

Abstract
Bacillus anthracis, the agent of anthrax, produces a poly-D-glutamic acid capsule that has been implicated in virulence. Many strains missing pXO2 (96 kb), which harbors the capsule biosynthetic operon capBCAD, but carrying pXO1 (182 kb) that harbors the anthrax toxin genes, are attenuated in animal models. Also, noncapsulated strains are readily phagocytosed by macrophage cell lines, whereas capsulated strains are resistant to phagocytosis. We show that a strain carrying both virulence plasmids but deleted specifically for capBCAD is highly attenuated in a mouse model for inhalation anthrax. The parent strain and capsule mutant initiated germination in the lungs, but the capsule mutant did not disseminate to the spleen. A mutant harboring capBCAD but deleted for the cap regulators acpA and acpB was also significantly attenuated, in agreement with the capsule-negative phenotype during in vitro growth. Surprisingly, an acpB mutant, but not an acpA mutant, displayed an elevated LD50 and reduced ability to disseminate, indicating that acpA and acpB are not true functional homologs and that acpB may play a larger role in virulence than originally suspected.
Keywords: anthracis, anthrax, capsule, infection, virulence
Introduction
The ability of Bacillus anthracis to cause anthrax disease has been primarily attributed to the production of a poly-D-glutamic acid capsule and two secreted toxins, anthrax lethal toxin and edema toxin (Thorne, 1993). These virulence factors are plasmid associated: pXO2 (96 kb; accession no. NC_002146) carries the capsule biosynthetic gene operon, capBCAD (accession no. NC_002146:52795–57483) (Makino et al, 1989; Uchida et al, 1993b), and the capsule gene regulators, acpA (accession no. NC_002146:68909–70360) (Vietri et al, 1995) and acpB (accession no. NC_002146:49418–50866) (Drysdale et al, 2004), while pXO1 (182 kb; accession no. NC_001496) harbors the toxin structural genes, cya (accession no. NC_001496:154224–156626) (Mock et al, 1988; Robertson et al, 1988; Tippetts and Robertson, 1988), pagA (accession no. NC_001496:133161–135455) (Vodkin and Leppla, 1983; Welkos et al, 1988), and lef (accession no. NC_001496:127442–129871) (Robertson and Leppla, 1986), and the toxin gene activator, atxA (accession no. NC_001496:150042–151469) (Uchida et al, 1993a; Koehler et al, 1994; Dai et al, 1995). This work focuses on the role of the B. anthracis capsule during infection in a mouse model for inhalation anthrax. The B. anthracis capsule is unusual in that it is composed solely of D-glutamic acid residues that are gamma-linked to form large homopolymers of over 215 kDa (γDPGA) (Record and Wallis, 1955). Although the structure of the capsule is known, the precise role of the capsule during infection has remained elusive (Record and Wallis, 1955; Zwartouw and Smith, 1956).
Most investigations of capsule function in animals have been performed using attenuated strains containing only one of the two large virulence plasmids (Ivins et al, 1986; Welkos, 1991). The reduced virulence of pXO1+ pXO2− strains has been attributed to the Cap− phenotype. However, pXO2 carries 85 predicted open reading frames (ORFs) and a pXO1+ pXO2+ capBCAD deletion mutant was not tested previously. pXO2 may carry factors other than the cap genes that contribute to the virulence of the organism. Welkos and co-workers (Welkos, 1991) identified transposon-derived mutants harboring insertions on pXO2 that produce wild-type levels of capsule when cultured in vitro but exhibit reduced virulence when injected subcutaneously in mice. They also demonstrated that differences in virulence between certain B. anthracis strains are partially linked to pXO2 (Welkos et al, 1993). In these experiments, the entire pXO2 plasmid was transduced from donor strains to pXO2-cured isolates and virulence was assessed by comparing LD50 values in a subcutaneous infection model (Welkos et al, 1993).
In vitro experiments have demonstrated that the B. anthracis capsule is antiphagocytic, similar to capsule function in other pathogenic bacteria. Noncapsulated bacilli are readily phagocytosed by tissue culture macrophages while capsulated bacilli are rarely phagocytosed (Keppie et al, 1953; Makino et al, 1989). In addition to its antiphagocytic properties, the γDPGA capsule is relatively nonimmunogenic (Leonard and Thorne, 1961; Schneerson et al, 2003). It has been hypothesized that the capsule enables the bacterium to avoid demise in the host because a strong humoral immune response is not generated against the outer surface of the bacteria. Interestingly, three different groups recently reported that conjugation of small glutamic acid polymers to different immunogenic molecules significantly enhances the immune response to glutamic acid (Rhie et al, 2003; Schneerson et al, 2003; Wang et al, 2004).
Small glutamic acid polypeptides may be important during anthrax infection. In vitro, the enzyme encoded by capD (formerly dep) depolymerizes large glutamic acid polymers into smaller fragments (Uchida et al, 1993b). Makino et al (2002) demonstrated that a capD-null mutant is attenuated when injected into mice intraperitoneally. Virulence is restored by coinjection of small glutamic acid polymers with the capD mutant, implicating a direct role for the homogenous, peptide fragments in vivo. The capD gene is the last gene of the capsule biosynthetic gene operon, capBCAD. The products of capB, capC, and capA are thought to mediate biosynthesis, transport, and attachment of large capsule polymers to the bacterial cell wall, although specific functions of the proteins encoded by these genes have not been demonstrated in B. anthracis.
We recently demonstrated that cap gene regulation in a pXO1+ pXO2+ strain is more complex than previously thought (Drysdale et al, 2004). Three genes encoding proteins with significant amino-acid homology play roles in positive regulation of the capsule biosynthetic gene operon. acpA and acpB are pXO2-encoded regulators with partially overlapping function. During in vitro culture, single acpA and acpB mutants synthesize capsule but a double mutant missing acpA and acpB is noncapsulated. atxA, originally identified as the anthrax toxin gene activator (Uchida et al, 1993a; Koehler et al, 1994; Dai et al, 1995), exerts its effect on cap expression via positive regulation of acpA and acpB. The molecular mechanism(s) by which these regulators activate gene transcription is not known. Epistatic relationships between the regulators and their effects on cap gene expression have been assessed in cells grown in batch culture, but the significance of the acpA and acpB genes during infection has not been determined.
Inhalational anthrax is characterized by rapid development of nonspecific, flu-like symptoms that, if left untreated, progress quickly to shock, respiratory distress, and death. Anthrax disease appears to result from a two-step process involving overwhelming bacterial replication and subsequent toxin production (Rhie et al, 2003). To investigate the precise roles of the capsule and the capsule gene regulators during infection, we assessed germination, dissemination, and virulence of mutants deleted for capsule biosynthetic and regulatory genes using a mouse model for inhalation anthrax. We show that the capsule is required for dissemination and the ability of B. anthracis to cause fatal disease.
Results
Virulence of B. anthracis capsule mutants
The virulence of isogenic B. anthracis mutants was assessed by measuring the lethal dose of spores that killed 50% of the mice (LD50) and the mean time of death (MTD) at 10-fold the LD50 (Table I). The parent strain for the study, UT500 (pXO1+ pXO2+), was constructed by transducing pXO2 into the Sterne strain as described previously (Bourgogne et al, 2003). The virulence of UT500 was comparable to the Ames strain of B. anthracis, a well-studied clinical isolate (Welkos et al, 1993; Read et al, 2003). The LD50 for UT500 was 4 × 103 spores and the MTD was 3 days, while the LD50 for Ames was 1 × 103 spores and the MTD was 2 days.
Table 1.
Strain LD50 values and MTD for infected mice
| Strain | Genotype | LD50 | MTD at 10 × LD50a |
|---|---|---|---|
| Ames | 1 × 103 | 2 days | |
| UT500 | 4 × 103 | 3 days | |
| UT538 | capBCAD | >5 × 107 | ND |
| UT526 | acpA acpB | >5 × 107 | ND |
| UT502 | acpA | 2 × 103 | 3 days |
| UT525 |
acpB |
3 × 105 |
6 days |
| aND indicates no deaths were observed using the 10 × LD50 dose. | |||
A mutant deleted for the entire capsule biosynthetic gene operon, UT538 (capBCAD), was completely attenuated; none of the mice died at the highest dose tested (5 × 107 spores). The same result was obtained for the regulatory mutant UT526 (acpA acpB). The acpA and acpB genes are partial functional homologs. During in vitro culture, the presence of either acpA or acpB is sufficient for capsule synthesis, but deletion of both genes results in noncapsulated cells (Drysdale et al, 2004). UT500 and UT502 (acpA) exhibited similar LD50 and MTD results. Surprisingly, the LD50 (3 × 105) and MTD (6 days) of UT525 (acpB) indicated that this mutant was less virulent than UT500.
Dissemination of B. anthracis to the spleen
Dissemination of B. anthracis was assessed as colony forming units (CFU) obtained from the spleens of infected mice at 3, 24, 48, and 72 h postinfection. For each experiment, groups of five mice were examined at each time point. Representative data obtained in one of three separate experiments are shown in Figure 1. Mice were infected with approximately 4 × 104 spores (10 × the LD50 of UT500) in order to compare the abilities of each strain to escape from the lungs and disseminate to the spleen. UT500 was detected in the spleens of some mice as early as 24 h after infection. At 72 h, high numbers of bacteria were found in the spleens of all the UT500-infected mice. In contrast, no CFU of UT538 (capBCAD) and UT526 (acpA acpB) were obtained from the spleens of infected mice at any time point tested. UT502 (acpA) disseminated and caused systemic infection similar to UT500. Dissemination of UT525 (acpB) occurred in few mice, relative to the parent strain, in accordance with the elevated LD50 of this mutant. In the experiment shown in Figure 1, one mouse succumbed to anthrax disease and no disseminated organisms were detected in the remaining four mice. However, in total, 15 mice were infected with UT525. Of these, two mice died of lethal anthrax disease and bacteria were found at 72 h postinfection in the spleens of three of the remaining mice.
Figure 1.
Dissemination of UT500 and isogenic mutant strains in mice. At time=0, mice were infected with 4 × 104 spores. Each bar represents data from one mouse. The y-axis is shown on a logarithmic scale and indicates CFU detected. Data shown are representative of at least three independent experiments. For UT500 and UT502 (acpA), five time points are shown to illustrate early dissemination, beginning 24 h postinfection for some mice. *One mouse died between 48 and 72 h.
We also evaluated germination of parent and mutant strains in the lungs of infected mice. The numbers of heat-resistant and heat-sensitive CFU in the lungs were determined at 3, 24, 48, and 72 h postinfection. There were no apparent germination differences between UT500 and the mutants. All of the strains germinated in the lungs. At 3 h postinfection, usually >95% of CFU detected were heat sensitive (data not shown), indicating that the inability of some mutants to cause disease is not due to an early germination problem in vivo.
The capsule phenotypes of all strains were stable. CFU isolated from infected mice displayed the original capsule phenotype associated with the strains when cultured in vitro: UT500, UT502, and UT525 were Cap+, while UT538 and UT526 were Cap− (data not shown).
Pathology
Histological examination of tissues from UT500- and UT538-infected mice revealed that at 72 h, mice infected with UT500 showed marked widespread necrotizing splenitis with hemorrhage and massive numbers of bacterial rods present (Figure 2C). The lungs from these mice were congested and exhibited mild, widespread fibrin exudation from septal capillaries. The nuclei in interalveolar septae appeared uniformly condensed, suggestive of cell death (Figure 2A). Bacterial rods were present in septal capillaries and in pulmonary arteries and veins.
Figure 2.
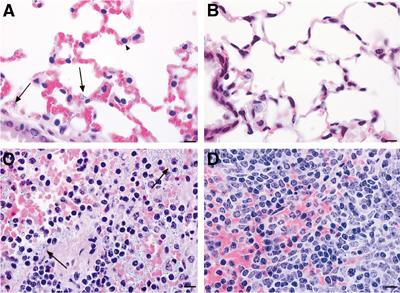
Pathology of UT500 and UT538 (capBCAD) in BALB/C mice. Lung (A, B) and spleen (C, D) from BALB/C mice 72 h after intratracheal inoculation. Tissues from UT500-infected mice are shown in (A, C). Tissues from UT538-infected mice are shown in (B, D). Note the presence of bacteria in the lung and spleen (arrows), and the condensed pulmonary septal nuclei (arrowhead) in the UT500-inoculated mice. Bar represents 10 μm.
In contrast to the UT500-infected mice, mice infected with UT538 (capBCAD) exhibited minimal pulmonary and splenic pathology at 72 h (Figure 2B and D). A few mice had a mild increase in splenic sinusoidal neutrophils. There was minimal disorganization of conducting airway epithelium, characterized by irregular spacing of epithelial cells and scattered foci of epithelial hyperchromasia and crowding, which was interpreted as regeneration. Bacteria were not seen in spleen or lung tissue from any of the UT538-infected mice.
Control of cap gene expression in vivo
B. anthracis virulence gene expression was assessed in infected mice using quantitative (Taqman) RT–PCR assays. Sensitivity testing of our assays revealed that we could detect as little as ∼100 pg of bacterial RNA when 60 000-fold more mammalian RNA was present in the same sample (data not shown). We tested for mRNA transcripts of capB, the first gene of the capsule biosynthetic gene operon, and pagA, the gene encoding the anthrax toxin protein, protective antigen, at 48 h postinfection in the lungs and spleens of mice infected with parent and mutant strains. As a control, we also tested for transcripts of the 16S rRNA gene. Transcripts of this gene are readily detected in vegetative cells of B. anthracis, but are not detected in spores. In our assay, 16S rRNA gene transcripts were not detectable in RNA isolated from 104 spores (data not shown).
All mice were infected with approximately 4 × 104 spores (10 × LD50 of UT500), as above, so that gene expression and dissemination in different strains could be compared. Data are shown in Table II. Detection of 16S rRNA gene transcripts was used as an indicator of the presence of vegetative cells in the lungs and spleens of infected mice. 16S rRNA gene transcripts were detected in lung and spleen tissue of all UT500- and UT502 (acpA)-infected mice in agreement with the dissemination experiments. In contrast, we were not able to detect 16S rRNA gene transcripts in the lungs of any UT538 (capBCAD)- or UT526 (acpA acpB)-infected mice, suggesting that the bacteria were not actively replicating. 16S rRNA gene transcripts were detected in some but not all UT525 (acpB)-infected mice. As described above, when mice were infected with UT525, only two of 15 animals died and CFU determinations revealed few mice with viable UT525 in the spleen. However, the detection of 16S rRNA gene transcripts in the spleens of four of five mice indicated that infection with UT525 resulted in dissemination in the majority of the mice. In general, the level of 16S RNA gene transcripts detected in the lungs and spleen of UT525 (acpB)-infected mice was 10- to 1000-fold lower than that observed in UT500-infected mice (data not shown). It is also important to note that the presence of 16S rRNA gene transcripts is not indicative of live vegetative cells. Considering the reduced virulence of UT525, it is likely that these organisms are cleared at a faster rate than UT500.
Table 2.
Detection of capB, pagA, and 16S rRNA gene transcripts in the lungs and spleens of infected mice at 48 h postinoculation
| Transcripts in lung |
Transcripts in spleen |
|||||
|---|---|---|---|---|---|---|
| Strain | 16S rRNA | capB | pagA | 16S rRNA | capB | pagA |
| UT500 | 4/4 | 2/4 | 2/4 | 4/4 | 3/4 | 2/4 |
| UT502 (acpA) | 4/4 | 3/4 | 3/4 | 4/4 | 3/4 | 2/4 |
| UT525 (acpB) | 2/5 | 0/5 | 0/5 | 4/5 | 0/5 | 0/5 |
| UT526 (acpA acpB) | 0/5 | 0/5 | 0/5 | NT | NT | NT |
| UT537 (capBCAD) | 0/4 | 0/4 | 0/4 | NT | NT | NT |
| NT=not tested. | ||||||
We also tested for capB and pagA transcripts to determine whether capsule and toxin gene expression occurred in the lungs and spleens of infected mice. We were able to detect capB and pagA transcripts in many UT500- and UT502 (acpA)-infected mice as expected. Neither transcript was found in the lungs of UT538 (capBCAD)- or UT526 (acpA acpB)-infected mice, almost certainly because the bacteria are not actively replicating after initial deposition in the lungs. We were also not able to detect capB or pagA transcripts in UT525 (acpB)-infected mice. Most likely, the bacteria were not present in a quantity sufficient for detection.
Capsule synthesis in vivo
The capsule of vegetative B. anthracis cells grown in vitro can be readily visualized by preparation of cells with India Ink. Capsule on the surface of the cells occludes the ink particles resulting in the appearance of a clear zone surrounding the bacilli.
To visualize capsule on B. anthracis cells grown in vivo, we added India Ink to homogenized lung and spleen tissues from mice infected with UT500 and UT538 (capBCAD). Although the capB gene was expressed by UT500 during infection, it was extremely difficult to identify capsulated organisms in tissues. Capsulated UT500 vegetative cells were only observed in tissues of mice that had recently succumbed to anthrax disease. Figure 3 shows capsulated cells in the lungs and spleen of a UT500-infected mouse. The tissues were harvested 5 min after the death of the animal. Vegetative B. anthracis cells from the lungs and spleen of mice infected with the capBCAD mutant UT538 were not visualized.
Figure 3.

Indian Ink preparations of vegetative UT500 and UT538 (capBCAD) B. anthracis in the lungs and spleen of BALB/C mice. Pictures were taken approximately 72 h postinfection.
Discussion
It has been commonly assumed that pXO1+ pXO2− B. anthracis strains are significantly reduced in virulence compared to pXO1+ pXO2+ strains because of the inability of pXO2− strains to produce the γDPGA capsule. Yet the 96-kb pXO2 plasmid contains 81 predicted ORFs in addition to the capsule biosynthesis operon. Moreover, previous experiments addressing pXO2 function in virulence have employed mouse models in which B. anthracis spores are delivered by interperitonial, subcutaneous, or intramuscular injection (Ivins et al, 1986; Welkos, 1991; Welkos et al, 1993; Makino et al, 2002). The physiological relevance of such delivery methods is questionable. Our data, derived from a newly established mouse model for inhalation anthrax (Lyons et al, 2004), demonstrate unequivocally that the capsule biosynthetic operon is essential for establishment of lethal anthrax infection. A pXO1+ pXO2+ strain deleted for only capBCAD was highly attenuated with an LD50 of greater than 5 × 107 spores. A B. anthracis mutant deleted for the cap regulators acpA and acpB was also unable to cause anthrax disease.
Our results indicate that virulence of UT500, the recombinant B. anthracis strain used in our studies, is comparable to that of the Ames strain, a clinical isolate with well-established virulence in animal models and recently associated with human cases (Read et al, 2003). UT500 was constructed by introducing pXO2 from the pXO1− pXO2+ Pasteur strain 6602 into the pXO1+ pXO2− Sterne strain 7702 (Bourgogne et al, 2003). We chose UT500 as the parent strain for our experiments because many gene regulation and animal studies have been performed with Sterne and Pasteur strain derivatives and we wished to be able to compare our results to previously published data.
The results from dissemination experiments indicate that fully virulent B. anthracis strains (UT500 and the acpA mutant UT502) escape from the lungs and replicate rapidly in the spleen. Despite the retrieval of mostly heat-sensitive bacteria from the lungs early in infection, microscopy revealed the presence of light-refractile spores in lung tissue (data not shown) and total CFU retrieved did not differ significantly from the initial infective dose. Capsulated vegetative cells were only apparent in the lung tissue at the end of the infection, following dissemination to the spleen. An increase in CFU in the lungs, again comprised mainly of heat-sensitive bacteria, was only observed shortly before death (Figure 1 and data not shown). Taken together, our findings suggest that bacteria may not be multiplying in lung tissue, but paused at an early germination stage. We hypothesize that anthrax disease results when a few bacteria escape from the lungs and then replicate rapidly to cause systemic infection. The increase in CFU in the lungs shortly before death may be due to secondary infection through the bloodstream. Inspection of a few mice that survived UT500 infection (initial dose of 104 spores) revealed no increase in lung CFU by day 14 postinfection. By day 48, the surviving mice had cleared the bacteria; no CFU were detected in the lungs or spleen (data not shown), suggesting that clearance of the bacteria from the lungs can occur but is a slow process.
Unlike UT500, the cap operon deletion mutant UT538 is unable to produce capsule under any in vitro growth condition tested. UT526, a mutant deleted for the capsule gene regulators acpA and acpB, is also unable to synthesize capsule in vitro. Both mutants were completely attenuated in the mouse model; they were not able to disseminate from the lungs to the spleen or blood even when the mice were given a large dose of spores (5 × 107 spores) (Figure 1 and data not shown). The attenuation of the capBCAD and acpA acpB mutants does not appear to be due to an early germination event. As was true for the parent strain and the acpA mutant, as early as 3 h after infection, most of the UT538 and UT526 CFU detected in the lungs of infected mice (>95%) were heat sensitive, indicating that the spores had begun to germinate (data not shown). Surprisingly, vegetative cells of the capBCAD and acpA acpB mutants were not visualized by microscopy in the lungs of infected mice 3, 48, or 72 h postinfection (Figure 3 and data not shown). Also, 16S rRNA gene, capB, and pagA transcripts could not be detected at 3, 4, or 72 h postinfection in mice infected with these mutants (Table II and data not shown). Taken together, these data indicate that the cap and acpA acpB mutants do not actively replicate in the lungs.
We also tested the ability of mice to clear the UT538 and UT526 spores from their lungs. The lungs of mice infected with approximately 3 × 107 spores of UT538 or UT526 contained average spore numbers of 5 × 104 and 1 × 105, respectively, at 48 days postinfection (data not shown). This finding is in agreement with previous reports indicating that spores remain in the lungs long after initial exposure (Henderson et al, 1956).
In the current model for capsule gene regulation, the acpA and acpB genes have partial functional similarity with regard to positive control of cap gene expression. While UT526 (acpA acpB) is noncapsulated during growth in vitro, mutants deleted for the individual genes are capsulated (Drysdale et al, 2004). Thus, we were not surprised to find that deletion of the acpA gene did not appear to affect virulence in our model. During growth in vitro, the acpA mutant UT502 produces capsule and toxin similar to UT500, and transcriptional profiling experiments have shown that acpA has only a two-fold effect on cap gene expression (Bourgogne et al, 2003). In contrast, the reduced virulence of the acpB mutant UT525 (acpB) was unexpected because, like UT502, this mutant produces capsule and toxin during growth in vitro (Drysdale et al, 2004). The predicted products of the capsule gene regulators, AcpA and AcpB, bear significant amino-acid homology (∼62%) to each other and to AtxA (∼50%), the global regulator of B. anthracis virulence genes. While partial functional redundancy between acpA and acpB during growth in vitro has been documented (Drysdale et al, 2004), clearly the functions of acpA and acpB during infection do not overlap completely. Our data indicate that in addition to controlling capBCAD, acpB regulates other genes that affect virulence in the mouse model. Future transcriptional profiling experiments will address this hypothesis.
There is renewed interest in the B. anthracis capsule with regard to immunoprotection. Although the γDPGA capsule itself appears to be relatively nonimmunogenic, a number of investigators have shown that glutamic acid polymers conjugated to various carriers can generate an immune response to glutamic acid (Rhie et al, 2003; Schneerson et al, 2003; Wang et al, 2004). Recently, Kozel et al (2004) demonstrated that a strong immune response is elicited against D-glutamic acid polymers when mice are coimmunized with agonist monoclonal antibodies (mAb) to CD40. These investigators produced mAbs specific for γDPGA and used the γDPGA mAb to detect soluble poly-glutamic acid in the serum of B. anthracis-infected mice. Moreover, the γDPGA mAb passively protected mice against lethal anthrax disease. These results reveal the potential of the B. anthracis capsule as a target for detection, treatment, and/or prevention of anthrax disease.
Our work demonstrates that the unique poly-D-glutamic acid capsule of B. anthracis is vital for dissemination from the lungs and hence persistence and survival in a mouse model for inhalation anthrax. The well-characterized and genetically complete parent strain, isogenic mutants, and murine model as used in our experiments provide a good starting point for evaluating the effect of specific B. anthracis genes on virulence. By employing mice, we were able to use the number of animals required for LD50 determinations for multiple B. anthracis mutants. In addition, the inbred nature of the mice removed potential quantitative trait differences in host factors that can influence host–pathogen interactions and lead to difficulty in data interpretation. Finally, the tools that are readily available for dissecting the murine host response also make this an attractive model for further studies evaluating B. anthracis–host interactions.
Materials and methods
Bacterial strains
Table III lists strains used in this study. Constructions of UT500, UT502 (acpA), UT525 (acpB), and UT526 (acpA acpB) have been described (Bourgogne et al, 2003; Drysdale et al, 2004). To construct the cap mutant, capBCAD coding sequences (from 3 bp upstream of capB to 43 bp downstream of capD) were replaced with an Ω-spectinomycin cassette using a method reported previously (Saile and Koehler, 2002). The gene replacement was constructed initially in a UM23C1-1td10 strain (Green et al, 1985) background. The capsule phenotypes of UM23C1-1td10 (Cap+), the capBCAD-null mutant (Cap−), and the mutant complemented with capBCAD in trans (Cap+) are shown in Supplementary data. The capBCDA-null mutation was subsequently transduced (Green et al, 1985) to UT500 with selection for Spcr isolates. Gene replacement was confirmed using the PCR. This strain was named UT538.
Table 3.
Strains used in this study; all strains are pXO1+ pXO2+
| Strain | Relevant characteristics | Reference |
|---|---|---|
| Ames | Virulent in ruminants rodents and humans | Read et al (2003) |
| UT500 | pXO2 from 6602 transduced into 7702 | Bourgogne et al (2003) |
| UT502 | acpA-null derivative of UT500 | Drysdale et al (2004) |
| UT525 | acpB-null derivative of UT500 | Drysdale et al (2004) |
| UT526 | acpA acpB-null derivative of UT500 | Drysdale et al (2004) |
| UT538 | capBCAD-null derivative of UT500 | This work |
Mouse infection
Female, 6-week old, BALB/C mice were purchased from Harlan Sprague Dawley (Madison, WI) and maintained as described previously (Lyons et al, 2004). Tracheal infections were performed under ABSL 3 containment. Mice were anesthetized with 400 μl avertin administered intraperitoneally. A small incision was made in the skin over the trachea and the salivary gland separated to expose the trachea. Sterile polypropylene tubing attached at one end to a bent 30-gauge needle and at the other end to a 1-ml syringe was used to inject 50-μl spore solutions. Incisions were closed using Super Glue (Manco Inc., Avon, OH). At 1 h after infection, two random mice were killed and the lungs were harvested to determine spore deposition.
LD50 determinations
For a single spore dose, groups of 10 mice were infected as described above. Doses ranged from 1 × 102 to 5 × 107 spores. LD50 values were calculated using the method described by Reed and Muench (1938).
B. anthracis germination and dissemination
For germination and dissemination experiments, mice were infected with approximately 4 × 104 spores (10 × LD50 of UT500). Germination was assessed by determining the numbers of heat-resistant and heat-sensitive CFU in the lungs of mice at 3, 48, and 72 h postinfection. The lungs were harvested from mice and placed in a microcentrifuge tube containing 1 ml of sterile phosphate-buffered saline (PBS) and 250 μl of 2.5-μm Silica/Zirconia beads (Biospec Products, Bartlesville, OK). Samples were homogenized for 90 s in a bead beater (Biospec Products). Tissue homogenates were divided in half. A 50 μl portion of one half was plated immediately onto Tryptic Soy Agar with 5% sheep blood (Remel, Lexena, KS) using an Autoplate 4000 (Spiral Biotech, Bethesda, MD). The other half was placed at 68°C for 40 min prior to plating. For some tissue samples, dilutions of the tissue homogenate were made in PBS prior to plating in order to obtain 20–200 CFU per plate. Colonies were enumerated using Qcount (Sprial Biotech). Dissemination was assessed by measuring CFU in the spleens of mice at 3, 48, and 72 h postinfection. The procedure for harvesting spleens was identical to that used for harvesting lungs.
Pathology
The lungs are removed from the thorax en bloc and inflated with 10% neutral buffered formalin via a tracheal cannula. The lungs and other tissues were fixed for 24–72 h and subsequently trimmed for paraffin embedding. The lungs were trimmed along the edges of the left main-stem bronchus, and the right cranial, middle, and caudal lobar bronchi. Paraffin-embedded tissues were sectioned at 5 μm and stained with hematoxylin and eosin for histology.
RNA purification
Mice were infected with approximately 4 × 104 spores and RNA was extracted from the lungs and spleens at 3 or 48 h postinfection. The lungs and spleens were harvested and placed immediately into 15-ml conical tubes containing 5 ml (for lungs) or 3 ml (for spleens) of prechilled Tri-Reagent (Molecular Research Center, Cincinnati, OH). Lung and spleen tissue was homogenized for 20 and 15 s, respectively, using a Pro200 Homogenizer (Proscientific, Oxford, CN). A measure of 1 and 0.6 ml of chloroform (Sigma Aldrich, St Louis, MO) was then added to the lung and spleen samples, respectively. The samples were vortexed for 30 s, placed at room temperature for 10 min, and then centrifuged at 3219 g for 20 min at 4°C. The top aqueous layer was removed and placed in a new conical tube. An equal volume of isopropanol was added to each sample. Tubes were inverted 10 times, placed at room temperature for 10 min, and then centrifuged as previously for 15 min. The isopropanol was removed and 1 ml of 75% ethanol was added to the RNA pellet. The pellet and ethanol were transferred to a 2-ml microcentrifuge tube and centrifuged at 16 000 g for 15 min. The ethanol was removed and the pellet air-dried for 10 min. The RNA pellet was resuspended in 100 μl of nuclease-free water and the RNA was further purified using the protocol and reagents of the Ribopure Bacteria Kit (Ambion, Austin, TX). DNase treatment of purified RNA was performed prior to Q-RT–PCR analysis using the DNase-Free Kit (Ambion, Austin, TX) according to the protocol of the supplier.
Quantitative real-time RT–PCR
Q-RT–PCR assays were performed using a 7700 Sequence Detector (Applied Biosystems) and a protocol described previously (Drysdale et al, 2004). Primer and probe sequences for the capB gene were as described (Drysdale et al, 2004). Primer sequences for pagA were (+)TCGTTCTTTGATATTGGTG (−)ATTGCGACCGTACTTG. The probe sequence for pagA was (5′FAM) AGTGTATCTGCAGGATTTAGTAATTCGA. Primer sequences for the 16S rRNA gene were (+)TTCGGGAGCAGAGTG (−)AACATCTCACGACACGAG. The probe sequence for the 16S rRNA gene was (5′FAM) CAGGTGGTGCATGGTTGTC.
Microscopy
Tissue samples were bead-beaten for 1 min in 1 ml of sterile PBS. Homogenized tissue samples were placed on glass microscope slides and coverslips were placed on top of the samples. India Ink (Becton Dickinson Microbiology Systems, Sparks, Md.; undiluted or diluted to 70% in sterile PBS) was added to the edges of the coverslips. Using an Olympus BH-2 microscope (Olympus, Orangeburg, NY), the capsule was visualized as the exclusion of ink particles around cells.
Supplementary Material
Supplementary Data
Acknowledgments
We thank Lucy Berliba, Kristin Garrison, Sonja Dean, and Amy Courtney for expert technical assistance. The assays, reactions, and initial data analysis for real-time RT–PCR were performed by the Quantitative Genomics Core Laboratory in the Department of Integrative Biology and Pharmacology at the University of Texas Health Sciences Center—Houston. This work was supported by Public Health Service grants AI33537 (TMK) and U54 AI057156 (TMK and CRL) from the National Institutes of Health and by Defense Advanced Research Projects Agency Contract N00178-01-C-3069 (CRL) from the Department of Defense.
References
- Arantes O, Lereclus D (1991) Construction of cloning vectors for Bacillus thuringiensis. Gene 108: 115–119 [DOI] [PubMed] [Google Scholar]
- Bourgogne A, Drysdale M, Hilsenbeck SG, Peterson SN, Koehler TM (2003) Global effects of virulence gene regulators in a Bacillus anthracis strain with both virulence plasmids. Infect Immun 71: 2736–2743 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y, Tenover FC, Koehler TM (2004) β-lactamase gene expression in a penicillin-resistant Bacillus anthracis strain. Antimicrob Agents Chemother 48: 4873–4877 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dai Z, Sirard JC, Mock M, Koehler TM (1995) The atxA gene product activates transcription of the anthrax toxin genes and is essential for virulence. Mol Microbiol 16: 1171–1181 [DOI] [PubMed] [Google Scholar]
- Drysdale M, Bourgogne A, Hilsenbeck SG, Koehler TM (2004) atxA controls Bacillus anthracis capsule synthesis via acpA and a newly discovered regulator, acpB. J Bacteriol 186: 307–315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Green BD, Battisti L, Koehler TM, Thorne CB, Ivins BE (1985) Demonstration of a capsule plasmid in Bacillus anthracis. Infect Immun 49: 291–297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Henderson DW, Peacock S, Belton FC (1956) Observations on the prophylaxis of experimental pulmonary anthrax in the monkey. J Hyg (Lond) 54: 28–36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivins BE, Ezzell JW Jr, Jemski J, Hedlund KW, Ristroph JD, Leppla SH (1986) Immunization studies with attenuated strains of Bacillus anthracis. Infect Immun 52: 454–458 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keppie J, Smith H, Harris-Smith W (1953) The chemical basis of the virulence of Bacillus anthracis. II. Some biological properties of bacterial products. Br J Exp Pathol 34: 486–496 [PMC free article] [PubMed] [Google Scholar]
- Koehler TM, Dai Z, Kaufman-Yarbray M (1994) Regulation of the Bacillus anthracis protective antigen gene: CO2 and a trans-acting element activate transcription from one of two promoters. J Bacteriol 176: 586–595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kozel TR, Murphy WJ, Brandt S, Blazar BR, Lovchik JA, Thorkildson P, Percival A, Lyons CR (2004) mAbs to Bacillus anthracis capsular antigen for immunoprotection in anthrax and detection of antigenemia. Proc Natl Acad Sci USA 101: 5042–5047 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leonard CG, Thorne CB (1961) Studies on the nonspecific precipitation of basic serum proteins with poly-gamma-glutamyl polypeptides. J Immunol 87: 175–188 [PubMed] [Google Scholar]
- Lyons CR, Lovchik JA, Hutt J, Lipsomb MF, Wang E, Heninger S, Berliba L, Garrison K (2004) Murine model of pulmonary anthrax: kinetics of dissemination, histopathology and mouse strain susceptibility. Infect Immun 72: 4801–4809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makino S, Uchida I, Terakado N, Sasakawa C, Yoshikawa M (1989) Molecular characterization and protein analysis of the cap region, which is essential for encapsulation in Bacillus anthracis. J Bacteriol 171: 722–730 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Makino S, Watarai M, Cheun HI, Shirahata T, Uchida I (2002) Effect of the lower molecular capsule released from the cell surface of Bacillus anthracis on the pathogenesis of anthrax. J Infect Dis 186: 227–233 [DOI] [PubMed] [Google Scholar]
- Mock M, Labruyere E, Glaser P, Danchin A, Ullmann A (1988) Cloning and expression of the calmodulin-sensitive Bacillus anthracis adenylate cyclase in Escherichia coli. Gene 64: 277–284 [DOI] [PubMed] [Google Scholar]
- Read TD, Peterson SN, Tourasse N, Baillie LW, Paulsen IT, Nelson KE, Tettelin H, Fouts DE, Eisen JA, Gill SR, Holtzapple EK, Okstad OA, Helgason E, Rilstone J, Wu M, Kolonay JF, Beanan MJ, Dodson RJ, Brinkac LM, Gwinn M, DeBoy RT, Madpu R, Daugherty SC, Durkin AS, Haft DH, Nelson WC, Peterson JD, Pop M, Khouri HM, Radune D, Benton JL, Mahamoud Y, Jiang L, Hance IR, Weidman JF, Berry KJ, Plaut RD, Wolf AM, Watkins KL, Nierman WC, Hazen A, Cline R, Redmond C, Thwaite JE, White O, Salzberg SL, Thomason B, Friedlander AM, Koehler TM, Hanna PC, Kolsto AB, Fraser CM (2003) The genome sequence of Bacillus anthracis Ames and comparison to closely related bacteria. Nature 423: 81–86 [DOI] [PubMed] [Google Scholar]
- Record BF, Wallis RG (1955) Physiochemical examination of polyglutamic acid from Bacillus anthracis grown in vivo. Biochem J 63: 443–447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed LJ, Muench H (1938) A simple method of estimating fifty per cent endpoints. Am J Hyg 27: 493–497 [Google Scholar]
- Rhie GE, Roehrl MH, Mourez M, Collier RJ, Mekalanos JJ, Wang JY (2003) A dually active anthrax vaccine that confers protection against both bacilli and toxins. Proc Natl Acad Sci USA 100: 10925–10930 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robertson DL, Leppla SH (1986) Molecular cloning and expression in Escherichia coli of the lethal factor gene of Bacillus anthracis. Gene 44: 71–78 [DOI] [PubMed] [Google Scholar]
- Robertson DL, Tippetts MT, Leppla SH (1988) Nucleotide sequence of the Bacillus anthracis edema factor gene (cya): a calmodulin-dependent adenylate cyclase. Gene 73: 363–371 [DOI] [PubMed] [Google Scholar]
- Saile E, Koehler TM (2002) Control of anthrax toxin gene expression by the transition state regulator abrB. J Bacteriol 184: 370–380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schneerson R, Kubler-Kielb J, Liu TY, Dai ZD, Leppla SH, Yergey A, Backlund P, Shiloach J, Majadly F, Robbins JB (2003) Poly(gamma-D-glutamic acid) protein conjugates induce IgG antibodies in mice to the capsule of Bacillus anthracis: a potential addition to the anthrax vaccine. Proc Natl Acad Sci USA 100: 8945–8950 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thorne CB (1993) Bacillus anthracis. In Bacillus subtilis and Other Gram Positive Bacteria: Biochemistry, Physiology, and Molecular Genetics, Sonenshein AL, Hoch JA, Losick R (eds) pp 113–124. Washington, DC: American Society for Microbiology [Google Scholar]
- Tippetts MT, Robertson DL (1988) Molecular cloning and expression of the Bacillus anthracis edema factor toxin gene: a calmodulin-dependent adenylate cyclase. J Bacteriol 170: 2263–2266 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchida I, Hornung JM, Thorne CB, Klimpel KR, Leppla SH (1993a) Cloning and characterization of a gene whose product is a trans-activator of anthrax toxin synthesis. J Bacteriol 175: 5329–5338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Uchida I, Makino S, Sasakawa C, Yoshikawa M, Sugimoto C, Terakado N (1993b) Identification of a novel gene, dep, associated with depolymerization of the capsular polymer in Bacillus anthracis. Mol Microbiol 9: 487–496 [DOI] [PubMed] [Google Scholar]
- Vietri NJ, Marrero R, Hoover TA, Welkos SL (1995) Identification and characterization of a trans-activator involved in the regulation of encapsulation by Bacillus anthracis. Gene 152: 1–9 [DOI] [PubMed] [Google Scholar]
- Vodkin MH, Leppla SH (1983) Cloning of the protective antigen gene of Bacillus anthracis. Cell 34: 693–697 [DOI] [PubMed] [Google Scholar]
- Wang TT, Fellows PF, Leighton TJ, Lucas AH (2004) Induction of opsonic antibodies to the gamma-D-glutamic acid capsule of Bacillus anthracis by immunization with a synthetic peptide-carrier protein conjugate. FEMS Immunol Med Microbiol 40: 231–237 [DOI] [PubMed] [Google Scholar]
- Welkos SL (1991) Plasmid-associated virulence factors of non-toxigenic (pX01−) Bacillus anthracis. Microb Pathogenesis 10: 183–198 [DOI] [PubMed] [Google Scholar]
- Welkos SL, Lowe JR, Eden-McCutchan F, Vodkin M, Leppla SH, Schmidt JJ (1988) Sequence and analysis of the DNA encoding protective antigen of Bacillus anthracis. Gene 69: 287–300 [DOI] [PubMed] [Google Scholar]
- Welkos SL, Vietri NJ, Gibbs PH (1993) Non-toxigenic derivatives of the Ames strain of Bacillus anthracis are fully virulent for mice: role of plasmid pX02 and chromosome in strain-dependent virulence. Microb Pathogenesis 14: 381–388 [DOI] [PubMed] [Google Scholar]
- Xu Y, Liang X, Chen Y, Koehler TM, Hook M (2004) Identification and biochemical characterization of two novel collagen binding MSCRAMMs of Bacillus anthracis. J Biol Chem 279: 51760–51768 [DOI] [PubMed] [Google Scholar]
- Zwartouw HT, Smith H (1956) Polyglutamic acid from Bacillus anthracis grown in vivo; structure and aggressin activity. Biochem J 63: 437–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Data