

Abstract
Stem cell biology has become an important field in regenerative medicine and tissue engineering therapy since the discovery and characterization of mesenchymal stem cells. Stem cell populations have also been isolated from human dental tissues, including dental pulp stem cells, stem cells from human exfoliated deciduous teeth, stem cells from apical papilla, dental follicle progenitor cells, and periodontal ligament stem cells. Dental stem cells are relatively easily obtainable and exhibit high plasticity and multipotential capabilities. The dental stem cells represent a gold standard for neural-crest-derived bone reconstruction in humans and can be used for the repair of body defects in low-risk autologous therapeutic strategies. The bioengineering technologies developed for tooth regeneration will make substantial contributions to understand the developmental process and will encourage future organ replacement by regenerative therapies in a wide variety of organs such as the liver, kidney, and heart. The concept of developing tooth banking and preservation of dental stem cells is promising. Further research in the area has the potential to herald a new dawn in effective treatment of notoriously difficult diseases which could prove highly beneficial to mankind in the long run.
Keywords: Dental stem cell, stem cell therapy, differentiation, regeneration, tissue engineering, tooth banking
Introduction
The tooth is composed of distinct tissues including the outer mineralized enamel layer; the adjacent mineralized dentin layer; the dental pulp containing blood vessels, nerves, and mesenchymal tissue; and root structures composed of dentin, cementum, and periodontal ligament (PDL), which secure teeth to the underlying alveolar bone. Dentin contains characteristic and distinctive tubules, produced by neural crest derived dental mesenchymal stem cells called odontoblasts, which persist in mature teeth and exhibit limited regenerative capacities to form reparative dentin in response to injury or disease. The dental pulp is composed of dental mesenchymal cells, nerves, and blood vessels that thread through the root canal. Teeth develop through continuous and reciprocal interactions between cranial neural crest-derived mesenchymal stem cells (MSCs) and oral-derived epithelial stem cells during early embryogenesis.1,2
Stem cells can be isolated from several oral tissues such as craniofacial bone, dental pulp, PDL, dental follicle, tooth germ, apical papilla, oral mucosa, gingival, and periosteum.3 The dental stem cells (DSCs) are post-natal stem cell populations that have MSC-like qualities, including the capacity for self-renewal and multilineage differentiation potential. These cells are derived from the neural crest, and thus have a different origin from bone-marrow-derived mesenchymal stem cells (BMMSCs), which are derived from mesoderm.4 Among oral tissue-derived stem cells, human dental pulp stem cells (hDPSCs) have been widely studied due to their great clinical potential, easy accessibility, and less invasive harvesting. These cells were found to form dentin-like tissue and to differentiate into osteoblast-like cells that formed bone in vitro. In the presence of specific stimuli, these DPSCs differentiated into several cell types, including neurons, adipocytes, and chondrocytes. Interestingly, vascular endothelial cells and DPSCs were found to synergistically differentiate into osteoblasts and endothelial cells, respectively.5,6
DSCs have been widely studied due to their great clinical potential, easy accessibility, and less invasive harvesting. Several preclinical investigations conducted so far indicated the extensive potential of the stem cell in tissue repair and regeneration of dental tissues, as well as other organs. This article focuses on the type of DSCs and its potential therapeutic applications in tissue engineering and regenerative medicine.
DSC
The dental pulp is a soft tissue of ectomesenchymal and mesenchymal origin, developing from the dental papilla. Stem cell populations can be isolated from different tissues of the oral and maxillofacial regions. They are stemmed from different developmental stages of the tooth. Around eight unique populations of dental tissue-derived MSCs have been isolated and characterized. Post-natal DPSCs were the first human dental MSCs to be identified from pulp tissue.7 Other dental MSC-like populations, such as stem cells from human exfoliated deciduous teeth (SHED),8 periodontal ligament stem cells (PDLSCs),9 dental follicle progenitor cells (DFPCs),10 alveolar bone-derived mesenchymal stem cells (ABMSCs),11 stem cells from the apical part of the human dental papilla (SCAP),12 tooth germ progenitor cells (TGPCs),13 and gingival mesenchymal stem cells (GMSCs),14 were also isolated and characterized (Figure 1).
Figure 1.
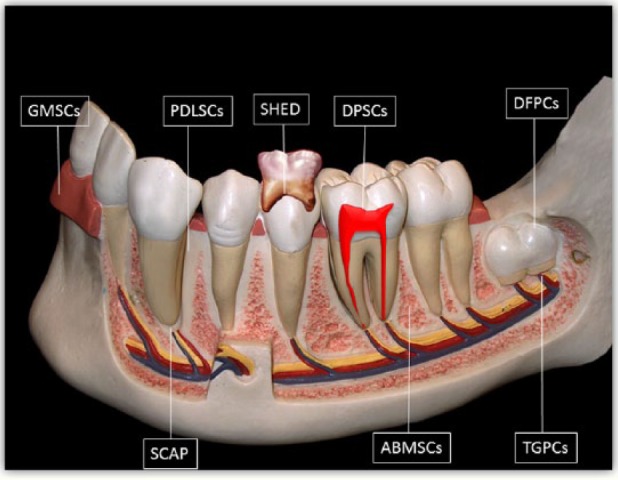
Schematic drawing illustrating sources of human dental tissue-derived MSCs.
ABMSCs: alveolar bone-derived mesenchymal stem cells; DFPCs: dental follicle progenitor cells; DPSCs: dental pulp stem cells; GMSCs: gingival mesenchymal stem cells; PDLSCs: periodontal ligament stem cells; SCAP: stem cells from the apical part of the human dental papilla; SHED: stem cells from human exfoliated deciduous teeth; TGPCs: tooth germ progenitor cells.
Dental pulp-derived stem cells such as human adult DPSCs and SHED are self-renewing MSCs residing within the perivascular niche of the dental pulp.7,8 They are thought to originate from the cranial neural crest, which expresses early markers for both MSCs and neuroectodermal stem cells.15 DPSC and SHED have been reported to demonstrate the ability to regenerate into various tissues. Recently it has been shown that implantation of DPSCs or SHED promotes functional recovery after spinal cord injury (SCI).16 DPSC also protect against ischemic brain injury in neonatal mice.17 DSC have comparable therapeutic potential similar to BMMSCs, and DSC is another alternative noninvasive source to be used for future regenerative therapies.
DPSC
DPSCs were the first type of DSC derived from dental pulp and was isolated by enzymatic digestion of the pulp tissue of the human-impacted third molars. These multipotential cells exhibited a typical fibroblast-like morphology.7 Isolation of DPSC was performed and with various differentiation media, their dentinogenic, osteogenic, adipogenic, neurogenic, chondrogenic, and myogenic differentiation potential were demonstrated.18,19 Even though there is no specific biomarker available for the identification of DPSC, these cells express several markers including the mesenchymal and bone-marrow stem cell markers, STRO-1, and CD146, as well as the embryonic stem cell (ESC) marker, OCT4. The candidate markers of DPSCs include STRO-1, CD29, CD44, CD73, CD90, CD105, CD146, CD166, and CD271.20
DPSC differentiate into adipogenic, chondrogenic, and osteogenic lineages, express epithelial markers and share common characteristics with neural stem cells; they are also able to differentiate, in vitro, into neural or vascular endothelial cells.21,22 DPSCs and endothelial cells have a synergistic effect in co-cultures, enhancing differentiation to osteogenic, odontogenic, and angiogenic phenotypes.23 hDPSCs which are grown by explant culture method have better proliferative capacity and also differentiate into various cell types of osteogenic, adipogenic, and myogenic lineages.24,25 The hDPSC cultures contain multipotent neural crest stem cell (NCSC) which can differentiate into a number of neural crest-derived cell lineages including melanocytes.26 Paino et al.27 showed that DPSC spontaneously differentiate in vitro towards the melanocytic lineage. DPSCs have shown the greatest potential to produce a high volume of mineralized matrix, suggesting that these cells also show promise for use in regenerative dental therapies.28 Dental pulp progenitors have not been clearly identified but some data suggest that pericytes, which are able to differentiate into osteoblasts, could also differentiate into odontoblasts.29
SHED
The stem cells isolated from the dental pulp of exfoliated deciduous teeth revealed their high proliferative and clonogenic nature. Miura et al.8 isolated mesenchymal progenitor stem cell from exfoliated deciduous teeth. These cells were named SHED (stem cells from human exfoliated deciduous teeth) and exhibited high plasticity since they could differentiate into neurons, adipocytes, osteoblasts, and odontoblasts. They differed from DPSCs as SHED were isolated from the pulp tissue of the crown of exfoliated deciduous teeth, and these stem cells did not grow as individual cells instead exhibited the growth in clusters forming several colonies which, after separation, grew as individual fibroblast-like cells. SHED demonstrated higher proliferation rate and a higher number of colony-forming cells compared to DPSCs with early expression of MSC markers (STRO-1 and CD146). They developed multiple cytoplasmic processes and expressed different neuronal and glial cell markers such as nestin when cultured with neurogenic inductive media, suggesting its neural crest origin. The transplantation of SHED into immunocompromised mice showed the formation of dentin-like tissues, which was immune-reactive to dentin-specific sialophospho protein antibody. This regenerated dentin was formed due to odontoblast-like cells indicating the odontogenic differentiation potential of SHED. SHED, unlike DPSCs, cannot be differentiated into osteoblasts or osteocytes but are able to induce the host cells to undergo osteogenic differentiation.8 SHED possess a higher proliferation rate, as well as high odontogenic and osteogenic differentiation potential, which make them distinct from the DPSC and represent the more immature form than DPSC.
PDLSC
The PDL is a specialized tissue located between the cementum and the alveolar bone and has a role of maintaining and supporting the teeth. Its continuous regeneration is thought to involve mesenchymal progenitors arising from the dental follicle. PDL contains different types of cells, which can be differentiated into cementoblasts and osteoblasts.30 The isolated PDLSCs demonstrated their fibroblast-like morphology and exhibited clonogenic nature.31 These cells showed a high rate of proliferation than DPSCs and demonstrated expression of STRO-1, CD146, and a tendon-specific transcription factor.31 It is thus obvious that PDL itself contains progenitors, which can be activated to self-renew and regenerate other tissues such as cementum and alveolar bone.9 PDLSCs possessed multilineage differentiation capabilities and were able to undergo osteogenic, adipogenic, and chondrogenic differentiations when cultured with the suitable inductive medium.9
DFPC
The dental follicle is ectomesenchymal in origin and surrounds the unerupted tooth just like a protective sac. It controls the osteoclastogenesis and osteogenesis processes during the tooth eruption and differentiates into the periodontium.32 DFPCs isolated from follicle of human third molars displayed fibroblast-like morphology and expressed various biomarkers such as Notch1, STRO-1, and nestin.10 The in vitro studies demonstrated the multilineage potential of DFPCs to undergo osteogenic, adipogenic, and neurogenic differentiation.33 These cells can be maintained in culture for at least 15 passages. STRO-1-positive dental follicle stem cells (DFSCs) can differentiate into cementoblasts in vitro and are able to form cementum in vivo.34 DFPC showed their potential to differentiate and express cementoblast markers under stimulation by BMP-2 and BMP-7 and enamel matrix derivatives (EMDs).34 Immortalized dental follicle cells are able to re-create a new PDL after in vivo implantation.35
ABMSC
Successful isolation and culture of human alveolar bone-derived mesenchymal stem cells (hABMSCs) were performed by Matsubara et al.11 The isolated cells exhibit a spindle-shaped fibroblast-like morphology, plastic adherence, and colony formation. These cells express the surface markers CD73, CD90, CD105, and STRO-1 but do not express the hematopoietic markers CD14, CD34, and CD45.11,36,37 ABMSCs can be differentiated into osteoblastic lineages with a high alkaline phosphatase (ALP) expression.11 Studies have shown that treatment of hABMSCs with dichloromethane fraction of Dipsaci Radix38 interferon-induced transmembrane protein 139 nicotine40 low-frequency pulsed electromagnetic fields,41 low-intensity pulsed ultrasound,42 low fluid dynamic shear stress,43 and orbital shear stress44 could enhance osteogenesis in these cells. They have chondrogenic and adipogenic differentiation potentials similar to those of other stem cell populations.45 Bioceramics may provide a good scaffold for ABMSC attachment, proliferation, migration, and differentiation for use in bone tissue engineering applications.46
SCAP
During the tooth development, the dental papilla develops into the dental pulp and subsequently contributes to the development of the root. The dental papilla progresses apically where it is loosely attached to the developing root.12 SCAP have been isolated and their potential to differentiate into odontoblasts was compared to that of the PDLSCs.12 SCAP express the early mesenchymal surface markers especially CD24, which could be a unique marker for their population.12,47 These cells demonstrate their capacity to undergo osteogenic, adipogenic, chondrogenic, and neurogenic differentiation when they are cultured in the suitable inductive media. After transplantation of SCAP into immunocompromised mice in an appropriate carrier matrix, a typical dentin-pulp-like structure was formed due to the presence of odontoblast-like cells.48 SCAP exhibit a higher proliferative rate and appears more effective than PDLSC for tooth formation and are easily accessible since they can be isolated from human third molars.
TGPC
TGPCs are novel stem cell population that were identified in the dental mesenchyme of the third molar tooth germ during the late bell stage.13 They can be expanded and maintained for nearly 60 population doublings, during which they retain their spindle-shaped morphology and high proliferation rate. TGPC express the MSC-associated markers STRO-1 and CDs and demonstrate a tendency for pluripotency-associated gene expression (nanog, oct4, sox2, klf4, and c-myc), indicating a mesenchymal phenotype.49,50 TGPC show a similar multilineage differentiation capacity to that of other dental MSCs, including the ability to differentiate into adipocytes, osteoblasts/odontoblasts, chondrocytes, and neurons. The hydroxyapatite (HA)/TGPC implants showed new bone formation in the presence of osteocytes in the newly formed bone matrix and a cuboid-shaped active osteoblast lining on the matrix surface. TGPCs can differentiate into cells with the morphological, phenotypic, and functional characteristics of hepatocytes in vitro suggesting that TGPC can be used to treat liver diseases.51,52
GMSC
The gingiva is a unique oral tissue overlaying the alveolar ridges and retromolar region that is recognized as a biological mucosal barrier and a distinct component of the oral mucosal immunity. GMSC can be obtained from gingival tissue that are easily accessible from the oral cavity with minimal discomfort.53 GMSC exhibit clonogenicity, self-renewal, and multipotent differentiation capacity, and these cells possess both stem cell-like- and immunomodulatory properties.14,54 GMSCs express CDs and display positive signals for Oct4, Sox2, Nanog, Nestin, SSEA-4, and Stro-1.55,56 Gingival tissues also exhibit scarless wound-healing properties and a regenerative capability with rapid constitution of the tissue architecture. Interestingly, GMSCs display stable phenotype and telomerase activity in long-term cultures and are not tumorigenic.57 Notably, GMSCs have demonstrated the capacity for self-renewal and the formation of connective tissue-like structures in vivo.14 GMSCs maintained the multilineage differentiation potential in the capacity to form mineral, fat, and cartilage-like matrix in vitro, compared with bulk-cultured growth factors. Studies also demonstrated that GMSCs possessed osteogenic potential in vivo after incubation under osteoinductive medium in vitro.56 These properties indicate that the clinical use of GMSCs is an attractive therapeutic option for tissue regeneration and repair.
Therapeutic potential of DSCs
The current understanding of the potential therapeutic applications of DSCs in various systems and diseases is summarized in Table 1.
Table 1.
Therapeutic potential of dental stem cells—systemic diseases.
| No. | Author | System involved | Type of DSC | Outcome |
|---|---|---|---|---|
| 1 | Young et al.58 | Neural system | DPSC | DPSC clones with high-nestin mRNA expression levels were found to successfully differentiate into Map2 and NF-positive neuronal-like cells. |
| 2 | Winderlich et al.59 | Neural system | DPSC | Adult human dental pulp stem cells express vascular endothelial growth factor-a that increases permeability across an in-vitro model of the blood–brain barrier. |
| 3 | Mita et al.60 | Neural system | SHED | Intranasal administration of SHED in mice resulted in substantially improved cognitive function in Alzheimer’s disease through factors that improved neuroprotection, axonal elongation, neurotransmission, the suppression of inflammation, and microglial regulation. |
| 4 | Shimojima et al.61 | Neural system | SHED | EAE, a mouse model of multiple sclerosis, treated with a single injection of SHED-CM significantly improved disease scores, reduced demyelination and axonal injury, and reduced inflammatory cell infiltration and pro-inflammatory cytokine expression in the spinal cord. |
| 5 | Yang et al.62 | Neural system | DPSC | Adult neuronal stem cells may be procured from third molars, and these cultured cells have potential for treatment of stroke-inflicted rats |
| 6 | Inoue et al.63 | Neural system | SHED, DPSC | SHED-CM promoted the migration and differentiation of endogenous neural progenitor cells, induced vasculogenesis, and ameliorated ischemic brain injury after pMCAO, as well as transplantation of DPSC. |
| 7 | Leong et al.64 | Neural system | DPSC | Intracerebral transplantation of human DPSC resulted in enhanced post-stroke functional recovery through non-neural replacement mechanisms. |
| 8 | Yamgata et al.65 | Neural system | SHED | SHED transplantation into the hypoxia ischemic–injured brain resulted in remarkable neurological and pathophysiological recovery in mice. |
| 9 | Mead et al.66 | Optic system | DPSC | Intravitreal transplants of DPSCs promoted significant neurotrophin-mediated retinal ganglion cell survival and axon regeneration after optic nerve injury in mice. |
| 10 | Gomes et al.67 | Optic system | DPSC | Tissue-engineered DPSC sheet was successful for the reconstruction of corneal epithelium in an animal model of total LSCD. |
| 11 | Syed-Picard et al.68 | Optic system | DPSC | DPSCs produced corneal stromal extracellular matrix without affecting corneal transparency or inducing immunological rejection when injected into mouse corneal stroma. |
| 12 | Yamaguchi et al.17 | Cardiac system | SHED | SHED-CM on myocardial injury in a mouse model of I/R decreased apoptosis and inflammatory cytokine levels, such as TNF-α, IL-6, and IL-β, thereby protecting the heart from acute ischemic injury. |
| 13 | Gandia et al.69 | Cardiac system | DPSC | Evidence of cardiac repair was noted by improved cardiac function, increase in the number of vessels, and a reduction in infarct size when DPSC expanded ex vivo was injected in a mice model. |
| 14 | Iohara et al.70 | Angiogenesis | DPSC | In models of mouse hind limb ischemia, local transplantation of this of side population derived from DPSC resulted in successful engraftment and an increase in the blood flow including high density of capillary formation. |
| 15 | Yang et al.71 | Muscular system | DPSC | Clones of DPSC aids in muscle regeneration by expressing human dystrophin and myosin heavy chain. |
| 16 | Tasli et al.72 | Muscular system | TGPC | Pluronic F68 increases the myogenic and neurogenic differentiation of TGPC, useful in functional skeletal and neural tissue engineering applications. |
| 17 | Cho et al.73 | Hepatic system | DPSC | Melatonin and DPSC when grafted in a liver cirrhosis mouse model significantly suppressed liver fibrosis and restored ALT, AST, and ammonia levels were observed |
| 18 | Ishikawa et al.74 | Autoimmune diseases | SHED | Single intravenous administration of serum-free CM from human deciduous dental pulp stem cells (SHED-CM) into anti-CAIA, a mouse model of RA, markedly improved the arthritis symptoms and joint destruction. |
| 19 | Wakayama et al.75 | Respiratory system | SHED | A single intravenous administration of either SHED or SHED-CM attenuated the lung injury and weight loss in BLM-treated mice that exhibit several pathogenic features associated with the human disease acute respiratory distress syndrome and improved their survival rate. |
| 20 | Kanafi et al.76 | Endocrine | SHED | ICC derived from SHED reverse STZ diabetes in mice without immunosuppression. |
| 21 | Kanafi et al.77 | Endocrine | DPSC and SHED | Observed a reversal of hyperglycemia to the normal level in the experimental diabetic mice. |
| 22 | Govindswamy et al.78 | Endocrine | DPSC | Differentiated into pancreatic cell lineage resembling islet-like cell aggregates, glucose-dependently released insulin, and C-peptide. |
| 23 | Annibali et al.79 | Bone | DPSC | Enhanced increase in the bone mineral density critical-sized bone defects in a rat calvarial critical defect model. |
| 24 | Honda et al.80 | Bone | DFSC | Bone formation with evidence of vascular invasion similar to intramembranous ossification. |
DPSC: dental pulp stem cell; SHED: stem cells from human exfoliated deciduous teeth; CM: conditioned medium; EAE: experimental autoimmune encephalomyelitis; pMCAO: permanent middle cerebral artery occlusion; LSCD: limbal stem cell deficiency; I/R: ischemia/reperfusion; TNF: tumor necrosis factor; IL: interleukin; TGPCs: tooth germ progenitor cells; CAIA: collagen type II antibody-induced arthritis; RA: rheumatoid arthritis; BLM: bleomycin; ICC: islet-like cell clusters; STZ: streptozotocin; DFSC: dental follicle stem cells.
The various cell types derived from DSCs are shown in Figure 2.
Figure 2.

The figure enumerates various cell types derived from dental stem cells: (a) cementoblast, (b) adipocyte, (c) odontoblasts, (d) neuronal cells, (e) myoblast, (f) chondrocyte, (g) pulp cells, (h) hepatocyte, (i) endothelial cell, (j) oteoblast, and (k) melanocyte.
Neurological disorders
DPSCs originate from the cranial neural crest and have neural characteristics such as the expression of neurotrophins.81 These diffusible peptides, secreted by neurons and neuron-supporting cells, serve as growth factors for the development, maintenance, repair, and survival of specific neuronal populations: in particular, brain-derived neurotrophic factor (BDNF), nerve growth factor (NGF), and glial-cell-derived neurotrophic factor (GDNF) produced by DPSCs have been shown to have crucial influence over neurons in the central nervous system (CNS), such as motor neurons and dopaminergic neurons of the substantia nigra.82 DPSCs are capable of influencing endogenous recruitment of neural stem cells and generated neurospheres.83 Therefore, DPSCs may represent a promising source in cell therapy for neurological disorders.84
The applications of DPSCS and SHEDs in SCI models have shown that the microenvironment of transplanted stem cells affects their capacity for differentiation; an injured spinal cord contains high levels of pro-inflammatory mediators which may activate the oligodendrocyte-specific differentiation cascade.15,85 Transplantation of hDPSCs into the completely transected adult rat spinal cord resulted in marked recovery of hind-limb locomotor functions. The hDPSCs exhibited neuroregenerative activities.15
The occlusion of a cerebral artery leads to ischemia in a restricted region of the CNS leading to stroke. Transplanting differentiated neural stem cells isolated from dental pulp improved motor disability and reduced infarct volume.86 Therapeutic translation studies of DPSCs to stroke treatment in a rodent cerebral ischemic model showed promising observations.64,87,88 Transplantation of porcine CD31~/CD146~ side population (SP) cells accelerated neovascularization of the ischemic zone and enhanced neuronal regeneration.87 Sugiyama et al.88 transplanted porcine CD31~/CD146~ SP cells migrated to the peri-infarct area, released neurotrophic factors, and promoted migration and differentiation of neural progenitor cells in the subventricular zone. The intracerebral transplantation of hDPSC after focal cerebral ischemia in a rodent model resulted in significant improvement in forelimb sensorimotor function. Improvements to function appeared to be mediated by DPSC-dependent paracrine effects.64 The therapeutic benefit of implanted rat DPSCs into the vitreous body of the eye after a surgically induced optic nerve crush injury promoted neurotrophin-mediated survival of rat ganglion cells and axon regeneration.66
Parkinson’s disease is a neurodegenerative disorder characterized by the progressive death of substantia nigra dopaminergic neurons that results in a regional loss of striatal dopamine (DA) levels. Nesti et al.89 investigated the neuroprotective effects of DPSC against MPP+ and rotenone in an in vitro model of Parkinson’s disease, using an indirect co-culture system with mesencephalic cell cultures. They found that the co-culture with DPSCs significantly attenuated MPP+ or rotenone-induced toxicity probably by the neuroprotection by the soluble factors such as BDNF and NGF, released by DPSC. Hence, DPSC can be viewed as possible candidates for studies on cell-based therapy in neurodegenerative disorders.
DPSC demonstrated that dental pulp-derived cell grafting promoted survival of injured motor neurons in a rat model of SCI.82 DPSC from both rats and humans produced and secreted neurotrophic factors, including NGF, BDNF, and GDNF, and these promoted the survival of sensory and dopaminergic neurons.90 DPSC exerted a neuroprotective effect in in-vitro models of Alzheimer’s and Parkinson’s diseases. The ability to produce and secrete growth factors is of prime importance, since these factors may induce the differentiation of endogenous cell types into those cells required at the place of injury or elicit secretion of other neurotrophic factors from endogenous cells to improve tissue regeneration.84 DPSC showed their applicability also at peripheral nervous system level for nerve injury treatment: they were loaded on poly(dl-lactic-co-glycolic acid) (PLGA) collagen and the scaffold was inserted in a model of facial nerve injury. The system allowed the reconnection of damaged axons (Figure 3).91
Figure 3.

Multilineage differentiation capacity, tissue regeneration, and potential clinical applications of human dental stem cells.
Angiogenesis and vasculogenesis
Vasculogenesis is a potential treatment for ischemic heart disease, and it is an exciting area of research in regenerative medicine. SP cells have been identified in human PDL cells and porcine dental pulp tissues.92 SP cells are highly enriched for stem cell activity.93,94 Iohara et al.70 have shown that SP of dental pulp cells has a property of vasculogenesis. They have isolated a highly vasculogenic subfraction of SP cells from dental pulp which are positive for CD31 and CD146 genes. Thus, they have further suggested that these SP cells are a new source of stem cells which stimulate angiogenesis/vasculogenesis in tissue and can be used in cell-based treatment of ischemic heart diseases.70 Under steady condition, EphB/ephrin-B molecule restricts DPSCs cells for their attachment, migration, and to maintain within their stem cell niche and thus contributing to the localization and maintenance of DPSCs within adult human teeth. Following injury, the mobilization of DPSC to the dentin surfaces may be mediated by EphB/ephrin-B interactions within the adult dental pulp tissue. Therefore, this result suggests a role of EphB/ephrin-B molecule in dental pulp development and regeneration.95
Liver disease
Liver cirrhosis, an irreversible fibrotic change to the liver, can lead to serious consequences such as impaired liver function, portal hypertension, and hepatocellular carcinoma. Liver transplantation is still the only treatment option to prevent more severe clinical course resulting from cirrhosis. Cell-based therapies have drawn attention as novel therapeutic alternatives to whole organ allograft96 SHED are feasible cell source for MSC-based therapy for both pediatric and adult patients with liver dysfunction.97 Stem cells from third molars were differentiated into hepatocytes in cell culture, and in an animal model of liver disease, they prevented liver fibrosis and increased levels of albumin and bilirubin.13,73 Cho et al.73 demonstrated that melatonin promotes hepatic differentiation of hDPSC by modulating the BMP, p38, ERK, and NF-jB pathways. Hence, they concluded that the combined treatment of grafted hDPSCs and melatonin could be a viable approach for the treatment of liver cirrhosis.
Diabetes mellitus
In diabetes, the autoimmune destruction of pancreatic β-cells or decreased sensitivity to insulin develops persistent hyperglycemia. The use of differentiated stem cells or islet transplantation for replenishing the lost insulin-producing cells could be an alternative approach to the conventional insulin-based therapy for diabetes.98 The potential of DPSC to differentiate into pancreatic cell lineage resembling islet-like cell aggregates was reported.78 Carnevale et al.99 reported that hDPSCs under appropriate stimuli express genes related to pancreatic β-cell development and function, including insulin, pancreatic, and duodenal homeobox-1. Kanafi et al.76 demonstrated the transplantation of islet-like cell clusters (ICCs) derived from hDPSC and SHED in diabetic mice. They noticed the reversal of hyperglycemia to the normal level in experimental diabetic mice. These observations suggested the use of dental pulp as an autologous stem cell therapy in diabetic patients.76,100
Regenerative ocular therapy
DSC, as an autologous stem cell source, has been successfully tested in corneal blindness. Since both cornea and DPSC share similar embryonic origin, DPSC differentiated effectively into keratocytes in vitro to generate a tissue-engineered corneal stromal-like tissue construct and to function as keratocytes in vivo without eliciting overt rejection.101 DPSC cultured on aligned nanofiber substrate-generated tissue-engineered, corneal stromal-like constructs, recaptiulated the tightly packed, aligned, parallel fibrillar collagen of native stromal tissue in mouse corneal stroma. These findings demonstrate a potential for the clinical application of DPSC in cellular or tissue engineering therapies for corneal stromal blindness.68 Studies have shown promising results using stem cells from exfoliated deciduous teeth (SHED) for corneal epithelium regeneration.67,102 SHED express markers similar to those of corneal limbal stem cells, and the delivery of cell sheets composed of SHED with and without the addition of amniotic membrane resulted in the regeneration of the corneal epithelium in total limbal stem cell deficiency rabbit models. Huang et al.103 reported that the inhibition of Wnt and bone morphogenetic protein signaling induced retinal cell differentiation from stem cells isolated from the PDL. Adult dental pulp cells isolated from third molars has the capability to differentiate into keratocytes, cells of the corneal stoma. After inducing differentiation in vitro, DPSC expressed molecules’ characteristic of keratocytes, keratocan, and keratan sulfate proteoglycans at both the gene and the protein levels. Intravitreal transplants of DPSC promoted significant neurotrophin-mediated retinal ganglion cell survival and axon regeneration after optic nerve injury in mice.66
Bone tissue engineering
DPSCs showed differentiation profiles similar to those shown during bone differentiation, and this event make them very interesting as a model to study the osteogenesis and the relationship with scaffolds.104,105 The osteogenic differentiation capacity of DPSC has been well demonstrated in vitro and in vivo, with strong ALP results106 and expression of bone-specific markers within newly formed bone.6,106 Enhanced mineralization, protein secretion, and an upregulated osteo-related gene profile resulted from immobilization and, interestingly, immobilization triggered osteogenic differentiation of DPSC without the use of induction factors in the medium.77
A tendency to increase the bone mineral density was observed when DPSCs were implanted in the granular deproteinized bovine bone (GDPB) scaffold.79 Rat DPSCs were utilized in a rat calvarial critical defect model in conjunction with a GDPB or beta tricalcium phosphate (β-TCP) scaffold. GDPB bone scaffolds with DPSC showed the potential to ameliorate bone regeneration process in the reconstruction of the calvarial defects.79 Lucaciu et al.107 used DFSCs from impacted teeth to improve bone regeneration on titanium implants surfaces. They observed spontaneous tendency for osteogenic differentiation and concluded that DFSCs could be used for improving bone regeneration on titanium implant surfaces. Maraldi et al.108 also utilized DPSCs in a rat calvarial critical-sized defect model. hDPSC-seeded collagen sponges showed almost complete bridging of the defect by 8 weeks. Regulation of DPSC differentiation is crucial for clinical use in cell therapies and regenerative medicine. The topographical design of biomaterials may be optimized to achieve this. The interaction of surface topographical parameters upon attachment, morphology, proliferation, and osteogenic differentiation of DPSCs with alterations in pillar topography revealed enhanced mineralization.109 The observations from the in vivo and in vitro studies showed that the addition of DPSC to scaffolds offers a great potential for clinical application in bone reconstruction.110
Therapeutic applications in dentistry
DSCs can be used in the repair of damaged dentin, pulp re-vascularization and regeneration, and for periodontal disorders (Table 2). The combination of DSCs with novel scaffolding materials might enable us to reach the goal of engineering oral tissues in the near future. Understanding the molecular mechanisms of tooth development and repair, utilizing emerging technologies in tissue engineering and biomaterials, and exploiting the potential of DSCs to form complex dental tissues are the foundations of regenerative dentistry. In tissue engineering, whole tooth regeneration, a current dental regenerative process under progress would reduce the difficulties associated with the presently offered dental treatments such as prosthesis, implants, and tooth transplantation.117
Table 2.
Therapeutic potential of dental stem cells in dentistry.
| No. | Author | Type of DSC | Oral site | Outcome |
|---|---|---|---|---|
| 1 | Yu et al.111 | GMSCs | Periodontal regeneration | GMSCs significantly enhanced the regeneration of the damaged periodontal tissue, including the alveolar bone, cementum, and functional PDL. |
| 2 | Li et al.112 | DPSCs | Periodontal regeneration | DPSCs from inflammatory site had a positive effect on regeneration of new bones to repair periodontal defects. |
| 3 | Zhu et al.113 | PDLSC + JBMSC | Periodontal regeneration | DLSCs and JBMSCs regenerated periodontal ligament-like fibers and mineralized matrix on the Ti scaffold surface, both in nude mice ectopic and minipig orthotropic transplantations. |
| 4 | Nagata et al.114 | PDLSC | Periodontal regeneration | PDLSC-CM enhanced periodontal regeneration by suppressing the inflammatory response via TNF-α production in mice model. |
| 5 | Lucaciu et al.107 | DFSC | Osseointegration | Dental follicle stem cells have a spontaneous tendency for osteogenic differentiation and can be used for improving bone regeneration on titanium implant surfaces. |
| 7 | Zhang et al.115 | GMSCs | Oral mucositis | Preconditioned GMSCs enhance mitigation of oral mucositis. |
| 8 | Gao et al.116 | GMSCs | Odontogenic regeneration | GMSCs showed enhanced odontogenic differentiation potential when induced with embryonic tooth germ cell-CM. |
DSC: dental stem cell; GMSC: gingival mesenchymal stem cell; DPSC: dental pulp stem cell; JBMSC: jaw bone mesenchymal stem cell; PDLSC: periodontal ligament stem cell; PDL: periodontal ligament; CM: conditioned medium; TNF: tumor necrosis factor.
Regenerative endodontic therapy
The current treatment modality of the infected root pulp is the removal of necrotic pup tissue and replacement with bioinert cements to obturate the root canals. Although the root canal treatment is effective in combating infection, it does not restore lost dental pulp tissue and the vitality of the tooth. The use of DPSC to regenerate healthy pulp tissue represents a simple and potentially very effective biological treatment. DPSC can be easily expanded in vitro and have been shown to reconstitute a pulp-like tissue ex vivo and in vivo.8,48,118–120
Complete pulp regeneration with neurogenesis and vasculogenesis occurred in an adult canine model with pulpectomy and with autogenous transplantation of pulp CD105 + SP cells with stromal cell-derived factor-1 (SDF-1).121 Another preclinical trial using autologous “mobilized” DPSCs to the pulpectomized teeth of dogs showed regeneration of pulp tissue with no adverse effects and the treated teeth showed recovery of pulp.122 Rosa et al.123 found that SHED survive and differentiate into odontoblasts when transplanted into full-length human root canals with injectable scaffolds. The pulp tissue generated under these experimental conditions contains functional odontoblasts capable of regenerating tubular dentin. Future regenerative endodontic protocol could be a combination of disinfection or debridement of infected root canal systems along with the use of stem cells, scaffolds, and growth factors to permit the revascularization of this pulp. The outcomes of the ongoing studies suggest that it might be feasible to restore viability in a necrotic young permanent tooth by engineering a new dental pulp. The potential impact of such therapy is immense and may allow for the completion and reinforcement of the tooth structure by biological regeneration in near future.124
Dentin regeneration
Regenerative property of the pulp-dentin complex mainly depends on the formation of tertiary dentin, reactionary dentin, and reparative dentin. DPSCs, a unique group of cells with clonogenic ability, high reproductive activity, and multiple differentiation potentials, are extremely crucial element for tertiary dentinogenesis.7,125–127 The DPSCs migrate, proliferate, and differentiate into odontoblasts, which then synthesize matrix to form the tertiary dentin at the damaged sites.128 There are two different approaches implemented in regeneration of dentin by the use of tissue engineering techniques.129,130 The first approach includes a device which can be used as a filling material into a deep cavity of tooth with partial layer of dentin on top of the pulp. In this process, they used some growth factors or molecules that can form reparative dentin.131,132 The second approach is to put scaffold on open pulp along with odontoblast-like cells to grow on it.133 These cells will synthesis reparative dentin. DPSCs have been cultured on a variety of scaffolds to engineer dentin tissues.134,135 However, the signaling pathways underlying the regulation of DPSCs in dentin regeneration remains largely unknown, limiting their effective application in dentin tissue engineering.
Regenerative periodontal therapy
The high prevalence of periodontal disease, the limited regenerative activity exhibited by the PDL, and the critical role of the PDL in maintaining tooth health and function have given more focus to the research on PDL tissue engineering. The challenges in regenerative periodontal therapy lie in the ability to induce the regeneration of a complex apparatus composed of different tissues, such as bone, cementum, and PDL. Despite the recent advancements in periodontal therapy, a complete regeneration of the damaged periodontium is still unattainable. Research findings established that the cells derived from the PDL can differentiate into osteoblasts or cementoblasts to contribute to periodontal regeneration.136 The multipotent PDLSCs were first isolated from the PDL of extracted teeth. These cells express MSC markers, such as STRO-1 and MCAM (CD146), and were shown to be clonogenic and able to differentiate into adipocytes, osteoblasts, and cementoblast-like cells both in vitro and in vivo.9,137,138
The multipotent differentiation properties of PDLSC for generating both hard and soft tissues were further demonstrated by constructing multilayered cell sheets supported by woven polyglycolic acid. Transplanted cell-seeded polyglycolic acid sheets regenerated new bone, cementum, and well-oriented collagen fibers when inserted into root surfaces. Efforts are under way to find an appropriate delivery system for engineering an efficient cell-based therapeutic tool for periodontal tissue regeneration.139 Flores et al.140 have found that fibrin gels carrying several layers of PDL cells can be successfully used as a delivery system for these cells. Collagen sponge scaffolds seeded with PDL cells were successfully tested for the regeneration of periodontal fenestration defects in beagle dogs.141 Another delivery system for PDLSC based on a combination of bovine bone with human dentin was shown to be effective because these stem cells formed a cementum-like complex in subcutaneous dorsum pockets of immunocompromised mice.142 Successful therapies for PDL tissue regeneration will not only facilitate the treatment of periodontal diseases but may also be used to improve current dental implant therapies.
Bioengineered tooth
The biological replacement tooth must include generation of a root and PDL with nerve and blood supplies. The bioengineering of a tooth from non-embryonic cells, one of the cell populations, either epithelial or mesenchymal, must be able to provide the inductive signals to the other.143 Embryonic tooth primordia have been generated in vitro from adult stem cells combined with instructive embryonic oral epithelium, adult epithelial cells combined with embryonic tooth inducing mesenchyme, or by the development of tooth germ models utilizing harvested cells from tooth buds combined with materials.118,144,145
The whole tooth regeneration by tissue engineering currently uses two methods: scaffold method and cell aggregates method. In scaffold method of tooth regeneration, the stem or precursor cells are arranged in proper spatial orientation using a biodegradable polymer membrane or collagen sponge scaffolds to generate an artificial tooth germ.143,146 The cell aggregates method, dental epithelial tissue, and mesenchymal cell pellets are dispersed in a well-controlled culture condition to create an artificial tooth germ. The tooth germ formed in this method mimics a tooth germ of the early inductive stage of tooth development where cell-to-cell and epithelial–mesenchymal interactions are predominant.147–150
Hung et al.151 were able to utilize DPSC to form tooth-like structures in rabbit alveolar sockets but there was no visible tooth eruption in any of the graft sites. In vivo study showed crown-like structures derived from DPSC pellet associated with adult rat apical bud cells with distinct regions of enamel, dentin, pre-dentin, and both ameloblast and odontoblast layers.152 The induced pluripotent stem cells (IPSCs) could generate dental epithelial cells, with embryonic-like properties, that could be recombined with the autologous DPSC and transplanted to form a tooth. The use of IPSCs from DPSCs may indeed provide a future solution to this problem.153 The bioengineered tooth germ primordium, isolated from a bioengineered tooth germ, could be transplanted into a tooth cavity after the extraction of a mandibular incisor and could develop into a tooth with typical spatial orientation of structures, such as enamel, dentin, dental pulp, root, blood vessels, PDL, and alveolar bone. These findings suggest the future possibility of successful generation of whole teeth by transplantation of bioengineered tooth germs into the adult oral environment.145,154
DSC banking
Since the DSCs have various clinical benefits and are applied in various fields of regenerative medicine, the preservation of DSCs for medical applications established the concept of “tooth bank.” Once stem-cell-containing tissues, such as pulp, apical papilla, PDL, follicle, gingiva, or the tooth itself, have been obtained from the patient, they can be preserved for many years to retain their regenerative potential for use in future regenerative therapies. The DSC is stored using either cryopreservation or magnetic freezing.155–157
Although tooth banking is currently not very popular, the trend is gaining acceptance mainly in the developed countries (Table 3). BioEden (Austin, Texas, USA) has international laboratories in the United Kingdom and Thailand. Stem-cell banking companies like Store-A-Tooth (Provia Laboratories, Littleton, Massachusetts, USA) and StemSave (StemSave Inc., New York, USA) are also expanding their presence internationally. The first commercial tooth bank was established as a venture company at Hiroshima University and the company was named as “Three Brackets.”158 Nagoya University (Kyodo, Japan) also came up with a tooth bank in 2007. Taipei Medical University in collaboration with Hiroshima University opened the nation’s first tooth bank in September 2008. The Norwegian Tooth Bank (a collaborative project between the Norwegian Institute of Public Health and the University of Bergen) set up in 2008 is collecting exfoliated primary teeth from 1,00,000 children in Norway. Stemade Biotech introduced the concept of DSC banking in India.
Table 3.
List of licensed dental stem cell banks.
| Country | Tooth bank name | Web address |
|---|---|---|
| Japan | Three brackets (Hiroshima University) | http://www.teethbank.jp/ |
| Teeth Bank Co., Japan | http://www.teethbank.jp/ | |
| Advanced Center for Tissue Engineering, Japan | http://www.acte-group.com/ | |
| USA | BioEDEN, USA | http://www.bioeden.com/ |
| Store-A-Tooth, USA | http://www.store-atooth.com/ | |
| StemSave, USA | http://www.stemsave.com/ | |
| India | Stemade Biotech Pvt., India | http://www.stemade.com/ |
| Norway | MoBaTann: Tooth biobank | http://www.uib.no/en/rg/biomaterial/64723/mobatann-tooth-biobank |
| The Norwegian Tooth Bank, Norway | http://www.fhi.no/morogbarn |
Potential limitations and challenges
Stem cell therapy represents a fascinating new approach for the repair of defective tissues or functions through the transplantation of live cells. However, multiple key parameters need to be optimized through clinical research such as the required stem cell density and availability, as well as appropriate strategies, for their use. Another issue is the availability of the cells over time, that is, DPSCs or exfoliated deciduous tooth stem cells are not available throughout a patient’s lifetime. Even though DSC banking may constitute a potential solution by cryopreserving them for future use, such a possibility is not only time-consuming and costly but limits their use in clinical applications.159 Culture conditions, dose of cell infusion, number of infusions, and route of cell delivery need to be optimized. Appropriate double-blind randomized clinical trials are still need to be performed to confirm the true regenerative power of these stem cells. The immune rejection is a major risk for cell transplantation, so biosecurity is a crucial point for cell therapy, requiring control of cell transformation and a protocol for cellular biobanking. The risk of transmission of bacterial, viral, and fungal or prion pathogens may lead to life-threatening reactions.160 Manufacturing of cell-based medicinal products inevitably does not include terminal sterilization, purification, viral removal, and inactivation. Hence, viral and microbial safety is a pivotal risk factor associated with the use of non-autologous cells including stem cells. Further research and understanding of the stem cell physiology may enhance development of novel and more competent therapeutic approaches and will help in fulfilling the huge impact that stem cell therapy will have for future healthcare.
Conclusion
Stem cell research has expanded at an exponential rate, though its therapeutic applications progress at a slower phase. Stem cell therapy has a promising future in tissue regeneration and the management of disease. It allows the repair of defective tissues or functions through the transplantation of autologus cells. Globally, the advances using stem cells in therapeutic, reconstructive, orthopedic, and cosmetic applications are the future of personalized and regenerative medicine. However, because stem cell technology is still in its infancy, interdisciplinary cooperation is needed to achieve successful clinical applications. DSCs have drawn attention in recent years because of their accessibility, plasticity, and high proliferative ability. Several types of DSCs have been identified, including DPSCs from adult human dental pulp, stem cells from human primary exfoliated deciduous teeth, PDLSCs, and DFSCs from human third molars. The DSCs can undergo self-renewal and have multipotent differentiation ability, but do not have the ethical issues associated with other sources of stem cells. The tissue engineering methodologies combined with an increased understanding of DSC biology will provide powerful tools for a wider spectrum of application of DSC in various therapeutic strategies. We anticipate that the next decade will bring great advances in stem cell and tissue engineering therapies.
Footnotes
Declaration of conflicting interests: The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding: The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant from Marine Biotechnology Program (20150220) funded by the Ministry of Oceans and Fisheries, Republic of Korea.
References
- 1. La Noce M, Mele L, Tirino V, et al. Neural crest stem cell population in craniomaxillofacial development and tissue repair. Eur Cell Mater 2014; 28: 348–357. [DOI] [PubMed] [Google Scholar]
- 2. Smith MM, Fraser GJ, Mitsiadis TA. Dental lamina as source of odontogenic stem cells: evolutionary origins and developmental control of tooth generation in gnathostomes. J Exp Zool B Mol Dev Evol 2009; 312B(4): 260–280. [DOI] [PubMed] [Google Scholar]
- 3. Egusa H, Sonoyama W, Nishimura M, et al. Stem cells in dentistry—part I: stem cell sources. J Prosthodont Res 2012; 56(3): 151–165. [DOI] [PubMed] [Google Scholar]
- 4. Komada Y, Yamane T, Kadota D, et al. Origins and properties of dental, thymic, and bone marrow mesenchymal cells and their stem cells. PLoS ONE 2012; 7(11): e46436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Kawai G, Ohno T, Kawaguchi T, et al. Human dental pulp facilitates bone regeneration in a rat bone defect model. Bone Tissue Regen Insights 2013; 4: 12. [Google Scholar]
- 6. D’Aquino R, Graziano A, Sampaolesi M, et al. Human postnatal dental pulp cells co-differentiate into osteoblasts and endotheliocytes: a pivotal synergy leading to adult bone tissue formation. Cell Death Differ 2007; 14(6): 1162–1171. [DOI] [PubMed] [Google Scholar]
- 7. Gronthos S, Mankani M, Brahim J, et al. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci USA 2000; 97(25): 13625–13630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Miura M, Gronthos S, Zhao M, et al. SHED: stem cells from human exfoliated deciduous teeth. Proc Natl Acad Sci USA 2003; 100(10): 5807–5812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Seo BM, Miura M, Gronthos S, et al. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004; 364(9429): 149–155. [DOI] [PubMed] [Google Scholar]
- 10. Morsczeck C, Gotz W, Schierholz J, et al. Isolation of precursor cells (PCs) from human dental follicle of wisdom teeth. Matrix Biol 2005; 24(2): 155–165. [DOI] [PubMed] [Google Scholar]
- 11. Matsubara T, Suardita K, Ishii M, et al. Alveolar bone marrow as a cell source for regenerative medicine: differences between alveolar and iliac bone marrow stromal cells. J Bone Miner Res 2005; 20(3): 399–409. [DOI] [PubMed] [Google Scholar]
- 12. Sonoyama W, Liu Y, Fang D, et al. Mesenchymal stem cell-mediated functional tooth regeneration in swine. PLoS ONE 2006; 1: e79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Ikeda E, Yagi K, Kojima M, et al. Multipotent cells from the human third molar: feasibility of cell-based therapy for liver disease. Differentiation 2008; 76(5): 495–505. [DOI] [PubMed] [Google Scholar]
- 14. Zhang Q, Shi S, Liu Y, et al. Mesenchymal stem cells derived from human gingiva are capable of immunomodulatory functions and ameliorate inflammation-related tissue destruction in experimental colitis. J Immunol 2009; 183(12): 7787–7798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Sakai K, Yamamoto A, Matsubara K, et al. Human dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat spinal cord by multiple neuro-regenerative mechanisms. J Clin Invest 2012; 122(1): 80–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Matsubara K, Matsushita Y, Sakai K, et al. Secreted ectodomain of sialic acid-binding Ig-like lectin-9 and monocyte chemoattractant protein-1 promote recovery after rat spinal cord injury by altering macrophage polarity. J Neurosci 2015; 35(6): 2452–2464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Yamaguchi S, Shibata R, Yamamoto N, et al. Dental pulp-derived stem cell conditioned medium reduces cardiac injury following ischemia-reperfusion. Sci Rep 2015; 5: 16295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Hilkens P, Gervois P, Fanton Y, et al. Effect of isolation methodology on stem cell properties and multilineage differentiation potential of human dental pulp stem cells. Cell Tissue Res 2013; 353(1): 65–78. [DOI] [PubMed] [Google Scholar]
- 19. Zhang W, Walboomers XF, Shi S, et al. Multilineage differentiation potential of stem cells derived from human dental pulp after cryopreservation. Tissue Eng 2006; 12(10): 2813–2823. [DOI] [PubMed] [Google Scholar]
- 20. Kawashima N. Characterisation of dental pulp stem cells: a new horizon for tissue regeneration? Arch Oral Biol 2012; 57(11): 1439–1458. [DOI] [PubMed] [Google Scholar]
- 21. Karaoz E, Demircan PC, Saglam O, et al. Human dental pulp stem cells demonstrate better neural and epithelial stem cell properties than bone marrow-derived mesenchymal stem cells. Histochem Cell Biol 2011; 136(4): 455–473. [DOI] [PubMed] [Google Scholar]
- 22. Laino G, Carinci F, Graziano A, et al. In vitro bone production using stem cells derived from human dental pulp. J Craniofac Surg 2006; 17(3): 511–515. [DOI] [PubMed] [Google Scholar]
- 23. Dissanayaka WL, Zhan X, Zhang C, et al. Coculture of dental pulp stem cells with endothelial cells enhances osteo-/odontogenic and angiogenic potential in vitro. J Endod 2012; 38(4): 454–463. [DOI] [PubMed] [Google Scholar]
- 24. Spath L, Rotilio V, Alessandrini M, et al. Explant-derived human dental pulp stem cells enhance differentiation and proliferation potentials. J Cell Mol Med 2010; 14(6B): 1635–1644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Pisciotta A, Carnevale G, Meloni S, et al. Human dental pulp stem cells (hDPSCs): isolation, enrichment and comparative differentiation of two sub-populations. BMC Dev Biol 2015; 15: 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Stevens A, Zuliani T, Olejnik C, et al. Human dental pulp stem cells differentiate into neural crest-derived melanocytes and have label-retaining and sphere-forming abilities. Stem Cells Dev 2008; 17(6): 1175–1184. [DOI] [PubMed] [Google Scholar]
- 27. Paino F, Ricci G, De Rosa A, et al. Ecto-mesenchymal stem cells from dental pulp are committed to differentiate into active melanocytes. Eur Cell Mater 2010; 20: 295–305. [DOI] [PubMed] [Google Scholar]
- 28. Davies OG, Cooper PR, Shelton RM, et al. A comparison of the in vitro mineralisation and dentinogenic potential of mesenchymal stem cells derived from adipose tissue, bone marrow and dental pulp. J Bone Miner Metab 2015; 33(4): 371–382. [DOI] [PubMed] [Google Scholar]
- 29. Tecles O, Laurent P, Zygouritsas S, et al. Activation of human dental pulp progenitor/stem cells in response to odontoblast injury. Arch Oral Biol 2005; 50(2): 103–108. [DOI] [PubMed] [Google Scholar]
- 30. McCulloch CA, Melcher AH. Cell density and cell generation in the periodontal ligament of mice. Am J Anat 1983; 167(1): 43–58. [DOI] [PubMed] [Google Scholar]
- 31. Gay IC, Chen S, MacDougall M. Isolation and characterization of multipotent human periodontal ligament stem cells. Orthod Craniofac Res 2007; 10(3): 149–160. [DOI] [PubMed] [Google Scholar]
- 32. Völlner F, Driemel O, Reichert T, et al. Differentiation and characterization of dental follicle precursor cells (PCs). Eur Cell Mater 2007; 14(Suppl. 2): S111. [PubMed] [Google Scholar]
- 33. Morsczeck C, Vollner F, Saugspier M, et al. Comparison of human dental follicle cells (DFCs) and stem cells from human exfoliated deciduous teeth (SHED) after neural differentiation in vitro. Clin Oral Investig 2010; 14(4): 433–440. [DOI] [PubMed] [Google Scholar]
- 34. Kemoun P, Laurencin-Dalicieux S, Rue J, et al. Human dental follicle cells acquire cementoblast features under stimulation by BMP-2/-7 and enamel matrix derivatives (EMD) in vitro. Cell Tissue Res 2007; 329(2): 283–294. [DOI] [PubMed] [Google Scholar]
- 35. Yokoi T, Saito M, Kiyono T, et al. Establishment of immortalized dental follicle cells for generating periodontal ligament in vivo. Cell Tissue Res 2007; 327(2): 301–311. [DOI] [PubMed] [Google Scholar]
- 36. Mason S, Tarle SA, Osibin W, et al. Standardization and safety of alveolar bone-derived stem cell isolation. J Dent Res 2014; 93(1): 55–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Park JC, Kim JC, Kim YT, et al. Acquisition of human alveolar bone-derived stromal cells using minimally irrigated implant osteotomy: in vitro and in vivo evaluations. J Clin Periodontol 2012; 39(5): 495–505. [DOI] [PubMed] [Google Scholar]
- 38. Kim BS, Kim YC, Zadeh H, et al. Effects of the dichloromethane fraction of Dipsaci Radix on the osteoblastic differentiation of human alveolar bone marrow-derived mesenchymal stem cells. Biosci Biotechnol Biochem 2011; 75(1): 13–19. [DOI] [PubMed] [Google Scholar]
- 39. Kim BS, Kim HJ, Kim JS, et al. IFITM1 increases osteogenesis through Runx2 in human alveolar-derived bone marrow stromal cells. Bone 2012; 51(3): 506–514. [DOI] [PubMed] [Google Scholar]
- 40. Kim BS, Kim SJ, Kim HJ, et al. Effects of nicotine on proliferation and osteoblast differentiation in human alveolar bone marrow-derived mesenchymal stem cells. Life Sci 2012; 90(3–4): 109–115. [DOI] [PubMed] [Google Scholar]
- 41. Lim K, Hexiu J, Kim J, et al. Effects of electromagnetic fields on osteogenesis of human alveolar bone-derived mesenchymal stem cells. Biomed Res Int 2013; 2013: 296019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Lim K, Kim J, Seonwoo H, et al. In vitro effects of low-intensity pulsed ultrasound stimulation on the osteogenic differentiation of human alveolar bone-derived mesenchymal stem cells for tooth tissue engineering. Biomed Res Int 2013; 2013: 269724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Lim KT, Kim J, Seonwoo H, et al. Enhanced osteogenesis of human alveolar bone-derived mesenchymal stem cells for tooth tissue engineering using fluid shear stress in a rocking culture method. Tissue Eng Part C Methods 2013; 19(2): 128–145. [DOI] [PubMed] [Google Scholar]
- 44. Lim KT, Hexiu J, Kim J, et al. Synergistic effects of orbital shear stress on in vitro growth and osteogenic differentiation of human alveolar bone-derived mesenchymal stem cells. Biomed Res Int 2014; 2014: 316803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Pekovits K, Kropfl JM, Stelzer I, et al. Human mesenchymal progenitor cells derived from alveolar bone and human bone marrow stromal cells: a comparative study. Histochem Cell Biol 2013; 140(6): 611–621. [DOI] [PubMed] [Google Scholar]
- 46. Kim BS, Kim JS, Chung YS, et al. Growth and osteogenic differentiation of alveolar human bone marrow-derived mesenchymal stem cells on chitosan/hydroxyapatite composite fabric. J Biomed Mater Res A 2013; 101(6): 1550–1558. [DOI] [PubMed] [Google Scholar]
- 47. Ten Cate AR. The development of the periodontium–a largely ectomesenchymally derived unit. Periodontol 1997; 13: 9–19. [DOI] [PubMed] [Google Scholar]
- 48. Huang GT, Yamaza T, Shea LD, et al. Stem/progenitor cell-mediated de novo regeneration of dental pulp with newly deposited continuous layer of dentin in an in vivo model. Tissue Eng Part A 2010; 16(2): 605–615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Yalvac M, Ramazanoglu M, Rizvanov A, et al. Isolation and characterization of stem cells derived from human third molar tooth germs of young adults: implications in neo-vascularization, osteo-, adipo-and neurogenesis. Pharmacogenomics J 2010; 10(2): 105–113. [DOI] [PubMed] [Google Scholar]
- 50. PamukÇUGÜVen E, YalvaÇ ME, Kayahan MB, et al. Human tooth germ stem cell response to calcium-silicate based endodontic cements. J Appl Oral Sci 2013; 21(4): 351–357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Yalvac ME, Yilmaz A, Mercan D, et al. Differentiation and neuro-protective properties of immortalized human tooth germ stem cells. Neurochem Res 2011; 36(12): 2227–2235. [DOI] [PubMed] [Google Scholar]
- 52. Dogan A, Yalvac ME, Sahin F, et al. Differentiation of human stem cells is promoted by amphiphilic pluronic block copolymers. Int J Nanomedicine 2012; 7: 4849–4860. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Mitrano TI, Grob MS, Carrion F, et al. Culture and characterization of mesenchymal stem cells from human gingival tissue. J Periodontol 2010; 81(6): 917–925. [DOI] [PubMed] [Google Scholar]
- 54. Du L, Yang P, Ge S. Isolation and characterization of human gingiva-derived mesenchymal stem cells using limiting dilution method. J Den Sci 2016; 11(3): 304–314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Tang L, Li N, Xie H, et al. Characterization of mesenchymal stem cells from human normal and hyperplastic gingiva. J Cell Physiol 2011; 226(3): 832–842. [DOI] [PubMed] [Google Scholar]
- 56. Wang F, Yu M, Yan X, et al. Gingiva-derived mesenchymal stem cell-mediated therapeutic approach for bone tissue regeneration. Stem Cells Dev 2011; 20(12): 2093–2102. [DOI] [PubMed] [Google Scholar]
- 57. Tomar GB, Srivastava RK, Gupta N, et al. Human gingiva-derived mesenchymal stem cells are superior to bone marrow-derived mesenchymal stem cells for cell therapy in regenerative medicine. Biochem Biophys Res Commun 2010; 393(3): 377–383. [DOI] [PubMed] [Google Scholar]
- 58. Young FI, Telezhkin V, Youde SJ, et al. Clonal heterogeneity in the neuronal and glial differentiation of dental pulp stem/progenitor cells. Stem Cells Int 2016; 2016: 1290561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Winderlich JN, Kremer KL, Koblar SA. Adult human dental pulp stem cells promote blood-brain barrier permeability through vascular endothelial growth factor-a expression. J Cereb Blood Flow Metab 2016; 36(6): 1087–1097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Mita T, Furukawa-Hibi Y, Takeuchi H, et al. Conditioned medium from the stem cells of human dental pulp improves cognitive function in a mouse model of Alzheimer’s disease. Behav Brain Res 2015; 293: 189–197. [DOI] [PubMed] [Google Scholar]
- 61. Shimojima C, Takeuchi H, Jin S, et al. Conditioned medium from the stem cells of human exfoliated deciduous teeth ameliorates experimental autoimmune encephalomyelitis. J Immunol 2016; 196(10): 4164–4171. [DOI] [PubMed] [Google Scholar]
- 62. Yang KL, Chen MF, Liao CH, et al. A simple and efficient method for generating Nurr1-positive neuronal stem cells from human wisdom teeth (tNSC) and the potential of tNSC for stroke therapy. Cytotherapy 2009; 11(5): 606–617. [DOI] [PubMed] [Google Scholar]
- 63. Inoue T, Sugiyama M, Hattori H, et al. Stem cells from human exfoliated deciduous tooth-derived conditioned medium enhance recovery of focal cerebral ischemia in rats. Tissue Eng Part A 2013; 19(1–2): 24–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Leong WK, Henshall TL, Arthur A, et al. Human adult dental pulp stem cells enhance poststroke functional recovery through non-neural replacement mechanisms. Stem Cells Transl Med 2012; 1(3): 177–187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Yamagata M, Yamamoto A, Kako E, et al. Human dental pulp-derived stem cells protect against hypoxic-ischemic brain injury in neonatal mice. Stroke 2013; 44(2): 551–554. [DOI] [PubMed] [Google Scholar]
- 66. Mead B, Logan A, Berry M, et al. Intravitreally transplanted dental pulp stem cells promote neuroprotection and axon regeneration of retinal ganglion cells after optic nerve injury. Invest Ophthalmol Vis Sci 2013; 54(12): 7544–7556. [DOI] [PubMed] [Google Scholar]
- 67. Gomes JA, Geraldes Monteiro B, Melo GB, et al. Corneal reconstruction with tissue-engineered cell sheets composed of human immature dental pulp stem cells. Invest Ophthalmol Vis Sci 2010; 51(3): 1408–1414. [DOI] [PubMed] [Google Scholar]
- 68. Syed-Picard FN, Du Y, Lathrop KL, et al. Dental pulp stem cells: a new cellular resource for corneal stromal regeneration. Stem Cells Transl Med 2015; 4(3): 276–285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Gandia C, Arminan A, Garcia-Verdugo JM, et al. Human dental pulp stem cells improve left ventricular function, induce angiogenesis, and reduce infarct size in rats with acute myocardial infarction. Stem Cells 2008; 26(3): 638–645. [DOI] [PubMed] [Google Scholar]
- 70. Iohara K, Zheng L, Wake H, et al. A novel stem cell source for vasculogenesis in ischemia: subfraction of side population cells from dental pulp. Stem Cells 2008; 26(9): 2408–2418. [DOI] [PubMed] [Google Scholar]
- 71. Yang R, Chen M, Lee CH, et al. Clones of ectopic stem cells in the regeneration of muscle defects in vivo. PLoS ONE 2010; 5(10): e13547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Tasli PN, Dogan A, Demirci S, et al. Myogenic and neurogenic differentiation of human tooth germ stem cells (hTGSCs) are regulated by pluronic block copolymers. Cytotechnology 2016; 68(2): 319–329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73. Cho YA, Noh K, Jue SS, et al. Melatonin promotes hepatic differentiation of human dental pulp stem cells: clinical implications for the prevention of liver fibrosis. J Pineal Res 2015; 58(1): 127–135. [DOI] [PubMed] [Google Scholar]
- 74. Ishikawa J, Takahashi N, Matsumoto T, et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental rheumatoid arthritis. Bone 2016; 83: 210–219. [DOI] [PubMed] [Google Scholar]
- 75. Wakayama H, Hashimoto N, Matsushita Y, et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating acute lung injury in mice. Cytotherapy 2015; 17(8): 1119–1129. [DOI] [PubMed] [Google Scholar]
- 76. Kanafi MM, Rajeshwari YB, Gupta S, et al. Transplantation of islet-like cell clusters derived from human dental pulp stem cells restores normoglycemia in diabetic mice. Cytotherapy 2013; 15(10): 1228–1236. [DOI] [PubMed] [Google Scholar]
- 77. Kanafi MM, Ramesh A, Gupta PK, et al. Dental pulp stem cells immobilized in alginate microspheres for applications in bone tissue engineering. Int Endod J 2014; 47(7): 687–697. [DOI] [PubMed] [Google Scholar]
- 78. Govindasamy V, Ronald VS, Abdullah AN, et al. Differentiation of dental pulp stem cells into islet-like aggregates. J Dent Res 2011; 90(5): 646–652. [DOI] [PubMed] [Google Scholar]
- 79. Annibali S, Bellavia D, Ottolenghi L, et al. Micro-CT and PET analysis of bone regeneration induced by biodegradable scaffolds as carriers for dental pulp stem cells in a rat model of calvarial “critical size” defect: preliminary data. J Biomed Mater Res B Appl Biomater 2014; 102(4): 815–825. [DOI] [PubMed] [Google Scholar]
- 80. Honda MJ, Imaizumi M, Suzuki H, et al. Stem cells isolated from human dental follicles have osteogenic potential. Oral Surg Oral Med Oral Pathol Oral Radiol Endod 2011; 111(6): 700–708. [DOI] [PubMed] [Google Scholar]
- 81. Chai Y, Jiang X, Ito Y, et al. Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development 2000; 127(8): 1671–1679. [DOI] [PubMed] [Google Scholar]
- 82. Nosrat IV, Widenfalk J, Olson L, et al. Dental pulp cells produce neurotrophic factors, interact with trigeminal neurons in vitro, and rescue motoneurons after spinal cord injury. Dev Biol 2001; 238(1): 120–132. [DOI] [PubMed] [Google Scholar]
- 83. Arthur A, Shi S, Zannettino AC, et al. Implanted adult human dental pulp stem cells induce endogenous axon guidance. Stem Cells 2009; 27(9): 2229–2237. [DOI] [PubMed] [Google Scholar]
- 84. Martens W, Bronckaers A, Politis C, et al. Dental stem cells and their promising role in neural regeneration: an update. Clin Oral Investig 2013; 17(9): 1969–1983. [DOI] [PubMed] [Google Scholar]
- 85. Yamamoto A, Sakai K, Matsubara K, et al. Multifaceted neuro-regenerative activities of human dental pulp stem cells for functional recovery after spinal cord injury. Neurosci Res 2014; 78: 16–20. [DOI] [PubMed] [Google Scholar]
- 86. Yang J, Guo L, Liu R, et al. Neuroprotective effects of VEGF administration after focal cerebral ischemia/reperfusion: dose response and time window. Neurochem Int 2012; 60(6): 592–596. [DOI] [PubMed] [Google Scholar]
- 87. Nakashima M, Iohara K, Sugiyama M. Human dental pulp stem cells with highly angiogenic and neurogenic potential for possible use in pulp regeneration. Cytokine Growth Factor Rev 2009; 20(5–6): 435–440. [DOI] [PubMed] [Google Scholar]
- 88. Sugiyama M, Iohara K, Wakita H, et al. Dental pulp-derived CD31(-)/CD146(-) side population stem/progenitor cells enhance recovery of focal cerebral ischemia in rats. Tissue Eng Part A 2011; 17(9–10): 1303–1311. [DOI] [PubMed] [Google Scholar]
- 89. Nesti C, Pardini C, Barachini S, et al. Human dental pulp stem cells protect mouse dopaminergic neurons against MPP+ or rotenone. Brain Res 2011; 1367: 94–102. [DOI] [PubMed] [Google Scholar]
- 90. Nosrat IV, Smith CA, Mullally P, et al. Dental pulp cells provide neurotrophic support for dopaminergic neurons and differentiate into neurons in vitro; implications for tissue engineering and repair in the nervous system. Eur J Neurosci 2004; 19(9): 2388–2398. [DOI] [PubMed] [Google Scholar]
- 91. Sasaki R, Aoki S, Yamato M, et al. PLGA artificial nerve conduits with dental pulp cells promote facial nerve regeneration. J Tissue Eng Regen Med 2011; 5(10): 823–830. [DOI] [PubMed] [Google Scholar]
- 92. Iohara K, Zheng L, Ito M, et al. Side population cells isolated from porcine dental pulp tissue with self-renewal and multipotency for dentinogenesis, chondrogenesis, adipogenesis, and neurogenesis. Stem Cells 2006; 24(11): 2493–2503. [DOI] [PubMed] [Google Scholar]
- 93. Zhou S, Schuetz JD, Bunting KD, et al. The ABC transporter Bcrp1/ABCG2 is expressed in a wide variety of stem cells and is a molecular determinant of the side-population phenotype. Nat Med 2001; 7(9): 1028–1034. [DOI] [PubMed] [Google Scholar]
- 94. Matsuzaki Y, Kinjo K, Mulligan RC, et al. Unexpectedly efficient homing capacity of purified murine hematopoietic stem cells. Immunity 2004; 20(1): 87–93. [DOI] [PubMed] [Google Scholar]
- 95. Stokowski A, Shi S, Sun T, et al. EphB/ephrin-B interaction mediates adult stem cell attachment, spreading, and migration: implications for dental tissue repair. Stem Cells 2007; 25(1): 156–164. [DOI] [PubMed] [Google Scholar]
- 96. Zhang Z, Wang FS. Stem cell therapies for liver failure and cirrhosis. J Hepatol 2013; 59(1): 183–185. [DOI] [PubMed] [Google Scholar]
- 97. Yamaza T, Alatas FS, Yuniartha R, et al. In vivo hepatogenic capacity and therapeutic potential of stem cells from human exfoliated deciduous teeth in liver fibrosis in mice. Stem Cell Res Ther 2015; 6: 171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Bhonde RR, Sheshadri P, Sharma S, et al. Making surrogate β-cells from mesenchymal stromal cells: perspectives and future endeavors. Int J Biochem Cell Biol 2014; 46: 90–102. [DOI] [PubMed] [Google Scholar]
- 99. Carnevale G, Riccio M, Pisciotta A, et al. In vitro differentiation into insulin-producing β-cells of stem cells isolated from human amniotic fluid and dental pulp. Dig Liver Dis 2013; 45(8): 669–676. [DOI] [PubMed] [Google Scholar]
- 100. Guimaraes ET, Cruz Gda S, Almeida TF, et al. Transplantation of stem cells obtained from murine dental pulp improves pancreatic damage, renal function, and painful diabetic neuropathy in diabetic type 1 mouse model. Cell Transplant 2013; 22(12): 2345–2354. [DOI] [PubMed] [Google Scholar]
- 101. Yam GH, Peh GS, Singhal S, et al. Dental stem cells: a future asset of ocular cell therapy. Expert Rev Mol Med 2015; 17: e20. [DOI] [PubMed] [Google Scholar]
- 102. Monteiro BG, Serafim RC, Melo GB, et al. Human immature dental pulp stem cells share key characteristic features with limbal stem cells. Cell Prolif 2009; 42(5): 587–594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Huang L, Liang J, Geng Y, et al. Directing adult human periodontal ligament-derived stem cells to retinal fate. Invest Ophthalmol Vis Sci 2013; 54(6): 3965–3974. [DOI] [PubMed] [Google Scholar]
- 104. Zhang W, Walboomers XF, van Kuppevelt TH, et al. The performance of human dental pulp stem cells on different three-dimensional scaffold materials. Biomaterials 2006; 27(33): 5658–5668. [DOI] [PubMed] [Google Scholar]
- 105. Liu J, Jin T, Chang S, et al. Matrix and TGF-beta-related gene expression during human dental pulp stem cell (DPSC) mineralization. In Vitro Cell Dev Biol Anim 2007; 43(3–4): 120–128. [DOI] [PubMed] [Google Scholar]
- 106. Lindroos B, Maenpaa K, Ylikomi T, et al. Characterisation of human dental stem cells and buccal mucosa fibroblasts. Biochem Biophys Res Commun 2008; 368(2): 329–335. [DOI] [PubMed] [Google Scholar]
- 107. Lucaciu O, Soritau O, Gheban D, et al. Dental follicle stem cells in bone regeneration on titanium implants. BMC Biotechnol 2015; 15: 114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108. Maraldi T, Riccio M, Pisciotta A, et al. Human amniotic fluid-derived and dental pulp-derived stem cells seeded into collagen scaffold repair critical-size bone defects promoting vascularization. Stem Cell Res Ther 2013; 4(3): 53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109. Kolind K, Kraft D, Boggild T, et al. Control of proliferation and osteogenic differentiation of human dental-pulp-derived stem cells by distinct surface structures. Acta Biomater 2014; 10(2): 641–650. [DOI] [PubMed] [Google Scholar]
- 110. Graziano A, d’Aquino R, Laino G, et al. Dental pulp stem cells: a promising tool for bone regeneration. Stem Cell Rev 2008; 4(1): 21–26. [DOI] [PubMed] [Google Scholar]
- 111. Yu X, Ge S, Chen S, et al. Human gingiva-derived mesenchymal stromal cells contribute to periodontal regeneration in beagle dogs. Cells Tissues Organs 2013; 198(6): 428–437. [DOI] [PubMed] [Google Scholar]
- 112. Li Y, Zhao S, Nan X, et al. Repair of human periodontal bone defects by autologous grafting stem cells derived from inflammatory dental pulp tissues. Stem Cell Res Ther 2016; 7(1): 141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Zhu B, Liu W, Liu Y, et al. Jawbone microenvironment promotes periodontium regeneration by regulating the function of periodontal ligament stem cells. Sci Rep 2017; 7: 40088. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 114. Nagata M, Iwasaki K, Akazawa K, et al. Conditioned medium from periodontal ligament stem cells enhances periodontal regeneration. Tissue Eng Part A. Epub ahead of print 27 January 2017. DOI: 10.1089/ten.TEA.2016.0274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. Zhang Q, Nguyen AL, Shi S, et al. Three-dimensional spheroid culture of human gingiva-derived mesenchymal stem cells enhances mitigation of chemotherapy-induced oral mucositis. Stem Cells Dev 2012; 21(6): 937–947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Gao Y, Zhao G, Li D, et al. Isolation and multiple differentiation potential assessment of human gingival mesenchymal stem cells. Int J Mol Sci 2014; 15(11): 20982–20996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Sharpe PT. Dental mesenchymal stem cells. Development 2016; 143(13): 2273–2280. [DOI] [PubMed] [Google Scholar]
- 118. Angelova Volponi A, Kawasaki M, Sharpe PT. Adult human gingival epithelial cells as a source for whole-tooth bioengineering. J Dent Res 2013; 92(4): 329–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Gronthos S, Brahim J, Li W, et al. Stem cell properties of human dental pulp stem cells. J Dent Res 2002; 81(8): 531–535. [DOI] [PubMed] [Google Scholar]
- 120. Cordeiro MM, Dong Z, Kaneko T, et al. Dental pulp tissue engineering with stem cells from exfoliated deciduous teeth. J Endod 2008; 34(8): 962–969. [DOI] [PubMed] [Google Scholar]
- 121. Iohara K, Imabayashi K, Ishizaka R, et al. Complete pulp regeneration after pulpectomy by transplantation of CD105+ stem cells with stromal cell-derived factor-1. Tissue Eng Part A 2011; 17(15–16): 1911–1920. [DOI] [PubMed] [Google Scholar]
- 122. Nakashima M, Iohara K. Mobilized dental pulp stem cells for pulp regeneration: initiation of clinical trial. J Endod 2014; 40(4. Suppl): S26–S32. [DOI] [PubMed] [Google Scholar]
- 123. Rosa V, Zhang Z, Grande RH, et al. Dental pulp tissue engineering in full-length human root canals. J Dent Res 2013; 92(11): 970–975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Potdar PD, Jethmalani YD. Human dental pulp stem cells: applications in future regenerative medicine. World J Stem Cells 2015; 7(5): 839–851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Sloan AJ, Smith AJ. Stem cells and the dental pulp: potential roles in dentine regeneration and repair. Oral Dis 2007; 13(2): 151–157. [DOI] [PubMed] [Google Scholar]
- 126. Patel M, Smith AJ, Sloan AJ, et al. Phenotype and behaviour of dental pulp cells during expansion culture. Arch Oral Biol 2009; 54(10): 898–908. [DOI] [PubMed] [Google Scholar]
- 127. Wang X, He H, Wu X, et al. Promotion of dentin regeneration via CCN3 modulation on Notch and BMP signaling pathways. Biomaterials 2014; 35(9): 2720–2729. [DOI] [PubMed] [Google Scholar]
- 128. Goldberg M, Lacerda-Pinheiro S, Jegat N, et al. The impact of bioactive molecules to stimulate tooth repair and regeneration as part of restorative dentistry. Dent Clin North Am 2006; 50(2): 277.x–298.x. [DOI] [PubMed] [Google Scholar]
- 129. Albuquerque MT, Valera MC, Nakashima M, et al. Tissue-engineering-based strategies for regenerative endodontics. J Dent Res 2014; 93(12): 1222–1231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Wang W, Dang M, Zhang Z, et al. Dentin regeneration by stem cells of apical papilla on injectable nanofibrous microspheres and stimulated by controlled BMP-2 release. Acta Biomater 2016; 36: 63–72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Chiang YC, Chang HH, Wong CC, et al. Nanocrystalline calcium sulfate/hydroxyapatite biphasic compound as a TGF-beta1/VEGF reservoir for vital pulp therapy. Dent Mater 2016; 32(10): 1197–1208. [DOI] [PubMed] [Google Scholar]
- 132. Zhang J, Liu X, Yu W, et al. Effects of human vascular endothelial growth factor on reparative dentin formation. Mol Med Rep 2016; 13(1): 705–712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Song JS, Takimoto K, Jeon M, et al. Decellularized human dental pulp as a scaffold for regenerative endodontics. J Dent Res. Epub ahead of print 1 February 2017. DOI: 10.1177/0022034517693606. [DOI] [PubMed] [Google Scholar]
- 134. El-Backly RM, Massoud AG, El-Badry AM, et al. Regeneration of dentine/pulp-like tissue using a dental pulp stem cell/poly(lactic-co-glycolic) acid scaffold construct in New Zealand white rabbits. Aust Endod J 2008; 34(2): 52–67. [DOI] [PubMed] [Google Scholar]
- 135. Wang J, Ma H, Jin X, et al. The effect of scaffold architecture on odontogenic differentiation of human dental pulp stem cells. Biomaterials 2011; 32(31): 7822–7830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Isaka J, Ohazama A, Kobayashi M, et al. Participation of periodontal ligament cells with regeneration of alveolar bone. J Periodontol 2001; 72(3): 314–323. [DOI] [PubMed] [Google Scholar]
- 137. Gronthos S, Mrozik K, Shi S, et al. Ovine periodontal ligament stem cells: isolation, characterization, and differentiation potential. Calcif Tissue Int 2006; 79(5): 310–317. [DOI] [PubMed] [Google Scholar]
- 138. Shi S, Bartold PM, Miura M, et al. The efficacy of mesenchymal stem cells to regenerate and repair dental structures. Orthod Craniofac Res 2005; 8(3): 191–199. [DOI] [PubMed] [Google Scholar]
- 139. Robey PG, Bianco P. The use of adult stem cells in rebuilding the human face. J Am Dent Assoc 2006; 137(7): 961–972. [DOI] [PubMed] [Google Scholar]
- 140. Flores MG, Hasegawa M, Yamato M, et al. Cementum-periodontal ligament complex regeneration using the cell sheet technique. J Periodontal Res 2008; 43(3): 364–371. [DOI] [PubMed] [Google Scholar]
- 141. Nakahara T, Nakamura T, Kobayashi E, et al. In situ tissue engineering of periodontal tissues by seeding with periodontal ligament-derived cells. Tissue Eng 2004; 10(3–4): 537–544. [DOI] [PubMed] [Google Scholar]
- 142. Yang Z, Jin F, Zhang X, et al. Tissue engineering of cementum/periodontal-ligament complex using a novel three-dimensional pellet cultivation system for human periodontal ligament stem cells. Tissue Eng Part C Methods 2009; 15(4): 571–581. [DOI] [PubMed] [Google Scholar]
- 143. Duailibi MT, Duailibi SE, Young CS, et al. Bioengineered teeth from cultured rat tooth bud cells. J Dent Res 2004; 83(7): 523–528. [DOI] [PubMed] [Google Scholar]
- 144. Zhang W, Abukawa H, Troulis MJ, et al. Tissue engineered hybrid tooth-bone constructs. Methods 2009; 47(2): 122–128. [DOI] [PubMed] [Google Scholar]
- 145. Ohazama A, Modino SA, Miletich I, et al. Stem-cell-based tissue engineering of murine teeth. J Dent Res 2004; 83(7): 518–522. [DOI] [PubMed] [Google Scholar]
- 146. Sumita Y, Honda MJ, Ohara T, et al. Performance of collagen sponge as a 3-D scaffold for tooth-tissue engineering. Biomaterials 2006; 27(17): 3238–3248. [DOI] [PubMed] [Google Scholar]
- 147. Honda MJ, Tsuchiya S, Sumita Y, et al. The sequential seeding of epithelial and mesenchymal cells for tissue-engineered tooth regeneration. Biomaterials 2007; 28(4): 680–689. [DOI] [PubMed] [Google Scholar]
- 148. Yamamoto H, Kim EJ, Cho SW, et al. Analysis of tooth formation by reaggregated dental mesenchyme from mouse embryo. J Electron Microsc 2003; 52(6): 559–566. [DOI] [PubMed] [Google Scholar]
- 149. Hu B, Nadiri A, Kuchler-Bopp S, et al. Tissue engineering of tooth crown, root, and periodontium. Tissue Eng 2006; 12(8): 2069–2075. [DOI] [PubMed] [Google Scholar]
- 150. Gao ZH, Hu L, Liu GL, et al. Bio-Root and Implant-Based Restoration as a Tooth Replacement Alternative. J Dent Res 2016; 95(6): 642–649. [DOI] [PubMed] [Google Scholar]
- 151. Hung CN, Mar K, Chang HC, et al. A comparison between adipose tissue and dental pulp as sources of MSCs for tooth regeneration. Biomaterials 2011; 32(29): 6995–7005. [DOI] [PubMed] [Google Scholar]
- 152. Yu J, Wang Y, Deng Z, et al. Odontogenic capability: bone marrow stromal stem cells versus dental pulp stem cells. Biol Cell 2007; 99(8): 465–474. [DOI] [PubMed] [Google Scholar]
- 153. Zhang W, Vazquez B, Yelick PC. Bioengineered post-natal recombinant tooth bud models. J Tissue Eng Regen Med 2017; 11(3): 658–668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Nakao K, Morita R, Saji Y, et al. The development of a bioengineered organ germ method. Nat Methods 2007; 4(3): 227–230. [DOI] [PubMed] [Google Scholar]
- 155. Oh YH, Che ZM, Hong JC, et al. Cryopreservation of human teeth for future organization of a tooth bank–a preliminary study. Cryobiology 2005; 51(3): 322–329. [DOI] [PubMed] [Google Scholar]
- 156. Gioventu S, Andriolo G, Bonino F, et al. A novel method for banking dental pulp stem cells. Transfus Apher Sci 2012; 47(2): 199–206. [DOI] [PubMed] [Google Scholar]
- 157. Tamaoki N, Takahashi K, Tanaka T, et al. Dental pulp cells for induced pluripotent stem cell banking. J Dent Res 2010; 89(8): 773–778. [DOI] [PubMed] [Google Scholar]
- 158. Kaku M, Kamata H, Kawata T, et al. Cryopreservation of PDL cells by use of program freezer with magnetic field for teeth banking. Den Japan 2007; 43: 82–86. [Google Scholar]
- 159. Inanc B, Elcin YM. Stem cells in tooth tissue regeneration–challenges and limitations. Stem Cell Rev 2011; 7(3): 683–692. [DOI] [PubMed] [Google Scholar]
- 160. Herberts CA, Kwa MS, Hermsen HP. Risk factors in the development of stem cell therapy. J Transl Med 2011; 9: 29. [DOI] [PMC free article] [PubMed] [Google Scholar]