

Abstract
Hepatocellular carcinoma (HCC) is a predominantly male disease in which the androgen receptor (AR) serves an important pathogenic role in hepatocarcinogenesis. Fatty acid metabolism also contributes to hepatocarcinogenesis and is associated with the prognosis of cancer. The present study aimed to investigate the effects of the AR on fatty acid metabolism-associated gene expression in human hepatoma cell lines. AR-expression plasmids or control plasmids were transiently transfected into the human HCC cell lines Huh7 and HepG2. After 48 h, cellular protein and RNA were extracted and the expression of AR was confirmed by western blotting. Complementary DNA was synthesized and subjected to a quantitative polymerase chain reaction-based array to examine the expression of 84 fatty acid metabolism-associated genes. Overexpression of AR significantly downregulated the expression of 11 fatty acid metabolism-associated genes in Huh7 cells and 35 in HepG2 cells. The overexpression of AR also resulted in the upregulation of 6 fatty acid metabolism genes in HepG2 cells; however, it had no effect in Huh7 cells. Acyl-coenzyme A (CoA) thioesterase 7 and acyl-CoA oxidase 3 were downregulated in both cell lines. In conclusion, upregulation of AR via overexpression led to the disturbance of fatty acid metabolism-associated gene expression in human HCC cells. Therefore, the AR may serve a role in hepatocarcinogenesis via the regulation of hepatocellular fatty acid metabolism.
Keywords: androgen receptor, fatty acid, liver cancer
Introduction
Steroid sex hormones serve major roles as tumor promoters by promoting cell cycle progression in several types of cancer, including hepatocellular carcinoma (HCC) (1–3). The sex hormone androgen and its associated androgen receptor (AR) are involved in the development of the lipogenic phenotype of cancer cells via the induction of proteins and enzymes, including fatty acid synthase (FASN), required for their autonomous nutrition (4).
A number of studies have implicated AR, a member of the nuclear steroid receptor superfamily, in hepatocarcinogenesis (1,3,5–7). AR is a transcription factor activated via ligand-dependent and ligand-independent mechanisms (8). Cytochrome P450, transforming growth factor β1, vascular endothelial growth factor A and heat shock protein family A member 5 are target genes of ARs in the liver (3,7,9,10). Crosstalk between ARs and liver X receptor (nuclear receptor subfamily 1 group H member 2) has also been implicated in maintaining cholesterol homeostasis (11). It has also been demonstrated that the activity of ARs is coupled with the upregulation of FASN (4).
Abnormalities in cellular metabolism are closely associated with HCC occurrence and development (4,12) and it has been demonstrated that FASN is highly expressed in a number of cancer types, including HCC (12). In HCC patients, abnormalities in plasma phospholipid fatty acid profiles are occasionally observed, which may be attributed to the alteration of intrinsic fatty acid metabolism caused by the cancer itself (13). Dietary fatty acids may be implicated in the risk of developing non-viral hepatitis-associated HCC (14); however, there is data to refute this hypothesis (13).
Fatty acid metabolism also serves a role in human carcinogenesis and is associated with a poor prognosis in HCC patients. The present study aimed to investigate the effects of AR expression on fatty acid metabolism-associated gene regulation in human HCC cell lines.
Materials and methods
Cell culture
The human HCC cell lines HepG2 and Huh7, as described previously (3), were purchased from the Japanese Collection of Research Bioresources Cell Bank (Ibaraki, Japan). The cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and grown at 37°C with 5% CO2.
Transfection and RNA extraction
A total of 4×105 cells per well were placed in 6-well plates 24 h prior to transfection. The cells were transfected with 0.4 µg human AR protein-expressing plasmid (pSG5-AR) or 0.4 µg control vectors, which were kindly provided by Professor J.T. Isaacs of The Johns Hopkins University School of Medicine (Baltimore, MD, USA) (15), using Effectene Transfection Reagent (Qiagen GmbH, Hilden, Germany) according to the manufacturer's protocol. At 48 h post-transfection, cellular RNA was collected using RNeasy (Qiagen GmbH), and RNA was stored at −80°C until use. RNA quality was measured by a NanoDrop spectrophotometer (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Complementary DNA (cDNA) synthesis and quantiative polymerase chain reaction (qPCR) arrays
cDNA synthesis was performed by an RT2 First Strand kit (Qiagen GmbH) using 1 µg of RNA per reaction. The cDNA synthesis reaction was incubated at 42°C for 15 min and then halted by heating at 95°C for 5 min. A human fatty acid metabolism PCR array (no. PAHS-007Z; Qiagen GmbH) was carried out according to the manufacturer's protocol. Briefly, amplification of cDNA was monitored with SYBR-Green by quantitative PCR analysis. PCR was performed in 25 µl of ROX PCR Master mix (Qiagen GmbH) containing primers and 1 µl of the reverse-transcription reaction mixture, using a 7300 Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.) according to the manufacturer's protocol. The PCR reaction was performed as follows: 95°C for 1 min, followed by 40 cycles of 95°C for 5 sec and 60°C for 30 sec. The data analysis was performed using the comparative threshold cycle method (16) and analyzed using PCR Array Data Analysis Software v2.1 (http://www.sabiosciences.com/pcrarraydataanalysis.php).
Western blot analysis
Cell lysates were prepared 48 h post transfection in 1X SDS sample buffer (Wako Pure Chemical Industries, Ltd., Osaka, Japan). Protein concentration was determined using the Bradford method. Cell lysate proteins (5 µg/well) were separated by 5–20% SDS-PAGE and transferred onto a nitrocellulose membrane (ATTO Corp., Tokyo, Japan). AR expression was analyzed by western blotting by incubating membranes with rabbit polyclonal antibodies against AR (no. sc-816; dilution, 1:1,000) or GAPDH (no. sc-25778; dilution, 1:2,000) (both from Santa Cruz Biotechnology Inc., Dallas, TX, USA) at 4°C for 16 h. Membranes were then incubated with anti-rabbit IgG HRP-linked secondary antibody (no. 7074; dilution, 1:3,500; Cell Signaling Technology, Inc., Danvers, MA, USA) at room temperature for 1 h. Proteins were visualized using an enhanced chemiluminescence western blotting substrate (GE Healthcare Life Sciences, Chalfont, UK) and scanned with an image analyzer (LAS-4000) and Image Gauge software (version 3.1) (both from Fujifilm, Tokyo, Japan). The relative quantity of protein was determined by normalizing to GAPDH.
Statistical analysis
For statistical analysis, two-tailed Student's t-tests were performed with DA Stats software version PAF01644 (NIFTY Corp.; Fujitsu). P<0.05 was considered to indicate a statistically significant difference.
Results
Overexpression of AR in human HCC cells
Overexpression of AR was induced in HepG2 and Huh7 cells to validate hypotheses of the effect of AR on the regulation of fatty acid metabolism-associated gene expression. In HepG2 cells transiently transfected with pSG5-AR, overexpression of AR was confirmed by western blot analysis (Fig. 1), and AR overexpression in Huh7 cells transiently transfected with pSG5-AR was demonstrated by western blotting in a previous study (3).
Figure 1.
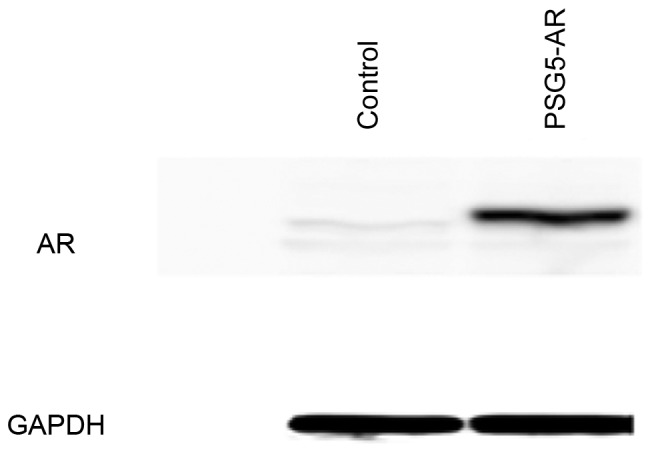
Overexpression of AR in HepG2 cells. Western blot analysis of AR and GAPDH expression level in HepG2 cells transiently transfected with the AR-expressing plasmid, pSG5-AR, or a control vector. AR, androgen receptor.
Downregulated fatty acid metabolism-associated genes in AR-overexpressing HepG2 cells
Fatty acid metabolism-associated gene expression profiles were examined using a qPCR-based array. Expression levels between HepG2 cells transiently overexpressing AR and control vector-transfected HepG2 cells were compared. In HepG2 cells, AR overexpression significantly downregulated the expression of 35 fatty acid metabolism-associated genes, including 6 acyl-coenzyme A (CoA) dehydrogenases [acyl-CoA dehydrogenase family member (ACAD)10, acyl-CoA dehydrogenase, very long chain, acyl-CoA dehydrogenase, C-2 to C-3 short chain (ACADS), ACAD9, ACADS branched (B) and enoyl-CoA hydratase and 3-hydroxyacyl CoA dehydrogenase (EHHADH)], 2 acyl-CoA oxidases [acyl-CoA oxidase (ACOX) 1 and ACOX3], 4 acyl-CoA synthetases [acyl-CoA synthetase long-chain family member (ACSL)1, acyl-CoA synthetase bubblegum family member (ACSBG)2, ACSBG1 and ACSL4], 4 acyl-CoA thioesterases [acyl-CoA thioesterase (ACOT)8, ACOT1, ACOT2 and ACOT7], 4 carnitine transferases [carnitine palmitoyltransferase (CPT)2, carnitine O-octanoyltranserase, carnitine O-acetyltransferase, CPT1B], 5 other fatty acid metabolism-associated genes [aldehyde dehydrogenase 2 family (mitochondrial) (ALDH2), 2,4-dienoyl CoA reductase 2, peroxisomal, enoyl CoA hydratase, short chain, 1, mitochondrial, enoyl-CoA δ isomerase 2 and methylmalonyl CoA mutase], 2 genes of fatty acid transport [CPT1A and solute carrier family 27 (fatty acid transporter), member 4], 4 genes of fatty acid biosynthesis regulation [protein kinase, cAMP-dependent, catalytic, α (PRKACA), protein kinase, AMP-activated, α 2 catalytic subunit, protein kinase, AMP-activated, γ 1 non-catalytic subunit and protein kinase, AMP-activated, α 1 catalytic subunit], 1 gene of ketogenesis and ketone body metabolism (3-hydroxybutyrate dehydrogenase, type 1) and 3 genes of triacylglycerol metabolism [glycerol-3-phosphatase dehydrogenase 1 (soluble), lipase, hormone-sensitive and glycerol kinase; Table I].
Table I.
Fatty acid metabolism-associated genes are significantly downregulated in androgen receptor-overexpressing HepG2 cells examined by polymerase chain reaction arrays.
| Symbol | Description | Fold change (vs. control) | P-value |
|---|---|---|---|
| GPD1 | Glycerol-3-phosphatase dehydrogenase 1 (soluble) | 0.40 | 0.0259 |
| ACAD10 | Acyl-CoA dehydrogenase family, member 10 | 0.44 | 0.0135 |
| LIPE | Lipase, hormone-sensitive | 0.50 | 0.0099 |
| ACADVL | Acyl-CoA dehydrogenase, very long chain | 0.54 | 0.0018 |
| CPT2 | Carnitine palmitoyltransferase 2 | 0.54 | 0.0181 |
| CROT | Carnitine O-octanoyltranserase | 0.54 | 0.0069 |
| CPT1A | Carnitine palmitoyltransferase 1A (liver) | 0.57 | 0.0039 |
| ACSL1 | Acyl-CoA synthetase long-chain family member 1 | 0.57 | 0.0375 |
| ACSBG2 | Acyl-CoA synthetase bubblegum family member 2 | 0.58 | 0.0096 |
| ACADS | Acyl-CoA dehydrogenase, C-2 to C-3 short chain | 0.58 | 0.0448 |
| ACAD9 | Acyl-CoA dehydrogenase family, member 9 | 0.59 | 0.0177 |
| ACADSB | Acyl-CoA dehydrogenase, short/branched chain | 0.60 | 0.0475 |
| ACOX1 | Acyl-CoA oxidase 1, palmitoyl | 0.60 | 0.0020 |
| ACOX3 | Acyl-CoA oxidase 3, pristanoyl | 0.60 | 0.0199 |
| PRKACA | Protein kinase, cAMP-dependent, catalytic, α | 0.61 | 0.0012 |
| PRKAA2 | Protein kinase, AMP-activated, α 2 catalytic subunit | 0.61 | 0.0347 |
| SLC27A4 | Solute carrier family 27 (fatty acid transporter), member 4 | 0.61 | 0.0428 |
| BDH1 | 3-hydroxybutyrate dehydrogenase, type 1 | 0.61 | 0.0130 |
| ACSBG1 | Acyl-CoA synthetase bubblegum family member 1 | 0.61 | 0.0233 |
| EHHADH | Enoyl-CoA, hydratase/3-hydroxyacyl CoA dehydrogenase | 0.62 | 0.0212 |
| ACSL4 | Acyl-CoA synthetase long-chain family member 4 | 0.63 | 0.0341 |
| ACOT8 | Acyl-CoA thioesterase 8 | 0.63 | 0.0191 |
| ACOT1 | Acyl-CoA thioesterase 1 | 0.64 | 0.0017 |
| ALDH2 | Aldehyde dehydrogenase 2 family (mitochondrial) | 0.64 | 0.0088 |
| PRKAG1 | Protein kinase, AMP-activated, γ 1 non-catalytic subunit | 0.64 | 0.0321 |
| CRAT | Carnitine O-acetyltransferase | 0.66 | 0.0002 |
| DECR2 | 2,4-dienoyl CoA reductase 2, peroxisomal | 0.66 | 0.0412 |
| ECHS1 | Enoyl CoA hydratase, short chain, 1, mitochondrial | 0.69 | 0.0368 |
| ECI2 | Enoyl-CoA δ isomerase 2 | 0.70 | 0.0347 |
| GK | Glycerol kinase | 0.72 | 0.0049 |
| ACOT2 | Acyl-CoA thioesterase 2 | 0.73 | 0.0252 |
| CPT1B | Carnitine palmitoyltransferase 1B (muscle) | 0.73 | 0.0038 |
| PRKAA1 | Protein kinase, AMP-activated, α 1 catalytic subunit | 0.76 | 0.0409 |
| MUT | Methylmalonyl CoA mutase | 0.81 | 0.0453 |
| ACOT7 | Acyl-CoA thioesterase 7 | 0.81 | 0.0098 |
CoA, conenzyme A.
Upregulated fatty acid metabolism-associated genes in AR-overexpressing HepG2 cells
In HepG2 cells, AR significantly upregulated the expression of 6 fatty acid metabolism-associated genes (Table II), including 3 fatty acid transports [Solute carrier family (SLC) 27A5, SLC27A2 and fatty acid binding protein 4, adipocyte], 1 fatty acid biosynthesis regulator [protein kinase, AMP-activated, γ 2 non-catalytic subunit (PRKAG2)], 1 acyl-CoA thioesterase (ACOT9) and 1 other fatty acid metabolism-associated gene [pyrophosphatase (inorganic) 1 (PPA1)].
Table II.
Fatty acid metabolism-associated genes are significantly upregulated in androgen receptor-overexpressing HepG2 cells examined by polymerase chain reaction arrays.
| Symbol | Description | Fold change (vs. control) | P-value |
|---|---|---|---|
| FABP4 | Fatty acid binding protein 4, adipocyte | 2.25 | 0.0015 |
| SLC27A2 | Solute carrier family 27 (fatty acid transporter), member 2 | 1.51 | 0.0167 |
| SLC27A5 | Solute carrier family 27 (fatty acid transporter), member 5 | 1.40 | 0.0156 |
| ACOT9 | Acyl-CoA thioesterase 9 | 1.25 | 0.0216 |
| PPA1 | Pyrophosphatase (inorganic) 1 | 1.23 | 0.0162 |
| PRKAG2 | Protein kinase, AMP-activated, γ 2 non-catalytic subunit | 1.20 | 0.0331 |
CoA, conenzyme A.
Upregulated and downregulated fatty acid metabolism-associated genes in AR-overexpressing Huh7 cells
In Huh7 cells, AR overexpression significantly downregulated the expression of the following 11 fatty acid metabolism-associated genes: 4 acyl-CoA synthetases [acyl-CoA synthetase medium-chain family member (ACSM) 2A, ACSL6, ACSL3 and ACSM3], 3 acyl-CoA thioesterases (ACOT7, ACOT9 and ACOT6), 1 acyl-CoA oxidase (ACOX3), 1 fatty acid transporter (SLC27A2), 1 fatty acid biosynthesis regulator (PRKAG2) and 1 regulator of ketogenesis and ketone body metabolism (3-hydroxybutyrate dehydrogenase, type 2; Table III). However, AR overexpression did not significantly upregulate the expression of any fatty acid metabolism-associated genes in Huh7 cells.
Table III.
Fatty acid metabolism-associated genes are significantly downregulated in androgen receptor-overexpressing Huh7 cells.
| Symbol | Description | Fold change (vs. control) | P-value |
|---|---|---|---|
| ACOX3 | Acyl-CoA oxidase 3, pristanoyl | 0.62 | 0.0180 |
| PRKAG2 | Protein kinase, AMP-activated, γ 2 non-catalytic subunit | 0.65 | 0.0402 |
| ACSM3 | Acyl-CoA synthetase medium-chain family member 3 | 0.66 | 0.0249 |
| SLC27A2 | Solute carrier family 27 (fatty acid transporter), member 2 | 0.69 | 0.0387 |
| BDH2 | 3-hydroxybutyrate dehydrogenase, type 2 | 0.69 | 0.0015 |
| ACSL3 | Acyl-CoA synthetase long-chain family member 3 | 0.87 | 0.0127 |
| ACOT6 | Acyl-CoA thioesterase 6 | 0.88 | 0.0020 |
| ACSL6 | Acyl-CoA synthetase long-chain family member 6 | 0.89 | 0.0052 |
| ACOT9 | Acyl-CoA thioesterase 9 | 0.91 | 0.0434 |
| ACOT7 | Acyl-CoA thioesterase 7 | 0.92 | 0.0480 |
| ACSM2A | Acyl-CoA synthetase medium-chain family member 2A | 0.93 | 0.0018 |
CoA, conenzyme A.
Discussion
The present study demonstrated that overexpression of AR alters fatty acid metabolism-associated gene expression in human HCC cell lines. Previous studies (17,18) demonstrated that in patients with HCC, ACADSB and EHHADH expression are downregulated in tissue isolated from cirrhotic livers compared with control tissues from healthy patients. Acyl-CoA oxidase deficient-mice spontaneously develop HCC (19). Li et al (20) reported that cluster of differentiation (CD) 147 expression significantly contributed to the reprogramming of fatty acid metabolism in HCC cells, and that CD147 downregulated CPT1A and ACOX1 by activating the p38 mitogen-activated protein kinase signaling pathway to inhibit fatty acid β-oxidation. A previous study in phosphatase and tensin homolog null nonalcoholic steatohepatitis liver tissue isolated from mice revealed that ACSL1 expression is decreased, and is almost non-existent in tumors isolated from these livers (21). A recent study (22) demonstrated that the presence of a specific mutation in ALDH2 (E487 K) led to increased protein turnover and promoted murine hepatocarcinogenesis. The presence of the DnaJ heat shock protein family (Hsp40) member B1-PRKACA chimeric transcript in fibrolamellar HCC suggests that this genetic alteration contributes to tumor pathogenesis (23). Therefore, the present study supports previous reports (17–23) (Table I).
Sakabe et al (24) reported that PRKAG2 enhances the effect of interferon (IFN)-α/fluorouracil (5-FU) and serves as a prognostic marker for IFN-α/5-FU therapy in HCC. The present study demonstrated that AR significantly upregulated the expression of PRKAG2 (Table II). ACOT7 has been implicated in the regulation of neuronal fatty acid metabolism to prevent neurotoxicity (25), and ACOX3 is expressed at particularly low levels in the liver (26). Notably, ACOT7 and ACOX3 were downregulated by the overexpression of AR in HepG2 and Huh7 cells (Tables I and III). These results support the results of a previous study by Bolton et al (27) that implicated AR in the regulation of cellular lipid metabolism.
In general, cancer cells synthesize a high amount of fatty acids via increased expression, and subsequent activity, of lipogenic enzymes to fulfill their high energy requirements and to maintain cell membranes for growth (4,28,29). In patients with HCC, the alterations in lipid and lipoprotein metabolism can be clearly observed (30).
AR has been demonstrated to be associated with hepato-carcinogenesis and HCC development (1,3,5–7,9,10). The present study revealed that AR could serve a role in hepatocarcinogenesis via the regulation of hepatocellular fatty acid metabolism. Hepatitis C virus (HCV) augments AR-mediated signaling (7) and replicates in hepatocytes using lipid droplets, which are used for the storage of neutral lipids (31). In a previous study, eradication of HCV using an interferon-free antiviral regimen resulted in rapid changes in the metabolic signaling pathway in the liver, which suggested that there was a direct effect of HCV replication on lipid homeostasis (32); however, further studies are required to determine potential indirect effects of HCV on lipid homeostasis. Hepatitis B virus (HBV) has also been revealed to augment AR-mediated signaling (5,6) and to induce hepatic steatosis by influencing fatty acid metabolism (33). HBV serves a more minor role in hepatic steatosis compared with HCV (34).
In conclusion, improving the understanding of the association between AR and fatty acid metabolism in hepatocarcinogenesis and the development of HCC may aid in the creation of novel therapeutic and prevention strategies.
Acknowledgements
The authors would like to thank Professor J.T. Isaacs (The Johns Hopkins University School of Medicine, Baltimore, MD, USA) for providing the plasmid pSG5-AR. The present work was supported by grants from the Ministry of Health, Labour and Welfare of Japan. This study was partially supported by the Research Program on Hepatitis from the Japan Agency for Medical Research and Development (AMED).
References
- 1.Nagasue N, Chang YC, Hayashi T, Galizia G, Kohno H, Nakamura T, Yukaya H. Androgen receptor in hepatocellular carcinoma as a prognostic factor after hepatic resection. Ann Surg. 1989;209:424–427. doi: 10.1097/00000658-198904000-00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Okitsu K, Kanda T, Imazeki F, Yonemitsu Y, Ray RB, Chang C, Yokosuka O. Involvement of interleukin-6 and androgen receptor signaling in pancreatic cancer. Genes Cancer. 2010;1:859–867. doi: 10.1177/1947601910383417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Jiang X, Kanda T, Nakamoto S, Miyamura T, Wu S, Yokosuka O. Involvement of androgen receptor and glucose-regulated protein 78 kDa in human hepatocarcinogenesis. Exp Cell Res. 2014;323:326–336. doi: 10.1016/j.yexcr.2014.02.017. [DOI] [PubMed] [Google Scholar]
- 4.Rochefort H, Chalbos D. The role of sex steroid receptors on lipogenesis in breast and prostate carcinogenesis: A viewpoint. Horm Cancer. 2010;1:63–70. doi: 10.1007/s12672-010-0009-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chiu CM, Yeh SH, Chen PJ, Kuo TJ, Chang CJ, Chen PJ, Yang WJ, Chen DS. Hepatitis B virus X protein enhances androgen receptor-responsive gene expression depending on androgen level. Proc Natl Acad Sci USA. 2007;104:2571–2578. doi: 10.1073/pnas.0609498104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zheng Y, Chen WL, Ma WL, Chang C, Ou JH. Enhancement of gene transactivation activity of androgen receptor by hepatitis B virus X protein. Virology. 2007;363:454–461. doi: 10.1016/j.virol.2007.01.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kanda T, Steele R, Ray R, Ray RB. Hepatitis C virus core protein augments androgen receptor-mediated signaling. J Virol. 2008;82:11066–11072. doi: 10.1128/JVI.01300-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ueda T, Bruchovsky N, Sadar MD. Activation of the androgen receptor N-terminal domain by interleukin-6 via MAPK and STAT3 signal transduction pathways. J Biol Chem. 2002;277:7076–7085. doi: 10.1074/jbc.M108255200. [DOI] [PubMed] [Google Scholar]
- 9.Gehlhaus M, Schmitt N, Volk B, Meyer RP. Antiepileptic drugs affect neuronal androgen signaling via a cytochrome P450-dependent pathway. J Pharmacol Exp Ther. 2007;322:550–559. doi: 10.1124/jpet.107.120303. [DOI] [PubMed] [Google Scholar]
- 10.Yoon G, Kim JY, Choi YK, Won YS, Lim IK. Direct activation of TGF-beta1 transcription by androgen and androgen receptor complex in Huh7 human hepatoma cells and its tumor in nude mice. J Cell Biochem. 2006;97:393–411. doi: 10.1002/jcb.20638. [DOI] [PubMed] [Google Scholar]
- 11.Krycer JR, Brown AJ. Cross-talk between the androgen receptor and the liver X receptor: Implications for cholesterol homeostasis. J Biol Chem. 2011;286:20637–20647. doi: 10.1074/jbc.M111.227082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hao Q, Li T, Zhang X, Gao P, Qiao P, Li S, Geng Z. Expression and roles of fatty acid synthase in hepatocellular carcinoma. Oncol Rep. 2014;32:2471–2476. doi: 10.3892/or.2014.3484. [DOI] [PubMed] [Google Scholar]
- 13.Qiu JF, Zhang KL, Zhang XJ, Hu YJ, Li P, Shang CZ, Wan JB. Abnormalities in plasma phospholipid fatty acid profiles of patients with hepatocellular carcinoma. Lipids. 2015;50:977–985. doi: 10.1007/s11745-015-4060-6. [DOI] [PubMed] [Google Scholar]
- 14.Koh WP, Dan YY, Goh GB, Jin A, Wang R, Yuan JM. Dietary fatty acids and risk of hepatocellular carcinoma in the Singapore Chinese health study. Liver Int. 2016;36:893–901. doi: 10.1111/liv.12978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Litvinov IV, Chang C, Isaacs JT. Molecular characterization of the commonly used human androgen receptor expression vector, pSG5-AR. Prostate. 2004;58:319–324. doi: 10.1002/pros.20027. [DOI] [PubMed] [Google Scholar]
- 16.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 17.Schröder PC, Segura V, Riezu JI, Sangro B, Mato JM, Prieto J, Santamaría E, Corrales FJ. A signature of six genes highlights defects on cell growth and specific metabolic pathways in murine and human hepatocellular carcinoma. Funct Integr Genomics. 2011;11:419–429. doi: 10.1007/s10142-011-0230-7. [DOI] [PubMed] [Google Scholar]
- 18.Suto K, Kajihara-Kano H, Yokoyama Y, Hayakari M, Kimura J, Kumano T, Takahata T, Kudo H, Tsuchida S. Decreased expression of the peroxisomal bifunctional enzyme and carbonyl reductase in human hepatocellular carcinomas. J Cancer Res Clin Oncol. 1999;125:83–88. doi: 10.1007/s004320050246. [DOI] [PubMed] [Google Scholar]
- 19.Meyer K, Lee JS, Dyck PA, Cao WQ, Rao MS, Thorgeirsson SS, Reddy JK. Molecular profiling of hepatocellular carcinomas developing spontaneously in acyl-CoA oxidase deficient mice: Comparison with liver tumors induced in wild-type mice by a peroxisome proliferator and a genotoxic carcinogen. Carcinogenesis. 2003;24:975–984. doi: 10.1093/carcin/bgg040. [DOI] [PubMed] [Google Scholar]
- 20.Li J, Huang Q, Long X, Zhang J, Huang X, Aa J, Yang H, Chen Z, Xing J. CD147 reprograms fatty acid metabolism in hepatocellular carcinoma cells through Akt/mTOR/SREBP1c and P38/PPARα pathways. J Hepatol. 2015;63:1378–1389. doi: 10.1016/j.jhep.2015.07.039. [DOI] [PubMed] [Google Scholar]
- 21.Muir K, Hazim A, He Y, Peyressatre M, Kim DY, Song X, Beretta L. Proteomic and lipidomic signatures of lipid metabolism in NASH-associated hepatocellular carcinoma. Cancer Res. 2013;73:4722–4731. doi: 10.1158/0008-5472.CAN-12-3797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jin S, Chen J, Chen L, Histen G, Lin Z, Gross S, Hixon J, Chen Y, Kung C, Chen Y, et al. ALDH2(E487K) mutation increases protein turnover and promotes murine hepatocarcinogenesis. Proc Natl Acad Sci USA. 2015;112:9088–9093. doi: 10.1073/pnas.1510757112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Honeyman JN, Simon EP, Robine N, Chiaroni-Clarke R, Darcy DG, Lim II, Gleason CE, Murphy JM, Rosenberg BR, Teegan L, et al. Detection of a recurrent DNAJB1-PRKACA chimeric transcript in fibrolamellar hepatocellular carcinoma. Science. 2014;343:1010–1014. doi: 10.1126/science.1249484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Sakabe T, Tsuchiya H, Kanki K, Azumi J, Gonda K, Mizuta Y, Yamada D, Wada H, Shomori K, Nagano H, Shiota G. Identification of the genes chemosensitizing hepatocellular carcinoma cells to interferon-α/5-fluorouracil and their clinical significance. PLoS One. 2013;8:e56197. doi: 10.1371/journal.pone.0056197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ellis JM, Wong GW, Wolfgang MJ. Acyl coenzyme A thioesterase 7 regulates neuronal fatty acid metabolism to prevent neurotoxicity. Mol Cell Biol. 2013;33:1869–1882. doi: 10.1128/MCB.01548-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zha S, Ferdinandusse S, Hicks JL, Denis S, Dunn TA, Wanders RJ, Luo J, De Marzo AM, Isaacs WB. Peroxisomal branched chain fatty acid beta-oxidation pathway is upregulated in prostate cancer. Prostate. 2005;63:316–323. doi: 10.1002/pros.20177. [DOI] [PubMed] [Google Scholar]
- 27.Bolton EC, So AY, Chaivorapol C, Haqq CM, Li H, Yamamoto KR. Cell- and gene-specific regulation of primary target genes by the androgen receptor. Genes Dev. 2007;21:2005–2017. doi: 10.1101/gad.1564207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Baron A, Migita T, Tang D, Loda M. Fatty acid synthase: A metabolic oncogene in prostate cancer? J Cell Biochem. 2004;91:47–53. doi: 10.1002/jcb.10708. [DOI] [PubMed] [Google Scholar]
- 29.Mounier C, Bouraoui L, Rassart E. Lipogenesis in cancer progression (review) Int J Oncol. 2014;45:485–492. doi: 10.3892/ijo.2014.2441. [DOI] [PubMed] [Google Scholar]
- 30.Jiang J, Nilsson-Ehle P, Xu N. Influence of liver cancer on lipid and lipoprotein metabolism. Lipids Health Dis. 2006;5:4. doi: 10.1186/1476-511X-5-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Miyanari Y, Atsuzawa K, Usuda N, Watashi K, Hishiki T, Zayas M, Bartenschlager R, Wakita T, Hijikata M, Shimotohno K. The lipid droplet is an important organelle for hepatitis C virus production. Nat Cell Biol. 2007;9:1089–1097. doi: 10.1038/ncb1631. [DOI] [PubMed] [Google Scholar]
- 32.Meissner EG, Lee YJ, Osinusi A, Sims Z, Qin J, Sturdevant D, McHutchison J, Subramanian M, Sampson M, Naggie S, et al. Effect of sofosbuvir and ribavirin treatment on peripheral and hepatic lipid metabolism in chronic hepatitis C virus, genotype 1-infected patients. Hepatology. 2015;61:790–801. doi: 10.1002/hep.27424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wu YL, Peng XE, Zhu YB, Yan XL, Chen WN, Lin X. Hepatitis B virus X protein induces hepatic steatosis by enhancing the expression of liver fatty acid binding protein. J Virol. 2015;90:1729–1740. doi: 10.1128/JVI.02604-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Haga Y, Kanda T, Sasaki R, Nakamura M, Nakamoto S, Yokosuka O. Nonalcoholic fatty liver disease and hepatic cirrhosis: Comparison with viral hepatitis-associated steatosis. World J Gastroenterol. 2015;21:12989–12995. doi: 10.3748/wjg.v21.i46.12989. [DOI] [PMC free article] [PubMed] [Google Scholar]