

ABSTRACT
A common pathological event among various neurodegenerative disorders (NDs) is the misfolding and aggregation of different proteins in the brain. This is thought to potentiate aberrant protein-protein interactions that culminate in the disruption of several biological processes and, ultimately, in neuronal cell loss. Although protein aggregates are a common hallmark in several disorders, the molecular pathways leading to their generation remain unclear. The misfolding and aggregation of α-Synuclein (aSyn) is the pathological hallmark of Parkinson disease (PD), the second most common age related ND. It has been postulated that oligomeric species of aSyn, rather than more mature aggregated forms of the protein, are the causative agents of cytotoxicity.
In recent years, we have been investigating the molecular mechanisms underlying the initial steps of aSyn accumulation in living cells. Using an unbiased genome-wide lentiviral RNAi screen we identified trafficking and kinase genes as modulators of aSyn oligomerization, aggregation, and toxicity. Among those, Rab8b, Rab11a, Rab13 and Slp5 were found to promote the clearance of aSyn inclusions and reduce aSyn toxicity. Moreover, we found that endocytic recycling and secretion of aSyn was enhanced upon expression of Rab11a or Rab13 in cells accumulating aSyn inclusions. Altogether, our findings suggest specific trafficking steps may prove beneficial as targets for therapeutic intervention in synucleinopathies, and should be further investigated in other models.
KEYWORDS: α-Synuclein, aggregation, neuronal trafficking, oligomerization, Parkinson disease, Rab GTPases, Slp5
Alpha-Synuclein (aSyn) is a neuronal protein that has been implicated in the etiology of several neurodegenerative disorders (NDs) known as synucleinopathies, that include Parkinson disease (PD), Dementia with Lewy bodies, and multiple system atrophy.1-3 The normal function of aSyn is still not fully understood, but it is thought to be involved in synaptic function and plasticity, cell differentiation, and vesicular trafficking. In disease, mutated or misfolded aSyn establishes aberrant protein-protein interactions, culminating with the disruption of several cellular pathways.4
Evidence from studies in PD patients and animal models suggests that oligomeric species of aSyn are toxic to the neurons, and that large cytoplasmic inclusions may be the result of a protective mechanism to avoid the accumulation of the more toxic species.5-7 In this context, modifying the oligomerization process of aSyn, either by inhibiting the initial interactions that drive the formation of oligomeric species, or by promoting the formation of cytoplasmic protein inclusions that consume oligomeric species, appear as promising strategies. However, promoting inclusion formation requires caution, as aggregates may also disrupt cellular functions, perhaps by physically clogging specific compartments in the cell. Overall, these concepts demand additional investigation.
In our recent study, we used cell-based models of synucleinopathy to investigate the molecular mechanisms underlying aSyn oligomerization. In particular, we screened a collection of shRNAs, targeting 76 genes associated with intracellular transport and 1311 genes involved in signal transduction players, to identify modifiers of aSyn oligomerization, using the bimolecular fluorescence complementation (BiFC) assay as a readout (Fig. 1). Through this approach5,8 we identified 9 genetic modifiers of aSyn oligomerization. Interestingly, the hits we identified were functionally related, and associated with neuronal trafficking processes. We then characterized these hits with respect to their effects on aSyn aggregation, toxicity and protein levels. After this first level of general characterization, we further investigated the mechanism of action of the hits by assessing their effects on aSyn secretion, a central aspect in the spreading of aSyn pathology.
Figure 1.
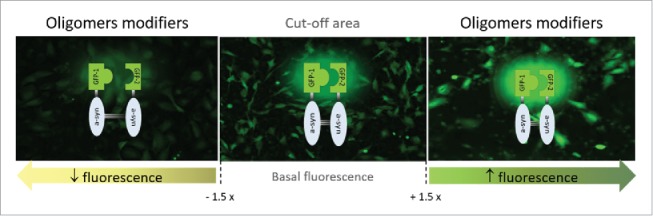
Strategy used for the identification of genetic modifiers of aSyn oligomerization in living cells. A human shRNA library targeting trafficking and phosphotransferase genes was screened using a stable cell line expressing aSyn-BiFC constructs, and basal fluorescent levels were determined (central image). Genes modifying aSyn oligomerization by at least 50% (left and right images) were identified using fluorescence microscopy analysis and were considered for further validation.
aSyn is secreted under physiological conditions, via non-classical exocytosis, in association with exosomes, and possibly via other less conventional mechanisms.9-12 This is also in agreement with the presence of aSyn in cerebrospinal fluid.13,14 Importantly, it remains to be demonstrated if aSyn secretion is detrimental, or if it is part of the normal biology of aSyn. If the latter is true, then secretion of certain forms of the protein need not to be toxic, and may be unrelated to the spreading of pathology. However, it was demonstrated that pathological and aggregated aSyn species can also be secreted, suggesting that aggregated and misfolded aSyn may be the key agent for propagation of aSyn pathology, possibly in a prion-like manner.15-17
Thus, in our study we selected 4 trafficking hits, based on the literature and on their relevance to secretory pathways: RAB8B, RAB11A, RAB13 and SYTL5. SYTL5 encodes for synaptotagmin-like protein 5 (Slp5), an effector of Rab27a. Ras-related protein in brain 8 (Rab8) is associated with actin and microtubule reorganization and with polarized trafficking to dynamic cell surface structures.18 In addition, it is able to reconstitute Golgi morphology in cellular models of PD19 and, as we independently showed, to rescue aSyn induced loss of dopaminergic neurons in Drosophila.20 Rab11a is a ubiquitously expressed protein with predominant localization at the endosomal recycling compartment/recycling endosome (ERC/RE). Strikingly, defects in trafficking from the ERC has been previously implicated in AD, HD and PD.21-23 Rab11a is involved in the process of exocytosis of aSyn via RE.23 Rab13 mediates trafficking between the trans-Golgi network and REs.24 Moreover, it has been associated with neuronal plasticity, neurite outgrowth, cell migration and regulation of tight junctions, all of which are important pathways in normal neuronal biology.25-27 Slp5 mediates the transport of vesicle-Rab27a complex along the cytoskeleton until the plasma membrane is reached, forming a docking/tethering complex that then releases the vesicles.28 For the purpose of our study, we focused on these 4 genes, but we believe that the remaining 5 genes identified in our RNAi-based screen deserve further examination, as they are thought to play relevant roles in neuronal cells (Table 1) and could, therefore, impact on aSyn accumulation.
Table 1.
Neuronal roles and effects of knockdown of the genes identified on aSyn oligomerization/aggregation.
| Effect of knockdown on aSyn |
||||||
|---|---|---|---|---|---|---|
| Oligomerization |
Aggregation |
|||||
| Hit | Role in neuronal processes | Fluorescence intensity | protein levels | Cytotoxicity | Inclusions per cell | Cytotoxicity |
| RAB39B |
|
↑ | ↔ | ↔ | ↑ | ↑ |
| CAMK1 | ↑ | ↑ | ↔ | ↑ | ↔ | |
| DYRK2 |
|
↑ | ↔ | ↔ | ↓ | ↔ |
| CC2D1A |
|
↓ | ↓ | ↔ | ↑ | ↔ |
| CLK4 |
|
↓ | ↓ | ↑ | ↑ | ↔ |
In our validation assays, we investigated the effect of the selected hits on aSyn accumulation, toxicity and secretion, assays that we have previously described.29 We found that silencing Rab8b, Rab11a and Rab13 rescued aSyn-induced toxicity and reduced the accumulation of both oligomeric and aggregated forms of aSyn. Although Slp5 rescues the toxicity associated both with aSyn oligomerization and aggregation, we found it to primarily affect the later stages of aggregation.
We analyzed the effect of overexpressing the hits on aSyn secretion using 2 approaches: by tracking a fluorescently-labeled transferrin (to investigate the endocytic recycling pathway), and by measuring aSyn levels on conditioned media, in the presence of aSyn oligomers or inclusions. Our main finding was that Rab11a and Rab13 increased aSyn release through the recycling endocytic route when aSyn inclusions were present in the cell. When oligomeric aSyn was present no differences in the release of aSyn were observed. Rab8 did not alter the release of aSyn in our models. Slp5 increased the release of aSyn in the oligomerization model, in a manner that was independent of the endocytic recycling pathway (Fig. 2). Given that Slp5 is an effector protein of Rab27a, involved in secretion through exosomes, our finding supports the idea that the release of aSyn can, at least in part, occur via exosomes, as other studies have suggested.12,30-33 Previously, we showed that Rab11 interacts with aSyn in vivo and modulates its secretion through a pathway that does not occur through exosomes.33 In the present study, we add that Rab11a assists the specific trafficking of aSyn oligomers and aggregates through the RE pathway.
Figure 2.

Genes found to modulate the accumulation and secretion of different aggregated species of aSyn. To study the effect of the genes tested on different steps of the process of aSyn aggregation, we used cell models of aSyn oligomerization and aggregation (dashed rectangles). In an unbiased large-scale RNAi screen, we identified 4 trafficking genes that modulate aSyn oligomerization, aggregation, toxicity and secretion.7 Rab8b is involved in polarized vesicular trafficking (endoplasmic reticulum to plasma membrane). Rab11a regulates endocytic recycling pathway and participates specifically in transferrin recycling. Rab13 regulates trans-Golgi network, recycling endosomes and cell/tight junctions. Slp5 is a Rab27a effector protein and plays a role in exosome release. We found that Rab8b, Rab11a and Rab13 overexpression inhibits oligomerization and aggregation of aSyn. Moreover, Rab11a and Rab13 increase aSyn secretion through the recycling endocytic (RE) route only when aSyn inclusions are present in the cell. Slp5 inhibits aSyn aggregation, but not oligomerization, and increases aSyn secretion in the cell model of aSyn oligomerization.
The common effect among the 4 trafficking hits was that Rab8b, Rab11a, Rab13 and Slp5 promoted similar effects in the aSyn aggregation cell model. Upon silencing, they increased the number of inclusions per cell. Conversely, upon overexpression, they reduced the percentage of cells with inclusions in 50%–90% and also reduced aSyn toxicity. Importantly, in cells with inclusions, the trafficking proteins co-localized with aSyn in inclusions. This could either be due to the recruitment of the various proteins into the inclusions, due to the sticky nature of the inclusions, or due to a cellular response in order to try to contain aSyn accumulation.7 This is in agreement with the interaction between aSyn and Rab8 in brain tissue from patients who showed Lewy body pathology but not in control tissue.34 In addition, Rab8 (as well as Rab3a and Rab5) co-immunoprecipitates with aSyn in the extracts from A30P transgenic mice.35 Rab proteins were also previously found to colocalize with aSyn inclusions in yeast cells, further supporting the possibility that aSyn, or other components of inclusions, might sequester Rab GTPases from their normal cellular functions. Also in yeast, deletion of Ypt6p, Ypt7p, and Ypt51p, homologues of mammalian Rab6, Rab7 and Rab5, respectively, that are involved in the endocytic pathway, increase aSyn aggregation.36 Other studies also reported the interaction between a PD-associated mutant version of aSyn (A30P) and Rab3a, Rab5 and Rab8, in transgenic mice.34,35
Trafficking processes govern the physiological homeostasis of neuronal cells in the brain, impacting on cell survival. Vesicular trafficking underlies the function of numerous essential cellular processes such as the export of newly synthesized proteins from the endoplasmic reticulum to the Golgi and to the cell surface, the recycling of membrane receptors, and the transportation and fusion of lysosomal vesicles for the degradation and recycling of cellular components. Thus, dysfunction of key intracellular trafficking processes may impact on normal neuronal function, especially in highly specialized cells such as dopaminergic neurons that appear particularly vulnerable in PD.37 The burden imposed by trafficking processes in dopaminergic neurons might be larger than in other neuron types, as these neurons are estimated to establish 1–2.5 million synapses per neuron in the striatum, and present complex axonal arborizations.38 Indeed, defects in exocytosis, endocytosis, sorting and recycling of endosomal receptors at synaptic transmission sites have already been associated with familial forms of PD. For example, the leucine-rich repeat kinase 2 (LRRK2) protein regulates the release of clathrin-coated endocytic vesicles, neuronal polarity and arborization. LRRK2 also plays roles in chaperone-mediated autophagy.39 Moreover, mutations in ATP6AP2, a gene required for receptor-mediated endocytosis, membrane trafficking and lysosomal degradation, cause X-linked Parkinsonism.40 In addition, mutations in vacuolar protein sorting 35 (VPS35), which mediates retrograde transport of endosomes to trans-Golgi network, cause late-onset PD.41
Our findings suggest that aSyn aggregation can interfere with intracellular trafficking and, in fact, disturbances in this process appear to be a common denominator among different forms of PD forms. Thus, it seems logical to hypothesize that future therapeutic strategies might be designed to target and correct neuronal trafficking defects, as this can relate to (i) autophagy-mediated protein degradation, known to be essential in maintaining the overall cellular proteostasis, and (ii) to the spreading of aSyn pathology in the brain. Additional studies in other cell and animal models will continue to shed light into the role intracellular trafficking plays in PD and other synucleinopathies.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
SAG was supported by Axa Research Fund (www.axa-research.org)) and Fundação para a Ciência e Tecnologia (www.fct.pt), SFRH/BD/79337/2011. TFO was supported by a grant from the Michael J. Fox Foundation (www.michaeljfox.org) and Parkinson's UK. TFO is currently supported by the DFG Center for Nanoscale Microscopy and Molecular Physiology of the Brain.
References
- [1].Maroteaux L, Campanelli JT, Scheller RH. Synuclein: a neuron-specific protein localized to the nucleus and presynaptic nerve terminal. J Neurosci 1988; 8(8):2804-15; PMID:3411354 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Spillantini MG, Crowther RA, Jakes R, Cairns NJ, Lantos PL, Goedert M. Filamentous alpha-synuclein inclusions link multiple system atrophy with Parkinson's disease and dementia with Lewy bodies. Neurosci Lett 1998; 251(3):205-8; PMID:9726379; https://doi.org/ 10.1016/S0304-3940(98)00504-7 [DOI] [PubMed] [Google Scholar]
- [3].Spillantini MG, Crowther RA, Jakes R, Hasegawa M, Goedert M. alpha-Synuclein in filamentous inclusions of Lewy bodies from Parkinson's disease and dementia with lewy bodies. Proc Natl Acad Sci U S A 1998; 95(11):6469-73; PMID:9600990; https://doi.org/ 10.1073/pnas.95.11.6469 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Burre J. The Synaptic Function of alpha-Synuclein. J Parkinsons Dis 2015; 5(4):699-713; PMID:26407041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Outeiro TF, Putcha P, Tetzlaff JE, Spoelgen R, Koker M, Carvalho F, Hyman BT, McLean PJ. Formation of toxic oligomeric alpha-synuclein species in living cells. PLoS One 2008; 3(4):e1867; PMID:18382657; https://doi.org/ 10.1371/journal.pone.0001867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Winner B, Jappelli R, Maji SK, Desplats PA, Boyer L, Aigner S, Hetzer C, Loher T, Vilar M, Campioni S, et al.. In vivo demonstration that alpha-synuclein oligomers are toxic. Proc Natl Acad Sci U S A 2011; 108(10):4194-9; PMID:21325059; https://doi.org/ 10.1073/pnas.1100976108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Goncalves SA, Macedo D, Raquel H, Simões PD, Giorgini F, Ramalho JS, Barral DC, Ferreira Moita L, Outeiro TF. shRNA-Based screen identifies endocytic recycling pathway components that act as genetic modifiers of alpha-synuclein aggregation, secretion and toxicity. PLoS Genet 2016; 12(4):e1005995; PMID:27123591; https://doi.org/ 10.1371/journal.pgen.1005995 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Goncalves SA, Matos JE, Outeiro TF. Zooming into protein oligomerization in neurodegeneration using BiFC. Trends Biochem Sci 2010; 35(11):643-51; PMID:20561791; https://doi.org/ 10.1016/j.tibs.2010.05.007 [DOI] [PubMed] [Google Scholar]
- [9].Lee HJ, Patel S, Lee SJ. Intravesicular localization and exocytosis of alpha-synuclein and its aggregates. J Neurosci 2005; 25(25):6016-24; PMID:15976091; https://doi.org/ 10.1523/JNEUROSCI.0692-05.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Danzer KM, Ruf WP, Putcha P, Joyner D, Hashimoto T, Glabe C, Hyman BT, McLean PJ. Heat-shock protein 70 modulates toxic extracellular alpha-synuclein oligomers and rescues trans-synaptic toxicity. FASEB J 2011; 25(1):326-36; PMID:20876215; https://doi.org/ 10.1096/fj.10-164624 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Sung JY, Park SM, Lee CH, Um JW, Lee HJ, Kim J, Oh YJ, Lee ST, Paik SR, Chung KC. Proteolytic cleavage of extracellular secreted {alpha}-synuclein via matrix metalloproteinases. J Biol Chem 2005; 280(26):25216-24; PMID:15863497; https://doi.org/ 10.1074/jbc.M503341200 [DOI] [PubMed] [Google Scholar]
- [12].Emmanouilidou E, Melachroinou K, Roumeliotis T, Garbis SD, Ntzouni M, Margaritis LH, Stefanis L, Vekrellis K. Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J Neurosci 2010; 30(20):6838-51; PMID:20484626; https://doi.org/ 10.1523/JNEUROSCI.5699-09.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].El-Agnaf OM, Salem SA, Paleologou KE, Cooper LJ, Fullwood NJ, Gibson MJ, Curran MD, Court JA, Mann DM, Ikeda S, et al.. Alpha-synuclein implicated in Parkinson's disease is present in extracellular biological fluids, including human plasma. FASEB J 2003; 17(13):1945-7; PMID:14519670 [DOI] [PubMed] [Google Scholar]
- [14].Lee PH, Lee G, Park HJ, Bang OY, Joo IS, Huh K. The plasma alpha-synuclein levels in patients with Parkinson's disease and multiple system atrophy. J Neural Transm (Vienna) 2006; 113(10):1435-9; PMID:16465458; https://doi.org/ 10.1007/s00702-005-0427-9 [DOI] [PubMed] [Google Scholar]
- [15].Kordower JH, Chu Y, Hauser RA, Freeman TB, Olanow CW. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson's disease. Nat Med 2008; 14(5):504-6; PMID:18391962; https://doi.org/ 10.1038/nm1747 [DOI] [PubMed] [Google Scholar]
- [16].Li JY, Englund E, Holton JL, Soulet D, Hagell P, Lees AJ, Lashley T, Quinn NP, Rehncrona S, Björklund A, et al., Lewy bodies in grafted neurons in subjects with Parkinson's disease suggest host-to-graft disease propagation. Nat Med 2008; 14(5):501-3; PMID:18391963; https://doi.org/ 10.1038/nm1746 [DOI] [PubMed] [Google Scholar]
- [17].Luk KC, Kehm V, Carroll J, Zhang B, O'Brien P, Trojanowski JQ, Lee VM. Pathological alpha-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science 2012; 338(6109):949-53; PMID:23161999; https://doi.org/ 10.1126/science.1227157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Hattula K, Furuhjelm J, Arffman A, Peränen J. A Rab8-specific GDP/GTP exchange factor is involved in actin remodeling and polarized membrane transport. Mol Biol Cell 2002; 13(9):3268-80; PMID:12221131; https://doi.org/ 10.1091/mbc.E02-03-0143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Rendon WO, Martínez-Alonso E, Tomás M, Martínez-Martínez N, Martínez-Menárguez JA. Golgi fragmentation is Rab and SNARE dependent in cellular models of Parkinson's disease. Histochem Cell Biol 2013; 139(5):671-84; PMID:23212845; https://doi.org/ 10.1007/s00418-012-1059-4 [DOI] [PubMed] [Google Scholar]
- [20].Yin G, Lopes da Fonseca T, Eisbach SE, Anduaga AM, Breda C, Orcellet ML, Szegő ÉM, Guerreiro P, Lázaro DF, Braus GH, et al.. alpha-Synuclein interacts with the switch region of Rab8a in a Ser129 phosphorylation-dependent manner. Neurobiol Dis 2014; 70:149-61; PMID:24983211; https://doi.org/ 10.1016/j.nbd.2014.06.018 [DOI] [PubMed] [Google Scholar]
- [21].Li X, Sapp E, Chase K, Comer-Tierney LA, Masso N, Alexander J, Reeves P, Kegel KB, Valencia A, Esteves M, et al.. Disruption of Rab11 activity in a knock-in mouse model of Huntington's disease. Neurobiol Dis 2009; 36(2):374-83; PMID:19699304; https://doi.org/ 10.1016/j.nbd.2009.08.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Greenfield JP, Leung LW, Cai D, Kaasik K, Gross RS, Rodriguez-Boulan E, Greengard P, Xu H. Estrogen lowers Alzheimer beta-amyloid generation by stimulating trans-Golgi network vesicle biogenesis. J Biol Chem 2002; 277(14):12128-36; PMID:11823458; https://doi.org/ 10.1074/jbc.M110009200 [DOI] [PubMed] [Google Scholar]
- [23].Liu J, Zhang JP, Shi M, Quinn T, Bradner J, Beyer R, Chen S, Zhang J. Rab11a and HSP90 regulate recycling of extracellular alpha-synuclein. J Neurosci 2009; 29(5):1480-5; PMID:19193894; https://doi.org/ 10.1523/JNEUROSCI.6202-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Nokes RL, Fields IC, Collins RN, Fölsch H. Rab13 regulates membrane trafficking between TGN and recycling endosomes in polarized epithelial cells. J Cell Biol 2008; 182(5):845-53; PMID:18779367; https://doi.org/ 10.1083/jcb.200802176 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Marzesco AM, Dunia I, Pandjaitan R, Recouvreur M, Dauzonne D, Benedetti EL, Louvard D, Zahraoui A. The small GTPase Rab13 regulates assembly of functional tight junctions in epithelial cells. Mol Biol Cell 2002; 13(6):1819-31; PMID:12058051; https://doi.org/ 10.1091/mbc.02-02-0029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Di Giovanni S, De Biase A, Yakovlev A, Finn T, Beers J, Hoffman EP, Faden AI. In vivo and in vitro characterization of novel neuronal plasticity factors identified following spinal cord injury. J Biol Chem 2005; 280(3):2084-91; PMID:15522871; https://doi.org/ 10.1074/jbc.M411975200 [DOI] [PubMed] [Google Scholar]
- [27].Wu C, Agrawal S, Vasanji A, Drazba J, Sarkaria S, Xie J, Welch CM, Liu M, Anand-Apte B, Horowitz A. Rab13-dependent trafficking of RhoA is required for directional migration and angiogenesis. J Biol Chem 2011; 286(26):23511-20; PMID:21543326; https://doi.org/ 10.1074/jbc.M111.245209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Fukuda M. Rab27 effectors, pleiotropic regulators in secretory pathways. Traffic 2013; 14(9):949-63; PMID:23678941; https://doi.org/ 10.1111/tra.12083 [DOI] [PubMed] [Google Scholar]
- [29].Lazaro DF, Rodrigues EF, Langohr R, Shahpasandzadeh H, Ribeiro T, Guerreiro P, Gerhardt E, Kröhnert K, Klucken J, Pereira MD, et al.. Systematic comparison of the effects of alpha-synuclein mutations on its oligomerization and aggregation. PLoS Genet 2014; 10(11):e1004741; PMID:25393002; https://doi.org/ 10.1371/journal.pgen.1004741 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Danzer KM, Kranich LR, Ruf WP, Cagsal-Getkin O, Winslow AR, Zhu L, Vanderburg CR, McLean PJ. Exosomal cell-to-cell transmission of alpha synuclein oligomers. Mol Neurodegener 2012; 7:42; PMID:22920859; https://doi.org/ 10.1186/1750-1326-7-42 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Alvarez-Erviti L, Seow Y, Schapira AH, Gardiner C, Sargent IL, Wood MJ, Cooper JM. Lysosomal dysfunction increases exosome-mediated alpha-synuclein release and transmission. Neurobiol Dis 2011; 42(3):360-7; PMID:21303699; https://doi.org/ 10.1016/j.nbd.2011.01.029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Ejlerskov P, Rasmussen I, Nielsen TT, Bergström AL, Tohyama Y, Jensen PH, Vilhardt F. Tubulin polymerization-promoting protein (TPPP/p25alpha) promotes unconventional secretion of alpha-synuclein through exophagy by impairing autophagosome-lysosome fusion. J Biol Chem 2013; 288(24):17313-35; PMID:NOT_FOUND; https://doi.org/ 10.1074/jbc.M112.401174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Chutna O, Gonçalves S, Villar-Piqué A, Guerreiro P, Marijanovic Z, Mendes T, Ramalho J, Emmanouilidou E, Ventura S, Klucken J, et al.. The small GTPase Rab11 co-localizes with alpha-synuclein in intracellular inclusions and modulates its aggregation, secretion and toxicity. Hum Mol Genet 2014; 23:6732-45; PMID:25092884 [DOI] [PubMed] [Google Scholar]
- [34].Dalfo E, Barrachina M, Rosa JL, Ambrosio S, Ferrer I. Abnormal alpha-synuclein interactions with rab3a and rabphilin in diffuse Lewy body disease. Neurobiol Dis 2004; 16(1):92-7; PMID:15207266; https://doi.org/ 10.1016/j.nbd.2004.01.001 [DOI] [PubMed] [Google Scholar]
- [35].Dalfo E, Gómez-Isla T, Rosa JL, Nieto Bodelón M, Cuadrado Tejedor M, Barrachina M, Ambrosio S, Ferrer I. Abnormal alpha-synuclein interactions with Rab proteins in alpha-synuclein A30P transgenic mice. J Neuropathol Exp Neurol 2004; 63(4):302-13; PMID:15099020; https://doi.org/ 10.1093/jnen/63.4.302 [DOI] [PubMed] [Google Scholar]
- [36].Soper JH, Kehm V, Burd CG, Bankaitis VA, Lee VM. Aggregation of alpha-synuclein in S. cerevisiae is associated with defects in endosomal trafficking and phospholipid biosynthesis. J Mol Neurosci 2011; 43(3):391-405; PMID:20890676; https://doi.org/ 10.1007/s12031-010-9455-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Matsuda W, Furuta T, Nakamura KC, Hioki H, Fujiyama F, Arai R, Kaneko T. Single nigrostriatal dopaminergic neurons form widely spread and highly dense axonal arborizations in the neostriatum. J Neurosci 2009; 29(2):444-53; PMID:19144844; https://doi.org/ 10.1523/JNEUROSCI.4029-08.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Hunn BH, Cragg SJ, Bolam JP, Spillantini MG, Wade-Martins R. Impaired intracellular trafficking defines early Parkinson's disease. Trends Neurosci 2015; 38(3):178-88; PMID:25639775; https://doi.org/ 10.1016/j.tins.2014.12.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Orenstein SJ, Kuo SH, Tasset I, Arias E, Koga H, Fernandez-Carasa I, Cortes E, Honig LS, Dauer W, Consiglio A, et al.. Interplay of LRRK2 with chaperone-mediated autophagy. Nat Neurosci 2013; 16(4):394-406; PMID:23455607; https://doi.org/ 10.1038/nn.3350 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Korvatska O, Strand NS, Berndt JD, Strovas T, Chen DH, Leverenz JB, Kiianitsa K, Mata IF, Karakoc E, Greenup JL, et al.. Altered splicing of ATP6AP2 causes X-linked parkinsonism with spasticity (XPDS). Hum Mol Genet 2013; 22(16):3259-68; PMID:23595882; https://doi.org/ 10.1093/hmg/ddt180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Vilarino-Guell C, Wider C, Ross OA, Dachsel JC, Kachergus JM, Lincoln SJ, Soto-Ortolaza AI, Cobb SA, Wilhoite GJ, Bacon JA, et al.. VPS35 mutations in Parkinson disease. Am J Hum Genet 2011; 89(1):162-7; PMID:21763482; https://doi.org/ 10.1016/j.ajhg.2011.06.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42]. Mata IF, Jang Y, Kim CH, Hanna DS, Dorschner MO, Samii A, Agarwal P, Roberts JW, Klepitskaya O, Shprecher DR, et al. The RAB39B p.G192R mutation causes X-linked dominant Parkinson's disease. Molecular neurodegeneration 2015; 10:50; https://doi.org/ 10.1186/s13024-015-0045-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43]. Wilson GR, Sim JC, McLean C, Giannandrea M, Galea CA, Riseley JR, Stephenson SE, Fitzpatrick E, Haas SA, Pope K, et al. Mutations in RAB39B cause X-linked intellectual disability and early-onset Parkinson disease with alpha-synuclein pathology. Am J Hum Genet 2014; 95:729–35; https://doi.org/ 10.1016/j.ajhg.2014.10.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44]. Yao Y, Cui X, Al-Ramahi I, Sun X, Li B, Hou J, Difiglia M, Palacino J, Wu ZY, Ma L, et al. A striatal-enriched intronic GPCR modulates huntingtin levels and toxicity. Elife 2015; 4; https://doi.org/ 10.7554/eLife.05449 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [45]. Ageta-Ishihara N, Takemoto-Kimura S, Nonaka M, Adachi-Morishima A, Suzuki K, Kamijo S, Fujii H, Mano T, Blaeser F, Chatila TA, et al. Control of cortical axon elongation by a GABA-driven Ca2+/calmodulin-dependent protein kinase cascade. J Neurosci 2009; 29:13720–9; https://doi.org/ 10.1523/JNEUROSCI.3018-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46]. Martinez J, Moeller I, Erdjument-Bromage H, Tempst P, Lauring B. Parkinson's disease-associated α-synuclein is a calmodulin substrate. J Biol Chem 2003; 278:17379–87; https://doi.org/ 10.1074/jbc.M209020200 [DOI] [PubMed] [Google Scholar]
- [47]. Slepak TI, Salay LD, Lemmon VP, Bixby JL. Dyrk kinases regulate phosphorylation of doublecortin, cytoskeletal organization, and neuronal morphology. Cytoskeleton (Hoboken) 2012; 69:514–27; https://doi.org/ 10.1002/cm.21021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48]. Martinelli N, Hartlieb B, Usami Y, Sabin C, Dordor A, Miguet N, Avilov SV, Ribeiro EA, Jr., Gottlinger H, Weissenhorn W. CC2D1A is a regulator of ESCRT-III CHMP4B. J Mol Biol 2012; 419:75–88; https://doi.org/ 10.1016/j.jmb.2012.02.044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49]. Manzini MC, Xiong L, Shaheen R, Tambunan DE, Di Costanzo S, Mitisalis V, Tischfield DJ, Cinquino A, Ghaziuddin M, Christian M, et al. CC2D1A regulates human intellectual and social function as well as NF-kappaB signaling homeostasis. Cell Rep 2014; 8:647–55; https://doi.org/ 10.1016/j.celrep.2014.06.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50]. Basel-Vanagaite L, Attia R, Yahav M, Ferland RJ, Anteki L, Walsh CA, Olender T, Straussberg R, Magal N, Taub E, et al. The CC2D1A, a member of a new gene family with C2 domains, is involved in autosomal recessive non-syndromic mental retardation. J Med Genet 2006; 43:203–10; https://doi.org/ 10.1136/jmg.2005.035709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51]. Jain P, Karthikeyan C, Moorthy NS, Waiker DK, Jain AK, Trivedi P. Human CDC2-like kinase 1 (CLK1): a novel target for Alzheimer's disease. Curr Drug Targets 2014; 15:539–50; https://doi.org/ 10.2174/1389450115666140226112321 [DOI] [PubMed] [Google Scholar]