

Abstract
An estimated 165 million children globally have stunted growth, and extensive growth data are available. Genome scale metabolic models allow the simulation of molecular flux over each metabolic enzyme, and are well adapted to analyze biological systems. We used a human genome scale metabolic model to simulate the mechanisms of growth and integrate data about breast-milk intake and composition with the infant’s biomass and energy expenditure of major organs. The model predicted daily metabolic fluxes from birth to age 6 months, and accurately reproduced standard growth curves and changes in body composition. The model corroborates the finding that essential amino and fatty acids do not limit growth, but that energy is the main growth limiting factor. Disruptions to the supply and demand of energy markedly affected the predicted growth, indicating that elevated energy expenditure may be detrimental. The model was used to simulate the metabolic effect of mineral deficiencies, and showed the greatest growth reduction for deficiencies in copper, iron, and magnesium ions which affect energy production through oxidative phosphorylation. The model and simulation method were integrated to a platform and shared with the research community. The growth model constitutes another step towards the complete representation of human metabolism, and may further help improve the understanding of the mechanisms underlying stunting.
Systems biology: Feeding breast milk to a computer model of a baby
A research group headed by Prof. Jens Nielsen from Chalmers University of Technology built a computer model of a newborn baby (0–6 months). The computer model simulated a baby’s metabolism of breastmilk, predicting growth in weight, fat and height. An optimization algorithm was used to identify the best flow of nutrients through the metabolic network, from milk to biomass. The researchers used the model to determine if the availability of any nutrient limits growth, e.g., essential amino acids. They found that all nutrients are available with good margin and that growth is limited by the production and consumption of energy. The model may be useful to better understand how malnutrition leads to stunted growth, which affects 165 million children worldwide. Additionally it represents a major advancement toward a complete computer model of the living human.
Introduction
Growth stunting affects 165 million children aged <5 years and is defined by height falling two standard deviations below a reference population median.1 Stunting is associated with decreased cognition, memory, and loco-motor skills, as well as increased mortality. Therefore it is important to understand its underlying mechanisms to identify targets for field studies, nutrition surveys, and policy interventions, to decrease the global burden of stunting. The process of stunting begins in the fetal stage and improved maternal nutrition is associated with increase birth weights.2 Infants grow at their highest rate during their first 6 months after birth, more than doubling their body weight from approximately 3 to 7 kg,3 and there is increasing realization that this is a critical time for growth.1 From the age of 6 months to 24 months nutrient intake is challenged by limited quality and quantity of complementary foods, and it is traditionally seen as one of the most critical periods.
Human tissues may be classified broadly as fat and lean mass. Fat mass is used for energy storage and requires low amounts of energy for maintenance. Lean mass consists mostly of water and proteins and is responsible for energy-demanding functions of the body. Many factors may affect the rate of growth including the lack of essential precursors, insufficient energy supply, co factor deficiencies, hormonal signals, or bio-physical constraints. During growth a series of simultaneous anabolic and catabolic biochemical reactions occur. The anabolic reactions use nutrients from food to synthesize macromolecules, e.g. DNA, phospholipids and proteins, and precursors, e.g. nucleotides and fatty acids. The catabolic reactions degrade nutrients to provide energy to fuel the anabolic reactions and to maintain the body’s functions.
Infant growth has been investigated by varied statistical and modeling methods. Weight changes during catch-up growth following malnutrition, has been modeled using the energy expenditure for tissue synthesis and for maintenance of the lean mass,4 and monthly protein storage and energy expenditure has been calculated in healthy infants.5 Maintenance energy is twice as high in infants as in adults, which has been modeled successfully as an effect of the relative organ weights,6 e.g. the brain constitutes 2% body mass (and uses 20% of the energy) in adults, compared with 10% body mass in infants. Both carbohydrates and fat may be used as energy sources, and the relative use of these nutrients and overall energy requirements was estimated by modeling, using interpolated CO2 excretion rates.7 Although these models provide important insights and predictions, they do not analyze the underlying biochemistry which limits the mechanistic interpretation of the results.
A genome scale metabolic model (GEM) is a mathematical representation of a metabolic network and they have been used to model the processes underlying biological phenotypes in many organisms.8 Although originally developed to study microbes, the method has been extended to human metabolism.9–11 The stoichiometry of all biochemical reactions are combined to an equation system, where the specific flux of each reaction is computationally determined. The cells are assumed to be in a quasi-steady-state, where the production rate of each molecule is matched with the consumption rate. This is justified by the dilution of metabolites due to growth being much smaller than the overall flux. Together with experimentally determined uptake rates it is possible to calculate the optimal allocation of fluxes for any given metabolic task, e.g. determine the maximum growth rate.
In this study we exploit the detailed knowledge on metabolic uptake rates that are available for exclusively breast-fed infants aged 0 to 6 months, to predict human growth. We combine the experience from traditional growth models with GEMs and are able to provide predictions of metabolic fluxes with enzyme level resolution on a day-to-day basis. The developed methods and computer code was integrated to a platform; Simulation Toolbox for Infant Growth with focus on metabolism (STIG-met), and shared with the research community.
Results
Modeling infant growth
We generated a mechanistic model of infant growth (Fig. 1a-b) based on a generic human GEM (HMR 2.00),12 that was augmented with biomass equations for lean and fat mass.13, 14 To specify the distribution of growth between fat and lean mass we calculated an age-dependent parameter based on published information on changes in infant body composition. The fat content of infants is 10% at birth, and increases to 25% during the first months.15 A consequence of the increase in relative fat mass, was that the fat content of newly formed biomass initially was high (40%) and gradually decreased to 25% (Fig. S1A-B). This was consistent with other studies of infant body composition despite some variation in timing and level of the effect.16–19 We also established an empirical relation between lean-mass and length from the composition data15 as [height in cm] = 30.6 × [lean mass in kg]0.45 (Fig. S1D).
Fig. 1.

Simulation of the metabolism of a growing infant from age 0 to 6 months. a The weight and fat mass of a healthy infant were chosen as the initial state of the simulation. Iterative simulations of the infant’s metabolism were performed, in which the current state was used as input to simulate the next state. The simulations were run in a human metabolic model (HMR 2.00) with the intake of metabolites from breast milk as uptake fluxes. The maintenance energy expenditure was calculated from the current body composition, with an additional term for physical activity, which was a function of both age and weight. The objective of the simulation was set to maximize growth, after expending the maintenance energy. Growth was defined as forming the biomass components (fat and lean mass), and the synthesizing energy required. The ratio of fat to total mass in the biomass composition was a function of age. b Simplified representation of the GEM (HMR 2.00) which contains 3800 Genes, 3000 unique metabolites and 8000 reactions out of which 1400 are cofactor dependent. c The weight increments from the simulation were expressed as a growth trajectory and compared with the 5th, 25th, 50th, 75th, and 95th percentile of WHO reference data.3 d The ratio of fat mass to total mass at each time step, compared with the mean and two standard deviations, from a previous study15
The molecular composition of human breast milk was used as input to the model. Published data about the age-dependent intake amount (l day−1)20, 21 and composition (mmol l−1)22, 23 of breast milk were integrated to calculate metabolic fluxes (mmol day−1). The intake of breast milk is initially low ( <100 ml/day), but rapidly increases over the first week to 500 ml, 700 ml at one month and 800 ml at six months of age (Fig. S1C). The molar ratio of most amino acids remains stable, but the protein content decreases from 20 gl−1 to 10 gl−1 over the first 2 months, and is replaced by increasing fat content, from 30 gl−1 to 40 gl−1.22 The human gut-microbiome is known to provide short chain fatty acids, generated by microbial fermentation of dietary polysaccharides24 for simplicity we avoid explicit modeling of the gut microbiome and treat polysaccharides as their glucose equivalents.
To obtain a mechanistic representation of maintenance energy during growth, we combined information about the age-dependent mass of the brain, heart, kidney, and liver, and the residual lean and fat mass with information about their specific energy expenditure (kcal kg−1day−1)6 (Fig. S2A-B). Most data about organ mass were collected from literature,25 but brain mass was estimated using World Health Organization (WHO) head circumference measures and an empirical formula.26 The estimated brain-mass values were in agreement with experimental measurements27 of brain volume that where available for the first 3 months (Fig. S2A). The sum of energy expenditure from all lean tissues was found to be proportional to the lean mass from birth to 6 months 59 kcal kg−1 lean mass day−1 (Fig. S2C). This corresponded to 53 kcal per kg body weight in the newborn, consistent with experimentally determined values (49 kcal).28 At age 5 months the maintenance energy plus the energy required for growth was estimated to 55 kcal per kg, consistent with the sleeping metabolic rate for infants of that age (60 kcal).29
We factored in the additional energy that is spent on tissue synthesis and physical activity. The growth-related energy expenditure may vary with protein (5.5 kcal/g) or fat synthesis (1.6 kcal/g).30 These energy costs were recalculated to ATP expenditure and integrated into the biomass equations. The high synthesis cost of protein partly compensated for the higher energy density of fat mass, caused by the absence of water and higher energy content. However synthesize of fat still required 5-fold more kcal/g.
The energy expenditure due to physical activity depends on both on developmental age and weight. Physical activity, as defined by the difference between sleeping and total metabolic rate, is around 14.4 kcal kg−1 in the 5-month-old infant,29 but setting this value as a constant for all ages would be overestimating the energy expenditure in the newborn, where it has been estimated to 2.4 kcal kg−1 31 in the prematurely born and 4.2 in the surgically newborn,32 as well as at 3 months where it has been estimated to 10 kcal kg−1 33 The degree of physical activity has previously been estimated using predicted metabolic rates at 1.5, 3, and 6 months.34 We use these predictions to estimate the intermediate age-dependent changes in physical activity (Fig. S2D).
To integrate these data with GEM simulations, we developed STIG-met, which included several independent software modules (MATLAB, MathWorks, Natick, MA, USA) that configured the parameters to the GEM, storage objects that held the simulated fluxes and weight increments, and a plotting function that generated human readable representations of the data. Further more, STIG-met compared the results to relevant literature values. The parameter modules were separated into a food module, which specifies the uptake fluxes for each metabolite (mmol/day); a fat module, which specifies the distribution between fat and lean mass in the newly formed tissue (g/g); and an activity module that specifies the energy spent on activity (kcal/kg). We made STIG-met available at the Human Metabolic Atlas repository (www.metabolicatlas.com).35
For simulation, the model iteratively solved a growth maximization problem for each day from birth to age 6 months. The GEM was constrained by milk intake and energy production, and the requirements for synthesis of fat and lean mass according to the age-related profile. The predicted increments in weight and fat mass were accumulated, and used by the model to calculate energy expenditure in the subsequent time step, i.e. day. The weight and fat mass growth was successfully described using the model (Fig. 1c-d), partly due to the introduction of the age dependent physical activity for a better fit to the reference trajectory, and that the growth of fat mass was controlled by a profile directly estimated from reference data. Female infants consume 7% less breast milk than males of the same age.36 We could accurately simulate female growth curves by adjusting milk intake and fat storage (Fig. S3). We also achieved acceptable simulations of the weight of boys up to 12 months, by extrapolating the milk intake (Fig. S4).
Energy is the main growth-limiting factor
The breast milk contains many metabolites that are essential for growth. We investigated the effect of the metabolites in the breast milk by perturbing them and evaluating the effect on predicted growth (Table S1). This sensitivity analysis listed lactose and oleate as the two most important metabolites, suggesting the hypothesis that energy is the main growth limiting factor, because these metabolites primarily serve as energy sources. To test this hypothesis we ran simulations with energy expenditure set to zero (Fig. 2a). This resulted in more than doubling of both fat and lean mass growth.
Fig. 2.

Energy is the main growth limiting factor. a Simulations were run with the energy for maintenance and activity set to zero. The optimal growth of lean and fat mass at each time point was calculated in 2 independent simulations and compared to reference data, calculated from the weight increment3 and the interpolated distribution between fat and lean mass.15 The maximum possible growth was on average 2.5 and 5.6 times higher than the reference growth for lean and fat mass respectively. The smallest difference between maximum and reference growth occurred from birth to age 2 months. b At 1 month age, the uptake of all amino acids from milk was higher than the amount of stored amino acid, except for the nonessential amino acid glycine
Essential amino acids cannot be synthesized by the human body and must be provided by the diet. Using the model we showed that the supply of theses metabolites did not limit growth under the studied conditions (Fig. 2b). Our sensitivity analysis showed that doubling the amount of any amino acid had negligible effect on growth (Table S1), in agreement with experimental results.37 Due to the energy limitation and excess amounts of amino acids, large proportions of the amino acids were converted to energy, observed by nitrogen (i.e. amino acid) retention <50% after age two months (Fig. S5). This result was consistent with the observation that the nitrogen retention is a function of the available energy.38 The high amount of degradation suggested that the exact mol ratio of amino acids in breast milk may be of less importance, since the distribution could be remodeled by selective degradation.
Linoleate and linolenate and their derivatives are essential fatty acids, and severe deficiency decrease growth in the rat.39 Our sensitivity analysis showed that increasing linoleate strongly increased growth (Table S1). However, we saw no growth limitation from these metabolites in the zero energy model (Fig. 2), which suggested that the sensitivity came from linoleate as an energy source. In rats the molecular composition of fat mass may adjust itself to the fat composition of the diet,40 further lowering the requirement of essential fatty acids. The non-essential fatty acids may be provided by the diet or synthesized de novo from e.g. carbohydrates or other fats. Since energy is limited and the energetic cost of lipid synthesis is high, the fatty acids are in general not broken down and resynthesized, consequently the distribution of fatty acids stored is expected to depend on the distribution in milk.
Although our simulations showed that essential amino and fatty acids were provided in sufficient quantities to ensure growth in the healthy breast-fed infant, these molecules cannot be synthesized by the mother and the content in breast milk may be affected by the mother’s diet. Malabsorption or competition for the nutrients by the gut microbiota may also skew the distribution of nutrients that enter the system. Our sensitivity analysis (Table S1) showed that reducing methionine or lysine to half of their concentrations has a significant negative effect on the growth during the first 3 months. These metabolites could therefore become growth-limiting if breast milk from the wrong time period were to be offered to the infants, since the protein content is twice as high during the first 2 months compared to the content at 5 months.22
The energy required for growth may be provided by fat or carbohydrates. In agreement with previous models of child growth,7 we observed that the primary energy source changed during infancy. The relative use of fat for energy generation may be determined from the respiratory quotient (RQ), defined as the ratio of CO2 produced to the O2 consumed. Energy provided by isolated macronutrient groups results in RQ 0.7 for fat and 1 for carbohydrates. The RQ was low, close to 0.7 in newborn, quickly rising to 0.92 during the first days after birth, and then slowly decreasing to 0.85 at age 4 months (Fig. S6A-B), in agreement with literature values.41, 42 We also observed a shift from glycolytic energy production toward beta oxidation of fat directly in the heat map of fluxes from different metabolic subsystems (Fig. S6C). This may be triggered by a higher degree of fat storage in early infancy and shifts in the macromolecular composition of the breast milk.
Growth sensitivity depends on the maintenance energy
The use of the energy intake in the infant is divided between maintenance energy and growth. A simplified relation between the total energy intake (T), maintenance energy (m) and growth (μ) is given in Eq. (1). Growth is sensitive to the total intake, which can be described by the energy elasticity of growth (), a measure of how a small change in T is amplified in μ. The elasticity depends on the ratio of maintenance energy to total energy (Eq. (2), see methods for derivation).
| 1 |
| 2 |
This means that if the maintenance energy is high, as in humans, then a small change in energy intake has a large effect on growth (), whereas if the maintenance energy is negligible, as in many microorganisms growing at their maximum rate, then growth is directly proportional to the total energy intake (). Assuming that maintenance requires 80% of the energy intake, then a 10% decrease in energy intake will decrease growth by 50%, but if maintenance is 0%, then growth will decrease by only 10%. The high-energy elasticity of growth stands in contrast to most other growth-related parameters, e.g. a growth-limiting amino acid, would have an elasticity of at most 1.
Simulated effects of food insecurity and disease
We investigated the effect of several variables related to growth by generating perturbed models (Fig. 3). We were particularly interested in the effects of over-feeding and underfeeding and differences in energy expenditure. Since height is a morphological property, we focus on the effects on weight in this study (Fig. 3a-b), we do however also attempt to study the effect on length (Fig. 3c-d) using its relationship with lean mass, which is explicitly modeled. The growth of all models, including the control, was lower than the WHO standard during the first 15 days after birth,3 due to low levels of milk intake. The discrepancy was likely caused by insufficient resolution in the WHO data, because independent studies of this age group confirmed that breast-fed infants lose 5% weight in the days following birth, and regain it around 10 days after birth.43
Fig. 3.

Perturbations to the reference model. The Energy spender model was defined by a 30% increase in the maintenance expenditure for lean mass. The “Starved” model had decreased milk intake to 50% from age 40–80 days. The overfed and underfed models had +10% and −10% change in intake of breast milk. The over active model spent twice as much energy on physical activity. a Body weight. b z-scores of weight, the number of standard deviations from the mean. c Height. d z-scores of height
Growth is sensitive to over-feeding and underfeeding. The sensitivity increased with age (Table S1), due to increasing energy expenditure caused by increased activity and to decreased specific milk uptake rate (Fig. S7). Our simulations showed that the weight and height remained around one standard deviation of the mean during the first 6 months in life for milk intake varied ±10% around the mean. It is possible that the model overestimated the weight gain of overfeeding, since fat storage likely is higher in an overfed infant. The current implementation of the model chooses the allocation of fat mass and lean mass based on population averages in that age, rather than the nutritional status. In rats excess intake of nutrients is predominately stored as fat after 50 days of age.44 This is likely also the reason behind the more extreme effects on height by the perturbations.
Changes in energy expenditure also affected growth. Many factors may contribute to elevated energy expenditure in humans, e.g. different types of disease,45 temperature,46 and physical activity. High-energy expenditure has been associated with growth faltering in rural populations,47 and chronic inflammation is associated with stunting.1 In our model, the energy expenditure for activity and maintenance only depended on the age and body composition, but in the living infant this may be subject to adaptation. We investigated perturbed models (Fig. 3) and the sensitivity of energy related variables (Table S2) and found that increased energy expenditure of lean mass had a strong negative effect on weight gain(elasticity 0.5–4, depending on age), albeit lower than for changes in the food supply. The effect of changes in synthesizing energy was lower (elasticity 0.1−0.2). The effect of physical activity was marked (elasticity up to 1.4), but came with a time delay due to the modeled increase in activity with age. We observed that the z-score of weight and height remained within 2 standard deviations of the mean for long periods of time regardless of the modeled perturbation, since growth is a cumulative function.
The rate of growth depended on the maintenance energy expenditure, which depended on the amount of lean mass. This relation suggests a self-regulatory mechanism, in which lower weight decreases the energy expenditure and enables increased catchup growth, as observed in the model of starvation (Fig. 3). This effect may be strong for stunted infants, who have low length for age. For simulations with birth weight at the 5th and 95th percentile we observed rapid convergence towards the norm (Fig. S8), it should be noted that the specific milk uptake rate of the underweight children is higher due to their lower weight, and that in the living child this may exceed the gastric or enzymatic capacity. This might be less effective in malnourished infants, with low weight for age, because energy-intensive organs will constitute a larger share of their lean mass. Previous work has shown that the maintenance energy expenditure in malnourished infants is high.4 Therefore, growth constitutes a risk of energy deficiency, since it irreversibly increases the infant’s maintenance energy demand. The growth of fat mass during the first months may have evolved to ensure that infants are provided with an energy buffer that ensures a secure commitment to growth in subsequent months.
Effect of mineral deficiency is linked to energy metabolism
Mineral cofactors are essential parts of the human biomass, and many enzymes rely on them for their catalytic activity, especially energy metabolism is well-known to be influenced by mineral deficiencies.48 We made use of our detailed model of metabolism to study the effects of different cofactor deficiencies. We collected information about the cofactor requirement for each enzyme present in the GEM and simulated cofactor deficiency by constraining the flux through reactions catalyzed by enzymes containing in turn: calcium, cobalt, copper, iron, magnesium, manganese, potassium, and zinc. We observed that deficiencies in magnesium, iron, and copper had the strongest effect on growth (Fig. 4a). These are involved in energy generation through oxidative phosphorylation (Fig. 4b) . The effect of cofactor deficiency was smaller for zinc which was not found to be involved in energy metabolism (Fig. 4c). The effect instead arose from its role in macromolecular synthesis. For the remaining cofactors there was no observed effect on growth, suggesting that they affect growth by other mechanisms of action. It is important to note that the simulations only cover metabolism, many cofactors are involved in other cellular functions that may affect growth, and there may also be enzymatic interactions that are not listed or still are unknown. The rate of relative weight loss decreased with time, mainly due to the decreasing growth rate in the control and the self-regulatory effect of lower weight. For Zn deficiency, this effect was sufficient to stabilize growth at 2 months of age.
Fig. 4.
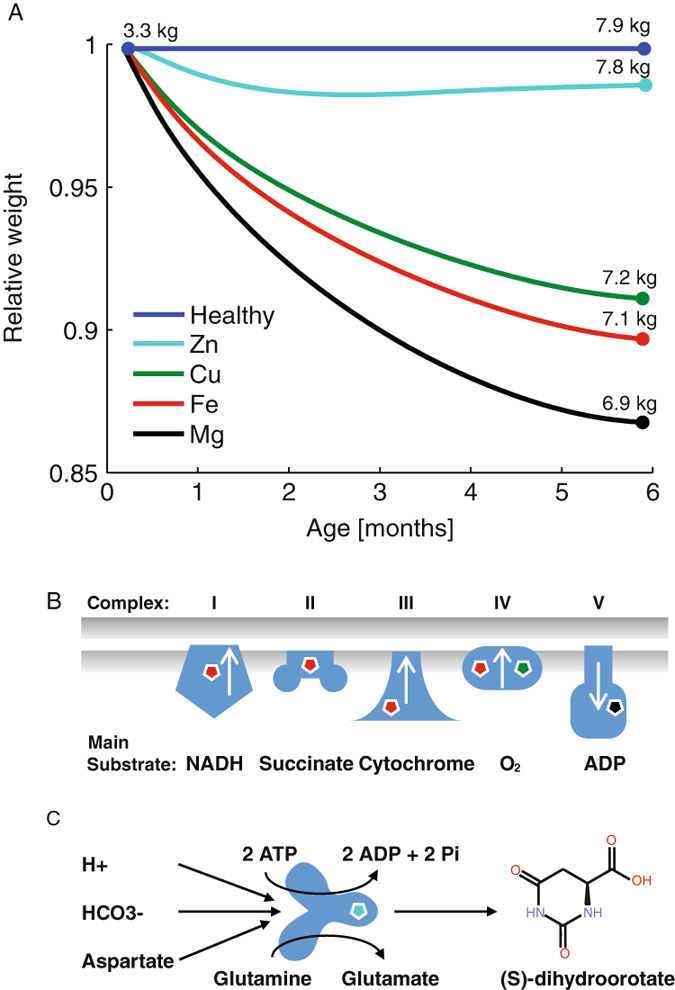
The metabolic effects of co factor deficiency was simulated by decreasing the fluxes through the reactions that depend on the cofactors to 95% optimal value. a The relative weight of infants with different cofactor deficiencies compared to healthy infants. b The strong decrease in growth for Fe, Cu, and Mg deficiency was caused by limitations in the energy generation, Fe and Cu are required for a functional electron transport chain67 and Mg is used by the ATP forming Complex V,68 i.e. ATP synthase. c The Zn deficiency affected the CAD protein (carbamoyl-phosphate synthetase 2, aspartate transcarbamoylase and dihydroorotase), limiting the synthesis of the nucleotide precursor (S)-dihydroorotic acid
In our simulation the effect of cofactor deficiencies is mediated through decreased metabolic flux over groups of enzymes containing these cofactors. In the living child this would result in a buildup of precursor metabolites. This could in turn decrease the absorption rate due to unfavorable metabolite gradients and potentially result in decrease appetite. A buildup of metabolites may also increase the lipid storage, as energy cost is >3-fold higher for synthesizing lean mass than for fat mass.30 It has previously been suggested that iron deficiency may cause obesity through limitations in energy metabolism.49
The detrimental effects of mineral deficiencies has been demonstrated in animal models, e.g. Fe, Mg, and Cu deficiency in new born pigs cause weight reduction to 76, 46, and 50% vs. control after 5–6 weeks.50, 51 The Fe deficiency is associated with underweight in human infants.52 Previous studies showed that children recovering from protein-calorie malnutrition had significantly faster weight gain with magnesium supplements,53 and infants with Cu deficiency experienced improved weight gains when taking Cu supplements.54 Mild zinc deficiency has also been associated with the low height percentiles.55 Although there is qualitative agreement, it is hard to establish a direct link between the in silico effects of cofactor deficiencies and clinical data. In the living child the likelihood of cofactor deficiency appears to be a more important factor than the severity when it occurs. Our model does not take bodily reserves in to account, but the healthy infant is born with Fe stores that persist for six months.56 The model does not take in to account the binding dynamics of cofactors, or the possibility of residual activity or cofactor substitutions. It does however showcase that also mild mineral deficiencies may have a strong influence on growth.
Discussion
We here integrated a GEM with anthropomorphic data to simulate infant growth, and modeled the enzymatic steps synthesizing biomass from food. We observed that essential amino and fatty acids were available in sufficient quantities to ensure growth in breast-fed infants, and that energy was the main growth-limiting factor, in agreement with literature. Disturbances to the energy balance had a strong effect on growth, e.g. disturbances in food supply, elevated maintenance expenditure or deficiencies in cofactors for the enzymes involved in oxidative phosphorylation. Maintenance energy could not be compromised in our model, implying that essential nutrients were degraded if energy was insufficient. Some studies of antibiotic usage during infancy imply increased weight and length,57 this may potentially be related to decreased energy expenditure for inflammation or increased nutrient utilization due to decreased competition from the gut microbiota.
A hypothesis that would require experimental validation is that supplementing breast milk with glucose solution potentially may rescue growth under energy-stressed conditions. Studies on piglets supplemented with 5% glucose solution showed no significant increase in growth,58 but lambs had improved growth rate when treated with glucose infusions compared to saline controls.59 Although our models showed that energy limited growth, further increased energy intake may not necessary translate into increased growth, because other factors, e.g. growth hormones and enzymatic capacity, may limit cell proliferation. Excessive energy intake may instead be stored as fat, converted to heat or not absorbed. It appears biologically sound for energy to be growth-limiting, because this ensures that all other nutrients will be present in sufficient quantities, which may simplify resource allocation.
There are several limitations to the current modeling approach. Many parameters were integrated, sometimes with marked uncertainty and high elasticity (Table S2). Due to the stepwise increase in body mass, the model did not account for increasing maintenance energy throughout the day due to growth, which may increase growth predictions with as much as 1% during the most growth-intensive periods. The brain mass, which is a major consumer of energy, was estimated from head circumference, because of absence of data; other studies suggest that the energy expenditure of the brain may change with age due to differences in neuronal levels.60 Although the specific energy expenditure of the other organs were assumed to be constant and the same as in adults, energy expenditure in infants may be higher e.g. due to increased metabolic load, or lower, e.g. due to the purity of the nutrient source. This may explain why the calculated energy expenditure in the newborn is slightly higher than suggested in some studies, 53 kcalkg−1 compared to 49 ± 6.28 The energy expenditure is complemented with an activity parameter with only partial empirical evidence. Therefore, the input and output of metabolites is expected to be in good agreement with the state in the healthy infant, but it cannot be excluded that the mechanistic reason may be partially mislabeled, e.g. increased energy expenditure of the brain being labeled as physical activity.
In this study the biomass equation was limited to the most abundant macromolecules. Therefore the modeled fluxes were dominated by transport reactions and energy metabolism. This is likely an accurate description of the growing infant, growth is primarily an assembly of nutrients from the diet. However, a more complete model would also include vitamins and other micro nutrients, which would require a greater share of the metabolic network to be used, albeit with much lower flux. A model including the division of labor between multiple tissues would also utilize a larger share of the network, e.g. through the production of ketones and recycling of lactate by the liver.9 In addition a more complete model would have more complex maintenance reactions, with separation into different metabolic tasks, weighted by importance. Other types of maintenance could also be included, e.g. spontaneous degradation of amino acids, that may narrow the gap between provided and used nutrients. Furthermore, a more complete model would need to consider fat allocation, including pure production of fat when the target for lean mass growth is satisfied, a more dynamic description of food intake, e.g. as a function of lean mass, and nutrient-dependent absorption quotients.
We here created the STIG-met modeling framework and integrated data from diverse sources. We focused on healthy males aged 0–6 months, and future studies may extend STIG-met to fetal growth and older childhood, provided that the metabolic content of the nutrient source (placental blood or diet) is known. This is required due to the model structure, where balances are included for each individual metabolite. The model is well-suited to be integrated with metabolic models of the gut microbiota, e.g. to simulate the host microbe interactions in short chain fatty acid and amino acid metabolism.24 With adequate data, the model could be used to investigate individual growth trajectories and potentially expose underlying reasons for malnutrition or obesity. As an integration of known modeling methods and parameter sets, STIG-met may be a major advancement toward a complete mechanistic model of the living human.
Methods
Model parameters
The GEM (HMR 2.00)12 was parameterized using literature data and used for growth simulations. The parameters were primarily related to breast milk, biomass, and energy expenditure.
The intake of breast milk (ml/day) was taken from 2 studies; a meta study of breastfeeding by WHO20 with intake determined at 1, 2, 3, 4, 5, and 6 months and a meta study of breast-milk intake at age 1, 2, 3, 5, and 7 days.21 The pchip (Piecewise Cubic Hermite Interpolating Polynomial) function (MATLAB, MathWorks, Natick, MA, USA) was used to interpolate the breast-milk intake between measured points. The protein content per liter milk (g/l) and amino acid composition (mmol/g) of the protein was taken from a study by Yamawaki et al.;22 the quantities were measured at age 1– 5, 6– 10, 11– 20, 21– 89, 90– 180, and 181– 365 days, and the amino acid content per liter milk was interpolated between the measured times. The lactose, glucose, and polysaccharide content (g/l) was taken from the same study and converted to mol/l. The polysaccharides were taken as their glucose equivalents. The fat content (g/l) was taken from this study, the fatty acid composition (mol/g) was however not determined and therefore taken from another study,23 measured at day 5 and 42. The digestible fraction of breast milk was taken as 93%61, and the intake of all nutrients were reduced to 93%.
Biomass composition was collected from several studies. Fat mass was modeled as the fatty acid composition (mmol/kg) of the adipose tissue of infants aged 0–2 years.14 The chosen water content of the lean mass was 80%, and the protein content 80% and glycogen content 4% of the dry weight, based on literature estimates.16 The RNA and DNA content were assumed to be 1% of the dry weight each. The amino acid composition was modeled as the fetal, with values for tryptophan and cysteine from the fetal pig, because these amino acids were not quantified in human.13 The fat mass per kg body weight (kg/kg) was taken from a multicenter study15 that had measurements at age 1 and 2 weeks, and then monthly up to 6 months of age. The data were fitted to a polynomial of degree 3, giving fat mass as a function of body weight. The fat mass at any time could then be estimated from the weight at that time, which was obtained from WHO data. The daily delta fat ratio was calculated as the increase in fat content divided by the increase in body weight for each day.
The resting energy expenditure for lean mass was estimated as 59 (kcal/kg lean mass) from the organ energy expenditure and composition. The energy expenditure for each organ (200 for liver, 240 for brain, 440 for heart and kidneys, 13 for skeletal muscle, 4.5 for adipose tissue, and 12 for the residuals all expressed as kcal per kg organ) was taken from a previous study,6 and the organ weight was taken from literature.25 Data for brain weight was not available and was estimated by an empirical relation between head circumference and brain weight,26 with head circumference data from WHO.62 A correction factor for converting the specific energy expenditure of adult to infant organs has been suggested, based on relative cellularity and quantified by the potassium lean mass ratio;6 however this factor was dropped in the present study, because the uncorrected energy expenditure was in better agreement with experimental values, and the potassium lean mass ratio remained constant over the study period.63 The energy expenditure for tissue synthesis was taken as 5.5 kcal/g lean mass and 1.6 kcal/g fat mass from literature.30 Additional energy expenditure for physical activity was calculated from the difference between sleeping energy expenditure and total energy expenditure, 14.4 kcal/kg in infants aged 4–6 months.29 This value was modulated with a factor between 0 and 1 based on estimated changes in physical activity from literature,34 and interpolated using a polynomial of degree 2. The energy expenditure was converted to 28.12 mmol ATP hydrolyzed per kcal; estimated by simulating the production of ATP from glucose with known energy content of 4 kcal/g. Conversion of fat (9 kcal/g) giving similar results.
Simulations
To simulate the growth curves, iterative growth maximization problems were solved using the RAVEN Toolbox for Flux Balance Analysis (FBA),64 resulting in predicted increments in lean and fat mass. The accumulated lean and fat mass were used to estimate the maintenance energy expenditure at the next time step, and the model was constrained to expend this energy through an artificial reaction hydrolyzing ATP to ADP. If there was no feasible solution to the growth maximization problem (due to maintenance energy exceeding the input), a minimization problem was solved, in which the energy was provided from the stored fat mass. For the standard growth curve, the ratio of fat in the biomass equation was chosen from the interpolated experimental data. For the lean and fat mass optimization problem, the maximization was set to produce either lean or fat mass. The z-scores of the growth curves were calculated using WHO data (available at the WHO web page http://www.who.int/childgrowth/standards, retrieved 2016-09-07).3 The fluxes depicted in the heat map (Fig. S6) were calculated using parsimonious FBA.65
The cofactors associated with reaction in the GEM were extracted from a protein database (UniProt)66 based on their gene association. The reactions in oxidative phosphorylation where manually curated with cofactor dependence from literature.67, 68 To estimate the growth curves with cofactor limitation, the following simulation was performed; for each time-step a simulation was run establishing the norm fluxes, and a second optimization problem was solved minimizing the flux through the cofactor affiliated reactions, while maintaining the optimal solution. The fluxes were constrained to a fraction of these fluxes and the growth optimization was rerun.
Sensitivity analysis
The sensitivity of the model parameters was investigated by calculating their elasticities E at different times in the simulation. Each parameter p was perturbed by a factor d = 1.001 (Eq. 3). The change in predicted growth rate was calculated by comparing the maximum growth rate in the perturbed state μ i to the non-perturbed state μ (Eq. 3).
| 3 |
The effects of nutrient variation (Table S1) was tested by perturbing the uptake rate by a factor 0.5 for severe nutrient limitation and 2 for nutrient exess.
To analyze the sensitivity of energy production the following formula was derived (Eq. 4), where μ is the growth rate, T is the energy production and m the maintenance.
| 4 |
Electronic supplementary material
Acknowledgements
The authors want to thank Shasha Jumbe, Elly Trepman, Elias Björnson, Jun Geng and Rasmus Ågren for valuable comments. The authors acknowledge funding from the Bill and Melinda Gates Foundation and the Knut and Alice Wallenberg Foundation.
Competing interests
The authors declare no competing interest.
Author contributions
A.N. constructed the model, performed model simulations, and analyzed the data. A.M. supervised the work. J.N. supervised the work and conceived the idea. A.N. drafted the paper. A.M. and J.N. edited the paper.
Footnotes
Electronic supplementary material
Supplementary Information accompanies the paper on the npj Systems Biology and Applications website (doi:10.1038/s41540-017-0004-5).
References
- 1.Prendergast AJ, Humphrey JH. The stunting syndrome in developing countries. Paediatr. Int. Child Health. 2014;34:250–265. doi: 10.1179/2046905514Y.0000000158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Gluckman PD, Hanson Ma. Living with the past: evolution, development, and patterns of disease. Science. 2004;305:1733–1736. doi: 10.1126/science.1095292. [DOI] [PubMed] [Google Scholar]
- 3.World Health Organization, WHO child growth standards length/height-for-age, weight-for-age, weight-for-length, weight-for-height and body mass index-for-age: methods and development. (World Health Organization, Geneva, 2006).
- 4.Fjeld CR, Schoeller DA, Brown KH. A new model for predicting energy requirements of children during catch-up growth developed using doubly labeled water. Pediatr. Res. 1989;25:503–508. doi: 10.1203/00006450-198905000-00016. [DOI] [PubMed] [Google Scholar]
- 5.Butte NF. Energy requirements of infants. Public Health Nutr. 2005;8:953–967. doi: 10.1079/phn2005790. [DOI] [PubMed] [Google Scholar]
- 6.Wang Z. High ratio of resting energy expenditure to body mass in childhood and adolescence: A mechanistic model. Am. J. Hum. Biol. 2012;24:460–467. doi: 10.1002/ajhb.22246. [DOI] [PubMed] [Google Scholar]
- 7.Jordan PN, Hall KD. Dynamic coordination of macronutrient balance during infant growth: insights from a mathematical model. Am. J. Clin. Nutr. 2008;87:692–703. doi: 10.1093/ajcn/87.3.692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lewis N. E., Nagarajan H., Palsson B. O. Constraining the metabolic genotype-phenotype relationship using a phylogeny of in silico methods. Nat. Rev. Mic. 10, 291–305 (2012). [DOI] [PMC free article] [PubMed]
- 9.Bordbar A. et al. A multi-tissue type genome-scale metabolic network for analysis of whole-body systems physiology, BMC Syst. Biol. 5, 180 (2011). [DOI] [PMC free article] [PubMed]
- 10.Thiele I, et al. A community-driven global reconstruction of human metabolism. Nat. Biotechnol. 2013;31:419–25. doi: 10.1038/nbt.2488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Mardinoglu A, et al. Integration of clinical data with a genome-scale metabolic model of the human adipocyte. Mol. Syst. Biol. 2013;9:649. doi: 10.1038/msb.2013.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mardinoglu A, et al. Genome-scale metabolic modelling of hepatocytes reveals serine deficiency in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014;5:3083. doi: 10.1038/ncomms4083. [DOI] [PubMed] [Google Scholar]
- 13.Wu G, Ott TL, Knabe DA, Bazer FW. Amino acid composition of the fetal pig. J. Nutr. 1999;129:1031–1038. doi: 10.1093/jn/129.5.1031. [DOI] [PubMed] [Google Scholar]
- 14.Sanjurjo P, et al. Fatty acid composition of skeletal muscle and adipose tissue in Spanish infants and children. Br. J. Nutr. 2006;95:168–173. doi: 10.1079/BJN20051605. [DOI] [PubMed] [Google Scholar]
- 15.Fields DA, et al. Longitudinal body composition data in exclusively breast-fed infants: a multicenter study. Obesity (Silver Spring). 2011;19:1887–1891. doi: 10.1038/oby.2011.11. [DOI] [PubMed] [Google Scholar]
- 16.Fomon SJ, Nelson SE. Body composition of the male and female reference infants. Annu. Rev. Nutr. 2002;22:1–17. doi: 10.1146/annurev.nutr.22.111401.145049. [DOI] [PubMed] [Google Scholar]
- 17.Roggero PM, et al. Quality of growth in exclusively breast-fed infants in the first six months of life: an Italian study. Pediatr. Res. 2010;68:542–4. doi: 10.1203/PDR.0b013e3181f85a20. [DOI] [PubMed] [Google Scholar]
- 18.Carberry AE, Colditz PB, Lingwood BE. Body composition from birth to 4.5 months in infants born to non-obese women. Pediatr. Res. 2010;68:84–88. doi: 10.1203/PDR.0b013e3181df5421. [DOI] [PubMed] [Google Scholar]
- 19.de Bruin NC, et al. Standards for total body fat and fat-free mass in infants. Arch. Dis. Child. 1996;74:386–99. doi: 10.1136/adc.74.5.386. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Butte N. F., Lopez-Alarcon M. G., Garza C. Nutrient Adequacy of Exclusive Breast Feeding for the Term Infant During the First Six Months of Life. (World Health Organization, 2002).
- 21.Hester SN, Hustead DS, Mackey AD, Singhal A, Marriage BJ. Is the macronutrient intake of formula-fed infants greater than breast-fed infants in early infancy? J. Nutr. Metab. 2012;2012:891201. doi: 10.1155/2012/891201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Yamawaki N, et al. Macronutrient, mineral and trace element composition of breast milk from Japanese women. J. Trace Elem. Med. Biol. 2005;19:171–181. doi: 10.1016/j.jtemb.2005.05.001. [DOI] [PubMed] [Google Scholar]
- 23.Peng YM, Zhang TY, Wang Q, Zetterström R, Strandvik B. Fatty acid composition in breast milk and serum phospholipids of healthy term Chinese infants during first 6 weeks of life. Acta Pædiatrica. 2007;96:1640–1645. doi: 10.1111/j.1651-2227.2007.00482.x. [DOI] [PubMed] [Google Scholar]
- 24.Shoaie S, et al. Quantifying diet-induced metabolic changes of the human gut Microbiome. Cell Metab. 2016;22:320–331. doi: 10.1016/j.cmet.2015.07.001. [DOI] [PubMed] [Google Scholar]
- 25.Pryce JW, et al. Reference ranges for organ weights of infants at autopsy: results of >1,000 consecutive cases from a single centre. BMC Clin. Pathol. 2014;14:18. doi: 10.1186/1472-6890-14-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cooke RWI, Lucas A, Yudkin PLN, Pryse-Davies J. Head circumference as an index of brain weight in the fetus and newborn. Early Hum. Dev. 1977;1:145–149. doi: 10.1016/0378-3782(77)90015-9. [DOI] [PubMed] [Google Scholar]
- 27.Holland D, Chang L, TM E, et al. STructural growth trajectories and rates of change in the first 3 months of infant brain development. JAMA Neurol. 2014;71:1266–1274. doi: 10.1001/jamaneurol.2014.1638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cai W, et al. Normal value of resting energy expenditure in healthy neonates. Nutrition. 2003;19:133–136. doi: 10.1016/S0899-9007(02)00929-2. [DOI] [PubMed] [Google Scholar]
- 29.Rising R, Duro D, Cedillo M, Valois S, Lifshitz F. Daily metabolic rate in healthy infants. J. Pediatr. 2003;143:180–185. doi: 10.1067/S0022-3476(03)00362-7. [DOI] [PubMed] [Google Scholar]
- 30.Towers HM, Schulze KF, Ramakrishnan R, Kashyap S. Energy expended by low birth weight infants in the deposition of protein and fat. Pediatr Res. 1997;41:584–589. doi: 10.1203/00006450-199704000-00021. [DOI] [PubMed] [Google Scholar]
- 31.Thureen PJ, Phillips RE, Baron KA, DeMarie MP, Hay WW. Direct measurement of the energy expenditure of physical activity in preterm infants. J. Appl. Physiol. 1998;85:223–230. doi: 10.1152/jappl.1998.85.1.223. [DOI] [PubMed] [Google Scholar]
- 32.Pierro A, et al. Partition of energy metabolism in the surgical newborn. J. Pediatr. Surg. 1991;26:581–586. doi: 10.1016/0022-3468(91)90712-3. [DOI] [PubMed] [Google Scholar]
- 33.Butte NF, et al. Energy requirements derived from total energy expenditure and energy deposition during the first 2 y of life. Am. J. Clin. Nutr. 2000;72:1558–1569. doi: 10.1093/ajcn/72.6.1558. [DOI] [PubMed] [Google Scholar]
- 34.Wells JC, Davies PS. Estimation of the energy cost of physical activity in infancy. Arch. Dis. Child. 1998;78:131–6. doi: 10.1136/adc.78.2.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Pornputtapong N., Nookaew I., Nielsen J. Human metabolic atlas: An online resource for human metabolism. Database, 2015. doi:10.1093/database/bav068. [DOI] [PMC free article] [PubMed]
- 36.Wallgren A. Breast-milk consumption of healthy full-term infants. Acta Pædiatrica. 1945;32:778–790. doi: 10.1111/j.1651-2227.1945.tb16864.x. [DOI] [PubMed] [Google Scholar]
- 37.Motil KJ, Sheng HP, Montandon CM, Wong WW. Human milk protein does not limit growth of breast-fed infants. J. Pediatr. Gastroenterol. Nutr. 1997;24:10–7. doi: 10.1097/00005176-199701000-00006. [DOI] [PubMed] [Google Scholar]
- 38.Grimble G., Preedy V. R., Watson R. Nutrition in the Infant: Problems and Practical Procedures . (Greenwich Medical Media, 2001).
- 39.Cunnane SC, Anderson MJ. Pure linoleate deficiency in the rat: influence on growth, accumulation of n-6 polyunsaturates, and [1-14C]linoleate oxidation. J. Lipid Res. 1997;38:805–812. [PubMed] [Google Scholar]
- 40.Cunnane SC, Trotti D, Ryan MA. Specific linoleate deficiency in the rat does not prevent substantial carbon recycling from [(14)C]linoleate into sterols. J. Lipid Res. 2000;41:1808–1811. [PubMed] [Google Scholar]
- 41.Jonxis J. H. P., Vlugt J. J., Groot C. J., Boersma E. R., Meijers E. D. K. The metabolic rate in praemature, dysmature and sick infants in relation to environmental temperature. In BT - Aspects of Praematurity and Dysmaturity: Groningen 10-12 May 1967. 201–209. (Springer, 1968).
- 42.Rising R., Sonmez G. T. Energy expenditure and physical activity in recovering malnourished infants. J. Nutr. Metab., 2010, doi:10.1155/2010/171490. [DOI] [PMC free article] [PubMed]
- 43.Macdonald PD, Ross SRM, Grant L, Young D. Neonatal weight loss in breast and formula fed infants. Arch. Dis. Child. - Fetal Neonatal Ed. 2003;88:F472–F476. doi: 10.1136/fn.88.6.F472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Drewry MM, Harris RBS, Martin RJ. Developmental changes in response to overfeeding: Effect on composition of gain, liver metabolism and adipocyte cellularity in rats. J. Nutr. 1988;118:194–198. doi: 10.1093/jn/118.2.194. [DOI] [PubMed] [Google Scholar]
- 45.Miles JM. Energy expenditure in hospitalized patients: Implications for nutritional support. Mayo Clin. Proc. 2016;81:809–816. doi: 10.4065/81.6.809. [DOI] [PubMed] [Google Scholar]
- 46.Brück K. Temperature regulation in the newborn infant (Part 1 of 3) Neonatology. 1961;3:65–81. doi: 10.1159/000239810. [DOI] [Google Scholar]
- 47.Butte NF, et al. Higher total energy expenditure contributes to growth faltering in breast-fed infants living in rural Mexico. J. Nutr. 1993;123:1028–35. doi: 10.1093/jn/123.6.1028. [DOI] [PubMed] [Google Scholar]
- 48.Huskisson E, Maggini S, Ruf M. The role of vitamins and minerals in energy metabolism and well-being. J. Int. Med. Res. 2007;35:277–289. doi: 10.1177/147323000703500301. [DOI] [PubMed] [Google Scholar]
- 49.Aigner E., Feldman A., Datz C. Obesity as an emerging risk factor for iron deficiency, Nutr. 6(9), 3587–3600 (2014). [DOI] [PMC free article] [PubMed]
- 50.Miller E. R., Ullrey D. E., Schmidt D. A., Luecke R. W., Hoefer J. A. Effects of nutrient deficiencies upon organ weights of the baby pig. J. Anim. Sci.26(5), 1046–1050 (1967). [DOI] [PubMed]
- 51.Hill GM, et al. A copper deficiency in neonatal pigs induced by a high zinc maternal diet. J. Nutr. 1983;113:867–872. doi: 10.1093/jn/113.4.867. [DOI] [PubMed] [Google Scholar]
- 52.Yang W, et al. Anemia, malnutrition and their correlations with socio-demographic characteristics and feeding practices among infants aged 0-18 months in rural areas of Shaanxi province in northwestern China: a cross-sectional study. BMC Public Health. 2012;12:1127. doi: 10.1186/1471-2458-12-1127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Nichols BL, Alvarado J, Hazlewood CF, Viteri F. Magnesium supplementation in protein-calorie malnutrition. Am. J. Clin. Nutr. 1978;31:176–188. doi: 10.1093/ajcn/31.1.176. [DOI] [PubMed] [Google Scholar]
- 54.Castillo-Duran C, Uauy R. Copper deficiency impairs growth of infants recovering from malnutrition. Am. J. Clin. Nutr. 1988;47:710–714. doi: 10.1093/ajcn/47.4.710. [DOI] [PubMed] [Google Scholar]
- 55.Gibson RS, et al. A growth-limiting, mild zinc-deficiency syndrome in some southern Ontario boys with low height percentiles. Am. J. Clin. Nutr. 1989;49:1266–1273. doi: 10.1093/ajcn/49.6.1266. [DOI] [PubMed] [Google Scholar]
- 56.Domellöf M. Iron requirements in infancy. Ann. Nutr. Metab. 2011;59:59–63. doi: 10.1159/000332138. [DOI] [PubMed] [Google Scholar]
- 57.Mbakwa CA, et al. Early life antibiotic exposure and weight development in children. J. Pediatr. 2016;176:105–113.e2. doi: 10.1016/j.jpeds.2016.06.015. [DOI] [PubMed] [Google Scholar]
- 58.Friend DW, Cunningham HM. The effect of water consumption on the growth, feed intake, and carcass composition of suckling piglets. Can. J. Anim. Sci. 1966;46:203–209. doi: 10.4141/cjas66-029. [DOI] [Google Scholar]
- 59.Leng RA, Economides S, Ball FM. The effect on growth of supplying glucose continuously into the duodenum of lambs on low protein diets. Proc. Aust. Soc. Anim. Prod. 1978;12:134. [Google Scholar]
- 60.Kuzawa CW, et al. Metabolic costs and evolutionary implications of human brain development. Proc. Natl. Acad. Sci. 2014;111:13010–13015. doi: 10.1073/pnas.1323099111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Draper H. Advances in Nutritional Research. (Springer, 1990). [DOI] [PubMed]
- 62.Organization, W. H. WHO Child Growth Standards: Growth Velocity Based on Weight, Length and Head Circumference. (World Health Organization, 2009).
- 63.Fomon S. J., Haschke F., Ziegler E. E., Nelson S. E. Body composition of reference children from birth to age 10 years. Am. J. Clin. Nutr.35: 1169-1175, 1982. [DOI] [PubMed]
- 64.Agren R, et al. The RAVEN toolbox and its use for generating a genome-scale metabolic model for penicillium chrysogenum. PLoS Comput. Biol. 2013;9:e1002980. doi: 10.1371/journal.pcbi.1002980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Lewis NE, et al. Omic data from evolved E. coli are consistent with computed optimal growth from genome-scale models. Mol. Syst. Biol. 2010;6:390. doi: 10.1038/msb.2010.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.UNIPROT Activities at the universal protein resource (UniProt) Nucleic Acids Res. 2014;42:D191–8. doi: 10.1093/nar/gkt1140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Roehm, K.-H. In eLS. (Wiley, 2001).
- 68.Ackerman, S. H., Tzagoloff, A. Function, Structure, and Biogenesis of Mitochondrial ATP Synthase, Vol. 80, 95–133. (Academic, 2005). [DOI] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.