

Abstract
Background
The Asian tiger mosquito (Aedes albopictus) is an invasive species and important arbovirus vector that was introduced into the U.S. in the 1980's where it continues to expand its range. Winter temperature is an important constraint to its northward expansion, with potential range limits located between the 0° and -5°C mean cold month isotherm. Connecticut is located within this climatic zone and therefore, Ae. albopictus was monitored statewide to assess its northern range expansion and to delineate where populations can stably persist.
Methodology/Principal findings
Ae. albopictus females were monitored at fixed trapping sites throughout Connecticut from June-October over a 20-year period, 1997–2016. In addition, Ae. albopictus larvae and pupae were collected from tire habitats and tires were retrieved from the field in the spring and flooded to evaluate overwintering success of hatching larvae. Ae. albopictus was first detected during statewide surveillance when a single adult female was collected in 2006. This species was not collected again until 2010 and was subsequently detected each successive year with increasing abundance and distribution except following the unusually cold winters of 2014 and 2015. Ae. albopictus mosquitoes were most abundant in urban and suburban locations along the southwestern shoreline of Connecticut; however, single specimens were occasionally detected in central parts of the state. Field-collected females were also screened for arbovirus infection yielding two isolations of Cache Valley virus and one isolation of West Nile virus, highlighting the threat posed by this mosquito. Ae. albopictus overwintered in Connecticut under mild winter conditions as shown by recovery of hatched larvae from field collected tires in spring and by early season detection of larvae and pupae.
Conclusions/Significance
This study documents the establishment and expansion of Ae. albopictus at the northern boundary of its range in the northeastern U.S. and provides a baseline for monitoring the future spread of this species anticipated under climate change.
Author summary
The Asian tiger mosquito (Aedes albopictus) is a highly invasive species and an important disease vector that is undergoing rapid range expansion in many countries including the U.S. Winter temperature is an important limit to its northward expansion with Connecticut situated near the northern boundary of its potential range. In this study, we sampled mosquitoes at fixed trapping sites located statewide to track the establishment and range expansion of Ae. albopictus in this region. In addition, mosquito larvae were monitored in tire habitats to evaluate overwintering success of local populations. From this effort, we describe the initial detection of Ae. albopictus in 2006, its annual reemergence and population expansion in southwestern Connecticut from 2010–2016, and its local overwintering under mild winter conditions. Together, this study documents population changes in Ae. albopictus at the northern boundary of its range and provides a baseline for monitoring future range expansion anticipated under climate change.
Introduction
The Asian tiger mosquito, Aedes albopictus (Skuse), is an invasive species that has spread from East Asia to the Americas, Africa, Europe, and the Middle East primarily by the global tire trade [1, 2]. This mosquito inhabits urbanized environments [3], develops in artificial, water-holding containers [4], and feeds readily on humans, making it a major pest species throughout its range [5–7]. Ae. albopictus is also an important vector of dengue virus (DENV), chikungunya virus (CHIKV), and Zika virus (ZIKV) in many parts of the world. This includes its primary role in outbreaks of DENV in China [8], CHIKV in the Indian Ocean Islands [9], ZIKV in central Africa [10], and autochthonous transmission of DENV and CHIKV in southern Europe [11–13]. In addition, Ae. albopictus is competent to transmit 23 other arboviruses [14]. This includes eastern equine encephalitis virus, West Nile virus (WNV), and La Crosse virus that have also been isolated from this species during field investigations in the U.S. [3, 15, 16].
In the continental U.S., the first established population of Ae. albopictus was documented in Houston, Texas in 1985 [17], followed by its rapid expansion throughout southeastern and parts of northeastern and northcentral U.S. [2, 18]. Ae. albopictus has been occasionally collected in locations as far north as Minnesota, New Hampshire, and Ontario, Canada, but these records most likely represent seasonal introductions rather than permanent resident populations [19, 20]. The northern distribution limit for overwintering populations was conservatively estimated to be at the 0°C cold-month isotherm based on its distribution in Asia [21]. The southern coast of Connecticut is at this thermal limit of Ae. albopictus and possesses suitable habitat for colonization by this species. In 2003, Ae. albopictus (a single female) was first collected during field studies in southern Connecticut but was not further detected that same year during statewide mosquito surveillance [22]. An infestation of this species was then discovered at a tire recycling plant in northeastern Connecticut in 2006, yet the population failed to overwinter and establish at the site [23]. Clearly, continued monitoring for this species is warranted in this region, particularly in urban and suburban locations and under milder winter conditions that are anticipated to increase in frequency due to climate change.
In this study, we document the northern range expansion of Ae. albopictus by continuous monitoring of mosquito populations over a 20 year period in Connecticut. Mosquitoes were collected at 91 fixed trapping locations from June-October, identified to the species level, and tested for arbovirus infection as a part of the statewide mosquito arbovirus surveillance program. Mosquito larvae and pupae were also monitored in a used tire habitat in Stratford, (Fairfield County) Connecticut at a known locality for Ae. albopictus reproduction. Tires were also retrieved from the field in the spring and flooded to evaluate overwintering success of Ae. albopictus. From this effort, we describe the initial detection of Ae. albopictus in 2006, its annual reemergence and population expansion in the state from 2010–2016, and its local overwintering under mild winter conditions.
Methods
Adult sampling
Mosquitoes were collected at 91 trapping locations statewide (Figs 1 and 2) as a part of the Connecticut Mosquito and Arbovirus Surveillance Program from June through October [24]. Thirty-six of the sites have been monitored since 1997 with the balance of sites in continuous operation since 2000. About half of the sites are located in surburban and urban locations, primarily in the southwestern and central parts of the state, which are focal areas for WNV. Specific trapping locales include neighborhood parks, school grounds, sewage treatment plants, municipal dumping stations, industrial parks, and fragmented woodlots. The remaining sites are located in more rural settings that are associated with eastern equine encephalitis virus transmission and include permanent fresh-water swamps and bogs, coastal salt marshes, and mixed woodlands.
Fig 1. County map of Connecticut showing geographic distribution of mosquito trapping sites and land use characteristics.

Black circles indicate locations where Ae. albopictus were collected from 2006–2016 and are scaled according to numbers collected at each site. Red arrow indicates location of the Stratford site. Inset map of northeastern U.S. shows approximate location of the 0°C and -5°C cold month isotherm.
Fig 2. Connecticut map showing geographic locations where Ae. albopictus were collected by year.

Open circles indicate location of all mosquito trapping sites and black circles depict trapping sites positive for Ae. albopictus. Number of Ae. albopictus collected is indicated for each year.
Each trapping site was sampled on average weekly and at least once every 10 days by setting a CO2-baited CDC light trap and a gravid trap (John W. Hock Co., Gainesville, FL) baited with a hay/yeast/lactalbumin infusion [25]. In addition, BG Sentinel (BGS) traps baited with the BG–Lure but without CO2 (Biogents AG, Regensburg, Germany) were added after the detection of Ae. albopictus at a site. Traps were operated overnight and adult mosquitoes were transported back to the laboratory alive in an ice chest. Mosquitoes were immobilized with dry ice, sorted, and identified to species on chill tables with the aid of a stereo microscope and taxonomic keys [22]. Female mosquitoes were combined into pools of 50 or fewer individuals by location, trap type, species, and collection date in 2 mL microcentrifuge tubes containing a copper BB, and stored at -80°C until virus testing.
Virus detection
Mosquito pools were prepared for virus isolation by adding 1 mL of PBS-G (phosphate buffered saline, 30% heat-inactivated rabbit serum, 0.5% gelatin, and antibiotic/antimycotic) to each tube. Mosquito pools were homogenized using the MM300 Mixer Mill (Retsch Laboratory, Hann, Germany) set for 4 min at 25 cycles/sec. Mosquito homogenates were centrifuged for 6 minutes at 7,000 rpm at 4°C and 100 μL of the supernatant was inoculated into Vero cell cultures- clone E6 (provided by Shirley Tirrell, Yale University) growing in 25 cm2 flasks. Vero cells were maintained at 37°C and 5% CO2 and examined daily for cytopathic effect from days 3–7 post-inoculation. RNA was extracted from infected cell supernatants and the corresponding mosquito pool using the viral RNA kit (Qiagen, Valencia, CA). Virus isolates were identified as WNV or Cache Valley virus (CVV) using PCR-based assays as previously described [26, 27].
Larval surveys
From 2013–2016, larvae and pupae were sampled from a location in Stratford, Connecticut. This site is a small woodlot, located in a mixed residential and commercial neighborhood with an abandoned pile of about 30 waste tires concealed in the woods. The majority of the tires at the pile were not available for oviposition and larval development because they were buried or completely filled with organic material. Seven tires were flagged, positioned upright, and sampled weekly from May-October. Water samples were removed from tires with a small plastic container and fresh water was added back to maintain water levels in the tires. Mosquito larvae were identified using taxonomic keys under a compound microscope and pupae were allowed to emerge as adults prior to their identification.
To evaluate mosquito overwintering from 2013–2016, four tires were retrieved from the Stratford site in late April, and residual water and sediments were removed and inspected for larvae. The tires were then placed in a greenhouse, exposed to natural light and photoperiod, oriented in the same position as in the field, and flooded with water. Tires were inspected daily and larvae were identified as described above. The tires were then emptied and re-flooded a second time after a 2–3 week period when larvae were no longer observed, to evaluate if there was a delayed hatch.
Data analysis
To assess the most effective method for collecting Ae. albopictus, we compared numbers of females collected in CDC-light, BGS, and gravid traps. Trap evaluations were performed only when all three trap types were in operation from 2010–2016 (n = 474 trap-nights). The mean number of Ae. albopictus collected per trap night was calculated for each trap type and compared by Krustal-Wallis, one-way analysis of variance (ANOVA) of ranks (Prism 7.0, GraphPad software). Dunn's multiple comparison post-tests were then applied to identify which pairs of trap types significantly differed from each other.
Daily weather summaries were obtained from the National Oceanic and Atmospheric Administration National Centers for Environmental Information (https://www.ncdc.noaa.gov/cdo-web/datatools/selectlocation) for four locations in coastal Fairfield and New Haven counties: Bridgeport, Success Hill (41.200°N, 73.157°W), Stamford, 5N (41.125°N, 73.548°W), Stratford, Sikorsky Airport (41.158°N, 73.129°W), and New Haven, Tweed Airport (41.264°N, 72.887°W). Mean monthly temperatures were calculated for the winter months (Fig 3), and the lowest temperature recorded at each site per month was noted.
Fig 3. Mean monthly temperatures recorded at weather stations in Bridgeport, New Haven, Stamford, and Stratford, Connecticut.

Results
Mosquito collection and virus detection
Figs 1 and 2 depict the spatial-temporal distribution and abundance of female Ae. albopictus collected during statewide mosquito trapping efforts in Connecticut. The first female Ae. albopictus was collected from a gravid trap located in New Haven County during 2006, with no additional collections until 2010 (N = 2) and 2011 (N = 12). The number of Ae. albopictus increased substantially during 2012 (N = 245, 11 locations) and 2013 (N = 547, 11 locations). During these years, positive sites were located in southwestern coastal Connecticut with the exception of one adult female collected in the central part of the state. Ae. albopictus collections decreased during 2014 (N = 133, 9 locations) and 2015 (N = 220, 10 locations) following winters with mean monthly temperatures well below the 0°C threshold and then increased sharply during 2016 (N = 936, 24 locations) following a more moderate winter (Fig 3). Ae. albopictus were detected primarily in densely-populated communities along the southwestern portion of the state; however, single specimens were also collected from a few sites in central Connecticut.
To evaluate the effectiveness of commonly implemented mosquito traps and methodologies, we compared Ae.albopictus collections for each trap configuration used in this study. The mean number of Ae. albopictus collected per trap night differed among the BGS trap with BG Lure (mean = 1.6/trap night), CDC light trap with CO2 (mean = 1.1/trap night) and gravid trap with hay infusion (mean = 0.4/trap night) (Krustal-Wallis ANOVA p<0.0001). Pairwise comparisons revealed significant differences in trapping efficiency between the BGS trap and gravid trap (Dunn's post-test p<0.0001) and the CDC-light trap and gravid trap (p<0.01), but not between the BGS and CDC-light traps (p = 0.7).
Female Ae. albopictus were processed and screened for arbovirus infection by Vero cell culture assay. There were two isolations of CVV from Ae. albopictus collected in Stratford in 2014 and one isolation of WNV from Bridgeport in 2016. WNV infection was then reconfirmed by directly testing the positive mosquito pool by real-time RT-PCR (Ct = 22.9).
Seasonal abundance
Ae. albopictus larvae and pupae were sampled weekly from tire habitats in Stratford, CT and compared to adult trap collections from the same site (Fig 4). Larvae and pupae were collected much earlier in the season during 2013 following a mild winter than in subsequent years (Fig 3). During 2013, the numbers of juvenile Ae. albopictus fluctuated throughout the season with peak abundances occurring during early May, August, and September. Adult collections lagged behind the larval- and pupal-cohorts with peaks occurring in early and late September. During 2014 and 2015, Ae. albopictus immatures and adults closely tracked each other, appearing later in the season and in fewer numbers than during 2013. Ae. albopictus populations rebounded during 2016; however, larvae appeared later in the season starting in July and peaked in late August to early September in parallel with adult collections.
Fig 4. Weekly collection of juvenile (larvae and pupae) and adult (females) Ae. albopictus from Stratford, CT.
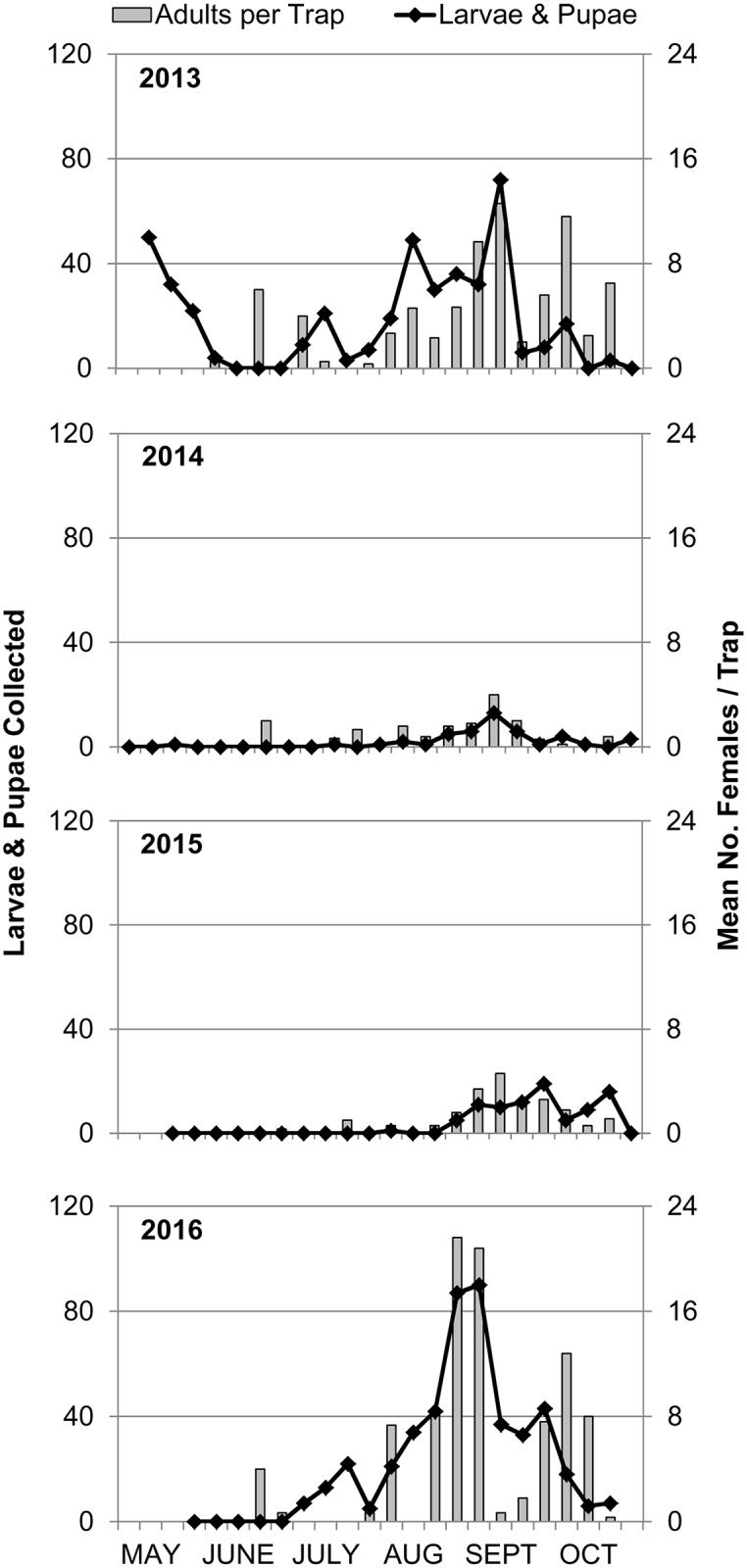
Evaluation of mosquito overwintering
To determine whether Ae. albopictus may survive winters in Connecticut, we retrieved four tires from the Stratford site in April and flooded them twice with water to recover hatched larvae. A total of 51 Ae. albopictus larvae were recovered during 2013, but none were collected from tires retrieved from the site in late April during 2014, 2015, or 2016. Other mosquito species hatching from the tires included Aedes japonicus (Theobald) (2013 N = 39 and 2014 N = 5) and Aedes triseriatus (Coquillett) (2013 N = 60 and 2014 N = 22); these species were not enumerated during 2015 and 2016.
Discussion
This study documents the establishment and expansion of Ae. albopictus at the northern boundary of its range in southern New England. This species was first detected during statewide mosquito surveillance in 2006 and then reemerged every summer starting in 2010. Ae. albopictus mosquitoes were most abundant in urban and suburban locations along the shoreline of southwestern Connecticut; however, single specimens were occasionally detected in central parts of the state. Mosquito abundance and the number of positive traps increased every year starting in 2010 except following the cold winters of 2013–2014 and 2014–2015. These changes in abundance and distribution are not an artifact of sampling effort because mosquito populations were consistently sampled at fixed trapping sites over a 20 year period. We conclude that Ae. albopictus is expanding northward in the northeastern U.S. and this trend is anticipated to accelerate under conditions of climate change as previously noted [28, 29].
We also showed that Ae. albopictus was able to overwinter in Connecticut during 2012–2013. Local overwintering was demonstrated by recovery of larvae hatching in late April from tires that were left outside during winter and by early seasonal detection of Ae. albopictus larvae and pupae. This finding, however, could not be reproduced during the following two winters that were exceptionally cold (January and February mean temperatures -2.2 to -7.2°C) or following a more mild winter during 2015–2016 (Fig 3). The performance of overwintering populations is affected by the number diapause-conditioned eggs laid during the previous fall, and overwintering temperatures and conditions impacting egg mortality. The lack of evidence for overwintering success observed in the spring of 2016 following a warmer winter may be explained, in part, by a lower abundance of females in the previous fall, leading to a lower number of diapaused eggs deposited for overwintering. It is also noteworthy that temperatures dropped to an absolute low of -21°C during a cold snap in February 2016 despite warmer mean temperatures for the entire month as shown in Fig 3. This is well below the 50% survival threshold estimated for diapausing Ae. albopictus eggs which ranges from -5 to -13°C for a 24 hour exposure period and approaches the -26°C supercooling point that is 100% lethal under brief exposures [30, 31].
The primary mechanism driving the annual reemergence of Ae. albopictus in Connecticut is unknown, but may be due to local overwintering of mosquito eggs, annual reintroduction of mosquitoes from southern locations, or some combination of both of these processes. Environmental conditions appeared to be lethal for diapausing eggs during three of the four years of this study based on recovery of Ae. albopictus from overwintering tires during the spring. Nevertheless, we cannot preclude the possibility that this species may survive at low levels in microhabitats that were not represented in our study. Ae. albopictus eggs were shown to survive winter temperatures as low as -19°C in northern Indiana, presumably due to the insulating effects of snow cover [32]. Snow drifts, sheltered structures, and urban heat transfers may enhance overwintering survival of this species at the northern distribution limit. Further studies on the field survival of diapause eggs under different microhabitat conditions and temperature regimes are needed to better understand the conditions affecting its overwintering success.
Ae. albopictus is readily dispersed over long distances by the transport of dormant eggs and emergent larvae and pupae in tires and other water-holding containers [33, 34]. This may be an important mechanism for reestablishing populations after severe winter conditions. Potential reintroduction would most likely be human mediated as Ae. albopictus is a short distance disperser where most females remain within 300 m of the larval habitat from which they emerge [35]. The observation of late season emergence of Ae. albopictus in our study sites may reflect either reintroduction of mosquitoes from more southerly locations or perhaps low levels of mosquito overwintering and hatching in the spring that could not be readily detected until later in the summer when the populations rebounded. We cannot distinguish from these possibilities based on our current data but show direct evidence for mosquito overwintering at least under mild winter conditions.
A number of studies have estimated the geographic distribution of Ae. albopictus by correlating current thermal conditions with mosquito distribution records and using this data to develop spatial models to predict its potential range. Our findings are in close agreement with predicted northern range limits along the southern margin of Connecticut, Rhode Island, and Massachusetts based on mean annual and winter temperatures [21, 28, 36]. Other studies show the distribution limit extending further north well into New England, including most of Connecticut [37, 38], yet we did not find stable populations in the interior part of the state. Mean winter temperature was identified as the most significant environmental factor predicting its current range within northeastern U.S. [28], with thermal limits estimated between the 0°C and -5°C cold month isotherm [21, 36]. The observed distribution of Ae. albopictus populations in Connecticut aligns more closely to the 0°C isotherm; however, we anticipate future range expansion as founding populations continue to build, along with milder winters and hotter summers that are projected to increase in frequency under climate change [39].
Our current trapping methods for mosquito monitoring rely mainly on the deployment of CO2-baited CDC light and gravid traps. CDC light traps are effective for collecting a diversity of crepuscular and nocturnal feeding Anopheline and Culicine species, whereas gravid traps are more effective for trapping gravid Culex species. Neither trap type is particularly effective for collecting diurnal feeding and container-inhabiting Aedes species so we supplemented our collections by deploying BGS traps that have shown promise for the collection of Ae. albopictus. In this study, the BGS trap did not significantly improve capture of Ae. albopictus over the CDC light trap but both of these traps were significantly better than the gravid trap. These findings contrast with trap evaluations that were performed in locations with more established Ae. albopictus populations [40, 41]. In these studies, BGS traps outperformed CDC light traps that may be explained by their combined use of CO2 and BG-lures and/or the overall density of Ae. albopictus at the trapping site.
In this study, we isolated WNV and CVV from Ae. albopictus collected in Connecticut, raising concerns about its role as a potential arbovirus vector. Both of these arboviruses have been detected in field-collected Ae. albopictus from other states, including Pennsylvania and New Jersey [3, 42, 43]. Ae. albopictus has also been implicated as a potential bridge vector of WNV based on its vector competence [44] and feeding behavior that occasionally includes blood meals from virus-competent birds [45, 46]. However, in another study, WNV infection was not detected in more than 30,000 Ae. albopictus collected in New Jersey despite concurrent WNV amplification in the region, suggesting a limited role for this species as a vector [43]. It is striking that we isolated both CVV and WNV from a limited sample of Ae. albopictus, highlighting its potential threat, but further sampling and testing are required to assess its overall contribution. In addition, exotic arboviruses such as CHIKV, DENV, and ZIKV are frequently introduced into the U.S. by infected travelers returning from endemic countries necessitating research on the vector competence and capacity of local Ae. albopictus populations for these pathogens. This is an important priority given the central role that this species had in the autochthonous transmission of DENV and CHIKV in unlikely places such as Hawaii, France, Italy, and Japan [11, 12, 47, 48].
Acknowledgments
We thank our support staff, specifically Angela Bransfield and Michael Misencik who provided technical assistance in the processing and virus testing of mosquitoes.
Data Availability
All relevant data are within the paper and its supporting information files.
Funding Statement
This work was supported in part by grants from the Centers for Disease Control and Prevention (U50/CCU116806-01-1), the US Department of Agriculture Hatch Funds (CONH00773) and Multistate Research Project (NE1443). The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Reiter P, Sprenger D. The used tire trade: a mechanism for the worldwide dispersal of container breeding mosquitoes. J Am Mosq Control Assoc. 1987;3(3):494–501. [PubMed] [Google Scholar]
- 2.Kraemer MU, Sinka ME, Duda KA, Mylne A, Shearer FM, Brady OJ, et al. The global compendium of Aedes aegypti and Ae. albopictus occurrence. Sci Data. 2015;2:150035 10.1038/sdata.2015.35 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Farajollahi A, Nelder MP. Changes in Aedes albopictus (Diptera: Culicidae) populations in New Jersey and implications for arbovirus transmission. J Med Entomol. 2009;46(5):1220–4. [DOI] [PubMed] [Google Scholar]
- 4.Unlu I, Farajollahi A, Strickman D, Fonseca DM. Crouching tiger, hidden trouble: urban sources of Aedes albopictus (Diptera: Culicidae) refractory to source-reduction. PLoS One. 2013;8(10):e77999 10.1371/journal.pone.0077999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Valerio L, Marini F, Bongiorno G, Facchinelli L, Pombi M, Caputo B, et al. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) in urban and rural contexts within Rome province, Italy. Vector Borne Zoonotic Dis. 2010;10(3):291–4. 10.1089/vbz.2009.0007 [DOI] [PubMed] [Google Scholar]
- 6.Munnoz J, Eritja R, Alcaide M, Montalvo T, Soriguer RC, Figuerola J. Host-feeding patterns of native Culex pipiens and invasive Aedes albopictus mosquitoes (Diptera: Culicidae) in urban zones from Barcelona, Spain. J Med Entomol. 2011;48(4):956–60. [DOI] [PubMed] [Google Scholar]
- 7.Faraji A, Egizi A, Fonseca DM, Unlu I, Crepeau T, Healy SP, et al. Comparative host feeding patterns of the Asian tiger mosquito, Aedes albopictus, in urban and suburban Northeastern USA and implications for disease transmission. PLoS Negl Trop Dis. 2014;8(8):e3037 10.1371/journal.pntd.0003037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Luo L, Liang HY, Hu YS, Liu WJ, Wang YL, Jing QL, et al. Epidemiological, virological, and entomological characteristics of dengue from 1978 to 2009 in Guangzhou, China. J Vector Ecol. 2012;37(1):230–40. 10.1111/j.1948-7134.2012.00221.x [DOI] [PubMed] [Google Scholar]
- 9.Reiter P, Fontenille D, Paupy C. Aedes albopictus as an epidemic vector of chikungunya virus: another emerging problem? Lancet Infect Dis. 2006;6(8):463–4. 10.1016/S1473-3099(06)70531-X [DOI] [PubMed] [Google Scholar]
- 10.Grard G, Caron M, Mombo IM, Nkoghe D, Mboui Ondo S, Jiolle D, et al. Zika virus in Gabon (Central Africa)—2007: a new threat from Aedes albopictus? PLoS Negl Trop Dis. 2014;8(2):e2681 10.1371/journal.pntd.0002681 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Succo T, Leparc-Goffart I, Ferre JB, Roiz D, Broche B, Maquart M, et al. Autochthonous dengue outbreak in Nimes, South of France, July to September 2015. Euro Surveill. 2016;21(21). [DOI] [PubMed] [Google Scholar]
- 12.Rezza G, Nicoletti L, Angelini R, Romi R, Finarelli AC, Panning M, et al. Infection with chikungunya virus in Italy: an outbreak in a temperate region. Lancet. 2007;370(9602):1840–6. 10.1016/S0140-6736(07)61779-6 [DOI] [PubMed] [Google Scholar]
- 13.Delisle E, Rousseau C, Broche B, Leparc-Goffart I, L'Ambert G, Cochet A, et al. Chikungunya outbreak in Montpellier, France, September to October 2014. Euro Surveill. 2015;20(17). [DOI] [PubMed] [Google Scholar]
- 14.Paupy C, Delatte H, Bagny L, Corbel V, Fontenille D. Aedes albopictus, an arbovirus vector: from the darkness to the light. Microbes Infect. 2009;11(14–15):1177–85. 10.1016/j.micinf.2009.05.005 [DOI] [PubMed] [Google Scholar]
- 15.Gerhardt RR, Gottfried KL, Apperson CS, Davis BS, Erwin PC, Smith AB, et al. First isolation of La Crosse virus from naturally infected Aedes albopictus. Emerg Infect Dis. 2001;7(5):807–11. 10.3201/eid0705.017506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Mitchell CJ, Niebylski ML, Smith GC, Karabatsos N, Martin D, Mutebi JP, et al. Isolation of eastern equine encephalitis virus from Aedes albopictus in Florida. Science. 1992;257(5069):526–7. [DOI] [PubMed] [Google Scholar]
- 17.Sprenger D, Wuithiranyagool T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J Am Mosq Control Assoc. 1986;2(2):217–9. [PubMed] [Google Scholar]
- 18.Moore CG, Mitchell CJ. Aedes albopictus in the United States: ten-year presence and public health implications. Emerg Infect Dis. 1997;3(3):329–34. 10.3201/eid0303.970309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Giordano BV, Gasparotto A, Hunter FF. A checklist of the 67 mosquito species of Ontario, Canada. J Am Mosq Control Assoc. 2015;31(1):101–3. 10.2987/14-6456R.1 [DOI] [PubMed] [Google Scholar]
- 20.Hahn MB, Eisen RJ, Eisen L, Boegler KA, Moore CG, McAllister J, et al. Reported Distribution of Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus in the United States, 1995–2016 (Diptera: Culicidae). J Med Entomol. 2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Nawrocki SJ, Hawley WA. Estimation of the northern limits of distribution of Aedes albopictus in North America. J Am Mosq Control Assoc. 1987;3(2):314–7. [PubMed] [Google Scholar]
- 22.Andreadis TG, Thomas MC, Shepard JJ. Identification Guide to the Mosquitoes of Connecticut. New Haven, CT: The Connecticut Agricultural Experiment Station; 2005. [Google Scholar]
- 23.Andreadis TG. Failure of Aedes albopictus to overwinter following introduction and seasonal establishment at a tire recycling plant in the northeastern USA. J Am Mosq Control Assoc. 2009;25(1):25–31. 10.2987/08-5813.1 [DOI] [PubMed] [Google Scholar]
- 24.Andreadis TG, Anderson JF, Vossbrinck CR, Main AJ. Epidemiology of West Nile virus in Connecticut: a five-year analysis of mosquito data 1999–2003. Vector Borne Zoonotic Dis. 2004;4(4):360–78. 10.1089/vbz.2004.4.360 [DOI] [PubMed] [Google Scholar]
- 25.Jackson BT, Paulson SL, Youngman RR, Scheffel SL, Hawkins B. Oviposition preferences of Culex restuans and Culex pipiens (Diptera: Culicidae) for selected infusions in oviposition traps and gravid traps. J Am Mosq Control Assoc. 2005;21(4):360–5. 10.2987/8756-971X(2006)21[360:OPOCRA]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 26.Tang Y, Anne Hapip C, Liu B, Fang CT. Highly sensitive TaqMan RT-PCR assay for detection and quantification of both lineages of West Nile virus RNA. J Clin Virol. 2006;36(3):177–82. 10.1016/j.jcv.2006.02.008 [DOI] [PubMed] [Google Scholar]
- 27.Andreadis TG, Armstrong PM, Anderson JF, Main AJ. Spatial-temporal analysis of Cache Valley virus (Bunyaviridae: Orthobunyavirus) infection in Anopheline and Culicine mosquitoes (Diptera: Culicidae) in the northeastern United States, 1997–2012. Vector Borne Zoonotic Dis. 2014;14(10):763–73. 10.1089/vbz.2014.1669 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Rochlin I, Ninivaggi DV, Hutchinson ML, Farajollahi A. Climate Change and Range Expansion of the Asian Tiger Mosquito (Aedes albopictus) in Northeastern USA: Implications for Public Health Practitioners. PLoS One. 2013;8(4):e60874 10.1371/journal.pone.0060874 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Ogden NH, Milka R, Caminade C, Gachon P. Recent and projected future climatic suitability of North America for the Asian tiger mosquito Aedes albopictus. Parasit Vectors. 2014;7:532 10.1186/s13071-014-0532-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hanson SM, Craig GB Jr. Relationship between cold hardiness and supercooling point in Aedes albopictus eggs. J Am Mosq Control Assoc. 1995;11(1):35–8. [PubMed] [Google Scholar]
- 31.Thomas SM, Obermayr U, Fischer D, Kreyling J, Beierkuhnlein C. Low-temperature threshold for egg survival of a post-diapause and non-diapause European aedine strain, Aedes albopictus (Diptera: Culicidae). Parasit Vectors. 2012;5:100 10.1186/1756-3305-5-100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hanson SM, Craig GB Jr. Aedes albopictus (Diptera: Culicidae) eggs: field survivorship during northern Indiana winters. J Med Entomol. 1995;32(5):599–604. [DOI] [PubMed] [Google Scholar]
- 33.Hawley WA. The biology of Aedes albopictus. J Am Mosq Control Assoc Suppl. 1988;1:1–39. [PubMed] [Google Scholar]
- 34.Reiter P. Aedes albopictus and the world trade in used tires, 1988–1995: the shape of things to come? J Am Mosq Control Assoc. 1998;14(1):83–94. [PubMed] [Google Scholar]
- 35.Medeiros MC, Boothe EC, Roark EB, Hamer GL. Dispersal of male and female Culex quinquefasciatus and Aedes albopictus mosquitoes using stable isotope enrichment. PLoS Negl Trop Dis. 2017;11(1):e0005347 10.1371/journal.pntd.0005347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kobayashi M, Nihei N, Kurihara T. Analysis of northern distribution of Aedes albopictus (Diptera: Culicidae) in Japan by geographical information system. J Med Entomol. 2002;39(1):4–11. [DOI] [PubMed] [Google Scholar]
- 37.Benedict MQ, Levine RS, Hawley WA, Lounibos LP. Spread of the tiger: global risk of invasion by the mosquito Aedes albopictus. Vector Borne Zoonotic Dis. 2007;7(1):76–85. 10.1089/vbz.2006.0562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife. 2015;4:e08347 10.7554/eLife.08347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Horton H, Yohe G, Easterling W, Kates R, Ruth M, Sussman E, et al. Northeast. 2014. In: Climate Change Impacts in the United States: the Third National Climate Assessment [http://s3.amazonaws.com/nca2014/high/NCA3_Climate_Change_Impacts_in_the_United%20States_HighRes.pdf]. U. S. Global Change Research Program; [371–95].
- 40.Meeraus WH, Armistead JS, Arias JR. Field comparison of novel and gold standard traps for collecting Aedes albopictus in Northern Virginia. J Am Mosq Control Assoc. 2008;24(2):244–8. 10.2987/5676.1 [DOI] [PubMed] [Google Scholar]
- 41.Farajollahi A, Kesavaraju B, Price DC, Williams GM, Healy SP, Gaugler R, et al. Field efficacy of BG-Sentinel and industry-standard traps for Aedes albopictus (Diptera: Culicidae) and West Nile virus surveillance. J Med Entomol. 2009;46(4):919–25. [DOI] [PubMed] [Google Scholar]
- 42.Holick J, Kyle A, Ferraro W, Delaney RR, Iwaseczko M. Discovery of Aedes albopictus infected with west nile virus in southeastern Pennsylvania. J Am Mosq Control Assoc. 2002;18(2):131 [PubMed] [Google Scholar]
- 43.Armstrong PM, Anderson JF, Farajollahi A, Healy SP, Unlu I, Crepeau TN, et al. Isolations of Cache Valley virus from Aedes albopictus (Diptera: Culicidae) in New Jersey and evaluation of its role as a regional arbovirus vector. J Med Entomol. 2013;50(6):1310–4. [DOI] [PubMed] [Google Scholar]
- 44.Turell MJ, O'Guinn ML, Dohm DJ, Jones JW. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J Med Entomol. 2001;38(2):130–4. 10.1603/0022-2585-38.2.130 [DOI] [PubMed] [Google Scholar]
- 45.Savage HM, Niebylski ML, Smith GC, Mitchell CJ, Craig GB Jr. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) at a temperate North American site. J Med Entomol. 1993;30(1):27–34. [DOI] [PubMed] [Google Scholar]
- 46.Richards SL, Ponnusamy L, Unnasch TR, Hassan HK, Apperson CS. Host-feeding patterns of Aedes albopictus (Diptera: Culicidae) in relation to availability of human and domestic animals in suburban landscapes of central North Carolina. J Med Entomol. 2006;43(3):543–51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Effler PV, Pang L, Kitsutani P, Vorndam V, Nakata M, Ayers T, et al. Dengue fever, Hawaii, 2001–2002. Emerg Infect Dis. 2005;11(5):742–9. 10.3201/eid1105.041063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kutsuna S, Kato Y, Moi ML, Kotaki A, Ota M, Shinohara K, et al. Autochthonous dengue fever, Tokyo, Japan, 2014. Emerg Infect Dis. 2015;21(3):517–20. 10.3201/eid2103/141662 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper and its supporting information files.