

Abstract
Fungal–bacterial interactions are highly diverse and contribute to many ecosystem processes. Their emergence under common environmental stress scenarios however, remains elusive. Here we use a synthetic microbial ecosystem based on the germination of Bacillus subtilis spores to examine whether fungal and fungal-like (oomycete) mycelia reduce bacterial water and nutrient stress in an otherwise dry and nutrient-poor microhabitat. We find that the presence of mycelia enables the germination and subsequent growth of bacterial spores near the hyphae. Using a combination of time of flight- and nanoscale secondary ion mass spectrometry (ToF- and nanoSIMS) coupled with stable isotope labelling, we link spore germination to hyphal transfer of water, carbon and nitrogen. Our study provides direct experimental evidence for the stimulation of bacterial activity by mycelial supply of scarce resources in dry and nutrient-free environments. We propose that mycelia may stimulate bacterial activity and thus contribute to sustaining ecosystem functioning in stressed habitats.
The maintenance of bacterial and fungal activity is essential for ecosystem functioning, particularly in dry soils where the two phyla co-exist. Here, Worrich and colleagues show experimentally that mycelia traffic water and nutrients and thereby stimulate bacterial activity in stressful conditions.
Fungi and bacteria co-inhabit a wide variety of environments1,2 and their interactions are significant drivers of important ecosystem functions and services3. In nature, they live in habitats exposed to fluctuating environmental conditions and frequently underlie strong selection pressure by limiting resources or drought4. To survive periods of stress, bacteria enter reversible states of low metabolic activity or dormancy5. Dormancy transiently disables bacterial functions while generating a seed bank of resting bacteria with the potential to be resuscitated in response to favourable environmental changes5,6. Fungi, by contrast, have been shown to possess higher resistance to drought and nutrient limitation due to efficient resource translocation between spatially separated sources and sink regions in their mycelia7,8,9,10. The redistribution of water in fungal mycelia was shown to enhance ecosystem resistance by preserving fungal carbon mineralization during drought11. While recent data suggest that mycelia create hospitable microhabitats for bacteria due to the exudation of carbonaceous compounds2,12,13 and a moistening of the surrounding substrate11, experimental evidence for a stimulation of bacterial activity in harsh environments is lacking.
During the past decade, powerful methods have been developed for the investigation of microbial interactions down to the single-cell scale14,15. Numerous studies revealed that resource exchange is a successful strategy in microbial communities to cope with unfavourable environmental conditions16,17,18,19. However, the role of resource transfer processes from fungi to bacteria for the maintenance of bacterial activity in dry and oligotrophic habitats remains to be elucidated.
In the present study, we use a synthetic microbial ecosystem to examine whether mycelia of fungi and oomycetes reduce water and nutrient stress for bacteria and thus enable bacterial activity in otherwise dry and oligotrophic environments. We introduce initially inactive spores of the soil-bacterium Bacillus subtilis into a water- and nutrient-free area and several hyphal model organisms, growing on a physically separated water and nutrient-rich agar piece, were allowed to overgrow the spore-bearing region. We find that the presence of mycelia enables both the germination of bacterial spores as well as vegetative growth near the hyphae. Spatially resolved secondary ion mass spectrometry in combination with stable isotope labelling reveals a long distance transport of labelled compounds in mycelia and a supply of water, carbon and nitrogen to the cells of B. subtilis located close to the hyphae. Spores more distant from the mycelium remain dormant and show no enrichment of the labelled substrates. Our results demonstrate that mycelium-forming fungi and oomycetes facilitate bacterial activity in dry and oligotrophic environments by providing nitrogen, carbon and water to bacteria and thus improving their habitat conditions.
Results
Mycelia affect spore germination and growth of B. subtilis
We developed a synthetic microbial ecosystem to assess whether fungal–bacterial interactions emerge in presence of drought and nutrient limitation stress conditions. Specifically, we tested to what degree the presence of mycelia evokes beneficial shifts in the habitat conditions for bacterial activity and growth. To this end, we placed spores of B. subtilis on silicon wafers (Fig. 1a) and analysed their germination and growth as an indicator for access to water and nutrients. The ascomycete Fusarium oxysporum, the basidiomycete Lyophyllum sp. Karsten, and the mycelium-forming oomycete Pythium ultimum were inoculated to a water and nutrient-rich agar patch physically separated from the spore-bearing silicon wafer (Fig. 1a). The mycelium overgrew the wafer within two (P. ultimum), four (F. oxysporum) and five days (Lyophyllum sp. Karsten) in direction of a second nutrient and water reservoir at the opposite side of the wafer. Subsequently, all biomass was detached from the wafer and the abundance of B. subtilis cells in presence and absence of mycelia determined by counting total colony forming units (c.f.u.; Fig. 2a). The number of total c.f.u. detached from mycelia-free controls 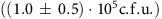 corresponded to the number of spores applied
corresponded to the number of spores applied 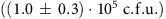 and thus excluded harmful wafer effects on spore germination (Fig. 2b). When hyphae overgrew the spore region, however, the total average c.f.u. numbers increased (Fig. 2b), indicating that mycelia supported growth of bacteria in dry and nutrient-free regions on the wafer. The highest c.f.u. numbers of B. subtilis were observed in presence of the oomycete P. ultimum
and thus excluded harmful wafer effects on spore germination (Fig. 2b). When hyphae overgrew the spore region, however, the total average c.f.u. numbers increased (Fig. 2b), indicating that mycelia supported growth of bacteria in dry and nutrient-free regions on the wafer. The highest c.f.u. numbers of B. subtilis were observed in presence of the oomycete P. ultimum
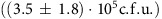 , followed by Lyophyllum sp. Karsten
, followed by Lyophyllum sp. Karsten 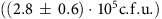 and F. oxysporum
and F. oxysporum
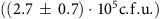 . Furthermore, we analysed the number of spores to obtain information on the fraction of sporulated and germinated cells relative to total cell numbers (Fig. 2a). In the control, we found that
. Furthermore, we analysed the number of spores to obtain information on the fraction of sporulated and germinated cells relative to total cell numbers (Fig. 2a). In the control, we found that 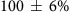 of the total c.f.u. remained in form of spores on the wafers. However, on P. ultimum wafers only
of the total c.f.u. remained in form of spores on the wafers. However, on P. ultimum wafers only 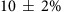 of the total cells were dormant (Fig. 2c). For F. oxysporum and Lyophyllum sp. Karsten the fraction of spores was higher (
of the total cells were dormant (Fig. 2c). For F. oxysporum and Lyophyllum sp. Karsten the fraction of spores was higher (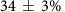 and
and 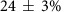 , respectively), but vegetative cells still accounted for the majority of the total c.f.u. (Fig. 2c). Microscopic analyses revealed the presence of vegetative B. subtilis cells always in close vicinity to the hyphae of P. ultimum (Fig. 1b,c magenta arrows). At locations more distant to hyphae however, B. subtilis persisted in the spore form (Fig. 1b,c yellow arrows).
, respectively), but vegetative cells still accounted for the majority of the total c.f.u. (Fig. 2c). Microscopic analyses revealed the presence of vegetative B. subtilis cells always in close vicinity to the hyphae of P. ultimum (Fig. 1b,c magenta arrows). At locations more distant to hyphae however, B. subtilis persisted in the spore form (Fig. 1b,c yellow arrows).
Figure 1. Synthetic microbial ecosystem reveals spore germination in presence of mycelia in dry and oligotrophic environments.

(a) Scheme and photographs of the setup employed to carry out the germination, growth and labelling experiments. The synthetic ecosystem is comprised of two agar plugs serving as water and nutrient sources (‘with') for the fungi or the oomycete inoculated on top of one of the agar plugs. A silicon wafer free of water and nutrients (‘without') placed in the middle between the two ‘with' zones served as carrier for spores of B. subtilis. An air gap between the ‘with' and ‘without' zone prevented the diffusion of water or substrates to the spore region. (b) Gradual enlargement of bright-field micrographs of the silicon wafer overgrown by mycelium of P. ultimum. In close vicinity to the hyphae (black arrow) rod-shaped, vegetative bacterial cells (magenta arrows) were found, whereas smaller, round-shaped spores (yellow arrows) were located more distantly. (c) Fluorescence micrograph of the 4′,6-diamidine-2-phenylindol (DAPI)-stained wafer showing P. ultimum hyphae containing nuclei (white arrows) and vegetative cells as well as spores of B. subtilis.
Figure 2. The presence of mycelium enables bacterial growth and spore germination.

(a) Scheme of the experimental procedure providing information about vegetative growth and germination in presence of mycelium. Total cell number was determined as c.f.u. after cell detachment from the wafer and plating on agar. Spores were obtained by counting c.f.u. after heat-inactivation of the vegetative cells. (b) c.f.u. of B. subtilis after detachment from the control wafer (no contact to mycelium) and wafers overgrown by P. ultimum, F. oxysporum or Lyophyllum sp. Karsten. The dashed line shows the number of c.f.u. applied to the wafer with the inoculum. In presence of mycelium, the number of total c.f.u. increased compared with the control. Bars show the average number of c.f.u. and error bars indicate the s.d. (c) Respective proportions of B. subtilis spores, calculated by dividing the number of spore c.f.u. by the total cell c.f.u. determined in b. The bars show the mean of the quotients for the three replicates and the s.d. The number of spores was different from the number of total cells for the respective strain but did not differ for the control.
Chemical mapping of B. subtilis and P. ultimum with ToF–SIMS
ToF–SIMS (time of flight- secondary ion mass spectrometry) analyses without prior isotope labelling were used to map the structural arrangement and elemental composition of the mycelium, vegetative bacterial cells and bacterial spores directly on the silicon wafer without previous isolation. This method prevented us from losing spatial and chemical information typically occurring during sample preparation in SIMS experiments20. The mass peaks corresponding to O−, OH−, PO2−, CH−, CN− and S− secondary ions were selected to represent the variance in composition of the biomass and its extracellular environment. To reduce the intensity modulation due to topography and effects related to the density of the sample, we normalized the intensity in respective ion yield distribution maps to total ion counts. As in microscopy, the ToF–SIMS analysis showed that vegetative cells of B. subtilis were located in close vicinity to the hyphae, whereas the spores of B. subtilis were randomly distributed regardless of the presence of hyphae (Fig. 3). The B. subtilis spores, which are known to be rich in sulfur compounds21, were observed as bright spots in the normalized intensity maps of O− and S− species (Fig. 3a,f) and vegetative cells were detected by their enhanced intensity of OH− ions (Fig. 3b), respectively. The PO2− and CH− ion signal intensities however, could not be used for detailed biomass analysis, as a diffuse, strong PO2− (Fig. 3c) and CH− (Fig. 3d) ion yield, distributed over the whole field of analysis, precluded clear discrimination between hyphae, spores and vegetative cells and pointed at the presence of extracellular material, possibly due to excretion, lysis of hyphae or damaged bacterial cells. By contrast, high secondary ion yields for CN− (Fig. 3e) reflected locally distinct protein contents and likely revealed locations of vegetative bacterial cells (white arrows) and residual hyphal fragments.
Figure 3. ToF–SIMS reveals sample composition via yield of secondary ion species.

Mass-resolved chemical map of P. ultimum hyphae as well as vegetative cells and spores of B. subtilis on top of the silicon wafer. Six negative secondary ion species were detected (a–f) in a 56 × 56 μm sample area. ToF–SIMS images produced were normalized by the total ion counts. The colour scale indicates the relative ion counts for each secondary ion species with warmer colours representing higher relative ion counts and cooler colours representing lower relative ion counts. Arrows in e point to vegetative cells (V). Scale bars, 10 μm.
NanoSIMS analysis of water and nutrient transfer
Building on the ToF–SIMS chemical mapping, we used nanoSIMS and isotope labelling approaches to obtain quantitative information on the nutrient and water transfer from the hyphae of P. ultimum to spores and vegetative cells of B. subtilis, respectively. The distribution of 18O, 13C and 15N in hyphae, vegetative cells and spores was quantified after the mycelium of P. ultimum had overgrown the spore-bearing wafer (Figs 4 and 5). P. ultimum was grown in parallel experiments with 18O-labelled water or with a combination of 13C-glucose and 15N-ammonium sulfate, to prevent slow development of hyphae due to strong kinetic isotope effects22,23. The localization of the biomass and its integrity was assessed by 12C14N− detection in two different fields of analysis (Fig. 4a,c). Both, vegetative cells and spores consistently provided strong 12C14N− signals, whereas hyphae of P. ultimum were found to be intact only in one of the two observation fields studied (Fig. 4a,c). As for microscopy and ToF–SIMS, vegetative cells of B. subtilis were either observed in close vicinity to the intact hyphae or lysed hyphal biomass. Strikingly, vegetative cells were consistently positioned longitudinally along the hyphae, forming a regular stratification typical for B. subtilis biofilms24 (Fig. 4c). Due to entire 12C and 14N replacement by their corresponding stable isotopes neither the hyphae nor the hyphae-attached vegetative cells were detected by the 12C14N− signal in 13C and 15N labelling experiments. High 12C14N− signals, however, were seen in spores distant from the hyphae as was further confirmed by the secondary electron image (Fig. 5a).
Figure 4. Water is transferred from hyphae to bacterial cells in dependence of the spatial organization.

NanoSIMS images of P. ultimum hyphae (H), B. subtilis spores (S) and vegetative cells (V) identified in the total biomass (12C14N−) images (a,c) of 18O-enriched samples. The ratio images of 18O/16O show the incorporation of label from water from the nutrient-rich zones into the biomass of P. ultimum and B. subtilis (b,d). The colour scale indicates the intensities of 12C14N− (a,c) and enrichment in 18O (b,d) with warmer colours representing higher secondary ion counts (a,c) or enrichment (b,d) levels and cooler colours representing lower values. Dashed lines indicate natural abundance of 18O. Images represent different fields of analysis corresponding to sample areas of 20 × 20 (a,b) and 30 × 30 μm (c,d). Scale bars, 4 μm.
Figure 5. Bacterial cells in close proximity to the hyphae receive carbon and nitrogen.

NanoSIMS images of P. ultimum hyphae (H), B. subtilis spores (S) and vegetative cells (V) identified in the secondary electron (SE) image (a) of 13C and 15N -enriched samples. The ratio images of 13C14N/12C14N (b) and 12C15N/12C14N (c) show the incorporation of label into the biomass of P. ultimum and B. subtilis. The colour scale indicates the relative enrichment in 13C and 15N (b,c) with warmer colours representing higher enrichment levels and cooler colours indicating lower values. The higher background signal for 13C on the left side of the hyphae (b) results from the topography-dependent re-deposition of sputtered sample material. Dashed lines indicate natural abundances of 13C and 15N. Vegetative cells and spores are framed in black and white, respectively. Images represent a field of analysis corresponding to a sample area of 40 × 40 μm. Scale bars, 4 μm.
Fungal biomass was found to be uniformly enriched in 18O except for small holes probably arising from ongoing hyphal disintegration (Fig. 4b). Vegetative cells in close vicinity to the hyphae showed higher enrichment in 18O (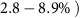 than spores located at longer distance to hyphae (
than spores located at longer distance to hyphae (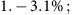 Fig. 6a) suggesting 18O transfer from the hyphae to vegetative cells. Compared with intact hyphae (
Fig. 6a) suggesting 18O transfer from the hyphae to vegetative cells. Compared with intact hyphae (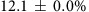 excess over natural abundance), 18O enrichment of B. subtilis cells however, was lower. This is presumably due to the much smaller diameter and the dilution of the signal attributed to the spores' own oxygen-containing molecules or the core water (which can account for up to 27–55% of the core wet weight25). Enrichment levels of 18O in spores was slightly higher in the 18O labelling experiments compared with non-labelled samples (Fig. 6a; Supplementary Fig. 1). Thus, some transport of water must have occurred via the gas phase. The transfer of carbon and nitrogen containing nutrients from hyphae to the bacterial cells was underpinned by significant 13C and 15N enrichment in hyphae and vegetative cells vicinal to hyphae yet not in spores more distant to hyphae (Fig. 5b,c). Relative to the 15N and 13C enrichment of the hyphae (15N:
excess over natural abundance), 18O enrichment of B. subtilis cells however, was lower. This is presumably due to the much smaller diameter and the dilution of the signal attributed to the spores' own oxygen-containing molecules or the core water (which can account for up to 27–55% of the core wet weight25). Enrichment levels of 18O in spores was slightly higher in the 18O labelling experiments compared with non-labelled samples (Fig. 6a; Supplementary Fig. 1). Thus, some transport of water must have occurred via the gas phase. The transfer of carbon and nitrogen containing nutrients from hyphae to the bacterial cells was underpinned by significant 13C and 15N enrichment in hyphae and vegetative cells vicinal to hyphae yet not in spores more distant to hyphae (Fig. 5b,c). Relative to the 15N and 13C enrichment of the hyphae (15N: 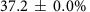 and 13C:
and 13C: 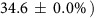 , vegetative bacterial cells were more enriched in 15N (
, vegetative bacterial cells were more enriched in 15N (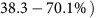 than in 13C (
than in 13C (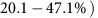 indicating both a lower dilution by spore-inherent nitrogen compounds and a higher nitrogen demand by germinating spores and growing cells of B. subtilis, respectively. By contrast, spores were 13C and 15N enriched by
indicating both a lower dilution by spore-inherent nitrogen compounds and a higher nitrogen demand by germinating spores and growing cells of B. subtilis, respectively. By contrast, spores were 13C and 15N enriched by 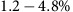 and
and 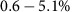 , respectively, and showed C and N isotope abundances comparable to non-labelled samples (Fig. 6b,c; Supplementary Fig. 2). The 18O and the joint 13C and 15N enrichments, hence, provided clear evidence of transfer of water and nutrients from hyphae to bacterial cells and the importance of the relative spatial organization of spores and hyphae on the wafer surface, respectively. The low levels of stable isotope incorporation into bacterial spores distant to the hyphae and the observation that vegetative cells were solely found in close proximity to hyphae indicate a very low air-borne transfer of the isotope labels (for example, by humidity or fungal volatiles) to the spores.
, respectively, and showed C and N isotope abundances comparable to non-labelled samples (Fig. 6b,c; Supplementary Fig. 2). The 18O and the joint 13C and 15N enrichments, hence, provided clear evidence of transfer of water and nutrients from hyphae to bacterial cells and the importance of the relative spatial organization of spores and hyphae on the wafer surface, respectively. The low levels of stable isotope incorporation into bacterial spores distant to the hyphae and the observation that vegetative cells were solely found in close proximity to hyphae indicate a very low air-borne transfer of the isotope labels (for example, by humidity or fungal volatiles) to the spores.
Figure 6. Vegetative cells are higher enriched in 13C, 15N and 18O compared with spores of B. subtilis.

Atom percent enrichment (APE) for (a) 18O, (b) 13C and (c) 15N by single cells and spores of B. subtilis measured with nanoSIMS in labelled (green) and non-labelled (blue) samples. Data are derived from different fields of analysis and from replicate wafers (for details see Supplementary Table 1). Dashed lines represent literature values for the stable isotope natural abundance.
Discussion
By verifying hyphal uptake, translocation and transfer of 13C, 15N and 18O labels to subsequently germinating Bacillus subtilis spores, our results demonstrate that mycelia enable bacterial activity in regions devoid of water and nutrients. Utilizing a synthetic microbial ecosystem and a novel combination of ToF- and nanoSIMS quantitative imaging of single-cell trophic interactions we showed that (i) the presence of fungi and oomycetes in an initially dry and nutrient-free environment induced the germination of metabolically inactive spores of B. subtilis to actively growing cells, (ii) only spores in close vicinity to the hyphae could germinate, and (iii) a transfer of nutrients and water from hyphae to the bacterial cells underlay this activation. Thus, our study provides for the first time direct experimental evidence for stimulation of bacterial activity by mycelial supply of scarce resources in dry and nutrient-free environments.
Within microbial communities competitive and cooperative interactions are often related to the excretion and uptake of metabolites from involved partners17,26,27. It is widely assumed that fungal exudates are a source of nutrients for bacteria in the mycosphere including organic acids28, sugars, polyols29 and amino acids29. In addition to known germinants such as amino acids and sugars30, the availability of water is required for spore germination. Water uptake by the spore enables the hydration of the spore core and represents a crucial step in the germination process30. Recently, fungal hyphae were shown to redistribute water and thereby compensate differences in the soil matric potential11. Although the authors provided evidence that such water redistribution drastically increased the functional ecosystemic stability under drought, the consequences of this underrated pathway for other soil organisms remained unexplored. Our results suggest that hyphal water translocation may not only provide fungi with a higher resistance to drought, but also facilitates bacterial activity in close proximity to the hyphae. Especially at low soil water potentials, the mycosphere may thus form a hotspot of microbial activity and significantly foster the maintenance of important ecosystem functions and services.
Three different strategies are described by which bacteria may derive nutrition from fungi: necrotrophy, extracellular biotrophy and endocellular biotrophy. In necrotrophic interactions, bacteria secrete molecules that kill the fungus and thus induce a release of nutrients. Extracellular biotrophs live in close proximity and use nutrients exuded from the hyphae, whereas endocellular biotrophs live inside the fungus and take up nutrients from the cytoplasm31. In accordance with existing literature, we never observed any endocellular occurance of B. subtilis. Hence, we consider endocellular biotrophy to be rather unlikely. Indeed, vegetative cells were always located tightly arranged in a longitudinal orientation along the hyphae resulting in maximum contact surface between the bacterial cell and the hyphae. This strongly indicates extracellular biotrophy based on the consumption of fungal exudates31. The regular cell chains observed along the hyphae (Fig. 5a) most probably arose from subsequent growth of the cells leading to a colonization of the hyphae as usually observed for the interaction of B. subtilis with plant roots32. In agriculture B. subtilis strains are routinely applied to soil because their biocontrol activity was shown to protect plants against fungal infestation33. Active killing of fungal hyphae in necrotrophic interactions was described for different Bacillus strains34,35 and most likely induced the disintegration of the hyphae observed in the course of our SIMS experiments. However, before germination the metabolically inactive spores cannot have evoked killing of the hyphae. Moreover, we observed vegetative cells along undamaged hyphae of P. ultimum still containing intact nuclei (Fig. 1c). This demonstrates that neither necrotrophy nor nutrients leaking from dead hyphal cells can be solely responsible for the stimulation of bacterial activity in the vicinity of mycelia. The dry conditions on the wafer strongly restricted the diffusion of exudates and thus only spores in close contact to the mycelium germinated. Indeed, already at distances of more than 1.7 μm spores did not germinate (cf. Fig. 5a). Thus, the close proximity between the mycelium and the bacterial cells constituted a prerequisite for diffusion-based transfer of resources as observed in different studies on nutrient exchange36 or chemical communication37 in microbial communities. However, under more humid conditions in soil, exudate diffusion probably extends beyond the direct vicinity of the hyphae.
Our observation that fungi support the growth of bacteria is in accordance with studies that showed an increased number of culturable bacteria in the vicinity of different fungal hyphae in soil microcosms12. Given the evidence that bacteria are attracted to the mycosphere38, it is tempting to speculate that the local conditions in these regions are often favourable for bacterial activity. Fungal mycelia were shown to change local habitat conditions and thereby affect bacteria by changing the water availability, the pH, the soil structure or by secreting inhibitory or stimulatory compounds39. Thereby they create, modify or destroy habitats and thus pursue ‘ecosystem engineering', which may influence the ecological success of other species40. In our experiments, representatives of different fungal phyla as well as a mycelium-forming oomycete could induce the germination and growth of B. subtilis by the supply of sufficient amounts of water and nutrients. The differences observed for total c.f.u. and the amount of germinated cells may be explained by the differences in the mycelial coverage of the wafer or differing cell wall compositions of oomycetes and fungi such as the lack of chitin in P. ultimum. Although the combination of the selected organisms may be artificial, we observed similar net effects for all three mycelial organisms likely indicating widespread nutrient and water transfer from mycelia to bacterial cells. The emergence of the interaction under the extremely harsh environmental conditions prevailing in the synthetic microcosms strongly suggests that the observed phenomenon could also arise under a wide range of less extreme conditions found in natural systems. The finding that mycelia facilitate bacterial activity by multiple resource supply may help to further understand the small-scale dynamics of bacterial activity in heterogeneous ecosystems exposed to fluctuating environmental conditions. The ability of mycelia to redistribute and transfer resources allows bacteria to occupy ecological niches which would otherwise be uninhabitable to them. For assessing the dynamics and stability of microbial ecosystem functioning, fungi and bacteria need to be considered as interconnected entities rather than autonomously operating components of the ecosystem.
Methods
Microcosm setup
Experiments were carried out in laboratory microcosms mimicking water and nutrient-rich (‘with') and depleted (‘without') zones (Fig. 1a). To create ‘with' zones, two agar plugs (Ø 1 cm) were cut out from an FAB-medium agar plate (medium composition adapted from41) supplemented with 5 mM glucose as sole carbon source. These two plugs were placed in a petri dish (Ø 6 cm) at a distance of 1.4 cm from each other. Between the two plugs, a silicon wafer (Plano, Wetzlar, Germany) was placed representing the ‘without' zone. This wafer served as a carrier for spores of B. subtilis, which were used as an indicator of changes in water and nutrient availability. Before its use, the wafer was cleaned with acetone, 70% ethanol and finally with autoclaved water and dried under a laminar flow box. Spore suspensions were bought from Merck (Darmstadt, Germany) and controlled microscopically for the absence of vegetative cells before use. Spores were washed three times with sterile-filtered, double-distilled water and in each washing step the supernatant was separated from the spores by centrifugation at 10,000g for 10 min. Subsequently, spores were resuspended in bidistilled water at a final concentration of 2.5·106 spores ml−1 and 40 μl of the suspension were applied the wafer. Wafers were dried in an oven for 30 min at 70 °C.
Quantification of total cells and spores
The effects of mycelia in dry and nutrient-poor environments on the overall cell number and the abundance of spores were quantified using the fungi F. oxysporum or Lyophyllum sp. Karsten, or the oomycete P. ultimum. Therefore, a small amount of agar covered with mycelium from freshly overgrown FAB–glucose agar plates was cut out and transferred onto one of the agar plugs in the microcosms (Fig. 1a). Microcosms were sealed with parafilm, placed in a plastic container and incubated at 25 °C in the dark. The experiments were run in triplicates and control wafers were included. The control wafers were also inoculated with the spore suspension and placed within the microcosms but close to the edge of each petri dish to avoid contact with the mycelium. This should guarantee that all samples were exposed to the same water vapor pressure in the microcosm atmosphere. Mycelium-overgrown wafers and controls were excised from the microcosms after two (P. ultimum), four (F. oxysporum) and five (Lyophyllum sp. Karsten) days and transferred to glass tubes containing 0.5 ml 1 × PBS buffer (10 mM, pH 7). Cell detachment was carried out by vortexing the tubes for 1 min. Immediately, 400 μl were pipetted into new glass tubes and heated to 80 °C for 20 min to kill the vegetative cells42. Colony forming units in the heat-treated and untreated suspensions were counted using the drop plate method as described earlier43. Briefly, 10-fold dilution series of the supernatant were prepared directly in 96-well microtiter plates and five microlitres of consecutive dilutions were dropped on FAB-glucose agar plates containing 0.2% cycloheximid to suppress growth of the fungi and the oomycete. Plates were incubated at 25 °C for 48 h. Droplets giving rise to 5–30 single colonies were used to determine c.f.u. numbers.
Visualization of spore germination in presence of mycelia
Setups with P. ultimum were used to visualize germination of spores of B. subtilis in water and nutrient-free regions of the wafer. After the mycelium had overgrown the wafers, hyphae around the wafers were cut and the wafers were directly observed under the microscope (Axio Imager Z2, Carl Zeiss) using bright field illumination. Pictures were acquired with a Zeiss Axiocam 506 colour camera. To prove hyphae disintegration, samples were also stained with 4′,6-diamidine-2-phenylindol (DAPI)–amended mountant containing 9 parts Citifluor mountant (Citifluor, Leicester, United Kingdom), 1 part PBS (1 × ), and 1 μg ml−1 DAPI. Epifluorescence images were acquired using DAPI filter settings.
ToF–SIMS analysis
In preparation for the envisaged nanoSIMS analysis a boron-doped silicon wafer overgrown with P. ultimum was analysed via ToF–SIMS technique employing a ToF-SIMS.5 (ION-TOF GmbH, Münster) instrument. The conditions of the ToF–SIMS experiment allowed for a detection of atomic and molecular secondary ions with m/z (mass to charge) ratio from 1 to about 500. The broad-range mass spectrum (1–500 amu) acquired for each raster point enabled us to compare the yield of different secondary ion species extracted from the sample. Detailed conditions of the analysis are specified as follows: the ToF–SIMS experiment has been performed using fast imaging mode of ToF-SIMS.5 operation in combination with delayed extraction44,45 of negative secondary ions providing the Mass Resolving Power (MRP) above 3,000 and lateral resolution of about 130 nm. Under these experimental conditions, the 30 keV NanoProbe LMIG source was delivering 0.02 pA of primary Bi3+ cluster ions in 100 ns pulses with 200 μs repetition period. The analysis has been done in 400 scans (planes) with 5 shots of Bi3+ primary cluster ions per pixel distributed randomly in 512 × 512 raster over 56 × 56 μm sample area. The lateral distribution of ion yield was analysed using the proprietary ION-TOF SurfaceLab 6.5 software. The accumulation of acquired scans was done after lateral drift correction and the resulted total stack was analysed for the lateral distribution of the ion yield. The ToF–SIMS images produced were normalized by the total ion image.
Analysis of water and nutrient transfer
13C-labelled glucose (99 atom%; Euriso-Top, Saarbrücken, Germany), 15N-labelled ammonium sulfate (99 atom%; Sigma, Munich, Germany) and 18O-labelled water (97 atom%; Campro Scientific, Berlin, Germany) were used for the labelling experiment. For the experiments with the labelled water, FAB-agar plates with 5 mM glucose were dried completely in an oven at 60 °C until only an agar foil was left. Thirty mg of this foil were transferred into an 5 ml Eppendorf tube and 2 ml of the labelled water were added. For the 13C and 15N labelling experiments FAB-agar plates without glucose and ammonium sulfate were prepared, dried and again 30 mg were transferred to a 5 ml Eppendorff tube. Subsequently, 2 mg of 13C labelled glucose, 4 mg of 15N labelled ammonium sulfate and 2 ml autoclaved bidistilled water were added. The tube contents were incubated at 95 °C with continuous shaking at 700 r.p.m. until the agar was completely dissolved. Subsequently, two droplets of 400 μl were introduced into the petri dishes at a distance of 1.4 cm from each other and dried for 3 min under the laminar flow box until it formed hard agar blocks. The spores to serve as inoculum were prepared as described above. For the nanoSIMS experiment a conductive chromium-coated, round-shaped silicon wafer (10 mm diameter) was used as a spore carrier. Experiments were carried out using P. ultimum and inoculation was conducted as described above. Each labelling experiment was performed in duplicates. After 48 h, the wafers were removed from the microcosms and the biomass was fixed in an atmosphere over a solution of a 10% paraformaldehyde and 37% ethanol at 30 °C for 2 h. Wafers were stored in a vacuum box until analysis. Parallel control experiments (n=2) were performed with non-labelled compounds to assess the natural isotopic composition of the samples.
NanoSIMS analysis
The wafers were analysed with a NanoSIMS-50L instrument (CAMECA (AMETEK), Gennevilliers Cedex, France) to assess the transfer of the stable isotopes from the hyphae to the bacterial cells. Depending on the arrangement of hyphae, vegetative cells and spores, fields of analysis of 20 × 20, 30 × 30 and 40 × 40 μm were selected. Measurements were performed in negative extraction mode employing a DC source of primary Cs+ ions. Detailed information on the analytical conditions for the measurements are described as follows: two replicate wafers labelled with 18O, and 13C/15N isotopes as well as two control wafers with natural isotopic composition were analysed. The 2 pA beam of Cs+ ions was focused into ∼70 nm spot at the sample surface during the analysis. The energy of Cs+ collision with the sample was set to 16 keV in the analysis mode. The sample was scanned in a 512 × 512 px raster over analysis areas of 20 × 20, 30 × 30 or 40 × 40 μm with 5 msec dwell time per pixel. Before the analysis with 16 keV Cs+ beam, the sample surface of 60 × 60 μm area was treated with 10 nA of low-energy (50 eV) Cs+ beam for 10 min. The low-energy implantation/deposition of cesium has been performed with the purpose to equilibrate the working function for negative secondary ions and to make the outermost layer of the sample available for the analysis avoiding its sputtering during high-energy implantation. The secondary ions were analysed for their mass and charge ratio (m/z). Using the seven available collectors, the following secondary ion species were detected: 12C− (collector-1), 13C− (collector-2), 16O− (collector-3), 18O− (collector-4), 12C15N− and 13C14N− (collector-5), 32S− (collector-6), 31P16O2− (collector-7). Two secondary ion species (12C15N− and 13C14N−) were detected using the collector-5 by switching the deflector voltage in combined analysis mode. The mass resolving power (MRP) was checked to be between 8,000 and 12,000 (M/ΔM) with the exit slit width of 40 μm, 20 μm wide entrance slit, 200 μm aperture slit and with the energy slit cutting 30% of secondary ions in high-energy tail of their energy distribution. It has been proven to get the sample sputtered completely within 35 scans upon the used analysis conditions; scans 2–30 were thus considered for the image and data analysis employing Look@NanoSIMS software (LANS)46 allowing for stacking the scans with lateral drift correction and quantitative analysis of isotopic ratios (18O/16O, 13C14N/12C14N and 12C15N/12C14N). Quantification of the atomic percent enrichment in 18O, 13C and 15N for hyphae, vegetative cells and spores was carried out by defining regions of interest around individual cells using the secondary electron images. The number of analysed fields and replicate wafers for each labelling experiment as well as the numbers of individual vegetative cells and spores used to quantify APE is specified in Supplementary Table 1.
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable request. The raw data sets for ToF- and nanoSIMS measurements have been deposited in the figshare repository under https://doi.org/10.6084/m9.figshare.4742857.v1 (url for ToF–SIMS data) and https://doi.org/10.6084/m9.figshare.4725409.v1 (url for nanoSIMS data), respectively.
Additional information
How to cite this article: Worrich, A. et al. Mycelium-mediated transfer of water and nutrients stimulates bacterial activity in dry and oligotrophic environments. Nat. Commun. 8, 15472 doi: 10.1038/ncomms15472 (2017).
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Material
Supplementary Figures and Supplementary Table
Acknowledgments
We acknowledge the Centre for Chemical Microscopy (ProVIS) at the Helmholtz Centre for Environmental Research supported by European Regional Development Funds (EFRE—Europe funds Saxony) for using their analytical facilities. This work was funded by the Helmholtz Association via the integrated project Controlling Chemicals' Fate of the research topic Chemicals In The Environment (CITE) within the research programme Terrestrial environment. The authors thank Prof J.D. van Elsas for kind provision of strain Lyophyllum sp. Karsten. We are grateful to Sophie Steigerwald, Manuel Trost, Rita Remer and Jana Reichenbach for skilled experimental help.
Footnotes
The authors declare no competing financial interests.
Author contributions All authors conceived the study framework and discussed the theoretical background. A.W., N.M., M.K. and L.Y.W. planned the study. A.W. and H.S. conducted the experiments and compiled the data. L.Y.W. and M.K. supervised the experiments. The manuscript was written by A.W. with extensive input from all authors.
References
- Wargo M. J. & Hogan D. A. Fungal—bacterial interactions: a mixed bag of mingling microbes. Curr. Opin. Microbiol. 9, 359–364 (2006). [DOI] [PubMed] [Google Scholar]
- Pion M. et al. Bacterial farming by the fungus Morchella crassipes. Proc. Biol. Sci. 280, 20132242 (2013). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boer W. d., Folman L. B., Summerbell R. C. & Boddy L. Living in a fungal world: impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 29, 795–811 (2005). [DOI] [PubMed] [Google Scholar]
- Pande S. et al. Metabolic cross-feeding via intercellular nanotubes among bacteria. Nat. Commun. 6, 6238 (2015). [DOI] [PubMed] [Google Scholar]
- Lennon J. T. & Jones S. E. Microbial seed banks: the ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 9, 119–130 (2011). [DOI] [PubMed] [Google Scholar]
- Schimel J., Balser T. C. & Wallenstein M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 88, 1386–1394 (2007). [DOI] [PubMed] [Google Scholar]
- de Vries F. T. et al. Land use alters the resistance and resilience of soil food webs to drought. Nat. Clim. Change 2, 276–280 (2012). [Google Scholar]
- Six J. Soil science: fungal friends against drought. Nat. Clim. Change 2, 234–235 (2012). [Google Scholar]
- Harms H., Schlosser D. & Wick L. Y. Untapped potential: exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 9, 177–192 (2011). [DOI] [PubMed] [Google Scholar]
- Allen M. F. Mycorrhizal fungi: highways for water and nutrients in arid soils. Vadose Zone J. 6, 291 (2007). [Google Scholar]
- Guhr A., Borken W., Spohn M. & Matzner E. Redistribution of soil water by a saprotrophic fungus enhances carbon mineralization. Proc. Natl Acad. Sci. USA 112, 14647–14651 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warmink J. A., Nazir R. & van Elsas J. D. Universal and species-specific bacterial ‘fungiphiles' in the mycospheres of different basidiomycetous fungi. Environ. Microbiol. 11, 300–312 (2009). [DOI] [PubMed] [Google Scholar]
- Nazir R., Warmink J. A., Boersma H. & van Elsas J. D. Mechanisms that promote bacterial fitness in fungal-affected soil microhabitats. FEMS Microbiol. Ecol. 71, 169–185 (2010). [DOI] [PubMed] [Google Scholar]
- Ponomarova O. & Patil K. R. Metabolic interactions in microbial communities: untangling the Gordian knot. Curr. Opin. Microbiol. 27, 37–44 (2015). [DOI] [PubMed] [Google Scholar]
- Musat N., Foster R., Vagner T., Adam B. & Kuypers M. M. Detecting metabolic activities in single cells, with emphasis on nanoSIMS. FEMS Microbiol. Rev. 36, 486–511 (2012). [DOI] [PubMed] [Google Scholar]
- Fetzer I. et al. The extent of functional redundancy changes as species' roles shift in different environments. Proc. Natl Acad. Sci. USA 112, 14888–14893 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morris B. E. L., Henneberger R., Huber H. & Moissl-Eichinger C. Microbial syntrophy: interaction for the common good. FEMS Microbiol. Rev. 37, 384–406 (2013). [DOI] [PubMed] [Google Scholar]
- McNally L., Viana M. & Brown S. P. Cooperative secretions facilitate host range expansion in bacteria. Nat. Commun. 5, 4594 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pande S. et al. Fitness and stability of obligate cross-feeding interactions that emerge upon gene loss in bacteria. ISME J. 8, 953–962 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watrous J. D. & Dorrestein P. C. Imaging mass spectrometry in microbiology. Nat. Rev. Microbiol. 9, 683–694 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonsen P. P. M., Spudich J. A., Nelson D. L. & Kornberg A. Biochemical studies of bacterial sporulation and germination XII. A sulfonic acid as a major sulfur compound of Bacillus subtilis spores. J. Bacteriol. 98, 62–68 (1969). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meckenstock R. U., Morasch B., Griebler C. & Richnow H. H. Stable isotope fractionation analysis as a tool to monitor biodegradation in contaminated acquifers. J. Contam. Hydrol. 75, 215–255 (2004). [DOI] [PubMed] [Google Scholar]
- Elsner M., Zwank L., Hunkeler D. & Schwarzenbach R. P. A new concept linking observable stable isotope fractionation to transformation pathways of organic pollutants. Environ. Sci. Technol. 39, 6896–6916 (2005). [DOI] [PubMed] [Google Scholar]
- Claessen D., Rozen D. E., Kuipers O. P., Sogaard-Andersen L. & van Wezel G. P. Bacterial solutions to multicellularity: a tale of biofilms, filaments and fruiting bodies. Nat. Rev. Microbiol. 12, 115–124 (2014). [DOI] [PubMed] [Google Scholar]
- Setlow P. Spores of Bacillus subtilis: their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 101, 514–525 (2006). [DOI] [PubMed] [Google Scholar]
- Freilich S. et al. Competitive and cooperative metabolic interactions in bacterial communities. Nat. Commun. 2, 589 (2011). [DOI] [PubMed] [Google Scholar]
- Phelan V. V., Liu W.-T., Pogliano K. & Dorrestein P. C. Microbial metabolic exchange—the chemotype-to-phenotype link. Nat. Chem. Biol. 8, 26–35 (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Hees P. A. W. et al. Oxalate and ferricrocin exudation by the extramatrical mycelium of an ectomycorrhizal fungus in symbiosis with Pinus sylvestris. New Phytol. 169, 367–378 (2006). [DOI] [PubMed] [Google Scholar]
- Sun Y.-P. et al. Exudation-reabsorption in a mycorrhizal fungus, the dynamic interface for interaction with soil and soil microorganisms. Mycorrhiza 9, 137–144 (1999). [Google Scholar]
- Setlow P. Spore germination. Curr. Opin. Microbiol. 6, 550–556 (2003). [DOI] [PubMed] [Google Scholar]
- Leveau J. H. J. & Preston G. M. Bacterial mycophagy: definition and diagnosis of a unique bacterial–fungal interaction. New Phytol. 177, 859–876 (2008). [DOI] [PubMed] [Google Scholar]
- Cao Y. et al. Bacillus subtilis SQR 9 can control Fusarium wilt in cucumber by colonizing plant roots. Biol. Fertil. Soils 47, 495–506 (2011). [Google Scholar]
- Leclère V. et al. Mycosubtilin overproduction by Bacillus subtilis BBG100 enhances the organism's antagonistic and biocontrol activities. Appl. Environ. Microbiol. 71, 4577–4584 (2005). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chérif M. et al. Ultrastructure and cytochemistry of in vitro interactions of the antagonistic bacteria Bacillus cereus X16 and B. thuringiensis 55T with Fusarium roseum var. sambucinum. J. Plant Pathol. 84, 83–93 (2002). [Google Scholar]
- Budi S. W. et al. Hydrolytic enzyme activity of Paenibacillus sp. strain B2 and effects of the antagonistic bacterium on cell integrity of two soil-borne pathogenic fungi. Appl. Soil Ecol. 15, 191–199 (2000). [Google Scholar]
- Kim H. J., Boedicker J. Q., Choi J. W. & Ismagilov R. F. Defined spatial structure stabilizes a synthetic multispecies bacterial community. Proc. Natl Acad. Sci. USA 105, 18188–18193 (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassler B. L. & Losick R. Bacterially speaking. Cell 125, 237–246 (2006). [DOI] [PubMed] [Google Scholar]
- Warmink J. A. & van Elsas J. D. Migratory response of soil bacteria to Lyophyllum sp. strain Karsten in soil microcosms. Appl. Environ. Microbiol. 75, 2820–2830 (2009). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johansson J. F., Paul L. R. & Finlay R. D. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol. Ecol. 48, 1–13 (2004). [DOI] [PubMed] [Google Scholar]
- Erwin D. H. Macroevolution of ecosystem engineering, niche construction and diversity. Trends Ecol. Evol. 23, 304–310 (2008). [DOI] [PubMed] [Google Scholar]
- Dechesne A. et al. Biodegradation in a partially saturated sand matrix: compounding effects of water content, bacterial spatial distribution, and motility. Environ. Sci. Technol. 44, 2386–2392 (2010). [DOI] [PubMed] [Google Scholar]
- Monteiro S. M. et al. A procedure for high-yield spore production by Bacillus subtilis. Biotechnol. Prog. 21, 1026–1031 (2005). [DOI] [PubMed] [Google Scholar]
- Chen C.-Y., Nace G. W. & Irwin P. L. A 6 × 6 drop plate method for simultaneous colony counting and MPN enumeration of Campylobacter jejuni, Listeria monocytogenes, and Escherichia coli. J. Microbiol. Methods 55, 475–479 (2003). [DOI] [PubMed] [Google Scholar]
- Lee J. L. S., Gilmore I. S., Fletcher I. W. & Seah M. P. Topography and field effects in the quantitative analysis of conductive surfaces using ToF-SIMS. Appl. Surf. Sci. 255, 1560–1563 (2008). [Google Scholar]
- Vanbellingen Q. P. et al. Time-of-flight secondary ion mass spectrometry imaging of biological samples with delayed extraction for high mass and high spatial resolutions. Rapid Commun. Mass Spectrom. 29, 1187–1195 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polerecky L. et al. Look@NanoSIMS—a tool for the analysis of nanoSIMS data in environmental microbiology. Environ. Microbiol. 14, 1009–1023 (2012). [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figures and Supplementary Table
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request. The raw data sets for ToF- and nanoSIMS measurements have been deposited in the figshare repository under https://doi.org/10.6084/m9.figshare.4742857.v1 (url for ToF–SIMS data) and https://doi.org/10.6084/m9.figshare.4725409.v1 (url for nanoSIMS data), respectively.