

Abstract
Background
Corn and soybean meal (SBM) are two of the most common feed ingredients used in pig feeds. However, a variety of antinutritional factors (ANFs) present in corn and SBM can interfere with the bioavailability of nutrients and have negative health effects on the pigs. In the present study, two-stage fermentation using Bacillus subtilis followed by Enterococcus faecium was carried out to degrade ANFs and improve the nutritional quality of corn and SBM mixed feed. Furthermore, the microbial composition and in vitro nutrient digestibility of inoculated mixed feed were determined and compared those of the uninoculated controls.
Results
During the fermentation process, B. subtilis and lactic acid bacteria (LAB) were the main dominant bacteria in the solid-state fermented inoculated feed, and fermentation produced a large amount of lactic acid (170 mmoL/kg), which resulted in a lower pH (5.0 vs. 6.4) than the fermented uninoculated feed. The amounts of soybean antigenic proteins (β-conglycinin and glycinin) in mixed feed were significantly decreased after first-stage fermentation with B. subtilis. Inoculated mixed feed following two-stage fermentation contained greater concentratioin of crude protein (CP), ash and total phosphorus (P) compared to uninoculated feed, whereas the concentrations of neutral detergent fiber (NDF), hemicellulose and phytate P in fermendted inoculated feed declined (P < 0.05) by 38%, 53%, and 46%, respectively. Notably, the content of trichloroacetic acid soluble protein (TCA-SP), particularly that of small peptides and free amino acids (AA), increased 6.5 fold following two-stage fermentation. There was no difference in the total AA content between fermented inoculated and uninoculated feed. However, aromatic AAs (Phe and Tyr) and Lys in inoculated feed increased, and some polar AAs, including Arg, Asp, and Glu, decreased compared with the uninoculated feed. In vitro dry matter and CP digestibility of inoculated feed improved (P < 0.05) compared with the uninoculated feed.
Conclusions
Our results suggest that two-stage fermentation using B. subtilis followed by E. faecium is an effective approach to improve the quality of corn-soybean meal mixed feed.
Electronic supplementary material
The online version of this article (doi:10.1186/s40104-017-0184-2) contains supplementary material, which is available to authorized users.
Keywords: Antinutritional factors (ANFs), In vitro digestibility, Mixed feed, Nutritional value, Two-stage fermentation
Background
Corn and soybean meal (SBM) are the most common feedstuffs used in pig production in China. However, conventional corn-SBM diets contain a variety of antinutritional factors (ANFs), such as soybean antigenic proteins, phytic acid, oligosaccharides, and other factors that can interfere with the bioavailability of nutrients and have negative health effects in pigs. Soybean antigenic proteins in the diets of weaned pigs provoked a transient hypersensitivity associated with the abnormal morphology of the small intestine [1]. These morphological changes can cause malabsorption syndrome, growth depression, and diarrhea [2, 3]. Phytate in diets may reduce mineral bioavailability and protein digestibility when it is fed to pigs [4]. Furthermore, high levels of soy oligosaccharides, in particular, stachyose and raffinose, can cause intestinal disorder in weaning piglets [5].
Previous research has indicated that fermentation can improve the nutritional quality of animal feed by increasing nutrient bioavailability and reducing ANFs [6]. In China, fermented feed is mainly produced through solid state fermentation (SSF), in which the focus is to decrease the ANFs in single feed ingredients, such as soybean meal [7], cottonseed meal [8], and rapeseed meal [9]. There have been few studies on the use of fermented mixed feed manufactured with SSF. However, the ability of SSF to effectively enhance the nutritional value of mixed feed should be further evaluated. Feeding pigs with fermented liquid feed (FLF) is a useful feeding strategy. Although the growth performance of piglets fed FLF compared with those fed dry feed or non-FLF has been shown to be variable, high lactic acid concentration and low pH in FLF can improve the gastrointestinal health of piglets [10]. In the present study, corn and SBM mixed feed was inoculated with Bacillus subtilis in the first stage of fermentation. The aim of the first stage was to decrease ANFs in mixed feed. Subsequently, Enterococcus faecium was used in the second stage fermentation to produce lactic acid and reduce the pH of the mixed feed.
Methods
Microorganisms and basal substrate
B. subtilis ZJ12-1 was isolated from a traditional fermented food (pickled vegetables). B subtilis ZJ12-1 was selected with a specific screening plate in which the soybean antigenic protein was the sole nitrogen source extracted from SBM. This strain was identified with Gram's dye and conventional biochemical tests including sugar fermentation, Voges-Proskauer, starch hydrolysis, gelatin liquefaction, salt tolerance etc., then confirmed with 16S rDNA sequencing (Additional file 1: Figures S1, S2, S3 and Table S1). E. faecium NCIMB 10415 was obtained from Baolai-leelai Bio-tech Co. Ltd. (Taian, China). E. faecium, which is an authorized feed additive for piglets in the EU and China, was isolated from healthy piglet intestines. Dried corn and SBM sieved through 40 mesh sieves were used in SSF.
Preparation of inoculated mixed feed
A schematic outline showing the manufacturing process of the two-stage fermented feed is provided in Fig. 1. Before fermentation, B. subtilis was cultured in Luria broth (LB) liquid medium at 37 °C for 12 h. E. faecium was cultured in de Man, Rogosa and Sharp (MRS) liquid medium at 37 °C for 16 h. The basal substrate (150 g) included 45% corn, 45% SBM and 10% wheat bran, which was mixed and placed in a 500 mL Erlenmeyer flask covered with cotton plugs and supplemented with sterile water to achieve a 40% moisture content. The wet mixed substrate was inoculated with B. subtilis (8.0 log cfu/g) and fermented at 37 °C for 24 h. After the first-stage of fermentation, the fermented mixture was transferred to a plastic bag equipped with a one-way valve (Rou Duoduo Biotechnology Co., Beijing, China), inoculated with E. faecium (8.0 log cfu/g), and incubated under anaerobic conditions at 37 °C (the second-stage of fermentation). Uninoculated flasks served as controls. In uninoculated samples, all the experimental procedures were the same as those for inoculated feed, except for the addition of sterile medium (LB and MRS) instead of inoculated bacteria. Inoculated and uninoculated samples (control) were set up in triplicate. After 48 h of anaerobic fermentation, wet samples (approximately 100 g) were collected and treated at 105 °C for 30 min to prevent continuous fermentation. Then, the inoculated and uninoculated samples were dried at 65 °C for 24 h, cooled and ground. Treated samples were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), in vitro digestibility, and chemical analysis. Moreover, the remaining inoculated feed continued to ferment under anaerobic conditions for 48 h (total 96 h) at 37 °C. Moisture samples were collected at different inoculation times for microbial, pH, and lactic acid analysis.
Fig. 1.
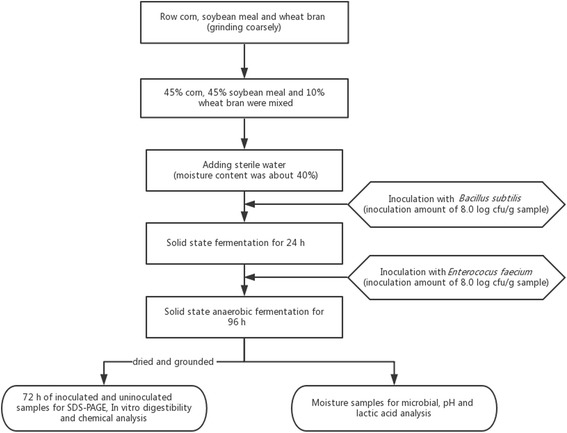
Schematic outline of the manufacturing process for two-stage fermented feed
Microbial determinations
Inoculated feed at different incubation times (0, 24, 48, 72, 96, and 120 h) were taken, 2 g of samples was diluted 1:9 (w/v) with sterile water. The suspension was homogenized in a stomacher blender (Interscience, St. Nom, France) for 2 min. Ten-fold dilutions were prepared in sterile water and 0.1 mL samples were plated on selective media. Lactic acid bacteria (LAB) were measured on MRS agar following anaerobic incubation at 37 °C for 2 d. The number of Enterobacteriaceae was determined on eosin methylene blue agar following aerobic incubation at 37 °C for 1 d. Yeasts and molds were counted on yeast extract peptone dextrose agar with 50 mg/L chloramphenicol (SR0177E, Oxoid LTD, Basingstoke, Hampshire, England) following aerobic incubation at 30 °C for 2 d. Nutrient Broth agar was used to count Bacillus spp. by morphological and biochemical identification after aerobic incubation at 37 °C for 1 d. The biochemical tests were the same as strain identification.
Chemical analysis
Uninoculated and inoculated (24 h with Bacillus subtilis and 48 h with Enterococcus faecium) feeds were analyzed for dry matter (DM), crude protein (CP), ether extract, neutral detergent fiber (NDF), acid detergent fiber (ADF), ash, calcium (Ca) and total phosphorus (P) using the AOAC International guidelines (2005). The trichloroacetic acid soluble protein (TCA-SP) of the sample was determined using the methods described by Ovissipour et al. [11]. The phytic acid content was measured according to the procedures described by Nair and Duvnjak [12]. Phytate-bound P was calculated as 28.2% of phytate. The amino acid profile was analyzed using an automatic amino acid analyzer (L-8900; Hitachi, Tokyo, Japan). Before analysis, the samples were hydrolyzed with 6 mol/L HCl at 110 °C for 24 h. Methionine and cysteine were analyzed as Met sulfone and cysteic acid after cold performic acid oxidation overnight before hydrolysis. The pH values (at different incubation times) were measured using a HI 99163 pH meter (Hanna instruments, Woonsocket, RI, USA) using 2 g of sample mixed with 18 mL of distilled water. The lactic acid content was determined using a lactic acid enzymology assay kit (Nanjing - Jiancheng Bio Co., Nanjing, China) according to the manufacturer's protocol. The contents of glycinin and β-conglycinin in uninoculated and inoculated feed were analyzed using an indirect competitive enzyme-linked immunosorbent assay (ELISA) kit (Longzhoufangke Bio Co., Beijing, China) according to the manufacturer's protocol.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE)
Soluble proteins in fermented uninoculated and inoculated feed were extracted according to the protocol described by Faurobert [13] with some minor modifications. The samples were ground finely to pass through a 60-mesh sieve, and 1.5 mL of Tris–HCl buffer (20 mmol/L, pH 8.3), including 0.1% SDS, 5 mmol/L dithiothreitol and 5 μg/mL protease inhibitor was added to each 0.1 g sample, and then homogenized on ice for 30 min. The homogenized samples were centrifuged at 14,000 × g for 10 min at 4 °C (5804R, Eppendorf, Germany), and the supernatants were transferred to Eppendorf tubes. The protein concentration in each sample was determined using the Bio-Rad Protein Assay Kit (Bio-Rad, USA). Soluble protein was fractionated using an SDS-PAGE system as previously described [14]. The electrophoresis system was based on 4 - 12% polyacrylamide gradient separating gels containing 0.1% SDS in Tris-glycine buffer. Approximately 20 μg of extracted protein sample was loaded into each well and separated at 65 mV for 120 min. A Thermo 26616 page ruler pre-stained protein ladder (10–170 kDa) was used as a size marker. After electrophoresis, the gel was stained using Coomassie Brilliant Blue R-250 (Bio-Rad, USA) for 45 min and de-stained with 7% acetic acid.
In vitro digestibility
An in vitro two-stage enzyme hydrolysis procedure was performed as described by Sakamoto et al. [15], with some modifications. In brief, fermented inoculated feed or uninoculated feed (3 g) was placed in 150 mL Erlenmeyer flasks. Thirty milliliters of 10,000 U/mL pepsin (activity: 3,000 U/mg, Sigma) solution (0.05 mol/L KCl-HCl buffer, pH 2.0) was mixed and incubated at 39 °C at 150 revolutions per min (rpm) for 4 h. The pH was adjusted to 7.0 with 1 mol/L NaOH, and 150 mg trypsin (activity: 250 U/mg, Sigma) was added to each sample, which were then mixed again, and incubated at 39 °C at 150 rpm for 4 h. After the digestion was complete, 5 mL of 20% sulfosalicylic acid was added and the samples were settled for 30 min. The digesta slurry samples were centrifuged at 3,000×g for 15 min, and the supernatants were discarded. The resulting pellets were dried at 105 °C for 4 h and analyzed in subsequent CP and AA assays. In vitro nutrient digestibility (%) = (original nutrient amount – residual nutrient amount) / original nutrient amount × 100%.
Statistical analysis
The data were analyzed by a one-way analysis of variance using the General Linear Models in SAS software (SAS, 1999). A value of 0.05 was used to indicate of a significant difference. The results are expressed as the means and standard deviations.
Results
Microbial composition, pH and lactic acid concentration during SSF
Figure 2a and b shows the microbial composition, pH and lactic acid dynamics during SSF. When the fermentation was prolonged, B. bacillus and LAB were the dominant bacteria present in the solid-state fermented inoculated feed (Fig. 2a). The initial density of B. subtilis was 8.0 log cfu/g. After 24 h of incubation, the density increased to 9.6 log cfu/g, and this level was maintained throughout the fermentation experiment. LAB naturally occurring in the mixed feed was low (<3.0 log cfu/g), and the density of LAB at 24 h was 8.1 log cfu/g after inoculation with E. faecium; this increased to 9.6 log cfu/g at 48 h in inoculated feed. During the subsequent fermentation period, the number of LAB was similar to that of B. subtilis. Notably, there was a proliferation of Enterobacteriaceae (potentially pathogens) during the first-stage of fermentation, which reached a maximum level (8.3 log cfu/g) after 24 h of incubation. However, the number of Enterobacteriaceae gradually decreased as the fermentation time increased, and the final count was below the level of detection (<3.0 log cfu/g). The two-stage process of B. subtilis and E. faecium fermentation had a significant effect on both the pH and lactic acid concentrations of the fermented substrate (Fig. 2b). There was a small increase in pH after incubation with B. subtilis (6.8 in the inoculated feed with 24 h fermentation vs. 6.4 in the raw mixed feed with 0 h fermentation). After inoculation with E. faecium, the pH gradually decreased from 6.8 to 5.0. Almost no change in the lactic acid content was observed during the first-stage of fermentation, and there was a gradual increase in the lactic acid content from 31 to 170 mmol/kg during the second-stage of anaerobic fermentation.
Fig. 2.
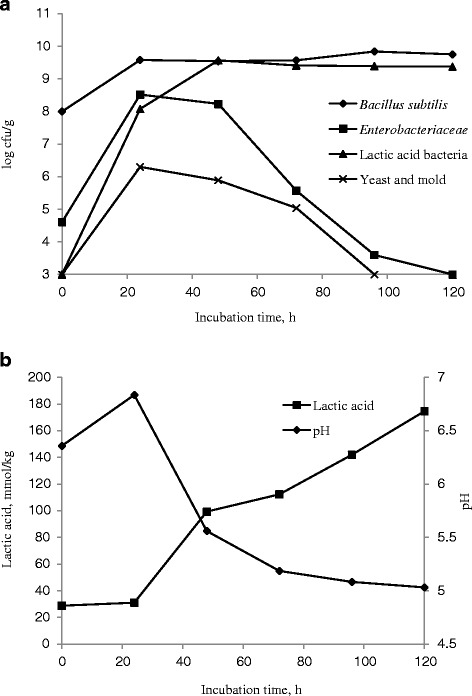
Microbial composition (log cfu/g) a, pH, and lactic acid concentration (mmol/kg) (b) in inoculated feed during solid-state fermentation, on a DM basis
Biodegradation of soybean antigenic proteins of mixed feed after fermentation
Our results showed that the protein profile corresponded to multiple bands in the range of 23–80 kDa in fermented uninoculated feed (Fig. 3). Subunits of soybean antigenic proteins, including β-conglycinin of α, α´ and β, and acidic and basic glycinin in soybean protein were separated. First-stage fermentation with B. subtilis significantly affected the characteristics of proteins in mixed feed. The α and α´ subunits of β-conglycinin and the acidic subunits of glycinin in the mixed feed were almost completely degraded during SSF. In contrast, fermentation increased the number of small peptides (<25 kDa) compared with the fermented uninoculated substrate. However, there was no effect on the protein profile during the second-stage of fermentation with E. faecium compared with the first-stage fermented feed. The contents of soybean antigenic protein in fermented uninoculated and inoculated feed are presented in Table 1. Both β-conglycinin and glycinin contents were significantly decreased after fermentation, and degradation of the antigenic protein had already occurred in the first-stage of fermentation.
Fig. 3.
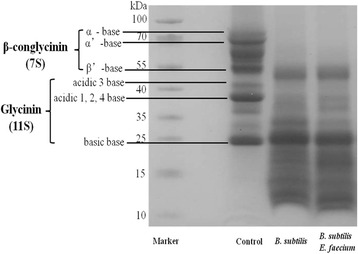
Distribution of peptides in inoculated and uninoculated feed. Marker: protein molecular weight markers (10–100 kDa); control: uninoculated mixed feed (corn 45%, soybean meal 45%, wheat bran 10%) was incubated at 37 °C for 72 h; B. subtilis: the mixed feed was inoculated with B. subtilis and fermented at 37 °C for 24 h; B. subtilis and E. faecium: first-stage fermented (B. subtilis) mixed feed was inoculated with E. faecium and fermented under anaerobic conditions at 37 °C for 48 h
Table 1.
Effect of fermentation on the concentration of soybean antigenic protein, as air-dry basis
| Item | Glycinin | β-conglycinin | ||
|---|---|---|---|---|
| Content, mg/g | Degradationa, % | Content, mg/g | Degradation, % | |
| Raw mixed feed | 63.74 | - | 31.76 | - |
| Uninoculated feedb | 61.02 | - | 32.15 | - |
| B. subtilis c | 7.97 | 86.94 | 6.98 | 78.28 |
| B. subtilis and E. faecium d | 8.47 | 86.12 | 7.12 | 77.53 |
aDegradation rate = soybean antigenic protein content in uninoculated feed – soybean antigenic protein content in inoculated feed) / soybean antigenic protein content in uninoculated feed × 100%
bUninoculated feed: sterile medium was added to mixed feed (45% corn, 45% soybean meal and 10% wheat bran) instead of inoculated bacteria, other experimental procedures were the same as those of inoculated mixed feed
c B. subtilis: mixed feed was inoculated with B. subtilis and fermented at 37 °C for 24 h
d B. subtilis and E. faecium: first-stage fermented (B. subtilis) mixed feed was inoculated with E. faecium and incubated under anaerobic conditions at 37 °C for 48 h
Chemical composition
The analyzed nutrient contents of the fermented uninoculated and inoculated feed after 72 h of incubation are presented in Table 2. Compared with uninoculated feed, the inoculated feed contained more CP, ash, and total P, whereas the concentrations of NDF, hemicellulose and phytate P declined (P < 0.05) by 39%, 53%, and 46%, respectively. Notably, the content of TCA-SP (<10 kDa) in uninoculated feed was 1.18%, which was increased 6.5-fold in inoculated feed. Inoculating with B. subtilis and E. faecium affected the AA composition patterns of mixed feed; content of some polar amino acids (Arg, Asp, and Glu) decreased, and aromatic amino acids (Phe and Tyr) and Lys increased. Compared to uninoculated feed, the total AA of inoculated feed increased by 7%; however, the difference was not significant.
Table 2.
Analyzed nutrient composition of fermented inoculated and uninoculated feed, as air-dry basis1
| Item | Inoculated feed | Uninoculated feed2 | AA compositon | Inoculated feed | Uninoculated feed |
|---|---|---|---|---|---|
| DM,% | 88.06 ± 1.02 | 89.09 ± 1.67 | Indispensable AA, % | ||
| CP,% | 27.61 ± 2.73a | 24.03 ± 1.93b | Arg | 1.01 ± 0.15b | 1.17 ± 0.19a |
| TCA-SP,% | 8.85 ± 1.19a | 1.18 ± 0.12b | His | 0.58 ± 0.08 | 0.52 ± 0.11 |
| Fat,% | 3.37 ± 0.65 | 3.67 ± 0.73 | Ile | 0.78 ± 0.13 | 0.78 ± 0.16 |
| NDF,% | 8.33 ± 0.95b | 13.64 ± 0.99a | Leu | 1.50 ± 0.18 | 1.46 ± 0.23 |
| ADF,% | 3.58 ± 0.40 | 3.49 ± 0.76 | Lys | 1.17 ± 0.08a | 0.99 ± 0.10b |
| Hemicellulose3,% | 4.75 ± 0.87b | 10.15 ± 0.56a | Met | 0.26 ± 0.05 | 0.23 ± 0.07 |
| Ash,% | 4.71 ± 0.51a | 3.77 ± 0.38b | Phe | 1.78 ± 0.26a | 0.86 ± 0.13b |
| Ca,% | 0.18 ± 0.03 | 0.17 ± 0.02 | Thr | 0.79 ± 0.12 | 0.75 ± 0.09 |
| Total P,% | 0.55 ± 0.05 | 0.49 ± 0.07 | Val | 1.06 ± 0.12 | 1.04 ± 0.17 |
| Phytate P,% | 0.21 ± 0.04b | 0.39 ± 0.04a | Dispensable AA,% | ||
| Asp | 1.68 ± 0.10b | 1.92 ± 0.17a | |||
| Ser | 0.79 ± 0.14 | 0.75 ± 0.16 | |||
| Glu | 3.23 ± 0.58 | 3.49 ± 0.44 | |||
| Gly | 0.85 ± 0.18 | 0.80 ± 0.13 | |||
| Ala | 0.98 ± 0.16 | 0.94 ± 0.10 | |||
| Cys | 0.48 ± 0.05a | 0.38 ± 0.06b | |||
| Tyr | 1.40 ± 0.21a | 0.67 ± 0.09b | |||
| Pro | 1.09 ± 0.17 | 1.17 ± 0.21 | |||
| Total AA | 19.56 ± 2.33 | 18.12 ± 2.47 | |||
1Values are means of three replicates per treatment. Means in a row without common superscript differ significantly (P < 0.05)
2Uninoculated feed: sterile medium was added to mixed feed (45% corn, 45% soybean meal and 10% wheat bran) instead of inoculated bacteria, other experimental procedures were the same as those of inoculated feed
3Hemicellulose = NDF-ADF
In vitro amino acid digestibility of the fermented samples
The results of in vitro AA digestibility of fermented inoculated feed with two-stage fermentation are presented in Table 3. In vitro CP and DM digestibility of inoculated feed were improved (P < 0.05) by 8% and 11%, respectively, compared with uninoculated substrate. In addition, the in vitro digestibility of 11 amino acids, including six essential amino acids (His, Ile, Leu, Met, Phe and Val), improved greatly (P < 0.05). Notably, the in vitro digestibility of three amino acids (His, Phe and Cys) increased by more than 10%.
Table 3.
In vitro CP and AA digestibility (%) of fermented inoculated feed and uninoculated1
| Item | Inoculated feed | Uninoculated feed2 |
|---|---|---|
| DM,% | 70.60 ± 2.87a | 59.33 ± 2.32b |
| CP,% | 86.28 ± 2.23a | 78.36 ± 2.04b |
| Indispensable AA,% | ||
| Arg | 82.50 ± 4.65 | 82.72 ± 3.87 |
| His | 84.91 ± 3.70a | 74.85 ± 3.46b |
| Ile | 80.49 ± 3.42a | 75.62 ± 2.44b |
| Leu | 77.30 ± 3.04a | 69.71 ± 2.81b |
| Lys | 84.59 ± 3.91 | 81.44 ± 3.60 |
| Met | 85.30 ± 3.96a | 70.31 ± 2.74b |
| Phe | 81.99 ± 4.25a | 65.64 ± 3.63b |
| Thr | 78.73 ± 4.12 | 75.03 ± 3.83 |
| Val | 80.74 ± 3.77a | 74.49 ± 3.48b |
| Mean | 81.29 ± 4.09a | 74.80 ± 3.21c |
| Dispensable AA,% | ||
| Asp | 83.14 ± 5.32 | 78.54 ± 4.97 |
| Ser | 77.86 ± 3.13 | 74.29 ± 3.74 |
| Glu | 85.13 ± 2.47a | 80.25 ± 3.02b |
| Gly | 80.78 ± 4.21 | 76.70 ± 4.08 |
| Ala | 84.53 ± 3.38a | 75.51 ± 3.66b |
| Cys | 79.74 ± 3.64a | 67.90 ± 3.87b |
| Tyr | 81.86 ± 3.43b | 72.28 ± 3.10c |
| Pro | 79.31 ± 4.28 | 75.41 ± 3.94 |
| Mean | 82.72 ± 3.11a | 77.16 ± 3.04b |
| Total AA,% | 82.15 ± 3.43a | 76.07 ± 3.35c |
1Values are means of three replicates per treatment. Means in a row without common superscript differ significantly (P < 0.05)
2Uninoculated feed: sterile medium was added to mixed feed (45% corn, 45% soybean meal and 10% wheat bran) instead of inoculated bacteria, other experimental procedures were the same as those of inoculated feed
Discussion
Interest in the fermentation of feed for improving the health of pigs increased dramatically after the European Union banned the use of antibiotics as antimicrobial growth promoters for swine [16, 17]. FLF usually contains >9 log cfu/g of LAB and a high concentration of lactic acid (>150 mmol/L), which can prevent the proliferation of spoilage organisms in the gastrointestinal tracts (GIT) of pigs, such as coliforms and Salmonella [18]. Additional advantages of feeding FLF include an increase in nutrient digestibility [19], improved intestinal morphology [20], and a reduction in dust levels in swine barns [21]. Feeding FLF has been shown to improve the performance of piglets and growing-finishing pigs [22], although the results showed high variation. In the present study, changes in the microbial composition with incubation time were determined. During the fermentation process, B. subtilis and LAB were the main dominant bacteria in the solid-state fermented feed. The final count of LAB (9.4 log cfu/g) in the present study was similar to that achieved with FLF. The B. subtilis count was >9.0 log cfu/g after the first stage of fermentation. However, the source of determined bacteria (exogenous addition or naturally occurring in the feed) is not clear. Notably, a proliferation of Enterobacteriaceae, mainly coliforms (potentially pathogens), also occurred at this stage. This result was consistent with that during the initial fermentation of FLF reported by Canibe and Jensen [10]. An increase in feed pH from 6.4 to 6.8 was observed during first-stage fermentation. This increase may be the result of fermentation with B. subtilis and other microbes, which introduce some new metabolites or changes in the chemical composition of the substrate. However, additional research is needed to determine the specific reasons for this increase. During the second stage of fermentation, a decrease in pH from 6.8 to 5.0 was most likely the result of increased lactic acid production. Several previously published studies showed that FLF had a pH of 3.8–4.5 [23–25], which was lower than that obtained in the present study. One explanation for the lower pH with FLF may be the difference in the composition of raw materials, since different ingredients have different buffer capacities. The pH may have decreased more rapidly when only the cereals were fermented because cereals have a lower buffering capacity than compound feed [10]. In the present study, the number of Enterobacteriaceae gradually decreased as the anaerobic incubation time increased, and the final count was reduced to levels below detection limits (<3.0 log cfu/g). Coliform was reduced mainly due to the low pH and increased lactic acid in fermented inoculated feed. Feeding fermented feed with low pH and high concentration of lactic acid can prevent the proliferation of pathogens (e.g. Enterobacteriaceae) along the GIT of piglets [18, 23].
Corn-SBM diet is the most commonly used feed for animal production in China. Corn, as the main energy feed, usually accounts for approximately 60% of the animals’ diet. Soybean meal is the most important plant protein feed for monogastric animals. Soybean antigenic proteins in the diet of weaned pigs, particularly glycinin and β-conglycinin, promote transient hypersensitivity, which may lead to morphological changes in the small intestine, including villi atrophy and crypt hyperplasia [1]. The use of solid-state fermentation to enhance the nutritional characteristics of raw plant materials has been proposed to improve the use of these materials in animal feeds. Several recent studies have shown that soy antigenic proteins could be degraded during fermentation [26, 27]; consequently, immunoreactivity and allergic reactions caused by soy products were reduced in human and animals [28, 29]. In the present study, the first stage of fermentation with B. subtilis significantly affected the characteristics of proteins in corn-SBM mixed feed. The α and α’ subunits of β-conglycinin and acidic subunits of glycinin of compound feed were almost completely degraded. This result was consistent with previous reports [14, 27]. ELISA analysis also showed that the contents of β-conglycinin and glycinin in mixed feed were degraded by 78 and 88%, respectively, after the first stage of fermentation. However, no degradation of soybean antigenic protein occurred during the second stage of anaerobic fermentation. This indicated that the decreased amount of antigenic protein in mixed feed may have been due to the hydrolysis of the proteolytic enzyme secreted by B. subtilis during the first stage of fermentation. B. subtilis secretes many proteolytic enzymes during fermentation, including aminopeptidases, serine endopeptidases, and metalloproteinases [30]. Recently, we showed that the enzyme activity of neutral protease was significantly increased during B. subtilis fermentation (unpublished data).
Inoculated mixed feed with two-stage fermentation contained greater concentrations of CP, ash, and total P than uninoculated feed, which was consistent with previous results for fermented compound feed [31], soybean meal [14, 32], rapeseed meal [33, 34] and cottonseed meal [35]. The loss of dry matter (mainly carbohydrates) in the fermented substrate contributing to a relative increase in the concentration of these nutrients was probably the reason for these results. Rozan et al. showed that the loss of dry matter during fermentation may explain the increase in total protein [36]. Because the CP increased after fermentation, the total AA content of inoculated feed would also be increased. However, the composition of amino acids differed between uninoculated and inoculated feed, and some polar amino acids in inoculated feed, such as Asp and Glu were decreased, whereas Tyr, Phe, and Lys increased compared with uninoculated feed. These results were similar to those of a previous study of fermented soybeans [14]. The total protein content in B. bacillus is 62.93%. Among them, the most abundant essential AAs are Leu, Lys, Phe, and Val [37], which are more than two times as much as the uninoculated feed. We hypothesized that part of the vegetable protein in mixed feed was used to synthesize microbial protein during SSF. However, the specific mechanism accounting for differences in amino acid composition between fermented inoculated and uninoculated feeds requires further study. Furthermore, inoculated feed exhibited an increase in TCA-SP (8.8%) compared to uninoculated feed (1.2%). An increase in TCA-SP was probably due to the hydrolysis of macromolecular proteins (especially antigenic proteins). TCA-SP was assumed to consist of small molecular peptides (2–20 amino acid residues) and free AAs [38]. Di- and tripeptides in TCA-SP can be directly absorbed in the animal gut system, and transport of AA in the form of small peptides was faster than their constituent AAs in the free form [39]. Furthermore, a decrease in NDF, hemicellulose, and phytic acid in FMF was observed after fermentation. This is might be due to the production of relevant enzymes by micro-organism, such as non-starch polysaccharide (NSP)-degrading enzymes and phytase, which caused the breakdown of these antinutritional substrates. Fermentation of rapeseed meal with Aspergillus niger was studied by Shi et al. [33], who showed that the levels of NDF, glucosinolates, and phytic acid in rapeseed meal declined by 14.45%, 43.72%, and 86.08%, respectively, after SSF. Pig feeding showed that fermented rapeseed meal had a greater P and energy digestibility than rapeseed meal [34]. Therefore, lower NDF and phytic acid indicate that inoculated mixed feed with two-stage fermentation may have higher nutrient digestibility compared with non-fermented mixed feed. The results of in vitro digestibility showed that two-stage fermentation with B. subtilis and E. faecium may improve the nutritional value of corn-SBM mixed feed.
Conclusions
Two-stage fermentation with B. subtilis followed by E. faecium effectively reduced ANFs (soy antigenic protein, NDF, and phytic acid) in corn-SBM mixed feed and increased the TCA-SP and CP content. Furthermore, the high lactic acid concentration and low pH in fermented inoculated feed inhibited the proliferation of Enterobacteriaceae. The results of in vitro digestion indicated that inoculated feed subjected to two-stage fermentation had higher DM and CP digestibility than fermented uninoculated feed. Therefore, two-stage fermented feed may be used as a novel feed ingredient in animal diets, especially for piglets. Our results suggest that the two-stage SSF method provides an effective approach for improving the quality of corn-soybean mixed feed.
Acknowledgments
The authors thank the Specialized Research Fund for the China Pig Modern Industrial Technology System Grant (CARS-36), the China and Zhejiang province Postdoctoral Science Foundation (518000-X91604, 518000-X81601) for supporting this study.
Funding
The design of the study and collection, analysis, and interpretation of data were supported by a China Pig Modern Industrial Technology System Grant (CARS-36), the China and Zhejiang province Postdoctoral Science Foundation (518000-X91604, 518000-X81601).
Availability of data and materials
The datasets generated and analyzed during the current study are not publicly available. Please contact the authors for data requests.
Author’s contributions
YZW and CYS conceived and designed the experiment. CYS and YZ carried out the experiment, including the solid-state fermentation, chemical analysis, and determination of in vitro digestibility. CYS analyzed the data and wrote the manuscript. ZQL verified the validity and checked the results. All authors read and approved the final version of this manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- AA
Amino acid
- ANFs
Antinutritional factors
- Ca
Calcium
- CP
Crude protein
- FLF
Fermented liquid feed
- LAB
Lactic acid bacteria
- NDF
Neutral detergent fiber
- NSP
Non-starch polysaccharide
- P
Phosphorus
- SBM
Soybean meal
- SDS-PAGE
Sodium dodecyl sulfate – polyacrylamide gel electrophoresis
- SSF
Solid state fermentation
- TCA-SP
Trichloroacetic acid soluble protein
Additional file
Strain identification information. (DOCX 1027 kb)
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s40104-017-0184-2) contains supplementary material, which is available to authorized users.
Contributor Information
Changyou Shi, Email: changyou523-@163.com.
Yu Zhang, Email: 494941469@qq.com.
Zeqing Lu, Email: 382609564@qq.com.
Yizhen Wang, Email: yzwang321@zju.edu.cn.
References
- 1.Li DF, Nelssen JL, Reddy PG, Blecha F, Hancock JD, Allee GL, et al. Transient hypersensitivity to soybean meal in the early-weaned pig. J Anim Sci. 1990;68:1790–1799. doi: 10.2527/1990.6861790x. [DOI] [PubMed] [Google Scholar]
- 2.Sun P, Li DF, Dong B, Qiao SY, Ma X. Effects of soybean glycinin on performance and immune function in early weaned pigs. Arch Anim Nutr. 2008;62:313–321. doi: 10.1080/17450390802066419. [DOI] [PubMed] [Google Scholar]
- 3.Sun P, Li DF, Li ZJ, Dong B, Wang FL. Effects of glycinin on IgE-mediated increase of mast cell numbers and histamine release in the small intestine. J Nutr Biochem. 2008;19:627–33. doi: 10.1016/j.jnutbio.2007.08.007. [DOI] [PubMed] [Google Scholar]
- 4.Guggenbuhl P, Simões-Nunes C. Effects of two phytases on the ileal apparent digestibility of minerals and amino acids in ileo-rectal anastomosed pigs fed on a maize rapeseed meal diet. Livest Sci. 2007;109:261–63. doi: 10.1016/j.livsci.2007.01.110. [DOI] [Google Scholar]
- 5.Zhang LY, Li DF, Qiao SY, Wang JT, Bai L, Wang ZY, et al. The effect of soybean galactooligosaccharides on nutrient and energy digestibility and digesta transit time in weaning piglets. Asian-Aust J Anim Sci. 2001;14:1598–1604. doi: 10.5713/ajas.2001.1598. [DOI] [Google Scholar]
- 6.Song YS, Pérez VG, Pettigrew JE, Martinez-Villaluenga C, de Mejia EG. Fermentation of soybean meal and its inclusion in diets for newly weaned pigs reduced diarrhea and measures of immunoreactivity in the plasm. Anim Feed Sci Technol. 2010;159:41–49. doi: 10.1016/j.anifeedsci.2010.04.011. [DOI] [Google Scholar]
- 7.Wang Y, Liu XT, Wang HL, Li DF, Piao XS, Lu WQ. Optimization of processing conditions for solid-state fermented soybean meal and its effects on growth performance and nutrient digestibility of weanling pigs. Livest Sci. 2014;170:91–99. doi: 10.1016/j.livsci.2014.07.020. [DOI] [Google Scholar]
- 8.Zhang WJ, Xu ZR, Zhao SH, Sun JY, Yang X. Development of a microbial fermentation process for detoxification of gossypol in cottonseed meal. Anim Feed Sci Technol. 2007;135:176–86. doi: 10.1016/j.anifeedsci.2006.06.003. [DOI] [Google Scholar]
- 9.Shi CY, He J, Yu J, Yu B, Huang Z, Mao XB, et al. Solid state fermentation of rapeseed cake with Aspergillus niger for degrading glucosinolates and upgrading nutritional value. J Anim Sci Biotechnol. 2015;6:13–20. doi: 10.1186/s40104-015-0015-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Canibe N, Jensen BB. Fermented liquid feed – microbial and nutritional aspects and impact on enteric diseases in pigs. Anim Feed Sci Technol. 2012;173:17–40. doi: 10.1016/j.anifeedsci.2011.12.021. [DOI] [Google Scholar]
- 11.Ovissipour M, Abedian A, Motamedzadegan A, Rasco B, Safari R, Shahiri H. The effect of enzymatic hydrolysis time and temperature on the properties of protein hydrolysates from Persian sturgeon (Acipenser persicus) viscera. Food Chem. 2009;115:238–42.12. Nair VC, Duvnjak Z. Reduction of phytic acid content in canola meal by Aspergillus ficuum in solid state fermentation process. Appl Microbiol Biotechnol. 1990;34:183–88. doi: 10.1007/BF00166777. [DOI] [Google Scholar]
- 12.Faurobert M. Application of two-dimensional gel electrophoresis to Prunus armeniaca leaf and bark tissues. Electrophoresis. 1997;17:170–73. doi: 10.1002/elps.1150180130. [DOI] [PubMed] [Google Scholar]
- 13.Hong KJ, Lee CH, Kim SW. Aspergillus oryzae GB-107 fermentation improves nutritional quality of food soybeans and feed soybean meals. J Med Food. 2004;7:430–35. doi: 10.1089/jmf.2004.7.430. [DOI] [PubMed] [Google Scholar]
- 14.Sakamoto K, Asano T, Furuya A, Takahashi S. Estimation of in vivo digestibility with the laying hen by an in vitro method using the intestinal fluid of the pig. Brit J Nutr. 1980;43:389–91. doi: 10.1079/BJN19800103. [DOI] [PubMed] [Google Scholar]
- 15.Plumed-Ferrer C, Von Wright A. Fermented pig liquid feed: nutritional, safety and regulatory aspects. J Appl Microbiol. 2009;106:351–68. doi: 10.1111/j.1365-2672.2008.03938.x. [DOI] [PubMed] [Google Scholar]
- 16.Missotten JAM, Michiels J, Ovyn A, De Smet S, Dierick NA. Fermented liquid feed for pigs. Arch Anim Nutr. 2010;64:437–66. doi: 10.1080/1745039X.2010.512725. [DOI] [PubMed] [Google Scholar]
- 17.van Winsen RL, Urlings BAP, Lipman LJA, Snijders JMA, Keuzenkamp D, Verheijden JHM. Effect of fermented feed on the microbial population of the gastrointestinal tracts of pigs. Appl Environ Microb. 2001;67:3071–76. doi: 10.1128/AEM.67.7.3071-3076.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lyberg K, Lundh T, Pedersen C, Lindberg JE. Influence of soaking, fermentation and phytase supplementation on nutrient digestibility in pigs offered a grower diet based on wheat and barley. Anim Sci. 2006;82:853–58. doi: 10.1017/ASC2006109. [DOI] [Google Scholar]
- 19.Hong TTT, Thuy TT, Passoth V, Lindberg JE. Gut ecology, feed digestion and performance in weaned piglets fed liquid diets. Livest Sci. 2009;125:232–37. doi: 10.1016/j.livsci.2009.04.013. [DOI] [Google Scholar]
- 20.Missotten JAM, Michiels J, Degroote J, Smet SD. Fermented liquid feed for pigs: an ancient technique for the future. J Anim Sci Biotechnol. 2015;6:4. doi: 10.1186/2049-1891-6-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Jensen BB, Mikkelsen LL. Feeding liquid diets to pigs. In: Garnsworthy PC, Wiseman J, editors. Recent Advances in Animal Nutrition. Nottingham, UK: Nottingham University Press; 1998. pp. 107–26. [Google Scholar]
- 22.Canibe N, Jensen BB. Fermented and non-fermented liquid feed to growing pigs: Effect on aspects of gastrointestinal ecology and growth performance. J Anim Sci. 2003;81:2019–31. doi: 10.2527/2003.8182019x. [DOI] [PubMed] [Google Scholar]
- 23.Canibe N, Jensen BB. Fermented liquid feed and fermented grain to piglets- effect on gastrointestinal ecology and growth performance. Livest Sci. 2007;108:232–35. doi: 10.1016/j.livsci.2007.01.078. [DOI] [PubMed] [Google Scholar]
- 24.Canibe N, Miettinen H, Jensen BB. Effect of adding Lactobacillus plantarum or a formic acid containing product to fermented liquid feed on gastrointestinal ecology and growth performance of piglets. Livest Sci. 2007;114:251–62. doi: 10.1016/j.livsci.2007.05.002. [DOI] [Google Scholar]
- 25.Aguirre L, Hebert EM, Garro MS, Giori GSD. Proteolytic activity of Lactobacillus strains on soybean proteins. LWT-Food Sci Technol. 2014;59:780–85. doi: 10.1016/j.lwt.2014.06.061. [DOI] [Google Scholar]
- 26.Chi CH, Cho SJ. Improvement of bioactivity of soybean meal by solid state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT-Food Sci Technol. 2016;68:619–25. doi: 10.1016/j.lwt.2015.12.002. [DOI] [Google Scholar]
- 27.Frias J, Song YS, Martínez-Villaluenga C, González DME, Vidal-Valverde C. Immunoreactivity and amino acid content of fermented soybean products. J Agric Food Chem. 2008;56:99–105. doi: 10.1021/jf072177j. [DOI] [PubMed] [Google Scholar]
- 28.Feng J, Liu X, Xu ZR, Lu YP, Liu YY. The effect of Aspergillus oryzae fermented soybean meal on growth performance, digestibility of dietary components and activities of intestinal enzymes in weaned piglets. Anim Feed Sci Technol. 2007;134:295–303. doi: 10.1016/j.anifeedsci.2006.10.004. [DOI] [Google Scholar]
- 29.Simonen M, Palva I. Protein secretion in Bacillus species. Microbiol Mol Biol Rev. 1993;57:109–137. doi: 10.1128/mr.57.1.109-137.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hu JK, Lu WQ, Wang CL, Zhu RH, Qiao JY. Characteristics of solid-state fermented feed and its effects on performance and nutrient digestibility in growing-finishing pigs. Asian-Aust J Anim Sci. 2008;21:1635–41. doi: 10.5713/ajas.2008.80032. [DOI] [Google Scholar]
- 31.Chen CC, Shih YC, Chiou PWS, Yu B. Evaluating nutritional quality of single stage- and two stage-fermented soybean meal. Asian-Aust J Anim Sci. 2010;23:598–606. doi: 10.5713/ajas.2010.90341. [DOI] [Google Scholar]
- 32.Chiang G, Lu WQ, Piao XS, Hu JK, Gong LM, Thacker PA. Effects of feeding solid-state fermented rapeseed meal on performance, nutrient digestibility, intestinal ecology and intestinal morphology of broiler chickens. Asian-Aust J Anim Sci. 2010;23:263–71. doi: 10.5713/ajas.2010.90145. [DOI] [Google Scholar]
- 33.Shi C, He J, Yu J, Yu B, Mao XB, Zheng P, et al. Amino acid, phosphorus, and energy digestibility of Aspergillus niger fermented rapeseed meal fed to growing pigs. J Anim Sci. 2015;93:2916–25. doi: 10.2527/jas.2014-8326. [DOI] [PubMed] [Google Scholar]
- 34.Sun H, Tang JW, Yao XH, Wu XF, Wang X, Feng J. Improvement of the Nutritional Quality of Cottonseed Meal by Bacillus subtilis and the Addition of Papain. Int J Agric Biol. 2012;14:563–68. [Google Scholar]
- 35.Rozan P, Villaume C, Bau HM, Schwertz A, Nicolas JP, Mejean L. Detoxication of rapeseed meal by Rhizopus oligosporus sp-T3: A first step towards rapeseed protein concentrate. Int J Food Sci Technol. 1996;31:85–90. doi: 10.1111/j.1365-2621.1996.17-315.x. [DOI] [Google Scholar]
- 36.Stokes JL, Gunness M. The amino acid composition of microorganisms. J Bacteriol. 1946;52:195–207. doi: 10.1128/JB.52.2.195-207.1946. [DOI] [PubMed] [Google Scholar]
- 37.Kuchroo CN, Fox PF. Soluble nitrogen in cheddar cheese: Comparison of extraction procedures. Milchwissenschaft. 1982;37:331–35. [Google Scholar]
- 38.Gilbert ER, Wong EA, Webb KE. Peptide absorption and utilization: Implications for animal nutrition and health. J Anim Sci. 2008;86:2135–55. doi: 10.2527/jas.2007-0826. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets generated and analyzed during the current study are not publicly available. Please contact the authors for data requests.