

Abstract
Background
In breast cancer, BRCA promoter hypermethylation and BRCA germline mutations are said to occur together rarely, but this property has not yet been translated into a clinical test. Our aim in this study was to investigate the diagnostic value of BRCA1/2 methylation in distinguishing breast carcinomas of BRCA1 and BRCA2 germline mutation carriers from sporadic breast carcinomas using a recently developed BRCA methylation assay based on methylation-specific multiplex ligation-dependent probe amplification (MS-MLPA).
Methods
MS-MLPAs were performed to assess BRCA1 and BRCA2 methylation in breast carcinoma tissues from 39 BRCA1 and 33 BRCA2 germline mutation carriers, 80 patients with sporadic breast cancer, and normal breast tissues from 5 BRCA1 and 4 BRCA2 mutation carriers and 5 nonmutation carriers.
Results
Methylation frequencies varied considerably between CpG sites across the BRCA1 and BRCA2 promoters. Some CpG sites were methylated more frequently in BRCA1/2-related than in sporadic carcinomas, whereas other CpG sites were methylated more frequently in sporadic carcinomas, with large variances in sensitivity and specificity as a consequence.
Conclusions
The diagnostic value of BRCA promoter methylation analysis in distinguishing BRCA1/2-related from sporadic breast carcinomas seems to be considerably dependent on the targeted CpG sites. These findings are important for adequate use of BRCA methylation analysis as a prescreening tool for BRCA germline genetic testing or to identify BRCAness patients who may benefit from targeted therapies such as poly(adenosine diphosphate-ribose) polymerase inhibitors.
Keywords: Breast cancer, BRCA, methylation, MS-MLPA
Background
Breast cancer is the most frequent cancer type in women worldwide [1]. In about 5–10%, breast cancer occurs in a hereditary setting, most commonly due to BRCA1 or BRCA2 germline mutations, which lead to a 40–80% lifetime risk of developing breast cancer as well as a 30–40% lifetime risk of ovarian cancer development [2–8]. Promoter hypermethylation plays an important role in carcinogenesis of several organs, including the breast, because hypermethylation of cytosine phosphate guanine (CpG) sites in promoter regions may lead to downregulation of tumor suppressor genes [9–15]. It has been proposed in the literature that BRCA promoter hypermethylation takes place almost exclusively in the sporadic setting and only rarely occurs in patients with an underlying BRCA1 or BRCA2 germline mutation [16–26]. This is potentially clinically important because promoter methylation assays could then serve as prescreening tests when a hereditary nature is suspected, obviating the need for germline mutation analysis in cases of promoter methylation. However, for routine testing, more confirmation is mandatory, such as with regard to the best CpG sites to target, and a robust assay needs to be at hand that works on small amounts of fragmented DNA from formalin-fixed, paraffin-embedded (FFPE) tumor material. The latter is also important in view of the growing need to test for BRCA1 and BRCA2 (BRCA1/2) promoter methylation as a sign of BRCAness, which may provide an indication for treatment with poly(adenosine diphosphate-ribose) polymerase (PARP) inhibitors [27–31]. Methylation-specific multiplex ligation-dependent probe amplification (MS-MLPA) is a rapid, robust, and inexpensive multiplex methylation test that works well on small amounts of DNA derived from FFPE tissues. The aim of this study was to investigate the diagnostic value of BRCA1/2 promoter methylation in distinguishing breast carcinomas from BRCA1 and BRCA2 germline mutation carriers (BRCA1/2-related breast carcinomas) and sporadic breast carcinomas using a recently developed BRCA methylation MS-MLPA assay. In other words, we sought to determine to what extent BRCA1/2 promoter methylation can be detected in BRCA1/2-related compared with sporadic breast carcinomas.
Methods
Patient material
FFPE tissues of 39 BRCA1 and 33 BRCA2 germline mutation-related breast cancer resection specimens (BRCA1/2-C) were derived from the pathology archives at the University Medical Center Utrecht, University Medical Center Groningen, VU University Medical Center Amsterdam, and local hospitals around Utrecht, The Netherlands. Also, FFPE tissues of prophylactic mastectomy specimens of five BRCA1 and four BRCA2 germline mutation carriers (BRCA1/2-N) were derived from the pathology archives of the University Medical Center Utrecht. BRCA status had been confirmed through mutation analysis at a medical genetics department within The Netherlands after informed consent. For comparison, FFPE tissues of 80 breast cancer resection (Sporadic-C) and 5 breast reduction samples (non-BRCA-related-N) from women not tested for a BRCA mutation were derived from the pathology archive of the University Medical Center Utrecht. These women did not receive BRCA germline mutation testing, because there was no clinical suspicion of a hereditary nature. No further inclusion or exclusion criteria were applied. From the tissue blocks, 4-μm-thick sections were cut and stained with hematoxylin and eosin. Tumor characterization, grading according to the modified Bloom-Richardson grading system [32], and scoring of immunohistochemical staining were performed by an experienced breast pathologist (PJvD), who was blinded to mutation status. Estrogen receptor (ER) and progesterone receptor (PR) immunohistochemical staining was considered positive when ≥10% of the tumor cells showed expression, regardless of intensity. Human epidermal growth factor receptor 2 (HER2) was scored according to the HercepTest scoring system (Dako, Glostrup, Denmark) for breast cancer, where only a 3+ score was considered positive. The clinicopathological characteristics are provided in Table 1.
Table 1.
Clinicopathological characteristics of included breast samples
| BRCA1-C | BRCA2-C | Sporadic-C | BRCA1-N | BRCA2-N | Non-BRCA- related-N | p ValueA | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | n | % | n | % | ||
| Number of samples (%) | 39 | 23.5 | 33 | 19.9 | 80 | 48.2 | 5 | 3 | 4 | 2.4 | 5 | 3 | |
| Age, years | 9.56°10−9*ǂ | ||||||||||||
| Median | 43 | N/A | 46 | N/A | 58 | N/A | 31 | N/A | 36.5 | N/A | 22 | N/A | 0.549B |
| Range | 30–80 | N/A | 21–69 | N/A | 29–86 | N/A | 29–33 | N/A | 35–38 | N/A | 18–52 | N/A | |
| GradeC | 0.00007*ǂǂ | ||||||||||||
| 1 | 0 | 0 | 1 | 3 | 20 | 25.3 | |||||||
| 2 | 9 | 23.1 | 10 | 30.3 | 25 | 31.6 | |||||||
| 3 | 30 | 76.9 | 22 | 66.7 | 34 | 43 | |||||||
| Tumor type | 0.230ǂǂ | ||||||||||||
| Ductal | 34 | 87.2 | 29 | 87.9 | 69 | 86.3 | |||||||
| Lobular | 2 | 5.1 | 2 | 6.1 | 10 | 12.5 | |||||||
| Other | 3 | 7.7 | 2 | 6.1 | 1 | 1.3 | |||||||
| ER | 0.0004*ǂǂǂ | ||||||||||||
| Negative | 26 | 66.7 | 8 | 24.2 | 23 | 28.8 | |||||||
| Positive | 13 | 33.3 | 25 | 75.8 | 57 | 71.3 | |||||||
| PR | 0.001*ǂǂǂ | ||||||||||||
| Negative | 29 | 74.4 | 17 | 51.5 | 30 | 37.5 | |||||||
| Positive | 10 | 25.6 | 16 | 48.5 | 50 | 62.5 | |||||||
| HER2 | 0.68ǂǂ | ||||||||||||
| Negative | 38 | 97.4 | 32 | 97 | 75 | 93.8 | |||||||
| Positive | 1 | 2.6 | 1 | 3 | 5 | 6.3 | |||||||
Abbreviations: BRCA1-C Breast carcinomas from BRCA1 germline mutation carriers, BRCA2-C Breast carcinomas from BRCA2 germline mutation carriers, BRCA1/2-N Normal breast tissue from BRCA1 and BRCA2 germline mutation carriers, ER Estrogen receptor, Non-BRCA-related-N Normal breast tissue from patients not known to have a BRCA1 or BRCA2 germline mutation, HER2 Human epidermal growth factor receptor 2, N/A Not available, PR Progesterone receptor, Sporadic-C Sporadic breast carcinoma
ATesting BRCA1-C and BRCA2-C together against Sporadic-C
BTesting BRCA1-N and BRCA2-N together against non-BRCA-related N
CFor 1 of 80 sporadic breast cancer cases, the grade was unknown
ǂMann-Whitney U test
ǂǂFisher’s exact test
ǂǂǂPearson’s chi-square test
*Statistically significant (two-sided p value <0.05)
DNA isolation
Normal breast and breast cancer tissues were harvested from 10×10-μm-thick and 4×4-μm-thick tissue sections, respectively. Areas with necrosis, preinvasive lesions, and extensive inflammation were avoided. DNA isolation was performed by overnight incubation at 56 °C in lysis buffer (50 mM Tris-HCl, pH 8.0, 0.5% Tween 20) with proteinase K (10 mg/ml; Roche, Basel, Switzerland). Proteinase K was deactivated by boiling for 10 minutes. After centrifugation for 2 minutes at 14,000 rpm, the supernatant was collected for further analysis. DNA content was measured using an ND-1000 spectrophotometer (NanoDrop Products, Wilmington, DE, USA).
Methylation analysis
Five microliters of supernatant with a DNA concentration between 50 and 500 ng/μl were used for MS-MLPA analysis according to the manufacturer’s instructions, using the ME053 BRCA1-BRCA2 X1-0914 methylation assay (MRC-Holland, Amsterdam, The Netherlands). When the DNA concentration exceeded 500 ng/μl, the input volume was adjusted proportionally. The ME053 methylation assay contains three and four probes to detect BRCA1 and BRCA2 promoter methylation, respectively, enabling methylation status determination of three CpG sites in the BRCA1 promoter region and five CpG sites in the BRCA2 promoter region (see Table 2 and Fig. 1 for further details). The MS-MLPA principle and analytical procedure are described elsewhere [33], and the technique has been shown to be reliable for methylation assessment [33–37]. Samples that were 100% methylated (SssI methyltransferase-treated MDA-MB-231 and A549 cells) were used as positive controls, and normal peripheral blood samples were used as negative controls. No template controls were included. Moreover, the methylation assay included two digestion (methylation) control probes.
Table 2.
Methylation probe characteristics of the methylation-specific multiplex ligation-dependent probe amplification assay (ME053 BRCA1-BRCA2 X1-0914; MRC-Holland)
| Gene | Probe ID | Length | Chromosome position | 5′ Probe | 3′ Probe | Start | End | CpG site | CpG loci ID |
|---|---|---|---|---|---|---|---|---|---|
| BRCA1 | BRCA1.1 | 165 | 17q21.31 | CCTCTGAGAGGCTGCTGCTTAGCGGTAGCCCCTT | GGTTTCCGTGGCAACGGAAAAGCGCGGGAATTACAGA | 41277415 | 41277483 | 41277429 | cg04110421 |
| BRCA1 | BRCA1.2 | 230 | 17q21.31 | CATGCATCTGAGAAACCCCACAGCCTGTCCCCCGTCCAGGAA | GTCTCAGCGAGCTCACGCCGCGCAGTCGCAGTTT | 41277407 | 41321886 | 41277395 | cg16630982 |
| BRCA1 | BRCA1.3 | 252 | 17q21.31 | GTGGGGTTTCTCAGATAACTGGGCCCCTGC | GCTCAGGAGGCCTTCACCCTCTGCTCTGGGTAAAGGT | 41277286 | 41277352 | 41277323 | cg08993267 |
| BRCA2 | BRCA2.1 | 130 | 13q13.1 | CCATCTTGTGGCGCGAGCTTCTGAAACTA | GGCGGCAGAGGCGGAGCCGCTGTGGCACTGCT | 32889614 | 32889670 | 32889621 | N/A |
| BRCA2 | BRCA2.2 | 149 | 13q13.1 | TGCGGGTTAGTGGTGGTGGTAGTGGGTT | GGGACGAGCGCGTCTTCCGCAGTCCCAGTCCAGCGTGG | 32889801 | 32889865 | 32889836 | N/A |
| BRCA2 | BRCA2.3 | 160 | 13q13.1 | CCTCTGAGAGGCTGCTGCTTAGCGGTAGCCCCTT | GGTTTCCGTGGCAACGGAAAAGCGCGGGAATTACAGA | 32889665 | 32889746 | 32889672 + 32889683 | N/A |
| BRCA2 | BRCA2.4 | 217 | 13q13.1 | CTTCCGGGTGGTGCGTGTGCTGCGTGTCGC | GTCACGGCGTCACGTGGCCAGCGCGGGCTTGT | 32889557 | 32889618 | 2889608 | N/A |
N/A Not available
The chromosomal locations are based upon GRCh37/hg19. The first nucleotides of the 5′ probe may not be complementary to the target DNA, because it contains the stuffer sequence. The CpG loci ID are as determined by the Infinium HumanMethylation450 BeadChip Array (Illumina, San Diego, CA, USA)
Fig. 1.
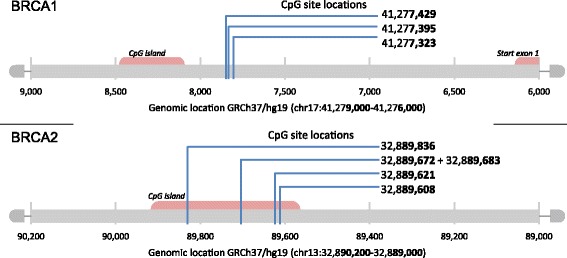
Overview of the targeted cytosine phosphate guanine (CpG) sites in the BRCA1 and BRCA2 promoter regions by methylation analysis of the selected studies
Coffalyser.Net software (MRC-Holland) was used for methylation data analysis. Quality control showed that the results of the control probes and control samples were adequate. The methylation percentage cutoff per probe was set at the highest methylation percentage value in normal breast tissues from nonmutation carriers (non-BRCA-related N), ranging from >15% to >19% (see also Fig. 2). Moreover, the cumulative methylation index (CMI) was calculated as the sum of the methylation percentage of all methylation probes. MS-MLPA analysis was performed by SV and CBM, who were blinded to mutation status.
Fig. 2.
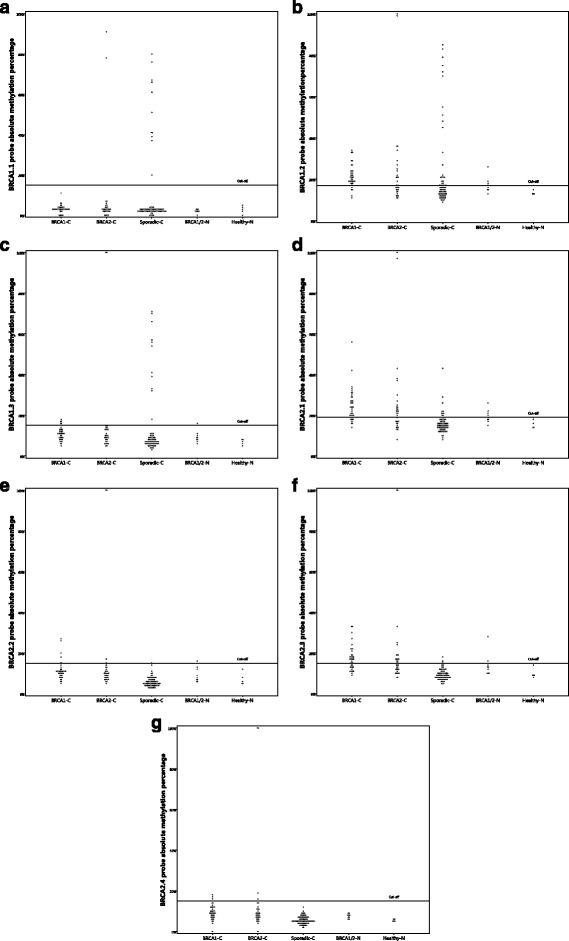
a–g Distribution of absolute methylation percentages for all BRCA1 and BRCA2 methylation probes. BRCA1-C Breast carcinomas from BRCA1 germline mutation carriers, BRCA2-C Breast carcinomas from BRCA2 germline mutation carriers, BRCA1/2-N Normal breast tissue from BRCA1 and BRCA2 germline mutation carriers, Sporadic-C Sporadic breast carcinoma
Statistical analysis
Statistical analyses were performed using IBM SPSS Statistics version 23.0 software (IBM, Armonk, NY, USA). Associations between absolute methylation percentages, CMI or age, and mutation status (BRCA1/2-related carcinomas versus sporadic carcinomas) were assessed by the Mann-Whitney U test. Associations between dichotomized BRCA promoter methylation and mutation status or other clinicopathological characteristics were assessed by Pearson’s chi-square test or Fisher’s exact test. Sensitivity and specificity were calculated. Correlations between CMI and age were assessed using the Spearman’s rho correlation coefficient. The level of significance used was set at a two-sided p value <0.05.
Correlation between BRCA1/2 methylation and messenger RNA expression
The Wanderer tool was used to assess the correlation between BRCA1/2 methylation and messenger RNA (mRNA) expression. This tool was created on the basis of data from The Cancer Genome Atlas (TCGA) Research Network [38]. The Infinium 450K HumanMethylation Array (Illumina, San Diego, CA, USA) was selected as the methylation data type, and Spearman’s correlation coefficient was selected as the correlation method.
Results
BRCA1 promoter methylation in BRCA1/2-related and sporadic breast carcinomas
The absolute methylation percentages and their distribution varied considerably between the three BRCA1 methylation probes (Table 3, Fig. 2). For the BRCA1.2 and BRCA1.3 probes, BRCA1/2-C showed significantly higher median methylation percentages than Sporadic-C (p = 0.00006 and p = 0.00003, respectively). The dichotomized results are shown in Table 4. BRCA1/2-C showed significantly less frequent methylation with the BRCA1.1 probe (p = 0.019), but significantly more frequent methylation with the BRCA1.2 probe (p = 0.000009). Methylation of at least one of the three BRCA1 methylation probes was seen in 46 (63.9%) of 72 BRCA1/2-C compared with 22 (27.5%) of 80 Sporadic-C (p = 0.000009). The sensitivity and specificity of the BRCA1 methylation probes in distinguishing BRCA1/2-C from Sporadic-C are shown in Table 5. The calculation of the sensitivity and specificity differs between probes owing to differences in methylation frequencies between BRCA1/2-C and Sporadic-C (see Table 4 and explanation in Table 5). The BRCA1.1 and BRCA1.3 probes showed good performance in ruling out BRCA1/2 germline mutations when methylation was detected (sensitivity 97.2% and 90.3%, respectively), although the specificity was poor because many Sporadic-C did not show methylation with these probes either (specificity for both 13.8%). The BRCA1.2 probe and the combination of the three BRCA1 probes (BRCA1 total) showed moderate sensitivity (both 63.9%) and specificity (72.5%) when used to rule in BRCA1/2 germline mutations when methylation was present.
Table 3.
BRCA promoter methylation percentages in BRCA1/2-related and sporadic breast carcinomas by methylation-specific multiplex ligation-dependent probe amplification
| Probe | BRCA1C-C, median % (range) | BRCA2-C, median % (range) | Sporadic-C, median % (range) | Test statistica | p Value |
|---|---|---|---|---|---|
| BRCA1.1 | 3 (0–11) | 3 (0–91) | 3 (0–80) | 2494.500 | 0.140 |
| BRCA1.2 | 21 (11–34) | 17 (11–100) | 15 (9–85) | 1795.500 | 0.00006* |
| BRCA1.3 | 11 (5–18) | 9 (5–100) | 7 (3–71) | 1760.600 | 0.00003* |
| BRCA2.1 | 24 (14–56) | 21 (8–100) | 15 (8–43) | 947.500 | 9.85°10−13* |
| BRCA2.2 | 11 (5–27) | 10 (5–100) | 5 (3–15) | 596.000 | 1.00°10−13* |
| BRCA2.3 | 17 (9–33) | 14 (8–100) | 9 (5–18) | 536.500 | 1.00°10−13* |
| BRCA2.4 | 9 (0–18) | 8 (0–100) | 5 (2–12) | 984.500 | 1.95°10−12* |
Abbreviations: BRCA1-C Breast carcinomas from BRCA1 germline mutation carriers, BRCA2-C Breast carcinomas from BRCA2 germline mutation carriers, Sporadic-C Sporadic breast carcinoma
aMann-Whitney U test for BRCA1-C and BRCA2-C together against Sporadic-C
*Statistically significant (two-sided p value <0.05)
Table 4.
Frequency of BRCA methylation (dichotomized results) in BRCA1/2-related and sporadic breast carcinomas by methylation-specific multiplex ligation-dependent probe amplification
| Probe | BRCA1-C, total n = 39 (%) | BRCA2-C, total n = 33 (%) | Sporadic-C, total n = 80 (%) | Test statistica | P-value | Total (%) | Cutoffb |
|---|---|---|---|---|---|---|---|
| BRCA1.1 | 0 (0) | 2 (6.1) | 11 (13.8) | 5.833c | 0.019* | 13 (8.6) | >15% |
| BRCA1.2 | 32 (82.1) | 14 (42.4) | 22 (27.5) | 20.296c | 0.000009* | 68 (44.7) | >17% |
| BRCA1.3 | 5 (12.8) | 2 (6.1) | 11 (13.8) | 0.589c | 0.465 | 18 (11.8) | >15% |
| BRCA1 totald | 32 (82.1) | 14 (42.4) | 22 (27.5) | 20.296c | 0.000009* | 68 (44.7) | |
| BRCA2.1 | 30 (76.9) | 18 (54.5) | 10 (12.5) | 47.117c | 2.93°10−12* | 58 (38.2) | >19% |
| BRCA2.2 | 4 (10.3) | 3 (9.1) | 0 (0) | 8.153e | 0.005* | 7 (4.6) | >15% |
| BRCA2.3 | 22 (56.4) | 14 (42.4) | 2 (2.5) | 45.600c | 1.65°10−12* | 38 (25) | >15% |
| BRCA2.4 | 3 (7.7) | 4 (12.1) | 0 (0.0) | 8.153e | 0.005* | 7 (4.6) | >15% |
| BRCA2 totald | 30 (76.9) | 20 (60.6) | 10 (12.5) | 51.432c | 0.029* | 60 (39.5) |
Abbreviations: BRCA1-C Breast carcinomas from BRCA1 germline mutation carriers, BRCA2-C Breast carcinomas from BRCA2 germline mutation carriers, Sporadic-C Sporadic breast carcinoma
aPearson’s chi-square test or Fisher’s exact test for BRCA1-C and BRCA2-C together against Sporadic-C
bCutoff based upon highest methylation percentage detected in normal breast tissue from nonmutation carriers
cPearson’s chi-square test
d BRCA1-C and BRCA2-C total entails the number (and percentage) of samples showing methylation in at least one of the BRCA1 or BRCA2 probes, respectively
eFisher’s exact test
*Statistically significant (two-sided p value <0.05)
Table 5.
Sensitivity and specificity for each methylation probe in distinguishing BRCA1/2-related from sporadic breast carcinomas
| Probe | Sensitivity | 95% CI | Specificity | 95% CI |
|---|---|---|---|---|
| BRCA1.1a | 70 of 72 (97.2%) | 90.3–99.6% | 11 of 80 (13.8%) | 7.1–23.3% |
| BRCA1.2b | 46 of 72 (63.9%) | 51.7–74.9% | 58 of 80 (72.5%) | 61.4–81.9% |
| BRCA1.3a | 65 of 72 (90.3%) | 90.0–96.0% | 11 of 80 (13.8%) | 7.1–23.3% |
| BRCA1 totalc | 46 of 72 (63.9%) | 51.7–74.9% | 58 of 80 (72.5%) | 61.4–81.9% |
| BRCA2.1b | 48 of 72 (66.7% | 54.6–77.3% | 70 of 80 (87.5%) | 78.2–93.8% |
| BRCA2.2b | 7 of 72 (9.7%) | 4.0–19.0% | 80 of 80 (100%) | 95.5–100% |
| BRCA2.3b | 36 of 72 (50.0%) | 38.0–62.0% | 78 of 80 (97.5%) | 91.3–99.7% |
| BRCA2.4b | 7 of 72 (9.7%) | 4.0–19.0% | 80 of 80 (100%) | 95.5–100% |
| BRCA2 totalc | 50 of 72 (69.4%) | 57.5–79.8% | 70 of 80 (87.5%) | 78.2–93.8% |
aSensitivity and specificity calculated as if BRCA1 promoter methylation would be performed to rule out BRCA germline mutations. True-positive: BRCA1/2-related cancers without BRCA1.1 or BRCA1.3 methylation. True-negative: sporadic cancers with BRCA1.1 or BRCA1.3 methylation
bSensitivity and specificity calculated as if BRCA1 promoter methylation would be performed to rule in BRCA germline mutations. True-positive: BRCA1/2-related cancers with BRCA1.2, BRCA2.1, BRCA2.2, BRCA2.3, or BRCA2.4 methylation. True-negative: sporadic cancers without BRCA1.2, BRCA2.1, BRCA2.2, BRCA2.3, or BRCA2.4 methylation
cSensitivity and specificity calculated as if BRCA1 promoter methylation would be performed to rule in BRCA germline mutations. True-positive: BRCA1/2-related cancers with methylation of at least one of the BRCA1 or BRCA2 probes. True-negative: sporadic cancers without methylation in any of the BRCA1 or BRCA2 probes
To evaluate the robustness of the MS-MLPA assay, we compared the results of the ME053 assay with another MS-MLPA assay tested on Sporadic-C in our laboratory, the ME001 assay (MRC-Holland) (C. B. Moelans, unpublished observations; data not shown). One of the BRCA1 methylation probes in this assay determines the methylation status of the same CpG site as the BRCA1.3 probe in the ME053 MS-MLPA assay. There was a strong correlation in dichotomized BRCA1 promoter methylation results in Sporadic-C between the two assays (Spearman’s rho correlation coefficient, 0.831; p = 1000°10−13). For absolute methylation percentages, the correlation was weaker but still significant (Spearman’s rho correlation coefficient, 0.379; p = 0.001). In general, the Sporadic-C showed slightly higher BRCA1 methylation percentages with the ME001 assay. In 4 of 80 cases, BRCA1 was methylated according to the ME001 assay but unmethylated according to the ME053 assay. However, methylation percentages in these cases were only slightly above the threshold of 15% (17–20%) with the ME001 assay.
BRCA2 promoter methylation in BRCA1/2-related and sporadic breast carcinomas
BRCA1/2-C showed significantly higher median methylation percentages for all BRCA2 methylation probes than Sporadic-C, although the absolute methylation percentages and their distribution varied considerably between the four BRCA2 methylation probes (Table 3, Fig. 2). Using dichotomized results, BRCA1/2-C showed significantly more frequent methylation in all four probes, as shown in Table 4. When the dichotomized results of the BRCA2 methylation probes were combined, 50 (69.4%) of 72 BRCA1/2-C showed methylation of at least one of the four BRCA2 methylation probes, compared with 10 (12.5%) of 80 Sporadic-C (p = 0.029). The sensitivity and specificity of the BRCA1 methylation probes in distinguishing BRCA1/2-C from Sporadic-C are shown in Table 5. The BRCA2.2 and BRCA2.4 probes showed excellent specificity (both 100%) when used to rule in BRCA1/2 germline mutations when methylation was detected because no Sporadic-C were methylated with these probes. However, the sensitivity was poor (both 9.7%) because few BRCA1/2-C showed methylation. The BRCA2.1 and BRCA2.3, as well as the combination of all four BRCA2 probes (BRCA2 total), showed moderate sensitivity (50.0–69.4%) and rather good specificity (87.5–97.5%) when used to rule in BRCA1/2 germline mutations when methylation was detected.
Correlation with clinicopathological variables
As shown in Table 1, BRCA1/2-C and Sporadic-C differed significantly with respect to age, grade, and ER and PR status. We analyzed whether the differences we observed in methylation frequencies between BRCA1/2-C and Sporadic-C may be related to these differences in clinicopathological variables (Tables 6, 7 and 8). In Sporadic-C, methylation of the BRCA1.1, BRCA1.2, and BRCA1.3 probes, separately as well as combined (BRCA1 total), was significantly more frequently detected in grade 3 tumors than in grades 1–2 tumors and in ER-negative than in ER-positive tumors. Methylation of the BRCA1.1 and BRCA1.3 probes was also significantly more frequently detected in PR-negative tumors. For BRCA2 methylation in Sporadic-C, there was a statistically significant association only with grade: Methylation of the BRCA2.1 probe and of all four BRCA2 probes combined was more frequently seen in grade 3 carcinomas. There were no statistically significant correlations between BRCA1 and BRCA2 methylation on the one hand and between tumor type (ductal versus lobular carcinomas) and HER2 status on the other hand. In BRCA1-C and BRCA2-C, there were no statistically significant associations between BRCA1 or BRCA2 methylation and clinicopathological variables. Moreover, no statistically significant correlation was found between CMI for BRCA1 and/or BRCA2 promoter methylation and age in BRCA1-C, BRCA2-C, Sporadic-C, BRCA1/2-N, and non-BRCA-related-N (Table 9).
Table 6.
Relationship between BRCA1/2 methylation and clinicopathological variables in sporadic breast carcinomas
| Sporadic carcinomas | Grades 1–2 vs. 3 | Ductal vs. lobular tumors | ER-positive vs. ER-negative | PR-positive vs. PR-negative | HER2-positive vs. HER2-negative | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Probe | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value |
| BRCA1.1 | 12.230b | 0.001* | 1.852b | 0.342 | 7.577b | 0.011* | 6.753b | 0.016* | 0.176b | 0.533 |
| BRCA1.2 | 8.190c | 0.006* | 0.026b | 1.000 | 6.689c | 0.014* | 3.762c | 0.071 | 2.825b | 0.125 |
| BRCA1.3 | 12.230b | 0.001* | 1.852b | 0.342 | 7.577b | 0.011* | 6.753b | 0.016* | 0.176b | 0.533 |
| BRCA1 totalc | 8.190c | 0.006* | 0.026b | 1.000 | 6.689c | 0.014* | 3.762c | 0.071 | 2.825b | 0.125 |
| BRCA2.1 | 6.577b | 0.015* | 1.659b | 0.345 | 0.706b | 0.462 | 0.274b | 0.736 | 3.688b | 0.115 |
| BRCA2.2 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| BRCA2.3 | 2.775b | 0.178 | 0.297b | 1.000 | 5.084b | 0.080 | 0.137b | 1.000 | 0.137b | 1.000 |
| BRCA2.4 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| BRCA2 totalc | 6.577b | 0.015* | 1.659c | 0.345 | 0.706c | 0.462 | 0.274c | 0.736 | 3.688c | 0.115 |
Abbreviations: ER Estrogen receptor, HER2 Human epidermal growth factor receptor 2, N/A Not applicable, because one of the two variables (either the methylation probe or the clinicopathological variable) was a constant, PR Progesterone receptor
aPearson’s chi-square test or Fisher’s exact test
bFisher’s exact test
cPearson’s chi-square test
*Statistically significant (two-sided p value <0.05)
Table 7.
Relationship between BRCA1/2 methylation and clinicopathological variables in BRCA1-related carcinomas
| BRCA1-related carcinomas | Grades 1–2 vs. 3 | Ductal vs. lobular tumors | ER-positive vs. ER-negative | PR-positive vs. PR-negative | HER2-positive vs. HER2-negative | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Probe | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value |
| BRCA1.1 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A |
| BRCA1.2 | 0.145b | 0.653 | 1.262b | 0.356 | 0.087b | 1.000 | 0.038b | 1.000 | 0.225b | 1.000 |
| BRCA1.3 | 0.031b | 1.000 | 0.342b | 1.000 | 0.115b | 1.000 | 0.096b | 1.000 | 0.151b | 1.000 |
| BRCA1 totalc | 0.145b | 0.653 | 1.262b | 0.356 | 0.087b | 1.000 | 0.038b | 1.000 | 0.225b | 1.000 |
| BRCA2.1 | 0.005b | 1.000 | 1.694b | 0.310 | 0.000b | 1.000 | 0.363b | 0.669 | 0.308b | 1.000 |
| BRCA2.2 | 0.009b | 1.000 | 0.265b | 1.000 | 3.482b | 0.099 | 0.001b | 1.000 | 0.117b | 1.000 |
| BRCA2.3 | 0.003b | 1.000 | 3.328b | 0.144 | 0.209bb | 0.740 | 0.070b | 1.000 | 0.793b | 1.000 |
| BRCA2.4 | 0.975b | 1.000 | 0.193b | 1.000 | 1.625b | 0.253 | 1.121b | 0.556 | 0.086b | 1.000 |
| BRCA2 totalc | 0.005b | 1.000 | 1.694b | 0.310 | 0.000b | 1.000 | 0.363b | 0.669 | 0.308b | 1.000 |
Abbreviations: BRCA1/2 BRCA1 and BRCA2, ER Estrogen receptor, HER2 Human epidermal growth factor receptor 2, N/A Not applicable, because one of the two variables (either the methylation probe or the clinicopathological variable) was a constant, PR Progesterone receptor
aPearson’s chi-square test or Fisher’s exact test
bFisher’s exact test
cPearson’s chi-square test
Table 8.
Relationship between BRCA1/2 methylation and clinicopathological variables in BRCA2-related carcinomas
| BRCA2-related carcinomas | Grades 1–2 vs. 3 | Ductal vs. lobular tumors | ER-positive vs. ER-negative | PR-positive vs. PR-negative | HER2-positive vs. HER2-negative | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Probe | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value | Test statistica | p Value |
| BRCA1.1 | 1.065b | 0.542 | 0.147b | 1.000 | 0.769b | 0.432 | 0.002b | 1.000 | 0.067b | 1.000 |
| BRCA1.2 | 0.062b | 1.000 | 0.057b | 1.000 | 0.248b | 0.695 | 0.730c | 0.491 | 0.760b | 1.000 |
| BRCA1.3 | 1.065b | 0.542 | 0.147b | 1.000 | 0.769b | 0.432 | 0.002b | 1.000 | 0.067b | 1.000 |
| BRCA1 totalc | 0.062b | 1.000 | 0.057b | 1.000 | 0.248b | 0.695 | 0.730bb | 0.491 | 0.760b | 1.000 |
| BRCA2.1 | 2.200c | 0.266 | 0.002b | 1.000 | 0.088b | 1.000 | 2.528c | 0.166 | 1.238b | 0.455 |
| BRCA2.2 | 1.650b | 0.534 | 0.229b | 1.000 | 0.149b | 1.000 | 0.437b | 0.601 | 0.103b | 1.000 |
| BRCA2.3 | 3.039b | 0.136 | 0.115b | 1.000 | 1.742b | 0.238 | 0.022c | 1.000 | 0.760b | 1.000 |
| BRCA2.4 | 0.569b | 0.586 | 0.317b | 1.000 | 0.001b | 1.000 | 0.004b | 1.000 | 0.142b | 1.000 |
| BRCA2 totalc | 3.110b | 0.132 | 0.057b | 1.000 | 0.916b | 0.431 | 0.863c | 0.481 | 1.587b | 0.394 |
Abbreviations: BRCA1/2 BRCA1 and BRCA2, ER Estrogen receptor, ER Estrogen receptor, HER2 Human epidermal growth factor receptor 2, PR Progesterone receptor
aPearson’s chi-square test or Fisher’s exact test
cPearson’s chi-square test
bFisher’s exact test
Table 9.
Correlation between age and cumulative methylation index for BRCA1/2 methylation
| Age | CMI for BRCA1 | CMI for BRCA2 | CMI for BRCA1 + BRCA2 | |||
|---|---|---|---|---|---|---|
| Spearman’s rho | p Value | Spearman’s rho | p Value | Spearman’s rho | p Value | |
| BRCA1-C | −0.297 | 0.066 | −0.287 | 0.077 | −0.275 | 0.090 |
| BRCA2-C | −0.019 | 0.918 | 0.003 | 0.989 | −0.035 | 0.846 |
| Sporadic-C | −0.153 | 0.175 | 0.003 | 0.982 | −0.120 | 0.289 |
| BRCA1/2-N | −0.252 | 0.513 | −0.467 | 0.205 | −0.417 | 0.265 |
| Non-BRCA-related-N | 0.300 | 0.624 | 0.100 | 0.873 | 0.100 | 0.873 |
Abbreviations: BRCA1/2 BRCA1 and BRCA2, BRCA1-C Breast carcinomas from BRCA1 germline mutation carriers, BRCA2-C Breast carcinomas from BRCA2 germline mutation carriers, BRCA1/2-N Normal breast tissue from BRCA1 and BRCA2 germline mutation carriers, CMI Cumulative methylation index, Non-BRCA-related-N Normal breast tissue from patients not known to have a BRCA1 or BRCA2 germline mutation, Sporadic-C Sporadic breast carcinoma
Correlation between age and CMI measured by Spearman’s rho correlation coefficient. CMI is calculated as the sum of the methylation percentage of all BRCA1 or BRCA2 methylation probes
Correlation between BRCA1/2 methylation and mRNA expression
Methylation of the evaluated CpG sites within the BRCA1 and BRCA2 promoters showed weak correlations with mRNA levels by TCGA data extraction through the Wanderer viewer. The Spearman’s correlation coefficients between BRCA1 methylation and mRNA expression were −0.203 for cg04110421 (targeted by the BRCA1.1 probe), −0.296 for cg16630982 (targeted by the BRCA1.2 probe), and −0.172 for cg08993267 (targeted by the BRCA1.3 probe). For BRCA2, the CpG loci identifiers from the TCGA data most closely located to our MS-MLPA targets were used. Therefore, the correlation between BRCA2 methylation and mRNA expression should be interpreted with caution. The Spearman’s correlation coefficients between BRCA2 methylation and mRNA expression were −0.014 for cg20073910 (82 and 69 bp from CpG sites targeted by the BRCA2.1 and BRCA2.4 probes, respectively), 0.067 for cg27253386 (80 and 69 bp from the CpG sites targeted by the BRCA2.3 probe), and −0.092 for cg08157964 (25 bp from the CpG site targeted by the BRCA2.2 probe).
BRCA promoter methylation in BRCA1/2-related and non-BRCA-related normal breast tissue
BRCA1/2-N samples showed statistically significant higher absolute methylation percentages for the BRCA2.3 and BRCA2.4 probes (p = 0.031 and p = 0.005, respectively) (Table 10, Fig. 2). There was a borderline significant trend of higher methylation percentages for the BRCA1.2, BRCA1.3, and BRCA2.1 probes in BRCA1/2-N samples than for the non-BRCA-related-N cases (Table 10, Fig. 2). If methylation cutoffs per probe were based upon the highest methylation percentage found in non-BRCA-related-N cases, 40% (two of five) and 60% (three of five) of BRCA1-N cases would have at least one methylated BRCA1 probe and one methylated BRCA2 probe, respectively. BRCA2-N cases would have methylation of at least one BRCA1 probe and one BRCA2 probe in 25% (one of four) and 50% (two of four) of cases, respectively (Table 11).
Table 10.
BRCA promoter methylation percentages in normal breast from BRCA1/2 germline mutation carriers and nonmutation carriers
| Probe | BRCA1/2-N, median % (range) | Non-BRCA-related-N, median % (range) | Test statistica | p Value |
|---|---|---|---|---|
| BRCA1.1 | 2 (0–3) | 3 (0–5) | 15.500 | 0.325 |
| BRCA1.2 | 17 (13–26) | 13 (13–17) | 8.500 | 0.057 |
| BRCA1.3 | 9 (6–16) | 7 (5–8) | 8.000 | 0.050 |
| BRCA2.1 | 19 (15–26) | 16 (14–19) | 8.500 | 0.060 |
| BRCA2.2 | 8 (6–16) | 6 (5–12) | 12.000 | 0.158 |
| BRCA2.3 | 13 (10–28) | 9 (8–14) | 6.500 | 0.031* |
| BRCA2.4 | 8 (6–9) | 5 (5–6) | 2.000 | 0.005* |
Abbreviations: BRCA1/2-N, Normal breast tissue from BRCA1 and BRCA2 germline mutation carriers; Non-BRCA-related-N, Normal breast tissue from patients not known to have a BRCA1 or BRCA2 germline mutation
aMann-Whitney U test for BRCA1/2-N together against non-BRCA-related-N
*Statistically significant (two-sided p value <0.05)
Table 11.
Frequency of BRCA methylation (dichotomized) in prophylactic mastectomies of BRCA1/2 germline mutation carriers by methylation-specific multiplex ligation-dependent probe amplification
| Probe | BRCA1, total n = 5 (%) | BRCA2, total n = 4 (%) | Total, n = 9 (%) | Cutoffa |
|---|---|---|---|---|
| BRCA1.1 | 0 (0) | 0 (0) | 0 (0) | >15% |
| BRCA1.2 | 2 (40) | 1 (25) | 3 (30) | >17% |
| BRCA1.3 | 1 (20) | 0 (0) | 1 (11.1) | >15% |
| BRCA1 totalb | 2 (40) | 1 (25) | 3 (30) | |
| BRCA2.1 | 2 (40) | 2 (50) | 4 (44.4) | >19% |
| BRCA2.2 | 1 (20) | 0 (0) | 1 (11.1) | >15% |
| BRCA2.3 | 2 (50) | 0 (0) | 2 (22.2) | >15% |
| BRCA2.4 | 0 (0) | 0 (0) | 0 (0) | >15% |
| BRCA2 totalb | 3 (60) | 2 (50) | 5 (55.5) |
aCutoff based upon highest methylation percentage detected in normal breast tissue from nonmutation carriers
b BRCA1 and BRCA2 total entails the number and percentage of samples showing methylation in at least one of the BRCA1 or BRCA2 probes, respectively
Discussion
The aim of this study was to investigate the diagnostic value of BRCA1/2 promoter methylation analysis using a new BRCA methylation MS-MLPA assay in distinguishing sporadic breast carcinomas from BRCA1 and BRCA2 germline mutation-related carcinomas in order to arrive at a clinically applicable prescreening test for BRCA1/2-related cancers. We observed considerably varying frequencies of BRCA promoter methylation between the targeted CpG sites across the BRCA1 and BRCA2 promoters. Some CpG sites were methylated more frequently in BRCA1/2-C than in Sporadic-C (those targeted by the BRCA1.2, BRCA2.1, BRCA2.2, BRCA2.3, and BRCA2.4 probes), whereas other CpG sites were methylated more frequently in Sporadic-C (those targeted by the BRCA1.1 and BRCA1.3 probes). In general, we observed frequent BRCA promoter methylation in BRCA1/2-C. At least 63.8% (46 of 72) of BRCA1/2-C and 12.5% (10 of 80) of Sporadic-C showed methylation of at least one of the targeted CpG sites in the BRCA1 or BRCA2 promoter. Interestingly, several BRCA1-C showed BRCA2 promoter methylation and vice versa. Sensitivity and specificity varied considerably between the probes. The best probes for ruling out Sporadic-C when methylation was detected were BRCA2.2 and BRCA2.4 (specificity 100%). However, many BRCA1/2-C would be missed because the sensitivity was poor (9.7%). The best probes for ruling out BRCA1/2-C when methylation was not detected were BRCA1.1 and BRCA1.3 (sensitivity 97.2% and 90.3%, respectively). However, many Sporadic-C would be misclassified as potentially BRCA1/2 germline mutation-related because the specificity was poor (both 13.8%). Sensitivity and specificity were most balanced when all four BRCA2 probes were used to rule in BRCA1/2 germline mutations when methylation was detected in at least one the BRCA2 probes (sensitivity 69.4%, specificity 87.5%). BRCA1 promoter methylation was more frequent in high-grade, ER-negative, and PR-negative tumors. This finding is in line with other reports in the literature because BRCA1 methylation has been more frequently described in triple-negative breast carcinomas [39, 40]. BRCA2 promoter methylation was more frequent in high-grade tumors but showed no other statistically significant clinicopathological associations.
In line with our findings, Daniels et al. [41] recently demonstrated that DNA methylation levels vary between CpG sites in the BRCA1 promoter. However, our findings do not support the general assumption and previous findings reported in the literature that BRCA promoter methylation and BRCA germline mutations are mutually exclusive. In most studies, none of the BRCA-related breast carcinomas showed BRCA promoter methylation [16, 21, 23–26]. Kontorovich et al. [20] observed BRCA1 promoter methylation in 3 (6.3%) of 48 BRCA1-related breast carcinomas, and Tapia et al. [17] observed BRCA1 promoter methylation in 2 (66.7%) of 3 observed BRCA1-related breast carcinomas. Differences in observed methylation frequencies could be related to the technique and specific CpG sites targeted, the quality of input material, and the determination of methylation cutoffs in subsequent analysis. It should be noted that some patients with a BRCA germline mutation may develop breast cancer through sporadic breast carcinogenetic mechanisms, which could affect methylation frequencies.
Whether BRCA promoter methylation may occur as a second hit in BRCA1/2-related breast carcinomas is still unclear. The main question is whether methylation really drives carcinogenesis or whether it can be considered a bystander. Interestingly, in our study, normal breast tissues from BRCA1/2 germline mutation carriers showed BRCA2 promoter methylation levels compared with normal breast tissues from patients without BRCA germline mutations, although the sample size was limited. Bijron et al. [42] described increased BRCA2 promoter methylation in normal and precursor fallopian tube tissues from BRCA germline mutation carriers compared with normal sporadic fallopian tube tissues. BRCA methylation might therefore play a role in carcinogenesis in a subset of BRCA germline mutation carriers.
To our knowledge, our present study is the largest one to date investigating both BRCA1 and BRCA2 promoter methylation in BRCA1 as well as BRCA2 germline mutation-related breast carcinomas. Moreover, this is the first MS-MLPA study to specifically test BRCA promoter methylation in BRCA1- and BRCA2-related breast carcinomas compared with sporadic breast carcinomas, as well as the first MS-MLPA study in which BRCA methylation levels have been investigated in normal breast tissues of BRCA carriers. We validated our results for one of the BRCA methylation probes by comparing them with data obtained from a previous MS-MLPA experiment using the commercially available ME001 MS-MLPA assay.
Our findings may have important implications for clinical practice, such as prescreening for BRCA germline genetic testing or eligibility for certain therapeutic strategies. BRCA1 promoter methylation analysis has been proposed as a cost-effective and reliable prescreening tool to exclude BRCA1 germline mutations in patients with breast cancer similar to MLH1 promoter methylation and Lynch syndrome [22, 43]. Moreover, recent research shows that breast and ovarian carcinomas with BRCA deficiencies, including BRCA methylation, may also benefit from PARP inhibitor therapy [30, 31, 44–49].
Although MS-MLPA has been shown to be a reliable tool to assess methylation in general, it targets single specific sites targetable by the HhaI methylation-sensitive restriction enzyme. For MS-MLPA to be a reliable prescreening tool for ruling in or ruling out BRCA germline mutations and/or determining sensitivity for targeted therapy, a review of existing literature and further research, preferably assessing all CpG sites in the BRCA promoter regions (e.g., by methylation-specific polymerase chain reaction), is needed to determine the most predictive CpG sites for each indication. The most predictive CpG sites should then be targetable by the HhaI methylation-sensitive restriction enzyme because otherwise MS-MLPA may not be the preferred methylation analytical technique in this context.
Conclusions
The diagnostic value of BRCA promoter methylation analysis in distinguishing BRCA1/2-related and sporadic breast carcinomas is considerably dependent on the targeted CpG sites. These findings are important for adequate use of BRCA methylation analysis as a prescreening tool for germline genetic testing or to identify patients who may benefit from targeted therapies such as PARP inhibitors, making their way to the clinic for breast cancer. Further research is needed to assess which other CpG sites are important in ruling in or ruling out BRCA germline mutations or determining sensitivity for targeted therapy.
Acknowledgements
We thank all collaborating pathology laboratories for making tumor tissues available.
Funding
This research was supported by no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Availability of data and materials
The datasets generated and/or analyzed during the current study are not publicly available, owing to privacy reasons for patients carrying BRCA1 or BRCA2 germline mutations, but they are available from the corresponding author on reasonable request.
Authors’ contributions
SV, CBM, and PJvD designed the study. SV and CBM carried out the experiments. SV analyzed the data and wrote the first manuscript draft. All authors read and approved the final manuscript.
Competing interests
MS-MLPA reagents were made available by MRC-Holland, Amsterdam, The Netherlands. The ME053 BRCA1-BRCA2 X1-0914 kit was designed by Lilit Atanesyan. MRC-Holland was not involved in the design and analysis of the experiments or in the writing of the submitted and published article.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Because coded archival pathological specimens were used, no ethical approval or specific prior informed consent was required according to Dutch legislation (Human Tissue and Medical Research Code of Conduct for Responsible Use 2011; https://www.federa.org/code-goed-gebruik-van-lichaamsmateriaal-2011 [in Dutch]).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- BRCA1/2
BRCA1 and BRCA2
- BRCA1-C
Breast carcinomas from BRCA1 germline mutation carriers
- BRCA2-C
Breast carcinomas from BRCA2 germline mutation carriers
- BRCA1/2-related
Related to a BRCA1 or BRCA2 germline mutation
- BRCA1/2-N
Normal breast tissue from BRCA1 and BRCA2 germline mutation carriers
- CMI
Cumulative methylation index
- CpG
Cytosine phosphate guanine
- ER
Estrogen receptor
- FFPE
Formalin-fixed paraffin-embedded
- Non-BRCA-related-N
Normal breast tissue from patients not known to have a BRCA1 or BRCA2 germline mutation
- HER2
Human epidermal growth factor receptor 2
- MS-MLPA
Methylation-specific multiplex ligation-dependent probe amplification
- PARP
Poly(adenosine diphosphate-ribose) polymerase
- PR
Progesterone receptor
- Sporadic-C
Sporadic breast carcinoma
- TCGA
The Cancer Genome Atlas
Contributor Information
Shoko Vos, Email: s.vos-4@umcutrecht.nl.
Cathy Beatrice Moelans, Email: c.b.moelans@umcutrecht.nl.
Paul Joannes van Diest, Email: p.j.vandiest@umcutrecht.nl.
References
- 1.Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, et al. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136:E359–86. doi: 10.1002/ijc.29210. [DOI] [PubMed] [Google Scholar]
- 2.Roy R, Chun J, Powell SN. BRCA1 and BRCA2: different roles in a common pathway of genome protection. Nat Rev Cancer. 2011;12:68–78. doi: 10.1038/nrc3181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Antoniou A, Pharoah PDP, Narod S, Risch HA, Eyfjord JE, Hopper JL, et al. Average risks of breast and ovarian cancer associated with BRCA1 or BRCA2 mutations detected in case series unselected for family history: a combined analysis of 22 studies. Am J Hum Genet. 2003;72:1117–30. doi: 10.1086/375033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Begg CB, Haile RW, Borg A, Malone KE, Concannon P, Thomas DC, et al. Variation of breast cancer risk among BRCA1/2 carriers. JAMA. 2008;299:194–201. doi: 10.1001/jama.2007.55-a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Paul A, Paul S. The breast cancer susceptibility genes (BRCA) in breast and ovarian cancers. Front Biosci. 2014;19:605–18. doi: 10.2741/4230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Venkitaraman AR. Cancer susceptibility and the functions of BRCA1 and BRCA2. Cell. 2002;108:171–82. doi: 10.1016/S0092-8674(02)00615-3. [DOI] [PubMed] [Google Scholar]
- 7.Chen S, Parmigiani G. Meta-analysis of BRCA1 and BRCA2 penetrance. J Clin Oncol. 2007;25:1329–33. doi: 10.1200/JCO.2006.09.1066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vos S, Van der Groep P, Van der Wall E, Van Diest PJ. Hereditary breast cancer syndromes: molecular pathogenesis and diagnostics. eLS. Chichester: John Wiley & Sons, Ltd; 2015. [Google Scholar]
- 9.Stefansson OA, Esteller M. Epigenetic modifications in breast cancer and their role in personalized medicine. Am J Pathol. 2013;183:1052–63. doi: 10.1016/j.ajpath.2013.04.033. [DOI] [PubMed] [Google Scholar]
- 10.Day TK, Bianco-Miotto T. Common gene pathways and families altered by DNA methylation in breast and prostate cancers. Endocr Relat Cancer. 2013;20:R215–32. doi: 10.1530/ERC-13-0204. [DOI] [PubMed] [Google Scholar]
- 11.Nowsheen S, Aziz K, Tran PT, Gorgoulis VG, Yang ES, Georgakilas AG. Epigenetic inactivation of DNA repair in breast cancer. Cancer Lett. 2014;342:213–22. doi: 10.1016/j.canlet.2012.05.015. [DOI] [PubMed] [Google Scholar]
- 12.Szyf M. DNA methylation signatures for breast cancer classification and prognosis. Genome Med. 2012;4:26. doi: 10.1186/gm325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jovanovic J, Rønneberg JA, Tost J, Kristensen V. The epigenetics of breast cancer. Mol Oncol. 2010;4:242–54. doi: 10.1016/j.molonc.2010.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Suijkerbuijk KPM, van Diest PJ, van der Wall E. Improving early breast cancer detection: focus on methylation. Ann Oncol. 2011;22:24–9. doi: 10.1093/annonc/mdq305. [DOI] [PubMed] [Google Scholar]
- 15.Widschwendter M, Jones PA. DNA methylation and breast carcinogenesis. Oncogene. 2002;21:5462–82. doi: 10.1038/sj.onc.1205606. [DOI] [PubMed] [Google Scholar]
- 16.Esteller M, Corn PG, Baylin SB, Herman JG. A gene hypermethylation profile of human cancer. Cancer Res. 2001;61:3225–9. [PubMed] [Google Scholar]
- 17.Tapia T, Smalley SV, Kohen P, Muñoz A, Solis LM, Corvalan A, et al. Promoter hypermethylation of BRCA1 correlates with absence of expression in hereditary breast cancer tumors. Epigenetics. 2008;3:157–63. doi: 10.4161/epi.3.3.6387. [DOI] [PubMed] [Google Scholar]
- 18.Dworkin AM, Spearman AD, Tseng SY, Sweet K, Toland AE. Methylation not a frequent “second hit” in tumors with germline BRCA mutations. Fam Cancer. 2009;8:339–46. doi: 10.1007/s10689-009-9240-1. [DOI] [PubMed] [Google Scholar]
- 19.Goodheart MJ, Rose SL, Hattermann-Zogg M, Smith BJ, De Young BR, Buller RE, et al. BRCA2 alteration is important in clear cell carcinoma of the ovary. Clin Genet. 2009;76:161–7. doi: 10.1111/j.1399-0004.2009.01207.x. [DOI] [PubMed] [Google Scholar]
- 20.Kontorovich T, Cohen Y, Nir U, Friedman E. Promoter methylation patterns of ATM, ATR, BRCA1, BRCA2 and P53 as putative cancer risk modifiers in Jewish BRCA1/BRCA2 mutation carriers. Breast Cancer Res Treat. 2009;116:195–200. doi: 10.1007/s10549-008-0121-3. [DOI] [PubMed] [Google Scholar]
- 21.Rennstam K, Ringberg A, Cunliffe HE, Olsson H, Landberg G, Hedenfalk I, et al. Genomic alterations in histopathologically normal breast tissue from BRCA1 mutation carriers may be caused by BRCA1 haploinsufficiency. Genes Chromosom Cancer. 2010;49:78–90. doi: 10.1002/gcc.20723. [DOI] [PubMed] [Google Scholar]
- 22.Lips EH, Mulder L, Oonk A, van der Kolk LE, Hogervorst FBL, Imholz ALT, et al. Triple-negative breast cancer: BRCAness and concordance of clinical features with BRCA1-mutation carriers. Br J Cancer. 2013;108:2172–7. doi: 10.1038/bjc.2013.144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Toffoli S, Bar I, Abdel-Sater F, Delrée P, Hilbert P, Cavallin F, et al. Identification by array comparative genomic hybridization of a new amplicon on chromosome 17q highly recurrent in BRCA1 mutated triple negative breast cancer. Breast Cancer Res. 2014;16:466. doi: 10.1186/s13058-014-0466-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Severson TM, Peeters J, Majewski I, Michaut M, Bosma A, Schouten PC, et al. BRCA1-like signature in triple negative breast cancer: molecular and clinical characterization reveals subgroups with therapeutic potential. Mol Oncol. 2015;9:1528–38. doi: 10.1016/j.molonc.2015.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tung N, Miron A, Schnitt SJ, Gautam S, Fetten K, Kaplan J, et al. Prevalence and predictors of loss of wild type BRCA1 in estrogen receptor positive and negative BRCA1-associated breast cancers. Breast Cancer Res. 2010;12:R95. doi: 10.1186/bcr2776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lisowska KM, Dudaladava V, Jarzab M, Huzarski T, Chmielik E, Stobiecka E, et al. BRCA1-related gene signature in breast cancer: the role of ER status and molecular type. Front Biosci. 2011;3:125–36. doi: 10.2741/e227. [DOI] [PubMed] [Google Scholar]
- 27.Lee JM, Ledermann JA, Kohn EC. PARP inhibitors for BRCA1/2 mutation-associated and BRCA-like malignancies. Ann Oncol. 2014;25:32–40. doi: 10.1093/annonc/mdt384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Livraghi L, Garber JE. PARP inhibitors in the management of breast cancer: current data and future prospects. BMC Med. 2015;13:188. doi: 10.1186/s12916-015-0425-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Dizdar O, Arslan C, Altundag K. Advances in PARP inhibitors for the treatment of breast cancer. Expert Opin Pharmacother. 2015;16:2751–8. doi: 10.1517/14656566.2015.1100168. [DOI] [PubMed] [Google Scholar]
- 30.Ledermann JA. PARP inhibitors in ovarian cancer. Ann Oncol. 2016;27(Suppl 1):i40–4. doi: 10.1093/annonc/mdw094. [DOI] [PubMed] [Google Scholar]
- 31.Moschetta M, George A, Kaye SB, Banerjee S. BRCA somatic mutations and epigenetic BRCA modifications in serous ovarian cancer. Ann Oncol. 2016;27:1449–55. doi: 10.1093/annonc/mdw142. [DOI] [PubMed] [Google Scholar]
- 32.Elston CW, Ellis IO. Pathological prognostic factors in breast cancer. I. The value of histological grade in breast cancer: experience from a large study with long-term follow-up. Histopathology. 1991;19:403–10. doi: 10.1111/j.1365-2559.1991.tb00229.x. [DOI] [PubMed] [Google Scholar]
- 33.Nygren AOH, Ameziane N, Duarte HMB, Vijzelaar RNCP, Waisfisz Q, Hess CJ, et al. Methylation-specific MLPA (MS-MLPA): simultaneous detection of CpG methylation and copy number changes of up to 40 sequences. Nucleic Acids Res. 2005;33:e128. doi: 10.1093/nar/gni127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Leong KJ, Wei W, Tannahill LA, Caldwell GM, Jones CE, Morton DG, et al. Methylation profiling of rectal cancer identifies novel markers of early-stage disease. Br J Surg. 2011;98:724–34. doi: 10.1002/bjs.7422. [DOI] [PubMed] [Google Scholar]
- 35.Paulsson K, An Q, Moorman AV, Parker H, Molloy G, Davies T, et al. Methylation of tumour suppressor gene promoters in the presence and absence of transcriptional silencing in high hyperdiploid acute lymphoblastic leukaemia. Br J Haematol. 2009;144:838–47. doi: 10.1111/j.1365-2141.2008.07523.x. [DOI] [PubMed] [Google Scholar]
- 36.Dikow N, Nygren AO, Schouten JP, Hartmann C, Krämer N, Janssen B, et al. Quantification of the methylation status of the PWS/AS imprinted region: comparison of two approaches based on bisulfite sequencing and methylation-sensitive MLPA. Mol Cell Probes. 2007;21:208–15. doi: 10.1016/j.mcp.2006.12.002. [DOI] [PubMed] [Google Scholar]
- 37.Suijkerbuijk KPM, Pan X, van der Wall E, van Diest PJ, Vooijs M. Comparison of different promoter methylation assays in breast cancer. Anal Cell Pathol (Amst) 2010;33:133–41. doi: 10.1155/2010/185219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Díez-Villanueva A, Mallona I, Peinado MA. Wanderer, an interactive viewer to explore DNA methylation and gene expression data in human cancer. Epigenetics Chromatin. 2015;8:22. doi: 10.1186/s13072-015-0014-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jacot W, Thezenas S, Senal R, Viglianti C, Laberenne AC, Lopez-Crapez E, et al. BRCA1 promoter hypermethylation, 53BP1 protein expression and PARP-1 activity as biomarkers of DNA repair deficit in breast cancer. BMC Cancer. 2013;13:523. doi: 10.1186/1471-2407-13-523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bal A, Verma S, Joshi K, Singla A, Thakur R, Arora S, et al. BRCA1-methylated sporadic breast cancers are BRCA-like in showing a basal phenotype and absence of ER expression. Virchows Arch. 2012;461:305–12. doi: 10.1007/s00428-012-1286-z. [DOI] [PubMed] [Google Scholar]
- 41.Daniels SL, Burghel GJ, Chambers P, Al-Baba S, Connley DD, Brock IW, et al. Levels of DNA methylation vary at CpG sites across the BRCA1 promoter, and differ according to triple negative and “BRCA-like” status, in both blood and tumour DNA. PLoS One. 2016;11:e0160174. doi: 10.1371/journal.pone.0160174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Bijron JG, van der Groep P, van Dorst EB, Seeber LMS, Sie-Go DMDS, Verheijen RHM, et al. Promoter hypermethylation patterns in fallopian tube epithelium of BRCA1 and BRCA2 germ line mutation carriers. Endocr Relat Cancer. 2012;19:69–81. doi: 10.1530/ERC-11-0338. [DOI] [PubMed] [Google Scholar]
- 43.Gausachs M, Mur P, Corral J, Pineda M, González S, Benito L, et al. MLH1 promoter hypermethylation in the analytical algorithm of Lynch syndrome: a cost-effectiveness study. Eur J Hum Genet. 2012;20:762–8. doi: 10.1038/ejhg.2011.277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Crafton SM, Bixel K, Hays JL. PARP inhibition and gynecologic malignancies: a review of current literature and on-going trials. Gynecol Oncol. 2016;142:588–96. doi: 10.1016/j.ygyno.2016.05.003. [DOI] [PubMed] [Google Scholar]
- 45.Alsop K, Fereday S, Meldrum C, deFazio A, Emmanuel C, George J, et al. BRCA mutation frequency and patterns of treatment response in BRCA mutation-positive women with ovarian cancer: a report from the Australian Ovarian Cancer Study Group. J Clin Oncol. 2012;30:2654–63. doi: 10.1200/JCO.2011.39.8545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Fong PC, Yap TA, Boss DS, Carden CP, Mergui-Roelvink M, Gourley C, et al. Poly(ADP)-ribose polymerase inhibition: frequent durable responses in BRCA carrier ovarian cancer correlating with platinum-free interval. J Clin Oncol. 2010;28:2512–9. doi: 10.1200/JCO.2009.26.9589. [DOI] [PubMed] [Google Scholar]
- 47.Banerjee S, Kaye SB, Ashworth A. Making the best of PARP inhibitors in ovarian cancer. Nat Rev Clin Oncol. 2010;7:508–19. doi: 10.1038/nrclinonc.2010.116. [DOI] [PubMed] [Google Scholar]
- 48.Konstantinopoulos PA, Spentzos D, Karlan BY, Taniguchi T, Fountzilas E, Francoeur N, et al. Gene expression profile of BRCAness that correlates with responsiveness to chemotherapy and with outcome in patients with epithelial ovarian cancer. J Clin Oncol. 2010;28:3555–61. doi: 10.1200/JCO.2009.27.5719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Veeck J, Ropero S, Setien F, Gonzalez-Suarez E, Osorio A, Benitez J, et al. BRCA1 CpG island hypermethylation predicts sensitivity to poly(adenosine diphosphate)-ribose polymerase inhibitors. J Clin Oncol. 2010;28:e563–6. doi: 10.1200/JCO.2010.30.1010. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and/or analyzed during the current study are not publicly available, owing to privacy reasons for patients carrying BRCA1 or BRCA2 germline mutations, but they are available from the corresponding author on reasonable request.