

Summary
The recent discovery of human-induced pluripotent stem cells (iPSC) has revolutionized the field of stem cells. iPSC have demonstrated that biological development is not an irreversible process and that mature adult somatic cells can be induced to become pluripotent. This breakthrough is projected to advance our current understanding of many disease processes and revolutionize the approach to effective therapeutics. Despite the great promise of iPSC, many translational challenges still remain. The authors review the basic concept of induction of pluripotency as a novel approach to understand cardiac regeneration, cardiovascular disease modeling, and drug discovery. They critically reflect on the current results of pre-clinical and clinical studies using iPSC for these applications with appropriate emphasis on the challenges facing clinical translation.
Key Words: cardiac regeneration, disease modeling, drug discovery, induced pluripotent stem cells
Abbreviations and Acronyms: ESC, embryonic stem cell(s); iCM, induced pluripotent stem cell–derived cardiomyocyte(s); iEC, induced pluripotent stem cell–derived endothelial cell(s); iPSC, induced pluripotent stem cell(s); MI, myocardial infarction; SCID, severe combined immunodeficient
Graphical abstract
Origin of the Induced Pluripotent Stem Cells
The totipotency of a fertilized egg confers a unique ability to divide and differentiate into all tissue types to form an entire organism. To identify the exact role of a mammalian cell nucleus undergoing embryonic development, somatic cell nuclear transfer experiments were conducted in frogs in the 1950s (1). These experiments offered a proof of principle that pluripotency can be conferred to somatic cells by transferring their nuclear contents into oocytes (2). In fact, the nuclei of differentiated mammalian cells possess the ability to become pluripotent upon transfer of the nucleus into an oocyte or fusion with embryonic stem cells (ESC) 3, 4. For many years, the major challenge was to reprogram the somatic cell nucleus without transferring its contents or using an oocyte (5). In 2006, Takahashi and Yamanaka (6) studied the effects of 24 transcription factors, which were known to confer pluripotency to early embryos and ESC. They succeeded in transforming the adult mouse fibroblasts into induced pluripotent stem cells (iPSC) using 4 select transcription factors: Oct4, Sox2, c-Myc, and Klf4 6, 7. These iPSC exhibited characteristics very similar to ESC. One year later, the same group generated human iPSC from human dermal fibroblasts using retroviral transduction with the same 4 transcription factors (8). These iPSC have morphology, cell surface markers, and genes characteristics similar to human ESC, exhibiting unlimited replication potential without telomere shortening or karyotype changes. The multilineage differentiation potential of iPSC has been confirmed both in vitro in embryoid bodies and in vivo based upon teratoma formation upon injection into severe combined immunodeficient (SCID) mice. Other groups reproduced the “stemness” or induced pluripotency of somatic cells using the same or slightly different transcription factors, demonstrating the robustness of this technique and revolutionizing the field of stem cell biology 9, 10, 11.
Since the ability to generate iPSC in culture from adult human skin fibroblasts has been established 8, 9, this pluripotent state has been induced in a variety of human cells including keratinocytes (12), T-lymphocytes (13), peripheral mononuclear blood cells (14), cord blood (15), placenta (16), neural stem cells (17), adipose tissue (18), and renal epithelial cells present in urine samples (19). Ectopic expression of different combinations of reprogramming factors has been used, with Oct4 being the most consistent across protocols. Furthermore, several protocols have been developed for directed in vitro differentiation of iPSC into spontaneously contractile cardiomyocytes, smooth muscle cells, and vascular endothelial cells 20, 21. The differentiation of iPSC into cardiomyocytes was confirmed by microscopic examination of beating colonies, immunostaining for cardiac proteins, electrophysiological testing, and real-time polymerase chain reaction analysis of cardiac markers 20, 21 (Figures 1A to 1C). The iPSC overcame the ethical concerns and immunogenicity of human ESC, thus becoming an attractive alternative for autologous tissue repair and regeneration as well as a source for allogeneic transplantation. The therapeutic potential of iPSC-derived cells has been tested in many pre-clinical studies with some encouraging results in murine and porcine models of myocardial infarction (MI) 16, 22, 23, 24, 25. Currently, the first human trials of iPSC-derived cells, aimed at establishing the safety of these cells, are enrolling patients with age-related macular degeneration in Japan (26).
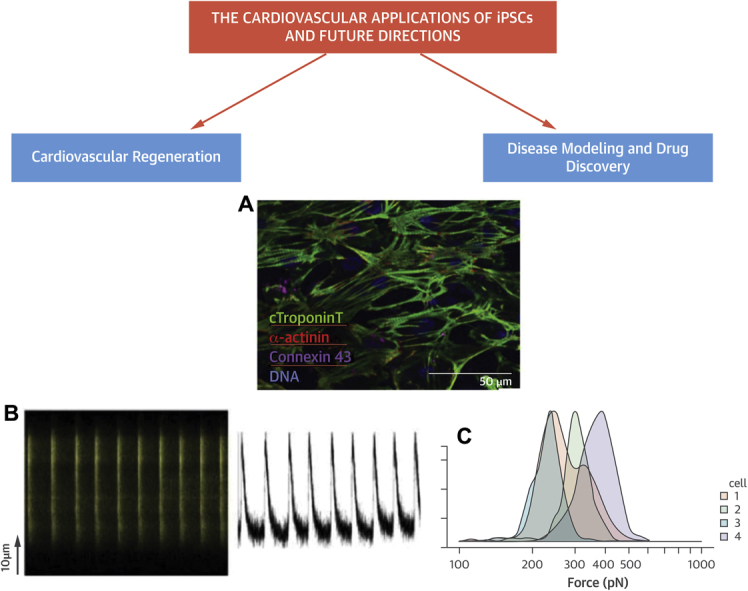
Figure 1.

The Cardiovascular Applications and Characteristics of iPSC-Derived Cardiomyocytes
(A) Immunostain of induced pluripotent stem cell (iPSC)-derived cardiomyocytes in cell culture demonstrate striated pattern of cardiac troponin T, α-actinin, connexin 43, and deoxyribonucleic acid (DNA). (B) Calcium metabolism and atomic force microscopy demonstrate live-cell calcium imaging of the contractile iPSC–derived cardiomyocytes, exhibiting regular and synchronous Ca2+ transients (left) with each contractile activity (right). (C) Histogram of atomic force microscopy evaluates the force exerted by each contraction of a representative iPSC-derived cardiomyocytes. The maximal contractile force measured for 4 representative beating cells was comparable to that of native cardiomyocytes (150 to ∼550 pN).
Beyond the potential for regenerative or transplantation therapies, iPSC offer an unprecedented opportunity to recapitulate both normal and pathologic human tissue formation in vitro, thereby providing novel cell-based biological models that enable better understanding of disease pathogenesis and drug discovery (27). From 2008 to 2015, more than 70 human iPSC-based disease models have been published in an exponential fashion (28). The ability to examine the direct effects and toxicity of new drugs on the patients’ own cells could also represent an invaluable tool for drug development and discovery. For these reasons, both Sir John Gurdon, who performed nuclear transfer experiments, and Shinya Yamanaka were awarded the Nobel Prize in Physiology or Medicine in 2012. This review discusses the current status of the cardiovascular applications of iPSC for cardiac and vascular regeneration, disease modeling, and drug discoveries with a special emphasis on the current challenges for clinical translation (Figure 1).
iPSC for Cardiovascular Regeneration
With the growing epidemic of heart failure, cardiac regeneration represents a major priority of regenerative medicine. The ability of iPSC to differentiate into autologous tissue-specific cells, similar to ESC but without the need to destroy a human embryo, is an important breakthrough in human stem cell biology. A number of pre-clinical studies have explored the effects of intramyocardial injection of induced pluripotent stem cell–derived cardiomyocytes (iCM) into murine and porcine models of MI (a complete recent list of pre-clinical studies is provided in Lalit et al. [29]). Nelson et al. (30) were the first to perform intramyocardial injection of iPSC-derived cells into a murine model of acute MI. They found improved left ventricular ejection fraction, fractional shortening, and regional wall motion on echocardiography 4 weeks after permanent coronary artery ligation when compared with the fibroblast-injected animals. Interestingly, they reported teratoma formation upon subcutaneous and intramyocardial injections of iPSC in immunodeficient animals but none in immunocompetent mice (30).
Other studies have reported formation of intramural teratomas in both immunocompetent and SCID mice after intramyocardial injection of undifferentiated iPSC 31, 32. Kim et al. (32) tracked the engraftment of iPSC at 4 weeks in a SCID murine model of MI. They demonstrated teratoma formation with no myocardial or endothelial cell differentiation despite the reported improvement in myocardial function and myocardial viability (32). This demonstrates a potential mechanistic role of cytokines released from the iPSC, which may restore cardiac function despite teratoma formation. Similar results were obtained with undifferentiated iPSC in a rat model of MI (33), which is consistent with the initial studies of undifferentiated murine ESC that resulted in teratoma formation 34, 35.
Repeat experiments using differentiated skeletal myoblast derived-iCM obtained from day 10 embryoid body–beating aggregates reported improved cardiac function with decreased infarct size and no evidence of tumorigenesis (36). Similar improvement in left ventricular function without teratoma formation was reported by the same group using cardiac progenitors harvested from embryoid body–beating aggregates of mesenchymal stem cells' derived-iPSC (37). Currently, it is widely thought that the pluripotent stem cell derivatives from either ESC or iPSC are still premature for clinical therapy because of the tumorigenic potential. Active research is ongoing using highly purified monolayer populations of differentiated iCM and methods to remove the undifferentiated iPSC (38). Through these efforts, the generation of >90% pure populations of iCM is increasingly routine in laboratories worldwide (39).
Using a single transcription factor (Oct3/4), Pasha et al were able to generate iPSC from mouse skeletal myoblasts that endogenously express 3 of the Yamanaka factors (40). They were also able to isolate cardiac progenitors from 5-day beating embryoid bodies. In this experiment, iPSC-derived cardiac progenitor cells showed sustained engraftment and gap junction formation upon transplantation into the infarcted murine heart (40). Another group reported significantly restored cardiac function in post-MI ventricular remodeling after intramyocardial injection of fetal-liver kinase 1–positive iPSC-derived cardiac progenitor cells into SCID mice (41). Based on the advantage conferred by epigenetic memory, cardiac progenitor cells expressing the surface marker Sca1 were used for induction of pluripotency to leverage their preferential cardiac differentiation potential. Compared with skin fibroblast-derived iPSC, cardiac progenitor cell–derived iPSC were more efficient in producing cardiomyocytes, smooth muscles cells, and endothelial cells (25). However, in vivo echocardiography and post-mortem infarct size analysis showed no significant difference between the 2 types of cells that were only marginally better than placebo at 8 weeks (25).
To improve the engraftment of iCM into the infarcted myocardium, Dai et al. (42) developed a homologous peritoneal patch seeded with iCM, CD31-selected iPSC-derived endothelial cells, and mouse embryonic fibroblasts (tri-cell patch). The tri-cell patch was used to cover the infarcted area instead of an intramyocardial injection into an infarcted myocardium 42, 43. Kawamura et al. (22) used a cell-sheet technique to ensure delivery of a large number of cells. Immunosuppressed minipigs (n = 6) were transplanted with human iCM in cell sheets, containing approximately 25 million cells, 4 weeks after MI. Significantly improved left ventricular ejection fraction and reduced end-systolic volume were noted in the iCM group. Despite these positive results over sham pigs, poor engraftment was noted by 8 weeks. The investigators proposed that the poor engraftment was due to insufficient immune suppression with a single agent (tacrolimus) in a xenogeneic transplantation model (22).
Beyond myocardial regeneration, many investigators have employed iPSC technology for inducing vascular regeneration in patients with coronary and peripheral artery disease where new vascular structures might overcome impaired perfusion caused by occlusive atherosclerotic disease. Most work in vascular regeneration has been done with adult stem cells (44). For peripheral artery disease, smaller clinical trials of adult stem cell therapy have shown promise in populations with different levels of disease severity 45, 46. Currently, a large National Institutes of Health/National Heart, Lung, and Blood Institute–sponsored Cardiovascular Cell Therapy Research Network trial using aldehyde dehydrogenase bright cells for intermittent claudication is ongoing (47). Stem cells obtained from adult tissues can be problematic, however. In elderly patients and those with comorbidities such as diabetes—the patients most likely to benefit from stem cell therapy—the quantity and function of stem cells can be impaired 48, 49. In contrast, iPSC can theoretically be derived from many tissue sources, thereby obviating the issue of cell quantity and quality. Furthermore, due to their pluripotency, iPSC can differentiate into the multiple cell types needed in ischemic tissue such as smooth muscle, pericyte, and cardiac cells in addition to the endothelial cells for regenerative purpose (30).
Although there are currently no clinical trials using iPSC for neovascularization, pre-clinical studies show great promise (50). Choi et al. (51) and Taura et al. (52) were among the first researchers to describe differentiation of iPSC into endothelial cells. Both groups found that the induced pluripotent stem cell–derived endothelial cells (iEC) were very similar to endothelial cells derived from human ESC. Since then, the iEC have been used in murine hindlimb ischemia models and have been found to improve limb perfusion at least in part through neovascularization. Compared with the human ESC, iEC appear to have similar therapeutic efficacy (53). Another potential use of the iEC in the setting of vascular insufficiency is for engineering better vascular conduits. Wang et al. (54) have demonstrated that functional smooth muscle cells can be derived from human iPSC and used on a scaffold to regenerate vascular tissue in vivo. Additional pre-clinical studies will demonstrate the potential efficacy of these novel therapeutic approaches.
Current Challenges for the iPSC in Cardiovascular Regeneration
Despite the promise of iPSC for regenerative medicine, there are still many roadblocks remaining that may hinder their clinical use.
Cell engraftment
Poor cell engraftment remains a major challenge for all stem cell therapies for cardiovascular applications. The inability of the transplanted cells to exhibit long-term survival and engraftment into the host myocardium with successful electromechanical integration into the complex cardiac syncytium has been a consistent finding throughout many studies 55, 56, 57, 58. Despite the fast pace of progress in the field of stem cell biology, there has been no improvement in engraftment over the past decade regardless of the types of stem cells used 25, 57, 59, 60. In many pre-clinical studies with positive functional results, the assessment of long-term, stable engraftment and electromechanical integration of the stem cells by the use of molecular reporter genes has yielded variable results 32, 59. In most of these studies, the mechanism underlying the positive results was postulated to be a “paracrine effect” 22, 32, 58. The shortcoming of the paracrine effect hypothesis is that it is difficult to measure this process in isolation. It is also challenging to pinpoint the specific biochemical factor(s) responsible for the salubrious actions of stem cells. Nevertheless, investigations are in order because if such a paracrine factor(s) is identified, this finding may obviate the need for cell therapy.
To address questions regarding the magnitude and mechanism of cardiac repair after injury in a practical way, multiple animal and clinical studies have tested the efficacy of different stem cell types for cardiac regeneration (58). Nevertheless, the limited engraftment of the stem cells and the inability to track the cells delivered in vivo have left many unanswered questions regarding the mechanism and description of the postulated benefit.
Although no direct correlative data exist, the poor cell engraftment may be associated with the low regenerative capacity of the heart. Earlier studies in late 1990s and early 2000s using evidence from small-animal experiments (61), clinical sex-mismatched heart and bone marrow transplantation studies (62), and the observation of dividing myocytes within the infarcted myocardium (63) and failing hearts (64) showed that the heart may possess significant regenerative capacity, contradicting the old dogma of the heart as a terminally differentiated organ. These studies, together with the concomitant isolation of different types of adult human stem cells 65, 66 and human ESC (67), established the foundation of the emerging field of regenerative cardiology. Despite the optimism that accompanied these studies, the rate and magnitude of ongoing turnover and cardiac regeneration have been controversial, with more recent studies showing that adult cardiomyocytes may possess very limited mitotic capability 68, 69. Bergmann et al. (68) calculated the post-natal human cardiomyocyte renewal at ∼1% annually at age 20, declining to 0.4% annually by age 70. More recently, van Berlo et al. (69) found that endogenous c-kit+ cells mainly regenerate endothelial cells with only a minor contribution toward cardiomyocyte renewal, estimated at a rate of 0.03% to 0.008%.
Another reason for the low engraftment could be the study design and route of administration of stem cells. Most stem cell studies attempted to deliver stem cells into an area of infarction, either directly through intramyocardial injection or indirectly through intracoronary infusion, to regenerate the myocardium. Such a design worked well with bone marrow transplantation but may not be suitable for a complex organ such as the heart, particularly in the setting of ischemic cardiomyopathy with impaired perfusion, increased oxidative stress, and inflammation. Even in the skin, which is a regenerative organ, skin burns are not cured by injecting stem cells into the skin but rather by using skin grafts. It may be necessary to modify current delivery techniques to address the complexity of the heart. Efforts are underway by several groups using different tissue-engineering techniques and cell derivatives 22, 42, 43.
Cell engraftment in studies of vascular regeneration poses a different challenge. In contrast to the low regenerative capacity of cardiomyocytes, endothelial cells show greater capacity for regeneration and repair in response to vascular injury via endothelial progenitor cells mostly derived from the bone marrow 70, 71. However, age and cardiovascular diseases such as coronary artery disease, cerebral vascular disease, congestive heart failure, and diabetes can significantly reduce both the quality and quantity of circulating endothelial progenitor cells. Though studies evaluating the nature of iPSC incorporation into native vascular endothelium are lacking, using embryonically derived endothelial cells (ESC-EC), Huang et al. (72) provided evidence of stem cell engraftment and neovascularization in a mouse ischemic limb model.
Technical issues with the induction of pluripotency, cardiac differentiation, and maturation
Reprograming somatic cells to a pluripotent state involves introducing transgenes, their derivative ribonucleic acid, or protein products with multiple passages in tissue culture. Several studies suggest that the reprogramming process induces genomic instability and down-regulates tumor suppressor genes, which can affect cellular integrity 73, 74, 75, 76. Comparing 12 human iPSC lines generated from the same primary fibroblast cell, Laurent et al. (76) found genetic aberrations in iPSC lines that were not present in the original fibroblast line. Also, Li et al. (75) calculated the degree of genetic heterogeneity among 24 iPSC clones obtained from skin fibroblasts of the same 8-week-old mouse and found 383 iPSC variants from the original 24 clones with a mean of 16.0 variants/clone and a range of 0 to 45, indicating a wide range of heterogeneity. For these reasons, iPSC are not approved for clinical use in the United States, and many have suggested that the study in the Japanese patients with age-related macular degeneration may be premature (77).
As the field of iPSC is only few years old, protocols for induction of pluripotency and cardiac differentiation are still evolving and there is a lack of consensus. Current iPSC lines have been generated using different cells of origin, different transcription factor combinations, different vectors (viral and nonviral), different time in culture, different number of passages, and different selection methods. Such inconsistency adds further layers of complexity to the endogenous heterogeneity created by the reprogramming process and expansion in culture, resulting in the absence of the uniformity needed for clinical therapeutic applications.
Pluripotency can be induced from a variety of human cells 12, 13, 14, 15, 16, 17, 18, 19, and the best cell of origin for induction of pluripotency for cardiac application is not known yet. Several studies have suggested that after induction of pluripotency, somatic cells retain some characteristics of their past identity, the so-called epigenetic memory (78). Although this epigenetic memory can be seen as a sign of incomplete programming, it can be used to facilitate differentiation toward the desired cell type more efficiently (27).
Another technical issue is the low efficiency of the reprogramming process, which ranges from as much as 4.4% with modified messenger ribonucleic acid and 1% with integrating viruses to as little as 0.001% with the nonintegrating Sendai virus, adenovirus, plasmids, and direct protein delivery (79). Efficiency decreases further if one does not use the oncogene c-myc (80). Currently, integrating viruses are falling out of favor because of the oncogenic potential resulting from incomplete proviral silencing. A recent comprehensive analysis of nonintegrating reprogramming methods for human iPSC was published to compare the quality of cells, reliability, efficiency, speed of colony emergence, aneuploidy rate, and hands-on time for each method (81).
Optimized protocols for cardiac differentiation have been developed that replicate the key commitment stages found during embryonic development (82). Whereas removal of the undifferentiated cells and purification of the iCM are becoming more standardized (83) (Figure 2), it is becoming evident that the iCM may represent immature and heterogeneous groups of cardiomyocytes 38, 84. A recent study calculated ventricular cardiomyocytes to be 50% of differentiated cardiomyoctes, with atrial cardiomyocytes being 37.5% and nodal cells 12.5% (85). The clinical use of this mixture of different iCM again may generate inconsistent results. A similar phenomenon is seen in the production of human iPSC-EC (iEC), which have been found to express multiple cell markers associated with lymphatic, venous, and arterial EC (86). In this study, higher levels of VEGF were associated with more iEC with an arterial phenotype and it was this arterial phenotype of cells that demonstrated the best in vivo performance in the development of mature and stable capillaries.
Figure 2.

Flow Cytometry Analysis of iCM Purity
Cardiac troponin T (cTnT), vascular adhesion molecule-1 (VCAM1), and cTnT/VCAM1 double-positive cells demonstrate high purity of induced pluripotent stem cell–derived cardiomyocytes (iCM). VCAM1 was shown to be a potent cell surface marker for robust, efficient, and scalable purification of cardiomyocytes from human embryonic stem cells/human induced pluripotent stem cells (83). FSC = forward-scattered light; LL = lower left; Q5 = human endothelial cells; UL = upper left; UR = upper right; SSC = side-scattered light.
Cardiomyocyte maturation is another important step for cell therapy. In vitro tissue culture conditions are obviously different from the environment of developing embryonic hearts. The tensile stretch exerted on the developing cardiomyocytes as well as the micro-environmental milieu of molecules and growth factors play a key role in cardiomyocytes’ growth and maturation. The current cell culture techniques on the flat surface of a petri dish or in suspension deprive the developing cardiomyocytes of important maturation signals. Newer culture techniques that offer optimal tensile structure for the developing cardiomyocytes, pulsed treatment with novel molecules and growth factors, and electrophysiological cues may enable selection of the iCM at a developmentally appropriate stage for therapeutic purposes (87).
Immunogenicity
One advantage of iPSC over other types of stem cells is their autologous source, with no expected immune rejection. This assumption is also supported by the lack of immune rejection upon administration of iPSC in many pre-clinical studies. However, a few studies have reported T-cell immune rejection of undifferentiated iPSC upon transplantation into syngeneic mice 88, 89. These studies raise the possibility that cell reprogramming, genetic instability, culture conditions, and frequent passages could alter iPSC in a way that renders them immunogenic. However, further studies using differentiated iPSC have shown negligible immunogenicity 90, 91. Taken together, the available evidence indicates that although immune rejection may be possible, it does not seem to be a consistent finding. The ongoing clinical trial in age-related macular degeneration in Japan will also address this question. However, we believe that further pre-clinical studies using differentiated cells are required to resolve the issue of immunogenicity.
Autologous versus allogeneic source
One great advantage of iPSC is the autologous source, which makes iPSC an inexhaustible source for cell therapies without immune rejection or ethical issues. However, there are several logistic issues with an autologous source. Induction of pluripotency followed by lineage-specific differentiation is a time-consuming process that requires multiple passages of iPSC followed by differentiation and purification, which may make autologous iPSC suitable only for chronic illnesses but not for acute conditions such as acute MI. Also, custom-made autologous cells would make clinical good manufacturing practice, quality assurance, regulatory compliance, and elimination of technical errors very expensive, limiting the reproducibility of the technique and its potential wide-scale clinical application (92). This paradigm, however, applies to almost all autologous cell therapies requiring in vitro expansion.
Whereas iPSC are traditionally considered a potential autologous therapy, the ability to identify iPSC genotypes and preselect matching donors provides a potential opportunity for allogeneic therapy. Several investigators have proposed the creation of large allogeneic libraries of clinical-grade quality iPSC and iPSC-derivatives 92, 93, 94. However, such cell/tissue libraries would require several thousand random donors to cover the human leukocyte antigen genotype variability within a specific population 94, 95. Despite logistic and financial difficulties, establishment of a blood-derived iPSC library has been initiated in Japan (92). This approach would provide safe off-the-shelf allografts of human leukocyte antigen–homozygous iPSC-derivatives ready for clinical use after excluding tumorigenic clones (96). Another advantage of the iPSC library is that the donors will be subjected to only minor procedures (e.g., blood sampling, skin biopsy) whereas the iPSC can be used in an unlimited number of matched recipients (93), addressing the tissue availability issue and potentially improving graft survival (97), but not completely eliminating the need for immunosuppressive medications.
Tumorigenicity
Tumorigenicity represents another potential obstacle to the clinical application of iPSC. In fact, the current functional gold standard test for pluripotency of human iPSC is teratoma formation upon injection into immunodeficient mice (7). Pluripotent origin, incomplete differentiation, and difficulty in eliminating all undifferentiated cells may lead to potential teratoma formation (7). However, advances in differentiation techniques and tissue culture conditions have mitigated this challenge, resulting in highly pure (>90%) iCM populations 38, 39.
In addition to the potential for teratoma formation by the undifferentiated cells, differentiated iPSC still carry an intrinsic risk of malignant transformation (98). The original use of retroviruses for induction of pluripotency with integration into the host deoxyribonucleic acid is a contributing factor. The overexpression of oncogenes, such as c-Myc, used in the original iPSC reprogramming protocol, carries another uncertainty. This has led to modifications of the original technique to induce pluripotency without using retroviral vectors (81). The current use of the ribonucleic acid nonintegrating modified Sendai virus eliminates the potential risk of genomic integration and maintains good transduction efficacy (99).
Future Directions for Cardiovascular Regeneration
Despite the limited innate ability of the injured myocardium and vasculature to regenerate, the implementation of iCM and iEC holds promise for regenerative medicine. Simpson et al. (100) succeeded in constructing a biologically active human pulmonary valve from a decellularized valve that was seeded with iPSC-derived mesenchymal stem cells. Also, combining bioengineering technology with iPSC-derivatives to construct engineered tissue may be more effective than direct intramyocardial injection to limit infarct expansion and adverse remodeling, leading to heart failure 22, 42.
Direct reprogramming or transdifferentiation, whereby terminally differentiated cells such as cardiac fibroblasts are changed into cardiomyocytes without first producing a pluripotent intermediate, may be an alternative strategy. Transdifferentiation has been known for decades. It was defined by Pritchard in 1978 as “the ability of a cell to lose a definitive characteristic and to acquire another feature characteristic of an alternate state” (quoted in [101]). The advent of iPSC technology has revolutionized the field of transdifferentiation by demonstrating that biological development is not an irreversible process. The discovery demonstrated that the individual cell function is largely dictated by changes in gene expression within each individual tissue or cell. Inspired by the experimental findings of Takahashi and Yamanaka (6), Ieda et al. (102) tested 14 key transcription factors for early heart development to reprogram fibroblasts into cardiomyocyte-like cells. They were able to convert mouse post-natal cardiac or dermal fibroblasts into functional beating cardiomyocytes in vitro using a combination of 3 transcription factors—Gata4, Mef2c, and Tbx5 (102). In 2012, in vivo delivery of Gata4, Mef2c, and Tbx5 transcription factors using a gene therapy approach converted mouse nonmyocytes into induced cardiomyocytes with improved cardiac function and reduced scar size after MI 103, 104. Compared with the in vitro induced cardiomyocytes, in vivo induced cells appeared more similar to endogenous cardiomyocytes, suggesting the importance of in vivo environmental cues such as electromechanical stimulation, signaling pathways, and extracellular matrix (105). This approach appears promising but carries significant limitations, including low efficiency, incomplete reprogramming, lack of robust experimental reproducibility, and the use of retroviral vectors in vivo (106). Greater understanding of the epigenetic changes associated with direct reprogramming is needed to enhance its therapeutic potential.
Similarly, Margariti et al. (107) have described partial reprogramming of fibroblasts directly into iEC using Oct4, Sox2, Klf4, c-Myc, and endothelial cell-specific media and culture conditions. These cells were able to repopulate decellularized vessel scaffolds with appropriate vascular structure and good attachment. However, this model still requires viral transduction. Moving away from this approach entirely, Lee et al. (108) have described the role of innate immunity in nuclear reprogramming and have gone on to demonstrate that using a toll-like receptor 3 agonist and exogenous endothelial cell growth factors, fibroblasts can be induced into EC (109). When transplanted into a murine ischemic hind limb model, these EC led to neovascularization and improved limb perfusion. Ultimately, safe, effective, and reproducible large-animal models will be necessary for eventual clinical translation.
iPSC for Disease Modeling and Drug Discoveries
iPSC have an emerging role in modeling human diseases at the cellular level and in facilitating new drug discoveries. Modeling of human diseases contributes to our understanding of the underlying pathophysiology, development of novel therapeutics, and evaluation of therapeutic efficacy and toxicity. Currently available disease models rely mainly on in vitro human cell lines and animal models for human diseases. The inherent limitations of this paradigm have hindered rapid advances in the understanding of many disease processes and their subsequent treatment. Similarly, multicenter clinical trials do not address the interindividual genetic variations that will necessitate larger numbers of participants with resultant increased time and cost.
The introduction of human iPSC as a source of in vitro disease models has added a new dimension to the current biomedical research tools. Shortly after the discovery of human iPSC, Park et al. (110) were able to prepare disease-specific iPSC lines in culture from patients with genetic disease, both with Mendelian and complex inheritance. Modeling rare genetic diseases such as long QT syndrome (111), LEOPARD (Lentigines, Electrocardiographic conduction abnormalities, Ocular hypertelorism, Pulmonary stenosis, Abnormalities of genitalia, Retardation of growth, Deafness) syndrome (112), and familial dilated cardiomyopathy (113) using patient-derived iPSC provided a deeper insight into the molecular mechanisms and signaling pathways responsible for disease phenotype. Obtaining peripheral blood cells or skin biopsy from patients with rare genetic heart diseases enables researchers to model the disease in vitro. The molecular mechanism of different diseases can be thoroughly studied at an individual cellular and genetic level in an unprecedented manner. Integration of patient-derived iPSC with the advances in bioengineering such as the disease-on-a-chip technology will take these models from an individual cell to the tissue level (114). With such advances, modeling genetic mitochondrial cardiomyopathy on a chip has become a reality (115).
With the development of successful cardiomyocyte differentiation protocols, Moretti et al. (111) modeled type 1 long QTc syndrome in vitro. They were able to describe a dominant negative trafficking defect that causes prolonged action potential in ventricular and atrial cardiomyocytes, which are exacerbated upon exposure to catecholamines and attenuated with beta-adrenergic blockers (111). The response to isoprenaline and metoprolol further validates this in vitro model as it correlates with the known effect of these medications in long QTc syndrome patients. iPSC-derived disease models not only include genetic abnormalities but also structural heart diseases such as the hypoplastic left heart syndrome (116). A full list of iPSC-derived cardiovascular disease models is shown in Table 1.
Table 1.
Disease Models and Drug Testing Studies of Cardiovascular Disease
| Mutation | Aim | Drug Tested | Number | First Author (Ref. #) | |
|---|---|---|---|---|---|
| Supravalvular aortic stenosis | ELN | Modeling + therapy | Elastin recombinant protein | 2 patients 2 control subjects | Ge et al. (128) |
| Hypoplastic left heart syndrome | N/A | Modeling | Isoproterenol | 1 patient 1 control subject | Jiang et al. (116) |
| ARVD | PKP2-2057del2 | Therapy | SB216763 | 2 patient 2 control subjects | Asimaki et al. (129) |
| ARVD | PKP2 L614P | Modeling | N/A | 1 patient 1 control subject | Ma et al. (130) |
| ARVD | PKP2 c.2484C>T PKP2 c.2013delC | Modeling | N/A | 2 patients 2 control subjects | Kim et al. (131) |
| Familial dilated cardiomyopathy | TNNT2-R173W point mutation | Therapy | Metoprolol | 4 patients 3 control subjects∗ | Sun et al. (113) |
| Hypertrophic cardiomyopathy | MYH7 Arg663His | Modeling | Verapamil, Diltiazem | 5 patients 5 control subjects∗ | Lan et al. (132) |
| LEOPARD syndrome | PTPN11 T468M | Modeling | N/A | 2 patients 2 control subjects | Carvajal-Vergara et al. (112) |
| Fredrick ataxia | FXN GAA triplet repeat expansion | Modeling | N/A | 2 patients 2 control subjects | Hick et al. (133) |
| Pompe syndrome | GAA | Therapy | rhGAA enzyme, 2-3-methyladenine, 3-L-carnitine | 2 patients 2 control subjects | Huang et al. (134) |
| Barth syndrome | TAZ c.517delG TAZ c.328T>C | Modeling + therapy | TAZ modRNA | 2 patients 3 control subjects∗ | Wang et al. (115) |
| CPVT | RyR2 P2328S | Modeling | Adrenaline | 1 patient 2 control subjects | Kujala et al. (135) |
| CPVT | RyR2 S406L | Therapy | Dantrolene | 1 patient 1 control subject | Jung et al. (136) |
| CPVT | CASQ2 D307H | Modeling | Isoproterenol | 2 patients∗ 3 control subjects | Novak et al. (137) |
| CPVT | RyR2 M4109R | Therapy | Flecainide, Thapsigargin | 1 patient 1 control subject | Itzhaki et al. (138) |
| LQT1 | KCNQ1 R190Q missense mutation | Modeling | Propranolol, Isoproterenol | 2 patients 2 control subjects | Moretti et al. (111) |
| LQT2 | KCNH2 A614V | Therapy | Nifedipine, Pinacidil, Ranolazine, Cisapride, IKr blocker E-4031 | 1 patient 1 control subject | Itzhaki et al. (139) |
| LQT2 | KCNH2 R176W | Modeling | Sotalol | 1 patients 3 control subjects | Lahti et al. (140) |
| LQT2 | KCNH2 G1681A | Therapy | PD118057, Nicorandil, ß-blocker | 2 patients∗ 2 control subjects | Matsa et al. (141) |
| Timothy syndrome (LQT8) | Calcium-channel gene G1216A | Therapy | Roscovitine | 2 patients 2 control subjects | Yazawa et al. (142) |
ARVD = arrhythmogenic right ventricular dysplasia; CASQ2 = calsequestrin 2; CPTV = catecholaminergic polymorphic ventricular tachycardia; ELN = elastin; FXN = frataxin; LEOPARD = Lentigines, Electrocardiographic conduction abnormalities, Ocular hypertelorism, Pulmonary stenosis, Abnormalities of genitalia, Retardation of growth, Deafness; LQT = long QTc syndrome; modRNA = modified RNA; N/A = not applicable; PKP2 = plakophilin-2; PTPN = protein-typrosine phosphatase nonreceptor; rhGAA = recombinant acid alpha-glucosidase; RYR2 = cardiac ryanodine receptor; TAZ = tafazzin.
Patients from the same family.
For drug testing, iPSC-derived disease models can serve as an additional in vitro arm to augment phase II and phase III clinical trials for dose optimization, efficacy, safety, side effects, and drug-drug interaction (28). The signal is measured in terms of physiologic effects such as shortening of the action potential duration, reduction in protein or gene expression, enzymatic activity, cell proliferation, apoptosis, or other measurable signal related to the disease process (117). Testing a small number of candidate drugs is an efficient way to monitor their therapeutic effects. Also, evaluating the effects of hundreds of medical compounds on patient-specific iPSC-derived cells in a systematic, less biased manner is possible using high-throughput drug screening systems (118). Lee et al. (118) tested the effect of 6,912 compounds on human iPSC-derived neural crest precursors. They found 8 compounds capable of rescuing the gene responsible for familial dysautonomia (118). Another big advantage of using iPSC for disease modeling and drug discovery is the very small number of patients needed to achieve valid conclusions about disease mechanisms and drug toxicity (Table 1). This will not preclude the need for eventual clinical trials as the final confirmatory step in drug discovery. However, the insights gained from these models are expected to increase the efficiency of the process by decreasing the need for some of the lengthy animal research. Furthermore, the ability to predict a patient’s response may enhance the trial design by shortening the duration and reducing the cost of randomized clinical trials, which may lead the transition to personalized medicine.
Similarly, high-throughput screening systems for drug discovery may be employed to investigate iPSC-derivatives from different tissue types for toxicity and side effects. Using iCM, Guo et al. (119) tested 88 drugs for in vitro arrythmogenicity potential. Navarette et al. (120) developed an in vitro system to screen for drug-induced arrhythmia using iCM from a skin biopsy of a 14-year-old volunteer. iCM are currently commercially available for such use. Pharmacokinetic data are also possible by evaluating hepatic metabolism in iPSC-derived hepatocytes. To validate such a notion, Takayma et al. (121) were able to assess interindividual differences in the hepatic metabolism and drug response using iPSC-derived hepatocyte-like cells derived from primary human hepatocytes of 12 donors. The cytochrome P450 metabolism of iPSC-derived hepatocyte-like cells was found to correlate highly with the primary human hepatocytes of origin, with preservation of interindividual differences (121).
Though it remains difficult to model adult-onset diseases such as atherosclerotic vascular disease using iPSC, Adams et al. (122) have found that iEC demonstrate functional features that may make them good candidates for understanding the role of EC in cardiovascular disease and for testing EC-based therapies. Specifically, the investigators demonstrated that in response to proinflammatory factors, iEC become activated and promote leukocyte transmigration in vitro. When subjected to flow dynamics in vitro that are known to promote or protect against atherosclerosis formation, iEC exhibit behavior similar to what is seen in human models of disease with different expression of mechanoregulated genes.
Limitations to iPSC-derived disease models
Many of the above-mentioned challenges for cardiovascular regeneration are also applicable to iPSC-derived disease models. Genetic alterations during the reprogramming process, incomplete reprogramming, changes induced during passages in culture, and differentiation-induced heterogeneous states of iCM development, including immature iCM, are limitations of iPSC-derived disease models (123). In addition, another set of challenges exists that is specific to disease modeling. For instance, not all human diseases can be modeled in vitro using iPSC. It is difficult to recapitulate diseases that depend on complex interactions of multiple genetic and environmental factors or on a long incubation period, such as atherosclerosis and congenital heart disease. At the drug screening level, it is not always possible to measure all toxicities or side effects at the cellular level. Investigation of medication-induced mood changes or long-term side effects in remote tissues will still need clinical human studies and post-marketing research. However, advances in bioengineering technologies such as microfluidics (124), high-throughput single-molecule optofluidic analysis (125), and disease on chip technologies 126, 127 are expected to overcome some of these limitations. These novel models and experimental constructs will perfuse drug solutions on iPSC-derivatives, including hepatic, renal, pulmonary, and cardiac cells in series or parallel, to simulate the multisystem interactions within the body and to assess important toxicities and/or side effects.
Summary and Conclusions
iPSC represent a promising tool for cardiovascular regeneration, disease modeling, and drug discovery. Despite some encouraging results in pre-clinical studies using iCM for cardiac regeneration, long-term engraftment remains challenging and the long-term results are unclear. It is possible that these issues may be ameliorated with the use of alternative delivery methods such as iPSC-based patches. Given the low engraftment and other limitations of iCM, some of the initial positive pre-clinical results with iCM should be interpreted with some skepticism. These studies will require long-term follow-up in pre-clinical models and eventually phase I clinical trials. As a new emerging technology, the application of iCM for the treatment of advanced heart failure is maturing rapidly by addressing the major extant challenges, including engraftment, immunogenicity, tumorigenicity, and genetic alterations from reprogramming 42, 43. Innovative molecular and bioengineering techniques to enhance cellular engraftment, reduce immunogenicity, eliminate tumorigenicity, identify paracrine factors, and generate mature and homogeneous iCM have a great potential to lead to successful salvage or regeneration of the injured myocardium and permanent restoration of cardiac function. Similarly, although no clinical trials of iPSC for vascular disease have been initiated, pre-clinical studies support optimism for this potential application. However, a host of issues similar to those that plague the application of iCM for cardiac regeneration in heart failure arise in the application of iEC to the field of vascular regeneration. There is still a strong need to identify novel methods to improve the yield of iPSC, to enhance production consistency and quality, and to improve regenerative effects following transplantation into tissues. The ability to model human disease in a petri dish provides a compelling biomedical rationale to pursue this pioneering research.
Footnotes
Supported by grants from the National Institutes of Health/National Heart, Lung, and Blood Institute Cardiovascular Cell Therapy Research Network (5UM1 HL113456-02, UM1-HL113530, 5UM1-HL087366-07), National Institute of Health HL-78825, National Heart, Lung, and Blood Institute Training Program in Mechanisms and Innovation in Vascular Disease (1T32HL098049-01A1), and National Institutes of Health/National Center for Advancing Translational Sciences (UL1-TR001427 and 1R01 HL125224). The authors have reported that they have no relationships relevant to the contents of this paper to disclose.
References
- 1.Briggs R., King T.J. Transplantation of living nuclei from blastula cells into enucleated frogs’ eggs. Proc Natl Acad Sci U S A. 1952;38:455–463. doi: 10.1073/pnas.38.5.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gurdon J.B., Elsdale T.R., Fischberg M. Sexually mature individuals of Xenopus laevis from the transplantation of single somatic nuclei. Nature. 1958;182:64–65. doi: 10.1038/182064a0. [DOI] [PubMed] [Google Scholar]
- 3.Cowan C.A., Atienza J., Melton D.A., Eggan K. Nuclear reprogramming of somatic cells after fusion with human embryonic stem cells. Science. 2005;309:1369–1373. doi: 10.1126/science.1116447. [DOI] [PubMed] [Google Scholar]
- 4.Tada M., Takahama Y., Abe K., Nakatsuji N., Tada T. Nuclear reprogramming of somatic cells by in vitro hybridization with ES cells. Curr Biol. 2001;11:1553–1558. doi: 10.1016/s0960-9822(01)00459-6. [DOI] [PubMed] [Google Scholar]
- 5.Hochedlinger K., Jaenisch R. Nuclear reprogramming and pluripotency. Nature. 2006;441:1061–1067. doi: 10.1038/nature04955. [DOI] [PubMed] [Google Scholar]
- 6.Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 7.Okita K., Ichisaka T., Yamanaka S. Generation of germline-competent induced pluripotent stem cells. Nature. 2007;448:313–317. doi: 10.1038/nature05934. [DOI] [PubMed] [Google Scholar]
- 8.Takahashi K., Tanabe K., Ohnuki M. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- 9.Yu J., Vodyanik M.A., Smuga-Otto K. Induced pluripotent stem cell lines derived from human somatic cells. Science. 2007;318:1917–1920. doi: 10.1126/science.1151526. [DOI] [PubMed] [Google Scholar]
- 10.Wernig M., Meissner A., Foreman R. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature. 2007;448:318–324. doi: 10.1038/nature05944. [DOI] [PubMed] [Google Scholar]
- 11.Park I.H., Zhao R., West J.A. Reprogramming of human somatic cells to pluripotency with defined factors. Nature. 2008;451:141–146. doi: 10.1038/nature06534. [DOI] [PubMed] [Google Scholar]
- 12.Aasen T., Raya A., Barrero M.J. Efficient and rapid generation of induced pluripotent stem cells from human keratinocytes. Nat Biotechnol. 2008;26:1276–1284. doi: 10.1038/nbt.1503. [DOI] [PubMed] [Google Scholar]
- 13.Brown M.E., Rondon E., Rajesh D. Derivation of induced pluripotent stem cells from human peripheral blood T lymphocytes. PLoS One. 2010;5:e11373. doi: 10.1371/journal.pone.0011373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Loh Y.H., Agarwal S., Park I.H. Generation of induced pluripotent stem cells from human blood. Blood. 2009;113:5476–5479. doi: 10.1182/blood-2009-02-204800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Giorgetti A., Montserrat N., Aasen T. Generation of induced pluripotent stem cells from human cord blood using OCT4 and SOX2. Cell Stem Cell. 2009;5:353–357. doi: 10.1016/j.stem.2009.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ge X., Wang I.N., Toma I. Human amniotic mesenchymal stem cell-derived induced pluripotent stem cells may generate a universal source of cardiac cells. Stem Cells Dev. 2012;21:2798–2808. doi: 10.1089/scd.2011.0435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kim J.B., Greber B., Arauzo-Bravo M.J. Direct reprogramming of human neural stem cells by OCT4. Nature. 2009;461:649–653. doi: 10.1038/nature08436. [DOI] [PubMed] [Google Scholar]
- 18.Sun N., Panetta N.J., Gupta D.M. Feeder-free derivation of induced pluripotent stem cells from adult human adipose stem cells. Proc Natl Acad Sci U S A. 2009;106:15720–15725. doi: 10.1073/pnas.0908450106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhou T., Benda C., Dunzinger S. Generation of human induced pluripotent stem cells from urine samples. Nat Protoc. 2012;7:2080–2089. doi: 10.1038/nprot.2012.115. [DOI] [PubMed] [Google Scholar]
- 20.Mauritz C., Schwanke K., Reppel M. Generation of functional murine cardiac myocytes from induced pluripotent stem cells. Circulation. 2008;118:507–517. doi: 10.1161/CIRCULATIONAHA.108.778795. [DOI] [PubMed] [Google Scholar]
- 21.Narazaki G., Uosaki H., Teranishi M. Directed and systematic differentiation of cardiovascular cells from mouse induced pluripotent stem cells. Circulation. 2008;118:498–506. doi: 10.1161/CIRCULATIONAHA.108.769562. [DOI] [PubMed] [Google Scholar]
- 22.Kawamura M., Miyagawa S., Miki K. Feasibility, safety, and therapeutic efficacy of human induced pluripotent stem cell-derived cardiomyocyte sheets in a porcine ischemic cardiomyopathy model. Circulation. 2012;126(Suppl 1):S29–S37. doi: 10.1161/CIRCULATIONAHA.111.084343. [DOI] [PubMed] [Google Scholar]
- 23.Xiong Q., Ye L., Zhang P. Functional consequences of human induced pluripotent stem cell therapy: myocardial ATP turnover rate in the in vivo swine heart with postinfarction remodeling. Circulation. 2013;127:997–1008. doi: 10.1161/CIRCULATIONAHA.112.000641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Li X., Zhang F., Song G. Intramyocardial injection of pig pluripotent stem cells improves left ventricular function and perfusion: a study in a porcine model of acute myocardial infarction. PLoS One. 2013;8:e66688. doi: 10.1371/journal.pone.0066688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sanchez-Freire V., Lee A.S., Hu S. Effect of human donor cell source on differentiation and function of cardiac induced pluripotent stem cells. J Am Coll Cardiol. 2014;64:436–448. doi: 10.1016/j.jacc.2014.04.056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kamao H., Mandai M., Okamoto S. Characterization of human induced pluripotent stem cell-derived retinal pigment epithelium cell sheets aiming for clinical application. Stem Cell Reports. 2014;2:205–218. doi: 10.1016/j.stemcr.2013.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sanchez Alvarado A., Yamanaka S. Rethinking differentiation: stem cells, regeneration, and plasticity. Cell. 2014;157:110–119. doi: 10.1016/j.cell.2014.02.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ko H.C., Gelb B.D. Concise review: drug discovery in the age of the induced pluripotent stem cell. Stem Cells Transl Med. 2014;3:500–509. doi: 10.5966/sctm.2013-0162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lalit P.A., Hei D.J., Raval A.N., Kamp T.J. Induced pluripotent stem cells for post-myocardial infarction repair: remarkable opportunities and challenges. Circ Res. 2014;114:1328–1345. doi: 10.1161/CIRCRESAHA.114.300556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nelson T.J., Martinez-Fernandez A., Yamada S., Perez-Terzic C., Ikeda Y., Terzic A. Repair of acute myocardial infarction by human stemness factors induced pluripotent stem cells. Circulation. 2009;120:408–416. doi: 10.1161/CIRCULATIONAHA.109.865154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ahmed R.P., Ashraf M., Buccini S., Shujia J., Haider H. Cardiac tumorigenic potential of induced pluripotent stem cells in an immunocompetent host with myocardial infarction. Regen Med. 2011;6:171–178. doi: 10.2217/rme.10.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kim P.J., Mahmoudi M., Ge X. Direct evaluation of myocardial viability and stem cell engraftment demonstrates salvage of the injured myocardium. Circ Res. 2015;116:e40–e50. doi: 10.1161/CIRCRESAHA.116.304668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Zhang Y., Wang D., Chen M., Yang B., Zhang F., Cao K. Intramyocardial transplantation of undifferentiated rat induced pluripotent stem cells causes tumorigenesis in the heart. PLoS One. 2011;6:e19012. doi: 10.1371/journal.pone.0019012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nussbaum J., Minami E., Laflamme M.A. Transplantation of undifferentiated murine embryonic stem cells in the heart: teratoma formation and immune response. FASEB J. 2007;21:1345–1357. doi: 10.1096/fj.06-6769com. [DOI] [PubMed] [Google Scholar]
- 35.Arai T., Kofidis T., Bulte J.W. Dual in vivo magnetic resonance evaluation of magnetically labeled mouse embryonic stem cells and cardiac function at 1.5 T. Magn Reson Med. 2006;55:203–209. doi: 10.1002/mrm.20702. [DOI] [PubMed] [Google Scholar]
- 36.Ahmed R.P., Haider H.K., Buccini S., Li L., Jiang S., Ashraf M. Reprogramming of skeletal myoblasts for induction of pluripotency for tumor-free cardiomyogenesis in the infarcted heart. Circ Res. 2011;109:60–70. doi: 10.1161/CIRCRESAHA.110.240010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Buccini S., Haider K.H., Ahmed R.P., Jiang S., Ashraf M. Cardiac progenitors derived from reprogrammed mesenchymal stem cells contribute to angiomyogenic repair of the infarcted heart. Basic Res Cardiol. 2012;107:301. doi: 10.1007/s00395-012-0301-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Burridge P.W., Matsa E., Shukla P. Chemically defined generation of human cardiomyocytes. Nat Methods. 2014;11:855–860. doi: 10.1038/nmeth.2999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Rulifson E., Matsuura Y., Ariyama M. Abstract 19831: in vivo molecular imaging of human pluripotent stem cell-derived cardiomyocytes in a murine myocardial injury model via a safe harbor integration of a reporter gene. Circulation. 2014;130:A19831. [Google Scholar]
- 40.Pasha Z., Haider H., Ashraf M. Efficient non-viral reprogramming of myoblasts to stemness with a single small molecule to generate cardiac progenitor cells. PLoS One. 2011;6:e23667. doi: 10.1371/journal.pone.0023667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mauritz C., Martens A., Rojas S.V. Induced pluripotent stem cell (iPSC)-derived Flk-1 progenitor cells engraft, differentiate, and improve heart function in a mouse model of acute myocardial infarction. Eur Heart J. 2011;32:2634–2641. doi: 10.1093/eurheartj/ehr166. [DOI] [PubMed] [Google Scholar]
- 42.Dai B., Huang W., Xu M. Reduced collagen deposition in infarcted myocardium facilitates induced pluripotent stem cell engraftment and angiomyogenesis for improvement of left ventricular function. J Am Coll Cardiol. 2011;58:2118–2127. doi: 10.1016/j.jacc.2011.06.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Huang W., Dai B., Wen Z. Molecular strategy to reduce in vivo collagen barrier promotes entry of NCX1 positive inducible pluripotent stem cells (iPSC(NCX1+)) into ischemic (or injured) myocardium. PLoS One. 2013;8:e70023. doi: 10.1371/journal.pone.0070023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Leeper N.J., Hunter A.L., Cooke J.P. Stem cell therapy for vascular regeneration: adult, embryonic, and induced pluripotent stem cells. Circulation. 2010;122:517–526. doi: 10.1161/CIRCULATIONAHA.109.881441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Botham C.M., Bennett W.L., Cooke J.P. Clinical trials of adult stem cell therapy for peripheral artery disease. Methodist Debakey Cardiovasc J. 2013;9:201–205. doi: 10.14797/mdcj-9-4-201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Cooke J.P., Losordo D.W. Modulating the vascular response to limb ischemia: angiogenic and cell therapies. Circ Res. 2015;116:1561–1578. doi: 10.1161/CIRCRESAHA.115.303565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Perin E.C., Murphy M., Cooke J.P. Rationale and design for PACE: patients with intermittent claudication injected with ALDH bright cells. Am Heart J. 2014;168:667–673. doi: 10.1016/j.ahj.2014.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Spinetti G., Kraenkel N., Emanueli C., Madeddu P. Diabetes and vessel wall remodelling: from mechanistic insights to regenerative therapies. Cardiovasc Res. 2008;78:265–273. doi: 10.1093/cvr/cvn039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Asahara T., Kawamoto A. Endothelial progenitor cells for postnatal vasculogenesis. Am J Physiol Cell Physiol. 2004;287:C572–C579. doi: 10.1152/ajpcell.00330.2003. [DOI] [PubMed] [Google Scholar]
- 50.Clayton Z.E., Sadeghipour S., Patel S. Generating induced pluripotent stem cell derived endothelial cells and induced endothelial cells for cardiovascular disease modelling and therapeutic angiogenesis. Int J Cardiol. 2015;197:116–122. doi: 10.1016/j.ijcard.2015.06.038. [DOI] [PubMed] [Google Scholar]
- 51.Choi K.D., Yu J., Smuga-Otto K. Hematopoietic and endothelial differentiation of human induced pluripotent stem cells. Stem Cells. 2009;27:559–567. doi: 10.1634/stemcells.2008-0922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Taura D., Sone M., Homma K. Induction and isolation of vascular cells from human induced pluripotent stem cells—brief report. Arterioscler Thromb Vasc Biol. 2009;29:1100–1103. doi: 10.1161/ATVBAHA.108.182162. [DOI] [PubMed] [Google Scholar]
- 53.Lai W.H., Ho J.C., Chan Y.C. Attenuation of hind-limb ischemia in mice with endothelial-like cells derived from different sources of human stem cells. PLoS One. 2013;8:e57876. doi: 10.1371/journal.pone.0057876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Wang Y., Hu J., Jiao J. Engineering vascular tissue with functional smooth muscle cells derived from human iPS cells and nanofibrous scaffolds. Biomaterials. 2014;35:8960–8969. doi: 10.1016/j.biomaterials.2014.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hendry S.L., 2nd, van der Bogt K.E., Sheikh A.Y. Multimodal evaluation of in vivo magnetic resonance imaging of myocardial restoration by mouse embryonic stem cells. J Thorac Cardiovasc Surg. 2008;136:1028–1037.e1. doi: 10.1016/j.jtcvs.2007.12.053. [DOI] [PubMed] [Google Scholar]
- 56.Hung T.C., Suzuki Y., Urashima T. Multimodality evaluation of the viability of stem cells delivered into different zones of myocardial infarction. Circ Cardiovasc Imaging. 2008;1:6–13. doi: 10.1161/CIRCIMAGING.108.767343. [DOI] [PubMed] [Google Scholar]
- 57.Terrovitis J., Stuber M., Youssef A. Magnetic resonance imaging overestimates ferumoxide-labeled stem cell survival after transplantation in the heart. Circulation. 2008;117:1555–1562. doi: 10.1161/CIRCULATIONAHA.107.732073. [DOI] [PubMed] [Google Scholar]
- 58.Sanganalmath S.K., Bolli R. Cell therapy for heart failure: a comprehensive overview of experimental and clinical studies, current challenges, and future directions. Circ Res. 2013;113:810–834. doi: 10.1161/CIRCRESAHA.113.300219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Li Z., Lee A., Huang M. Imaging survival and function of transplanted cardiac resident stem cells. J Am Coll Cardiol. 2009;53:1229–1240. doi: 10.1016/j.jacc.2008.12.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Keith M.C., Bolli R. “String theory” of c-kit(pos) cardiac cells: a new paradigm regarding the nature of these cells that may reconcile apparently discrepant results. Circ Res. 2015;116:1216–1230. doi: 10.1161/CIRCRESAHA.116.305557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Orlic D., Kajstura J., Chimenti S. Bone marrow cells regenerate infarcted myocardium. Nature. 2001;410:701–705. doi: 10.1038/35070587. [DOI] [PubMed] [Google Scholar]
- 62.Quaini F., Urbanek K., Beltrami A.P. Chimerism of the transplanted heart. N Engl J Med. 2002;346:5–15. doi: 10.1056/NEJMoa012081. [DOI] [PubMed] [Google Scholar]
- 63.Beltrami A.P., Urbanek K., Kajstura J. Evidence that human cardiac myocytes divide after myocardial infarction. N Engl J Med. 2001;344:1750–1757. doi: 10.1056/NEJM200106073442303. [DOI] [PubMed] [Google Scholar]
- 64.Kajstura J., Leri A., Finato N., Di Loreto C., Beltrami C.A., Anversa P. Myocyte proliferation in end-stage cardiac failure in humans. Proc Natl Acad Sci U S A. 1998;95:8801–8805. doi: 10.1073/pnas.95.15.8801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Pittenger M.F., Mackay A.M., Beck S.C. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284:143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 66.Messina E., De Angelis L., Frati G. Isolation and expansion of adult cardiac stem cells from human and murine heart. Circ Res. 2004;95:911–921. doi: 10.1161/01.RES.0000147315.71699.51. [DOI] [PubMed] [Google Scholar]
- 67.Thomson J.A., Itskovitz-Eldor J., Shapiro S.S. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282:1145–1147. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- 68.Bergmann O., Bhardwaj R.D., Bernard S. Evidence for cardiomyocyte renewal in humans. Science. 2009;324:98–102. doi: 10.1126/science.1164680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.van Berlo J.H., Kanisicak O., Maillet M. c-kit+ cells minimally contribute cardiomyocytes to the heart. Nature. 2014;509:337–341. doi: 10.1038/nature13309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Asahara T., Murohara T., Sullivan A. Isolation of putative progenitor endothelial cells for angiogenesis. Science. 1997;275:964–967. doi: 10.1126/science.275.5302.964. [DOI] [PubMed] [Google Scholar]
- 71.Asahara T., Masuda H., Takahashi T. Bone marrow origin of endothelial progenitor cells responsible for postnatal vasculogenesis in physiological and pathological neovascularization. Circ Res. 1999;85:221–228. doi: 10.1161/01.res.85.3.221. [DOI] [PubMed] [Google Scholar]
- 72.Huang N.F., Niiyama H., Peter C. Embryonic stem cell-derived endothelial cells engraft into the ischemic hindlimb and restore perfusion. Arterioscler Thromb Vasc Biol. 2010;30:984–991. doi: 10.1161/ATVBAHA.110.202796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Hussein S.M., Batada N.N., Vuoristo S. Copy number variation and selection during reprogramming to pluripotency. Nature. 2011;471:58–62. doi: 10.1038/nature09871. [DOI] [PubMed] [Google Scholar]
- 74.Lister R., Pelizzola M., Kida Y.S. Hotspots of aberrant epigenomic reprogramming in human induced pluripotent stem cells. Nature. 2011;471:68–73. doi: 10.1038/nature09798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Li C., Klco J.M., Helton N.M. Genetic heterogeneity of induced pluripotent stem cells: results from 24 clones derived from a single C57BL/6 mouse. PLoS One. 2015;10:e0120585. doi: 10.1371/journal.pone.0120585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Laurent L.C., Ulitsky I., Slavin I. Dynamic changes in the copy number of pluripotency and cell proliferation genes in human ESCs and iPSCs during reprogramming and time in culture. Cell Stem Cell. 2011;8:106–118. doi: 10.1016/j.stem.2010.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cyranoski D. Stem cells cruise to clinic. Nature. 2013;494:413. doi: 10.1038/494413a. [DOI] [PubMed] [Google Scholar]
- 78.Kim K., Doi A., Wen B. Epigenetic memory in induced pluripotent stem cells. Nature. 2010;467:285–290. doi: 10.1038/nature09342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Robinton D.A., Daley G.Q. The promise of induced pluripotent stem cells in research and therapy. Nature. 2012;481:295–305. doi: 10.1038/nature10761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Nakagawa M., Koyanagi M., Tanabe K. Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat Biotechnol. 2008;26:101–106. doi: 10.1038/nbt1374. [DOI] [PubMed] [Google Scholar]
- 81.Schlaeger T.M., Daheron L., Brickler T.R. A comparison of non-integrating reprogramming methods. Nat Biotechnol. 2015;33:58–63. doi: 10.1038/nbt.3070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kattman S.J., Witty A.D., Gagliardi M. Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac differentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell. 2011;8:228–240. doi: 10.1016/j.stem.2010.12.008. [DOI] [PubMed] [Google Scholar]
- 83.Uosaki H., Fukushima H., Takeuchi A. Efficient and scalable purification of cardiomyocytes from human embryonic and induced pluripotent stem cells by VCAM1 surface expression. PLoS One. 2011;6:e23657. doi: 10.1371/journal.pone.0023657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lian X., Zhang J., Azarin S.M. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat Protocols. 2013;8:162–175. doi: 10.1038/nprot.2012.150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Fuerstenau-Sharp M., Zimmermann M.E., Stark K. Generation of highly purified human cardiomyocytes from peripheral blood mononuclear cell-derived induced pluripotent stem cells. PLoS One. 2015;10:e0126596. doi: 10.1371/journal.pone.0126596. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Rufaihah A.J., Huang N.F., Kim J. Human induced pluripotent stem cell-derived endothelial cells exhibit functional heterogeneity. Am J Transl Res. 2013;5:21–35. [PMC free article] [PubMed] [Google Scholar]
- 87.Ribeiro A.J., Ang Y.S., Fu J.D. Contractility of single cardiomyocytes differentiated from pluripotent stem cells depends on physiological shape and substrate stiffness. Proc Natl Acad Sci U S A. 2015;112:12705–12710. doi: 10.1073/pnas.1508073112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Zhao T., Zhang Z.N., Rong Z., Xu Y. Immunogenicity of induced pluripotent stem cells. Nature. 2011;474:212–215. doi: 10.1038/nature10135. [DOI] [PubMed] [Google Scholar]
- 89.de Almeida P.E., Meyer E.H., Kooreman N.G. Transplanted terminally differentiated induced pluripotent stem cells are accepted by immune mechanisms similar to self-tolerance. Nat Commun. 2014;5:3903. doi: 10.1038/ncomms4903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Araki R., Uda M., Hoki Y. Negligible immunogenicity of terminally differentiated cells derived from induced pluripotent or embryonic stem cells. Nature. 2013;494:100–104. doi: 10.1038/nature11807. [DOI] [PubMed] [Google Scholar]
- 91.Guha P., Morgan J.W., Mostoslavsky G., Rodrigues N.P., Boyd A.S. Lack of immune response to differentiated cells derived from syngeneic induced pluripotent stem cells. Cell Stem Cell. 2013;12:407–412. doi: 10.1016/j.stem.2013.01.006. [DOI] [PubMed] [Google Scholar]
- 92.Turner M., Leslie S., Martin N.G. Toward the development of a global induced pluripotent stem cell library. Cell Stem Cell. 2013;13:382–384. doi: 10.1016/j.stem.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 93.Taylor C.J., Peacock S., Chaudhry A.N., Bradley J.A., Bolton E.M. Generating an iPSC bank for HLA-matched tissue transplantation based on known donor and recipient HLA types. Cell Stem Cell. 2012;11:147–152. doi: 10.1016/j.stem.2012.07.014. [DOI] [PubMed] [Google Scholar]
- 94.Okita K., Matsumura Y., Sato Y. A more efficient method to generate integration-free human iPS cells. Nat Methods. 2011;8:409–412. doi: 10.1038/nmeth.1591. [DOI] [PubMed] [Google Scholar]
- 95.Gourraud P.A., Gilson L., Girard M., Peschanski M. The role of human leukocyte antigen matching in the development of multiethnic “haplobank” of induced pluripotent stem cell lines. Stem Cells. 2012;30:180–186. doi: 10.1002/stem.772. [DOI] [PubMed] [Google Scholar]
- 96.Tsuji O., Miura K., Okada Y. Therapeutic potential of appropriately evaluated safe-induced pluripotent stem cells for spinal cord injury. Proc Natl Acad Sci U S A. 2010;107:12704–12709. doi: 10.1073/pnas.0910106107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Takemoto S.K., Terasaki P.I., Gjertson D.W., Cecka J.M. Twelve years' experience with national sharing of HLA-matched cadaveric kidneys for transplantation. N Engl J Med. 2000;343:1078–1084. doi: 10.1056/NEJM200010123431504. [DOI] [PubMed] [Google Scholar]
- 98.Lee A.S., Tang C., Rao M.S., Weissman I.L., Wu J.C. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat Med. 2013;19:998–1004. doi: 10.1038/nm.3267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Ban H., Nishishita N., Fusaki N. Efficient generation of transgene-free human induced pluripotent stem cells (iPSCs) by temperature-sensitive Sendai virus vectors. Proc Natl Acad Sci U S A. 2011;108:14234–14239. doi: 10.1073/pnas.1103509108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Simpson D.L., Wehman B., Galat Y. Engineering patient-specific valves using stem cells generated from skin biopsy specimens. Ann Thorac Surg. 2014;98:947–954. doi: 10.1016/j.athoracsur.2014.04.075. [DOI] [PubMed] [Google Scholar]
- 101.Parker K.K., Norenberg M.D., Vernadakis A. “Transdifferentiation” of C6 glial cells in culture. Science. 1980;208:179–181. doi: 10.1126/science.6102413. [DOI] [PubMed] [Google Scholar]
- 102.Ieda M., Fu J.D., Delgado-Olguin P. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell. 2010;142:375–386. doi: 10.1016/j.cell.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Qian L., Huang Y., Spencer C.I. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature. 2012;485:593–598. doi: 10.1038/nature11044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Song K., Nam Y.J., Luo X. Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature. 2012;485:599–604. doi: 10.1038/nature11139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Srivastava D., Yu P. Recent advances in direct cardiac reprogramming. Curr Opin Genet Dev. 2015;34:77–81. doi: 10.1016/j.gde.2015.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Yoshida Y., Yamanaka S. Labor pains of new technology: direct cardiac reprogramming. Circ Res. 2012;111:3–4. doi: 10.1161/CIRCRESAHA.112.271445. [DOI] [PubMed] [Google Scholar]
- 107.Margariti A., Winkler B., Karamariti E. Direct reprogramming of fibroblasts into endothelial cells capable of angiogenesis and reendothelialization in tissue-engineered vessels. Proc Natl Acad Sci U S A. 2012;109:13793–13798. doi: 10.1073/pnas.1205526109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Lee J., Sayed N., Hunter A. Activation of innate immunity is required for efficient nuclear reprogramming. Cell. 2012;151:547–558. doi: 10.1016/j.cell.2012.09.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sayed N., Wong W.T., Ospino F. Transdifferentiation of human fibroblasts to endothelial cells: role of innate immunity. Circulation. 2015;131:300–309. doi: 10.1161/CIRCULATIONAHA.113.007394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Park I.H., Arora N., Huo H. Disease-specific induced pluripotent stem cells. Cell. 2008;134:877–886. doi: 10.1016/j.cell.2008.07.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Moretti A., Bellin M., Welling A. Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N Engl J Med. 2010;363:1397–1409. doi: 10.1056/NEJMoa0908679. [DOI] [PubMed] [Google Scholar]
- 112.Carvajal-Vergara X., Sevilla A., D'Souza S.L. Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syndrome. Nature. 2010;465:808–812. doi: 10.1038/nature09005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Sun N., Yazawa M., Liu J. Patient-specific induced pluripotent stem cells as a model for familial dilated cardiomyopathy. Sci Transl Med. 2012;4:130ra47. doi: 10.1126/scitranslmed.3003552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Bhatia S.N., Ingber D.E. Microfluidic organs-on-chips. Nat Biotechnol. 2014;32:760–772. doi: 10.1038/nbt.2989. [DOI] [PubMed] [Google Scholar]
- 115.Wang G., McCain M.L., Yang L. Modeling the mitochondrial cardiomyopathy of Barth syndrome with induced pluripotent stem cell and heart-on-chip technologies. Nat Med. 2014;20:616–623. doi: 10.1038/nm.3545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Jiang Y., Habibollah S., Tilgner K. An induced pluripotent stem cell model of hypoplastic left heart syndrome (HLHS) reveals multiple expression and functional differences in HLHS-derived cardiac myocytes. Stem Cells Transl Med. 2014;3:416–423. doi: 10.5966/sctm.2013-0105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Schenone M., Dancik V., Wagner B.K., Clemons P.A. Target identification and mechanism of action in chemical biology and drug discovery. Nat Chem Biol. 2013;9:232–240. doi: 10.1038/nchembio.1199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Lee G., Ramirez C.N., Kim H. Large-scale screening using familial dysautonomia induced pluripotent stem cells identifies compounds that rescue IKBKAP expression. Nat Biotechnol. 2012;30:1244–1248. doi: 10.1038/nbt.2435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Guo L., Coyle L., Abrams R.M., Kemper R., Chiao E.T., Kolaja K.L. Refining the human iPSC-cardiomyocyte arrhythmic risk assessment model. Toxicol Sci. 2013;136:581–594. doi: 10.1093/toxsci/kft205. [DOI] [PubMed] [Google Scholar]
- 120.Navarrete E.G., Liang P., Lan F. Screening drug-induced arrhythmia [corrected] using human induced pluripotent stem cell-derived cardiomyocytes and low-impedance microelectrode arrays. Circulation. 2013;128(Suppl 11):S3–S13. doi: 10.1161/CIRCULATIONAHA.112.000570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Takayama K., Morisaki Y., Kuno S. Prediction of interindividual differences in hepatic functions and drug sensitivity by using human iPS-derived hepatocytes. Proc Natl Acad Sci U S A. 2014;111:16772–16777. doi: 10.1073/pnas.1413481111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Adams W.J., Zhang Y., Cloutier J. Functional vascular endothelium derived from human induced pluripotent stem cells. Stem Cell Reports. 2013;1:105–113. doi: 10.1016/j.stemcr.2013.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Soldner F., Jaenisch R. iPSC disease modeling. Science. 2012;338:1155–1156. doi: 10.1126/science.1227682. [DOI] [PubMed] [Google Scholar]
- 124.Psaltis D., Quake S.R., Yang C. Developing optofluidic technology through the fusion of microfluidics and optics. Nature. 2006;442:381–386. doi: 10.1038/nature05060. [DOI] [PubMed] [Google Scholar]
- 125.Kim S., Streets A.M., Lin R.R., Quake S.R., Weiss S., Majumdar D.S. High-throughput single-molecule optofluidic analysis. Nat Methods. 2011;8:242–245. doi: 10.1038/nmeth.1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Agarwal A., Goss J.A., Cho A., McCain M.L., Parker K.K. Microfluidic heart on a chip for higher throughput pharmacological studies. Lab Chip. 2013;13:3599–3608. doi: 10.1039/c3lc50350j. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Grosberg A., Alford P.W., McCain M.L., Parker K.K. Ensembles of engineered cardiac tissues for physiological and pharmacological study: heart on a chip. Lab Chip. 2011;11:4165–4173. doi: 10.1039/c1lc20557a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Ge X., Ren Y., Bartulos O. Modeling supravalvular aortic stenosis syndrome with human induced pluripotent stem cells. Circulation. 2012;126:1695–1704. doi: 10.1161/CIRCULATIONAHA.112.116996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Asimaki A., Kapoor S., Plovie E. Identification of a new modulator of the intercalated disc in a zebrafish model of arrhythmogenic cardiomyopathy. Sci Transl Med. 2014;6:240ra74. doi: 10.1126/scitranslmed.3008008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ma D., Wei H., Lu J. Generation of patient-specific induced pluripotent stem cell-derived cardiomyocytes as a cellular model of arrhythmogenic right ventricular cardiomyopathy. Eur Heart J. 2013;34:1122–1133. doi: 10.1093/eurheartj/ehs226. [DOI] [PubMed] [Google Scholar]
- 131.Kim C., Wong J., Wen J. Studying arrhythmogenic right ventricular dysplasia with patient-specific iPSCs. Nature. 2013;494:105–110. doi: 10.1038/nature11799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Lan F., Lee A.S., Liang P. Abnormal calcium handling properties underlie familial hypertrophic cardiomyopathy pathology in patient-specific induced pluripotent stem cells. Cell Stem Cell. 2013;12:101–113. doi: 10.1016/j.stem.2012.10.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Hick A., Wattenhofer-Donze M., Chintawar S. Neurons and cardiomyocytes derived from induced pluripotent stem cells as a model for mitochondrial defects in Friedreich's ataxia. Dis Model Mech. 2013;6:608–621. doi: 10.1242/dmm.010900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Huang H.P., Chen P.H., Hwu W.L. Human Pompe disease-induced pluripotent stem cells for pathogenesis modeling, drug testing and disease marker identification. Hum Mol Genet. 2011;20:4851–4864. doi: 10.1093/hmg/ddr424. [DOI] [PubMed] [Google Scholar]
- 135.Kujala K., Paavola J., Lahti A. Cell model of catecholaminergic polymorphic ventricular tachycardia reveals early and delayed afterdepolarizations. PLoS One. 2012;7:e44660. doi: 10.1371/journal.pone.0044660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Jung C.B., Moretti A., Mederos y Schnitzler M. Dantrolene rescues arrhythmogenic RYR2 defect in a patient-specific stem cell model of catecholaminergic polymorphic ventricular tachycardia. EMBO Mol Med. 2012;4:180–191. doi: 10.1002/emmm.201100194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Novak A., Barad L., Zeevi-Levin N. Cardiomyocytes generated from CPVTD307H patients are arrhythmogenic in response to β-adrenergic stimulation. J Cell Mol Med. 2012;16:468–482. doi: 10.1111/j.1582-4934.2011.01476.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Itzhaki I., Maizels L., Huber I. Modeling of catecholaminergic polymorphic ventricular tachycardia with patient-specific human-induced pluripotent stem cells. J Am Coll Cardiol. 2012;60:990–1000. doi: 10.1016/j.jacc.2012.02.066. [DOI] [PubMed] [Google Scholar]
- 139.Itzhaki I., Maizels L., Huber I. Modelling the long QT syndrome with induced pluripotent stem cells. Nature. 2011;471:225–229. doi: 10.1038/nature09747. [DOI] [PubMed] [Google Scholar]
- 140.Lahti A.L., Kujala V.J., Chapman H. Model for long QT syndrome type 2 using human iPS cells demonstrates arrhythmogenic characteristics in cell culture. Dis Model Mech. 2012;5:220–230. doi: 10.1242/dmm.008409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Matsa E., Rajamohan D., Dick E. Drug evaluation in cardiomyocytes derived from human induced pluripotent stem cells carrying a long QT syndrome type 2 mutation. Eur Heart J. 2011;32:952–962. doi: 10.1093/eurheartj/ehr073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Yazawa M., Hsueh B., Jia X. Using induced pluripotent stem cells to investigate cardiac phenotypes in Timothy syndrome. Nature. 2011;471:230–234. doi: 10.1038/nature09855. [DOI] [PMC free article] [PubMed] [Google Scholar]