redundant functions may substitute for you in other regions of the embryo. Intriguingly, expression of another Scube gene in mouse—Scube1—is observed in many embryonic tissues known to require Hedgehog signaling for their proper development, including the ventral forebrain, limb bud, somites, and developing gonad [48]. These results suggest that an additional zebrafish Scube gene may also play a role in the development of other areas of the embryo where Hedgehog signaling is active. Moreover, interactions between SCUBE proteins may be important for Hedgehog signaling; biochemical analysis suggests that SCUBE1 and SCUBE2 proteins can interact to form both homodimers and heterodimers [52].
In addition to its role in promoting Hedgehog signaling in the developing muscle pioneers, ventral neural tube, and dorsal aorta, the expression pattern of you suggests that the gene may act in other cell types and perhaps in other pathways. During gastrulation, when Hedgehog signaling is required for specification of muscle pioneers [28], you is widely expressed. In 24-hpf embryos, however, you is expressed strongly in specific regions in the forebrain, midbrain, and hindbrain, and dorsally along the length of the spinal cord. Some of these expression domains overlap with regions of Hedgehog signaling, whereas others do not. One region where Hedgehog activity and you expression intersect at later embryonic stages is the cerebellum, where Hedgehog signaling plays a well-defined role in the proliferation of granule cell precursors in mammals [61,62]. In the trunk and tail, however, you expression in the dorsal spinal cord corresponds neither with known sources of Hedgehog signals nor with cells that require Hedgehog signaling for their proper development. This result suggests that you may function in other pathways later in development. Future studies will define the role of you in the Hedgehog pathway and address the possibility that you and other Scube family genes also function in other signaling pathways.
Materials and Methods
Fish strains
Zebrafish embryos were maintained at 28.5 °C and were staged according to [63]. Wild-type embryos were derived from the WIK strain. All phenotypic analysis of you mutants was performed with embryos homozygous for the youty97 allele [22].
Genetic mapping
The mapping panel was generated by crossing youty97/+ individuals with wild-type fish from the WIK strain. youty97 heterozygotes in the F1 generation were intercrossed, and mutant and wild-type embryos in the F2 progeny were collected at 3–4 dpf for mapping. Genomic DNA was prepared from these embryos as described [64]. Primer sequences for SSLP markers were obtained from the MGH zebrafish database (http://zebrafish.mgh.harvard.edu). For initial localization of you, bulked segregant analysis (reviewed in [65]) was performed on DNA pools from 20 mutant and 20 wild-type embryos. Putative zebrafish orthologs of hedgehog-related genes were identified by reciprocal BLAST analysis and localized to the Heat Shock Panel as previously described [47].
BAC screening, chromosome walking, and BAC sequencing
The CHORI211 BAC library was screened by PCR to identify positive BAC clones (http://www.rzpd.de). BAC end sequences were obtained from the Sanger Institute database (http://trace.ensembl.org, and PCR primers were designed to amplify regions of these sequences. PCR amplicons were sequenced from homozygous wild-type and mutant embryos to identify nucleotide differences that generated restriction enzyme fragment length polymorphisms. These polymorphisms were tested on the mapping panel, and markers showing tighter linkage with you were iteratively screened against the BAC library until a contiguous stretch of genomic sequence with ends that flanked you was identified. The BAC zC93A15 was subcloned into pBluescript SK+ (Stratagene, La Jolla, California, United States) following double digest with either Pst I and EcoR I or Xba I and Xho I. Sequences were analyzed on a 3730 DNA Analyzer (Applied Biosystems, Foster City, California, United States). Sequences generated from this BAC were used in iterated searches against the zebrafish whole-genome shotgun assembly to nucleate contigs of genomic sequence. Sequencing primers were designed from these contigs and used to generate additional sequence data for zC93A15.
Plasmid constructs
A full-length you clone in pBluescript SK− (Stratagene) was isolated from a 15–19-hpf cDNA library (gift of Bruce Appel and Judith Eisen). A cDNA clone harboring a frameshift mutation that is predicted to truncate the you protein at amino acid residue 34 was isolated from the same library and was used as a control in overexpression experiments. A modified version of the pCS2+ expression vector was generated by cloning a 41-bp fragment into its EcoR I and Xba I sites. This stuffer fragment abolished the endogenous EcoR I, Stu I, Xho I, and Xba I restriction digest sites of pCS2+, and introduced Xba I, Sac I, Apa I, Pst I, and Xho I recognition sequences in a 5′ to 3′ orientation with respect to the SP6 promoter. Wild-type and mutant you clones were subcloned into the Xba I and Xho I sites of this modified pCS2+ vector. In situ probes for you were generated by linearizing this vector with Xba I, followed by antisense RNA synthesis with T3 polymerase. Synthetic you mRNA for injections was generated by digestion with Not I, followed by transcription using the SP6 mMessage mMachine kit (Ambion, Austin, Texas, United States).
In situ hybridization, antibody labeling, and genotyping
Probe synthesis, in situ hybridizations, and immunohistochemistry were performed using standard protocols. Embryos from you/+ intercrosses were genotyped after in situ hybridization and antibody labeling as described [66]. Other probes used were zebrafish myod [67], ptc1 [68], nkx2.2 [44], ehh [20], shh [69], and monoclonal antibody 4D9 [29]. Genotyping was performed by scoring a polymorphism in AI722938 (primer 1, GTGAAAGCAAAAAGCAAGCA; primer 2, GCACTGCATTATGTTTGTGGA; followed by a Hinf I digest).
Microinjections
Embryos were injected through their chorions with 500 pl of solution at the 1–4-cell stage as described [70]. RNA was diluted in 0.2 M KCl with 5 mg/ml Phenol Red prior to injection. MOs were obtained from Gene Tools (Philomath, Oregon, United States). A MO targeted to the you translational initiation site (5′- GCCGTACAGTCCAAACAGCTCCCAT-3′) or a 5-bp mismatch control MO (5′-GCCcTAg AGTCg AAACAcCTg CCAT-3′) was diluted in a 1x Danieau's solution with Phenol Red at 5 mg/ml prior to microinjection. Sequences for MOs targeting ptc1 and ptc2 were obtained from [28].
Transplantations
Cellular transplantations were done according to standard methods [22]. Embryos derived from you/+ intercrosses were injected at the 1–4-cell stage with a 1% solution of Oregon Green 488 dextran (Molecular Probes, Eugene, Oregon, United States). Approximately 50–100 cells were removed from labeled donors in late blastula and early gastrula periods (4–5.3 hpf) and transplanted near the margin of unlabeled sibling hosts. Labeled donor embryos were allowed to develop until 24 hpf. Genotypes of donor embryos derived from you/+ intercrosses were determined by PCR. Genotypes of host embryos were determined by staining with Engrailed antibody. Donor cells in chimeras with wild-type cells transplanted into you mutant hosts were obtained either from WIK intercrosses or from genotypically wild-type embryos in you/+ intercrosses.
Supporting Information
Accession Numbers
The you cDNA sequence has been deposited in GenBank (http://www.ncbi.nlm.nih.gov/Genbank/index.html) under accession number AY741664.
The LocusLink (http://www.ncbi.nlm.nih.gov/LocusLink/) accession numbers for the genes and gene products discussed in this paper are Attractin (LocusID 8455), C1r (LocusID 715), C1s (LocusID 716), con/disp1 (LocusID 378448), Cubilin (LocusID 8029), ehh (LocusID 30299), Engrailed (LocusID 30244), Gas1 (LocusID 14451), Hedgehog (LocusID 42737), Hip1 (LocusID 15245), Megalin (LocusID 14725), myod (LocusID 30513), nkx2.2 (LocusID 30697), Patched (LocusID 35851), prox1 (LocusID 30679), ptc1 (LocusID 30181), ptc2 (LocusID 30189), sea urchin Fibropellin (LocusID 373313), smu/smoh (LocusID 30225), syu/ssh (LocusID 30269), Tolloid (LocusID 42945), tout velu (LocusID 36614), twhh (LocusID 30444), ubo/prdm1 (LocusID 323473), yot/gli2 (LocusID 30154), human LMO1 (LocusID 4004), human SCUBE1 (LocusID 80274), human SCUBE2 (LocusID 57758), human ST5 (LocusID 6764), human STK33 (LocusID 65975), mouse SCUBE1 (LocusID 64706), and mouse SCUBE2 (LocusID 56788).
Acknowledgments
We thank Alex Schier and members of our lab for helpful discussions and comments on the manuscript, and Nipam Patel for supplying the 4D9 antibody. This work was supported by National Institutes of Health grant R01 RR12349 (WST) and a predoctoral fellowship from the Howard Hughes Medical Institute (IGW).
Competing interests. The authors have declared that no competing interests exist.
Abbreviations
- BAC
bacterial artificial chromosome
- con
chameleon
- hpf
hours post fertilization
- MHB
midbrain-hindbrain boundary
- MO
morpholino oligonucleotide
- SSLP
simple sequence length polymorphism
Author contributions. IGW and WST conceived and designed the experiments. IGW performed the experiments. IGW and WST analyzed the data and wrote the paper.
Citation: Woods IG, Talbot WS (2005) The you gene encodes an EGF-CUB protein essential for hedgehog signaling in zebrafish. PLoS Biol 3(3): e66.
References
- McMahon AP, Ingham PW, Tabin CJ. Developmental roles and clinical significance of hedgehog signaling. Curr Top Dev Biol. 2003;53:1–114. doi: 10.1016/s0070-2153(03)53002-2. [DOI] [PubMed] [Google Scholar]
- Hahn H, Wicking C, Zaphiropoulous PG, Gailani MR, Shanley S, et al. Mutations of the human homolog of Drosophila patched in the nevoid basal cell carcinoma syndrome. Cell. 1996;85:841–851. doi: 10.1016/s0092-8674(00)81268-4. [DOI] [PubMed] [Google Scholar]
- Johnson RL, Rothman AL, Xie J, Goodrich LV, Bare JW, et al. Human homolog of patched, a candidate gene for the basal cell nevus syndrome. Science. 1996;272:1668–1671. doi: 10.1126/science.272.5268.1668. [DOI] [PubMed] [Google Scholar]
- Xie J, Murone M, Luoh SM, Ryan A, Gu Q, et al. Activating Smoothened mutations in sporadic basal-cell carcinoma. Nature. 1998;391:90–92. doi: 10.1038/34201. [DOI] [PubMed] [Google Scholar]
- Goodrich LV, Milenkovic L, Higgins KM, Scott MP. Altered neural cell fates and medulloblastoma in mouse patched mutants. Science. 1997;277:1109–1113. doi: 10.1126/science.277.5329.1109. [DOI] [PubMed] [Google Scholar]
- Pietsch T, Waha A, Koch A, Kraus J, Albrecht S, et al. Medulloblastomas of the desmoplastic variant carry mutations of the human homologue of Drosophila patched. Cancer Res. 1997;57:2085–2088. [PubMed] [Google Scholar]
- Xie J, Johnson RL, Zhang X, Bare JW, Waldman FM, et al. Mutations of the PATCHED gene in several types of sporadic extracutaneous tumors. Cancer Res. 1997;57:2369–2372. [PubMed] [Google Scholar]
- Thayer SP, di Magliano MP, Heiser PW, Nielsen CM, Roberts DJ, et al. Hedgehog is an early and late mediator of pancreatic cancer tumorigenesis. Nature. 2003;425:851–856. doi: 10.1038/nature02009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belloni E, Muenke M, Roessler E, Traverso G, Siegel-Bartelt J, et al. Identification of Sonic hedgehog as a candidate gene responsible for holoprosencephaly. Nat Genet. 1996;14:353–356. doi: 10.1038/ng1196-353. [DOI] [PubMed] [Google Scholar]
- Ming JE, Kaupas ME, Roessler E, Brunner HG, Golabi M, et al. Mutations in PATCHED-1, the receptor for SONIC HEDGEHOG, are associated with holoprosencephaly. Hum Genet. 2002;110:297–301. doi: 10.1007/s00439-002-0695-5. [DOI] [PubMed] [Google Scholar]
- Bellaiche Y, The I, Perrimon N. Tout-velu is a Drosophila homologue of the putative tumour suppressor EXT-1 and is needed for Hh diffusion. Nature. 1998;394:85–88. doi: 10.1038/27932. [DOI] [PubMed] [Google Scholar]
- The I, Bellaiche Y, Perrimon N. Hedgehog movement is regulated through tout velu-dependent synthesis of a heparan sulfate proteoglycan. Mol Cell. 1999;4:633–639. doi: 10.1016/s1097-2765(00)80214-2. [DOI] [PubMed] [Google Scholar]
- Chen Y, Struhl G. Dual roles for patched in sequestering and transducing Hedgehog. Cell. 1996;87:553–563. doi: 10.1016/s0092-8674(00)81374-4. [DOI] [PubMed] [Google Scholar]
- Lee CS, Buttitta L, Fan CM. Evidence that the WNT-inducible growth arrest-specific gene 1 encodes an antagonist of sonic hedgehog signaling in the somite. Proc Natl Acad Sci U S A. 2001;98:11347–11352. doi: 10.1073/pnas.201418298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang PT, McMahon AP. Vertebrate Hedgehog signalling modulated by induction of a Hedgehog-binding protein. Nature. 1999;397:617–621. doi: 10.1038/17611. [DOI] [PubMed] [Google Scholar]
- Lewis PM, Dunn MP, McMahon JA, Logan M, Martin JF, et al. Cholesterol modification of sonic hedgehog is required for long-range signaling activity and effective modulation of signaling by Ptc1. Cell. 2001;105:599–612. doi: 10.1016/s0092-8674(01)00369-5. [DOI] [PubMed] [Google Scholar]
- Zeng X, Goetz JA, Suber LM, Scott WJ, Schreiner CM, et al. A freely diffusible form of Sonic hedgehog mediates long-range signalling. Nature. 2001;411:716–720. doi: 10.1038/35079648. [DOI] [PubMed] [Google Scholar]
- Willnow TE, Hilpert J, Armstrong SA, Rohlmann A, Hammer RE, et al. Defective forebrain development in mice lacking gp330/megalin. Proc Natl Acad Sci U S A. 1996;93:8460–8464. doi: 10.1073/pnas.93.16.8460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCarthy RA, Barth JL, Chintalapudi MR, Knaak C, Argraves WS. Megalin functions as an endocytic sonic hedgehog receptor. J Biol Chem. 2002;277:25660–25667. doi: 10.1074/jbc.M201933200. [DOI] [PubMed] [Google Scholar]
- Currie PD, Ingham PW. Induction of a specific muscle cell type by a hedgehog-like protein in zebrafish. Nature. 1996;382:452–455. doi: 10.1038/382452a0. [DOI] [PubMed] [Google Scholar]
- Devoto SH, Melancon E, Eisen JS, Westerfield M. Identification of separate slow and fast muscle precursor cells in vivo, prior to somite formation. Development. 1996;122:3371–3380. doi: 10.1242/dev.122.11.3371. [DOI] [PubMed] [Google Scholar]
- van Eeden FJ, Granato M, Schach U, Brand M, Furutani-Seiki M, et al. Mutations affecting somite formation and patterning in the zebrafish, Danio rerio . Development. 1996;123:153–164. doi: 10.1242/dev.123.1.153. [DOI] [PubMed] [Google Scholar]
- Blagden CS, Currie PD, Ingham PW, Hughes SM. Notochord induction of zebrafish slow muscle mediated by Sonic hedgehog. Genes Dev. 1997;11:2163–2175. doi: 10.1101/gad.11.17.2163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du SJ, Devoto SH, Westerfield M, Moon RT. Positive and negative regulation of muscle cell identity by members of the hedgehog and TGF-beta gene families. J Cell Biol. 1997;139:145–156. doi: 10.1083/jcb.139.1.145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis KE, Currie PD, Roy S, Schauerte H, Haffter P, et al. Control of muscle cell-type specification in the zebrafish embryo by Hedgehog signalling. Dev Biol. 1999;216:469–480. doi: 10.1006/dbio.1999.9519. [DOI] [PubMed] [Google Scholar]
- Barresi MJ, Stickney HL, Devoto SH. The zebrafish slow-muscle-omitted gene product is required for Hedgehog signal transduction and the development of slow muscle identity. Development. 2000;127:2189–2199. doi: 10.1242/dev.127.10.2189. [DOI] [PubMed] [Google Scholar]
- Roy S, Wolff C, Ingham PW. The u-boot mutation identifies a Hedgehog-regulated myogenic switch for fiber-type diversification in the zebrafish embryo. Genes Dev. 2001;15:1563–1576. doi: 10.1101/gad.195801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wolff C, Roy S, Ingham PW. Multiple muscle cell identities induced by distinct levels and timing of hedgehog activity in the zebrafish embryo. Curr Biol. 2003;13:1169–1181. doi: 10.1016/s0960-9822(03)00461-5. [DOI] [PubMed] [Google Scholar]
- Patel NH, Martin-Blanco E, Coleman KG, Poole SJ, Ellis MC, et al. Expression of engrailed proteins in arthropods, annelids, and chordates. Cell. 1989;58:955–968. doi: 10.1016/0092-8674(89)90947-1. [DOI] [PubMed] [Google Scholar]
- Hatta K, Bremiller R, Westerfield M, Kimmel CB. Diversity of expression of engrailed-like antigens in zebrafish. Development. 1991;112:821–832. doi: 10.1242/dev.112.3.821. [DOI] [PubMed] [Google Scholar]
- Stickney HL, Barresi MJ, Devoto SH. Somite development in zebrafish. Dev Dyn. 2000;219:287–303. doi: 10.1002/1097-0177(2000)9999:9999<::AID-DVDY1065>3.0.CO;2-A. [DOI] [PubMed] [Google Scholar]
- Chen W, Burgess S, Hopkins N. Analysis of the zebrafish smoothened mutant reveals conserved and divergent functions of hedgehog activity. Development. 2001;128:2385–2396. doi: 10.1242/dev.128.12.2385. [DOI] [PubMed] [Google Scholar]
- Varga ZM, Amores A, Lewis KE, Yan YL, Postlethwait JH, et al. Zebrafish smoothened functions in ventral neural tube specification and axon tract formation. Development. 2001;128:3497–3509. doi: 10.1242/dev.128.18.3497. [DOI] [PubMed] [Google Scholar]
- Hammerschmidt M, Bitgood MJ, McMahon AP. Protein kinase A is a common negative regulator of Hedgehog signaling in the vertebrate embryo. Genes Dev. 1996;10:647–658. doi: 10.1101/gad.10.6.647. [DOI] [PubMed] [Google Scholar]
- Norris W, Neyt C, Ingham PW, Currie PD. Slow muscle induction by Hedgehog signalling in vitro. J Cell Sci. 2000;113:2695–2703. doi: 10.1242/jcs.113.15.2695. [DOI] [PubMed] [Google Scholar]
- Schauerte HE, van Eeden FJ, Fricke C, Odenthal J, Strahle U, et al. Sonic hedgehog is not required for the induction of medial floor plate cells in the zebrafish. Development. 1998;125:2983–2993. doi: 10.1242/dev.125.15.2983. [DOI] [PubMed] [Google Scholar]
- Karlstrom RO, Talbot WS, Schier AF. Comparative synteny cloning of zebrafish you-too: Mutations in the Hedgehog target gli2 affect ventral forebrain patterning. Genes Dev. 1999;13:388–393. doi: 10.1101/gad.13.4.388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karlstrom RO, Tyurina OV, Kawakami A, Nishioka N, Talbot WS, et al. Genetic analysis of zebrafish gli1 and gli2 reveals divergent requirements for gli genes in vertebrate development. Development. 2003;130:1549–1564. doi: 10.1242/dev.00364. [DOI] [PubMed] [Google Scholar]
- Nakano Y, Kim HR, Kawakami A, Roy S, Schier AF, et al. Inactivation of dispatched 1 by the chameleon mutation disrupts Hedgehog signalling in the zebrafish embryo. Dev Biol. 2004;269:381–392. doi: 10.1016/j.ydbio.2004.01.022. [DOI] [PubMed] [Google Scholar]
- Sekimizu K, Nishioka N, Sasaki H, Takeda H, Karlstrom RO, et al. The zebrafish iguana locus encodes Dzip1, a novel zinc-finger protein required for proper regulation of Hedgehog signaling. Development. 2004;131:2521–2532. doi: 10.1242/dev.01059. [DOI] [PubMed] [Google Scholar]
- Wolff C, Roy S, Lewis KE, Schauerte H, Joerg-Rauch G, et al. iguana encodes a novel zinc-finger protein with coiled-coil domains essential for Hedgehog signal transduction in the zebrafish embryo. Genes Dev. 2004;18:1565–1576. doi: 10.1101/gad.296004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baxendale S, Davison C, Muxworthy C, Wolff C, Ingham PW, et al. The B-cell maturation factor Blimp-1 specifies vertebrate slow-twitch muscle fiber identity in response to Hedgehog signaling. Nat Genet. 2004;36:88–93. doi: 10.1038/ng1280. [DOI] [PubMed] [Google Scholar]
- Odenthal J, van Eeden FJ, Haffter P, Ingham PW, Nusslein-Volhard C. Two distinct cell populations in the floor plate of the zebrafish are induced by different pathways. Dev Biol. 2000;219:350–363. doi: 10.1006/dbio.1999.9589. [DOI] [PubMed] [Google Scholar]
- Barth KA, Wilson SW. Expression of zebrafish nk2.2 is influenced by sonic hedgehog/vertebrate hedgehog-1 and demarcates a zone of neuronal differentiation in the embryonic forebrain. Development. 1995;121:1755–1768. doi: 10.1242/dev.121.6.1755. [DOI] [PubMed] [Google Scholar]
- Chen JN, Haffter P, Odenthal J, Vogelsang E, Brand M, et al. Mutations affecting the cardiovascular system and other internal organs in zebrafish. Development. 1996;123:293–302. doi: 10.1242/dev.123.1.293. [DOI] [PubMed] [Google Scholar]
- Kelly PD, Chu F, Woods IG, Ngo-Hazelett P, Cardozo T, et al. Genetic linkage mapping of zebrafish genes and ESTs. Genome Res. 2000;10:558–567. doi: 10.1101/gr.10.4.558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woods IG, Kelly PD, Chu F, Ngo-Hazelett P, Yan YL, et al. A comparative map of the zebrafish genome. Genome Res. 2000;10:1903–1914. doi: 10.1101/gr.10.12.1903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimmond S, Larder R, Van Hateren N, Siggers P, Hulsebos TJ, et al. Cloning, mapping, and expression analysis of a gene encoding a novel mammalian EGF-related protein (SCUBE1) Genomics. 2000;70:74–81. doi: 10.1006/geno.2000.6370. [DOI] [PubMed] [Google Scholar]
- Amid C, Bahr A, Mujica A, Sampson N, Bikar SE, et al. Comparative genomic sequencing reveals a strikingly similar architecture of a conserved syntenic region on human chromosome 11p15.3 (including gene ST5) and mouse chromosome 7. Cytogenet Cell Genet. 2001;93:284–290. doi: 10.1159/000056999. [DOI] [PubMed] [Google Scholar]
- Bork P, Beckmann G. The CUB domain. A widespread module in developmentally regulated proteins. J Mol Biol. 1993;231:539–545. doi: 10.1006/jmbi.1993.1305. [DOI] [PubMed] [Google Scholar]
- Grimmond S, Larder R, Van Hateren N, Siggers P, Morse S, et al. Expression of a novel mammalian epidermal growth factor-related gene during mouse neural development. Mech Dev. 2001;102:209–211. doi: 10.1016/s0925-4773(00)00586-4. [DOI] [PubMed] [Google Scholar]
- Yang RB, Ng CK, Wasserman SM, Colman SD, Shenoy S, et al. Identification of a novel family of cell-surface proteins expressed in human vascular endothelium. J Biol Chem. 2002;277:46364–46373. doi: 10.1074/jbc.M207410200. [DOI] [PubMed] [Google Scholar]
- Wu BT, Su YH, Tsai MT, Wasserman SM, Topper JN, et al. A novel secreted, cell-surface glycoprotein containing multiple EGF-like repeats and one CUB domain is highly expressed in primary osteoblasts and bones. J Biol Chem. 2004;279:37485–37490. doi: 10.1074/jbc.M405912200. [DOI] [PubMed] [Google Scholar]
- Christensen EI, Birn H. Megalin and cubilin: Multifunctional endocytic receptors. Nat Rev Mol Cell Biol. 2002;3:256–266. doi: 10.1038/nrm778. [DOI] [PubMed] [Google Scholar]
- Pons S, Trejo JL, Martinez-Morales JR, Marti E. Vitronectin regulates Sonic hedgehog activity during cerebellum development through CREB phosphorylation. Development. 2001;128:1481–1492. doi: 10.1242/dev.128.9.1481. [DOI] [PubMed] [Google Scholar]
- Blaess S, Graus-Porta D, Belvindrah R, Radakovits R, Pons S, et al. Beta1-integrins are critical for cerebellar granule cell precursor proliferation. J Neurosci. 2004;24:3402–3412. doi: 10.1523/JNEUROSCI.5241-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baier H, Klostermann S, Trowe T, Karlstrom RO, Nusslein-Volhard C, et al. Genetic dissection of the retinotectal projection. Development. 1996;123:415–425. doi: 10.1242/dev.123.1.415. [DOI] [PubMed] [Google Scholar]
- Brand M, Heisenberg CP, Warga RM, Pelegri F, Karlstrom RO, et al. Mutations affecting development of the midline and general body shape during zebrafish embryogenesis. Development. 1996;123:129–142. doi: 10.1242/dev.123.1.129. [DOI] [PubMed] [Google Scholar]
- Karlstrom RO, Trowe T, Klostermann S, Baier H, Brand M, et al. Zebrafish mutations affecting retinotectal axon pathfinding. Development. 1996;123:427–438. doi: 10.1242/dev.123.1.427. [DOI] [PubMed] [Google Scholar]
- van Eeden FJ, Granato M, Schach U, Brand M, Furutani-Seiki M, et al. Genetic analysis of fin formation in the zebrafish, Danio rerio . Development. 1996;123:255–262. doi: 10.1242/dev.123.1.255. [DOI] [PubMed] [Google Scholar]
- Dahmane N, Ruiz-i-Altaba A. Sonic hedgehog regulates the growth and patterning of the cerebellum. Development. 1999;126:3089–3100. doi: 10.1242/dev.126.14.3089. [DOI] [PubMed] [Google Scholar]
- Wechsler-Reya RJ, Scott MP. Control of neuronal precursor proliferation in the cerebellum by Sonic Hedgehog. Neuron. 1999;22:103–114. doi: 10.1016/s0896-6273(00)80682-0. [DOI] [PubMed] [Google Scholar]
- Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embryonic development of the zebrafish. Dev Dyn. 1995;203:253–310. doi: 10.1002/aja.1002030302. [DOI] [PubMed] [Google Scholar]
- Talbot WS, Schier AF. Positional cloning of mutated zebrafish genes. Methods Cell Biol. 1999;60:259–286. doi: 10.1016/s0091-679x(08)61905-6. [DOI] [PubMed] [Google Scholar]
- Postlethwait JH, Talbot WS. Zebrafish genomics: From mutants to genes. Trends Genet. 1997;13:183–190. doi: 10.1016/s0168-9525(97)01129-3. [DOI] [PubMed] [Google Scholar]
- Sirotkin HI, Dougan ST, Schier AF, Talbot WS. bozozok and squint act in parallel to specify dorsal mesoderm and anterior neuroectoderm in zebrafish. Development. 2000;127:2583–2592. doi: 10.1242/dev.127.12.2583. [DOI] [PubMed] [Google Scholar]
- Weinberg ES, Allende ML, Kelly CS, Abdelhamid A, Murakami T, et al. Developmental regulation of zebrafish MyoD in wild-type, no tail and spadetail embryos. Development. 1996;122:271–280. doi: 10.1242/dev.122.1.271. [DOI] [PubMed] [Google Scholar]
- Concordet JP, Lewis KE, Moore JW, Goodrich LV, Johnson RL, et al. Spatial regulation of a zebrafish patched homologue reflects the roles of sonic hedgehog and protein kinase A in neural tube and somite patterning. Development. 1996;122:2835–2846. doi: 10.1242/dev.122.9.2835. [DOI] [PubMed] [Google Scholar]
- Krauss S, Concordet JP, Ingham PW. A functionally conserved homolog of the Drosophila segment polarity gene hh is expressed in tissues with polarizing activity in zebrafish embryos. Cell. 1993;75:1431–1444. doi: 10.1016/0092-8674(93)90628-4. [DOI] [PubMed] [Google Scholar]
- Westerfield M. The zebrafish book. Eugene: University of Oregon Press; 1994. 1 v. [Google Scholar]